
Tyrphostin AG1024 Suppresses Coronaviral Replication by Downregulating JAK1 via an IR/IGF-1R Independent Proteolysis Mediated by Ndfip1/2_NEDD4-like E3 Ligase Itch


Abstract
1. Introduction
2. Results
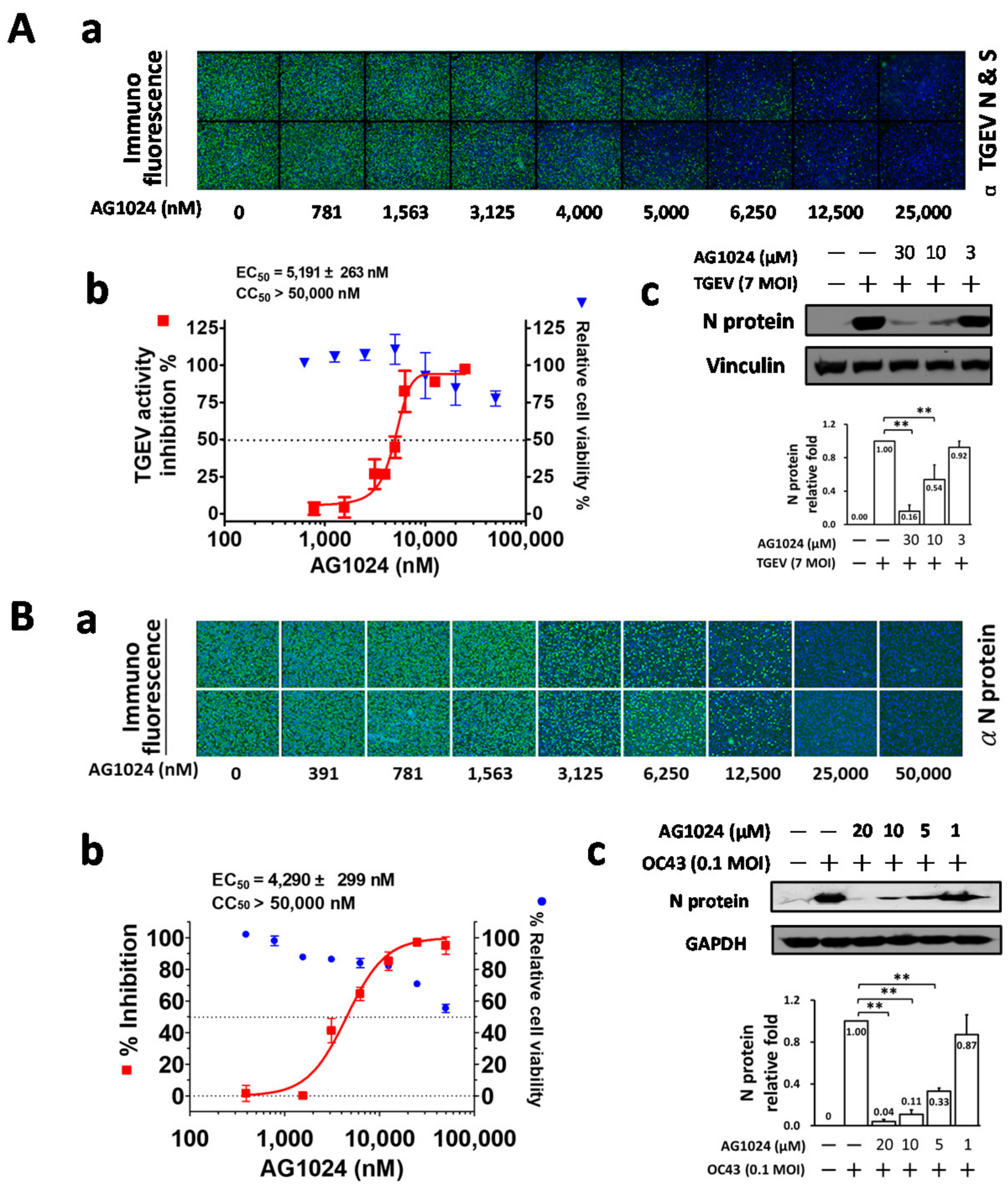
2.1. Tyrphostin AG1024 Inhibited TGEV and HCoV-OC43 Coronaviral Activities
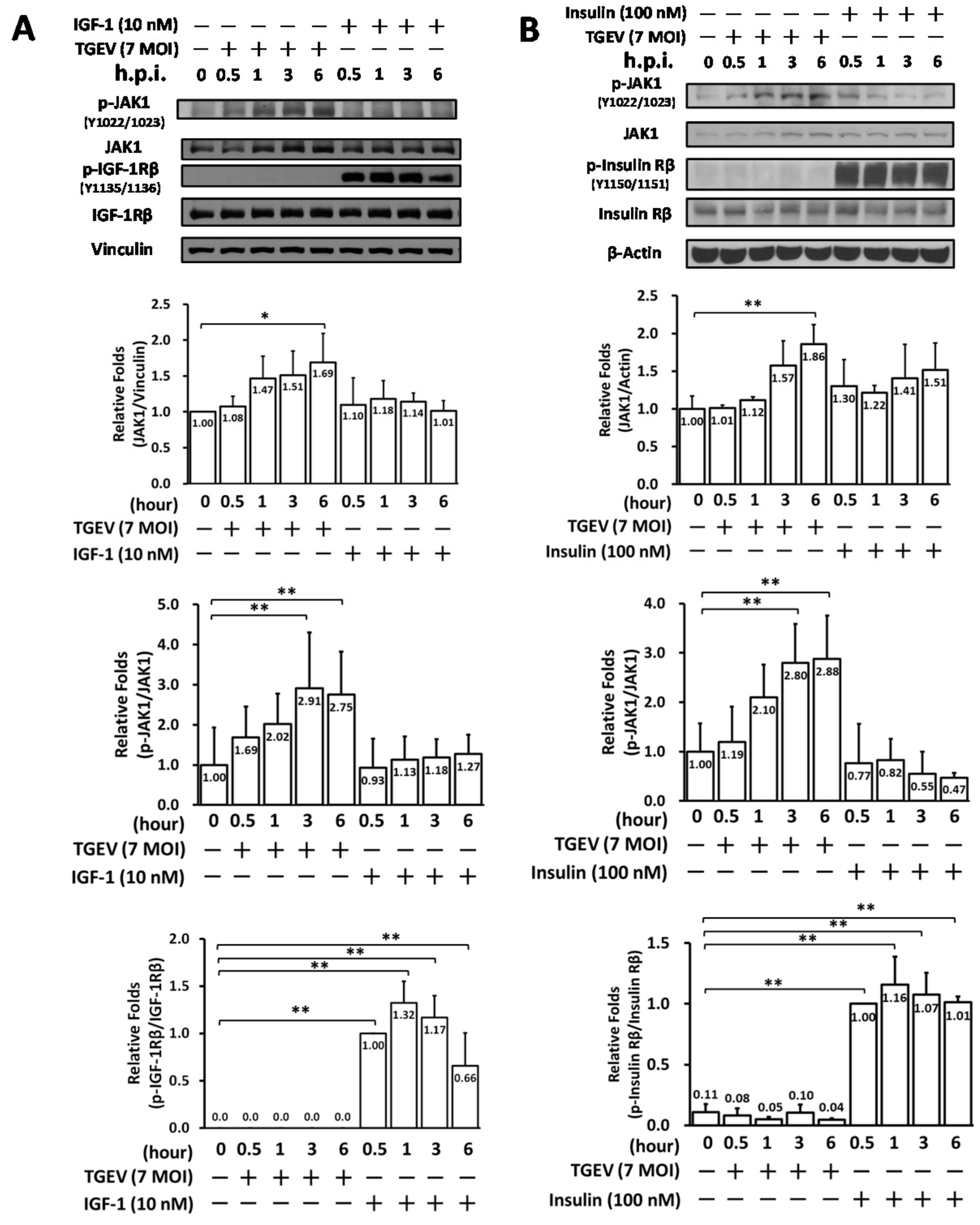
2.2. Coronaviral Inhibition of Tyrphostin AG1024 Is IR/IGF-1R Signaling Independent
2.3. AG1024 Treatment Resulted in JAK1 Diminishment/Downregulation and Inhibited Viral N Protein Expression in TGEV Infected ST Cells
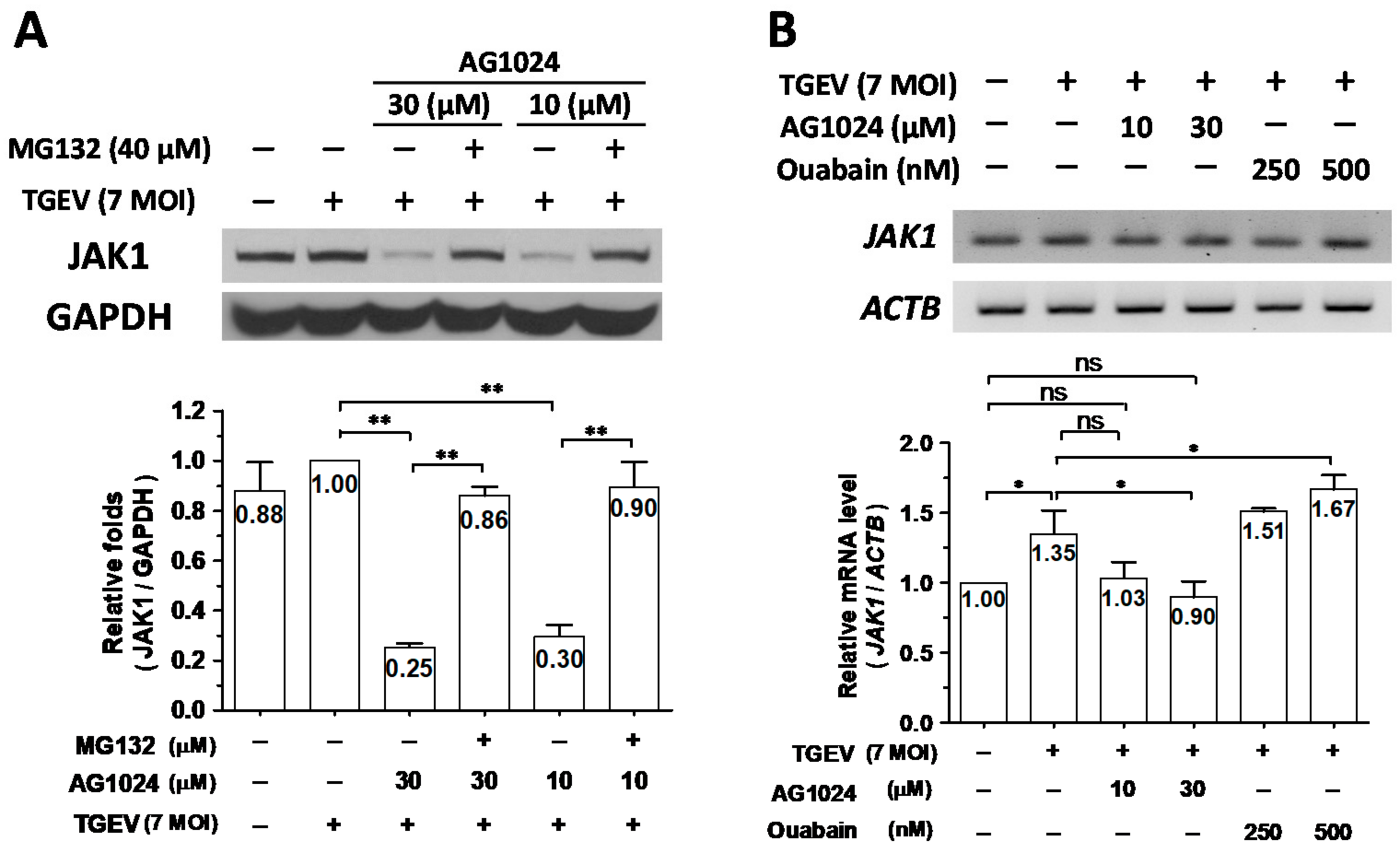
2.4. AG1024 Diminished JAK1 Protein Levels Mainly through Proteolysis
2.5. Downregulation of JAK1 by AG1024 Induced Proteolysis Was Associated with Ndfip1/2
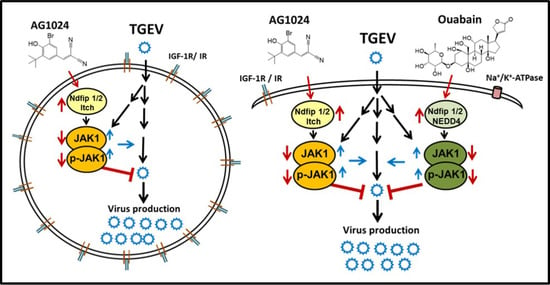
2.6. Downregulation of JAK1 by AG1024 Induced Proteolysis Was Mediated by NEDD4-like E3 Ligase Itch
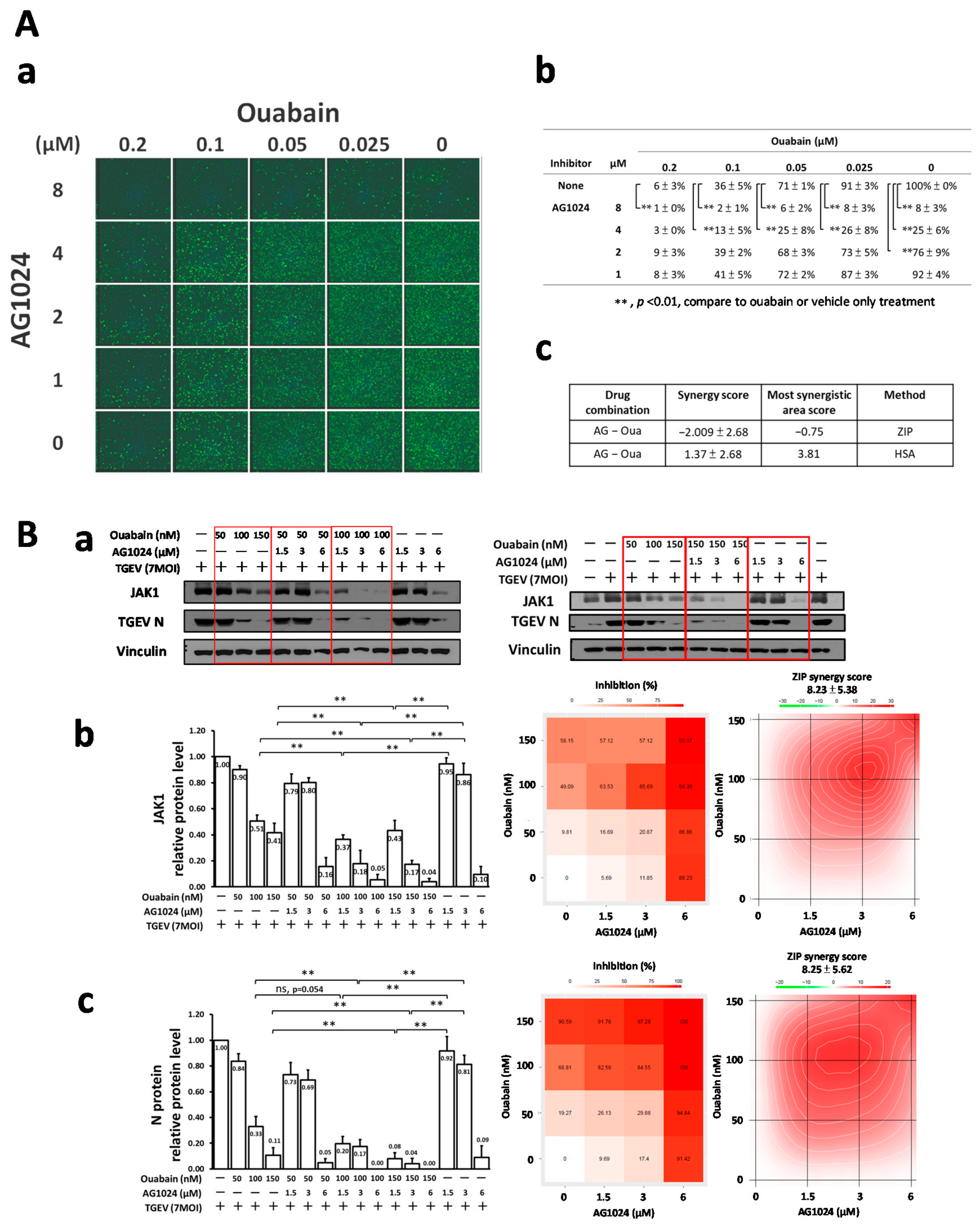
2.7. Combined Treatment of AG1024 and Ouabain Had an Additive Effect on Both Anti-Viral Activity and JAK1 Downregulation
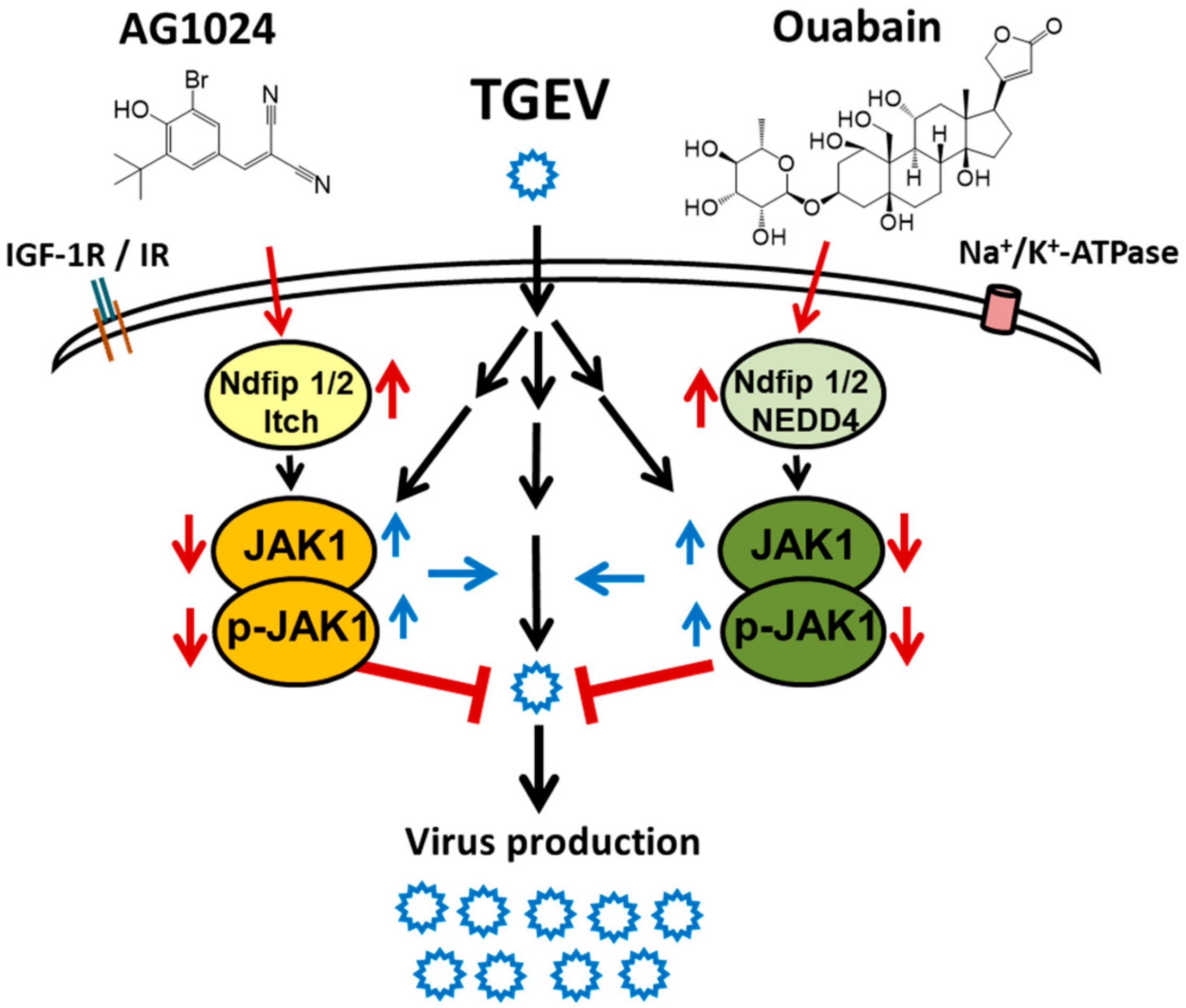
3. Discussions
4. Materials and Methods
4.1. Cells, Viruses, Immunofluorescence Assay (IFA), and Cytotoxicity Assays
4.2. Chemicals and Antibodies for Western Blot Analyses
4.3. RNA Isolation, Semi-Quantitative Reverse-Transcriptase Polymerase Chain Reaction (Semi-RT-PCR)
4.4. Gene Silence
4.5. Drug Combination Study
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Satarker, S.; Tom, A.A.; Shaji, R.A.; Alosious, A.; Luvis, M.; Nampoothiri, M. JAK-STAT Pathway Inhibition and their Implications in COVID-19 Therapy. Postgrad. Med. 2021, 133, 489–507. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Banerjee, S.; Mondal, A.; Chakraborty, U.; Pumarol, J.; Croley, C.R.; Bishayee, A. Targeting the JAK/STAT Signaling Pathway Using Phytocompounds for Cancer Prevention and Therapy. Cells 2020, 9, 1451. [Google Scholar] [CrossRef] [PubMed]
- Zouein, F.A.; Duhe, R.J.; Booz, G.W. JAKs go nuclear: Emerging role of nuclear JAK1 and JAK2 in gene expression and cell growth. Growth Factors 2011, 29, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Gadina, M.; Hilton, D.; Johnston, J.A.; Morinobu, A.; Lighvani, A.; Zhou, Y.J.; Visconti, R.; O’Shea, J.J. Signaling by type I and II cytokine receptors: Ten years after. Curr. Opin. Immunol. 2001, 13, 363–373. [Google Scholar] [CrossRef]
- O’Leary, C.E.; Riling, C.R.; Spruce, L.A.; Ding, H.; Kumar, S.; Deng, G.; Liu, Y.; Seeholzer, S.H.; Oliver, P.M. Ndfip-mediated degradation of Jak1 tunes cytokine signalling to limit expansion of CD4+ effector T cells. Nat. Commun. 2016, 7, 11226. [Google Scholar] [CrossRef]
- Lee, Y.; Hyung, S.W.; Jung, H.J.; Kim, H.J.; Staerk, J.; Constantinescu, S.N.; Chang, E.J.; Lee, Z.H.; Lee, S.W.; Kim, H.H. The ubiquitin-mediated degradation of Jak1 modulates osteoclastogenesis by limiting interferon-beta-induced inhibitory signaling. Blood 2008, 111, 885–893. [Google Scholar] [CrossRef]
- Wu, C.; Guan, Q.; Wang, Y.; Zhao, Z.J.; Zhou, G.W. SHP-1 suppresses cancer cell growth by promoting degradation of JAK kinases. J. Cell. Biochem. 2003, 90, 1026–1037. [Google Scholar] [CrossRef]
- Yang, C.W.; Hsu, H.Y.; Chang, H.Y.; Lee, Y.Z.; Lee, S.J. Natural cardenolides suppress coronaviral replication by downregulating JAK1 via a Na+/K+-ATPase independent proteolysis. Biochem. Pharmacol. 2020, 180, 114122. [Google Scholar] [CrossRef]
- Sluimer, J.; Distel, B. Regulating the human HECT E3 ligases. Cell. Mol. Life Sci. CMLS 2018, 75, 3121–3141. [Google Scholar] [CrossRef]
- Zou, X.; Levy-Cohen, G.; Blank, M. Molecular functions of NEDD4 E3 ubiquitin ligases in cancer. Biochim. Biophys. Acta BBA-Rev. Cancer 2015, 1856, 91–106. [Google Scholar] [CrossRef]
- An, H.; Krist, D.T.; Statsyuk, A.V. Crosstalk between kinases and Nedd4 family ubiquitin ligases. Mol. BioSyst. 2014, 10, 1643–1657. [Google Scholar] [CrossRef]
- George, A.J.; Hoffiz, Y.C.; Charles, A.J.; Zhu, Y.; Mabb, A.M. A Comprehensive Atlas of E3 Ubiquitin Ligase Mutations in Neurological Disorders. Front. Genet. 2018, 9, 29. [Google Scholar] [CrossRef]
- Párrizas, M.; Gazit, A.; Levitzki, A.; Wertheimer, E.; LeRoith, D. Specific inhibition of insulin-like growth factor-1 and insulin receptor tyrosine kinase activity and biological function by tyrphostins. Endocrinology 1997, 138, 1427–1433. [Google Scholar] [CrossRef]
- Deutsch, E.; Maggiorella, L.; Wen, B.; Bonnet, M.L.; Khanfir, K.; Frascogna, V.; Turhan, A.G.; Bourhis, J. Tyrosine kinase inhibitor AG1024 exerts antileukaemic effects on STI571-resistant Bcr-Abl expressing cells and decreases AKT phosphorylation. Br. J. Cancer 2004, 91, 1735–1741. [Google Scholar] [CrossRef][Green Version]
- Von Willebrand, M.; Zacksenhaus, E.; Cheng, E.; Glazer, P.; Halaban, R. The tyrphostin AG1024 accelerates the degradation of phosphorylated forms of retinoblastoma protein (pRb) and restores pRb tumor suppressive function in melanoma cells. Cancer Res. 2003, 63, 1420–1429. [Google Scholar]
- Nan, Y.; Wu, C.; Zhang, Y.-J. Interplay between Janus Kinase/Signal Transducer and Activator of Transcription Signaling Activated by Type I Interferons and Viral Antagonism. Front. Immunol. 2017, 8, 1758. [Google Scholar] [CrossRef]
- Johnson, K.E.; Knipe, D.M. Herpes simplex virus-1 infection causes the secretion of a type I interferon-antagonizing protein and inhibits signaling at or before Jak-1 activation. Virology 2010, 396, 21–29. [Google Scholar] [CrossRef]
- De-Simone, F.I.; Sariyer, R.; Otalora, Y.L.; Yarandi, S.; Craigie, M.; Gordon, J.; Sariyer, I.K. IFN-Gamma Inhibits JC Virus Replication in Glial Cells by Suppressing T-Antigen Expression. PLoS ONE 2015, 10, e0129694. [Google Scholar] [CrossRef]
- Van Tol, S.; Atkins, C.; Bharaj, P.; Johnson, K.N.; Hage, A.; Freiberg, A.N.; Rajsbaum, R. VAMP8 Contributes to the TRIM6-Mediated Type I Interferon Antiviral Response during West Nile Virus Infection. J. Virol. 2020, 94, e01454-19. [Google Scholar] [CrossRef]
- Lee, Y.H.; Yun, Y. HBx protein of hepatitis B virus activates Jak1-STAT signaling. J. Biol. Chem. 1998, 273, 25510–25515. [Google Scholar] [CrossRef]
- Chen, D.Y.; Khan, N.; Close, B.J.; Goel, R.K.; Blum, B.; Tavares, A.H.; Kenney, D.; Conway, H.L.; Ewoldt, J.K.; Chitalia, V.C.; et al. SARS-CoV-2 disrupts proximal elements in the JAK-STAT pathway. J. Virol. 2021, 95, e00862-21. [Google Scholar] [CrossRef]
- Stebbing, J.; Sánchez Nievas, G.; Falcone, M.; Youhanna, S.; Richardson, P.; Ottaviani, S.; Shen, J.X.; Sommerauer, C.; Tiseo, G.; Ghiadoni, L.; et al. JAK inhibition reduces SARS-CoV-2 liver infectivity and modulates inflammatory responses to reduce morbidity and mortality. Sci. Adv. 2021, 7, eabe4724. [Google Scholar] [CrossRef]
- Yang, C.W.; Chang, H.Y.; Lee, Y.Z.; Hsu, H.Y.; Lee, S.J. The cardenolide ouabain suppresses coronaviral replication via augmenting a Na+/K+-ATPase-dependent PI3K_PDK1 axis signaling. Toxicol. Appl. Pharmacol. 2018, 356, 90–97. [Google Scholar] [CrossRef]
- Soucy, T.A.; Smith, P.G.; Milhollen, M.A.; Berger, A.J.; Gavin, J.M.; Adhikari, S.; Brownell, J.E.; Burke, K.E.; Cardin, D.P.; Critchley, S.; et al. An inhibitor of NEDD8-activating enzyme as a new approach to treat cancer. Nature 2009, 458, 732–736. [Google Scholar] [CrossRef]
- Liu, Y.-M.; HuangFu, W.-C.; Huang, H.-L.; Wu, W.-C.; Chen, Y.-L.; Yen, Y.; Huang, H.-L.; Nien, C.-Y.; Lai, M.-J.; Pan, S.-L.; et al. 1,4-Naphthoquinones as inhibitors of Itch, a HECT domain-E3 ligase, and tumor growth suppressors in multiple myeloma. Eur. J. Med. Chem. 2017, 140, 84–91. [Google Scholar] [CrossRef]
- Rossi, M.; Rotblat, B.; Ansell, K.; Amelio, I.; Caraglia, M.; Misso, G.; Bernassola, F.; Cavasotto, C.N.; Knight, R.A.; Ciechanover, A.; et al. High throughput screening for inhibitors of the HECT ubiquitin E3 ligase ITCH identifies antidepressant drugs as regulators of autophagy. Cell Death Dis. 2014, 5, e1203. [Google Scholar] [CrossRef]
- Wu, L.; Grigoryan Arsen, V.; Li, Y.; Hao, B.; Pagano, M.; Cardozo Timothy, J. Specific Small Molecule Inhibitors of Skp2-Mediated p27 Degradation. Chem. Biol. 2012, 19, 1515–1524. [Google Scholar] [CrossRef]
- Yang, Y.; Yan, W.; Liu, Z.; Wei, M. Skp2 inhibitor SKPin C1 decreased viability and proliferation of multiple myeloma cells and induced apoptosis. Braz. J. Med. Biol. Res. 2019, 52, e8412. [Google Scholar] [CrossRef] [PubMed]
- Bjorklund, C.C.; Kang, J.; Amatangelo, M.; Polonskaia, A.; Katz, M.; Chiu, H.; Couto, S.; Wang, M.; Ren, Y.; Ortiz, M.; et al. Iberdomide (CC-220) is a potent cereblon E3 ligase modulator with antitumor and immunostimulatory activities in lenalidomide- and pomalidomide-resistant multiple myeloma cells with dysregulated CRBN. Leukemia 2020, 34, 1197–1201. [Google Scholar] [CrossRef] [PubMed]
- Witalisz-Siepracka, A.; Klein, K.; Prinz, D.; Leidenfrost, N.; Schabbauer, G.; Dohnal, A.; Sexl, V. Loss of JAK1 Drives Innate Immune Deficiency. Front. Immunol. 2018, 9, 3108. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Cui, D.; Chen, X.; Xiong, X.; Zhao, Y. PROTACs: An Emerging Targeting Technique for Protein Degradation in Drug Discovery. BioEssays 2018, 40, e1700247. [Google Scholar] [CrossRef]
- Hughes, P.; Marshall, D.; Reid, Y.; Parkes, H.; Gelber, C. The costs of using unauthenticated, over-passaged cell lines: How much more data do we need? Biotechniques 2007, 43, 575–586. [Google Scholar] [CrossRef]
- Yang, C.W.; Lee, Y.Z.; Kang, I.J.; Barnard, D.L.; Jan, J.T.; Lin, D.; Huang, C.W.; Yeh, T.K.; Chao, Y.S.; Lee, S.J. Identification of phenanthroindolizines and phenanthroquinolizidines as novel potent anti-coronaviral agents for porcine enteropathogenic coronavirus transmissible gastroenteritis virus and human severe acute respiratory syndrome coronavirus. Antivir. Res. 2010, 88, 160–168. [Google Scholar] [CrossRef]
- Hsu, H.Y.; Yang, C.W.; Lee, Y.Z.; Lin, Y.L.; Chang, S.Y.; Yang, R.B.; Liang, J.J.; Chao, T.L.; Liao, C.C.; Kao, H.C.; et al. Remdesivir and Cyclosporine Synergistically Inhibit the Human Coronaviruses OC43 and SARS-CoV-2. Front. Pharmacol. 2021, 12, 706901. [Google Scholar] [CrossRef]
- Yang, C.W.; Yang, Y.N.; Liang, P.H.; Chen, C.M.; Chen, W.L.; Chang, H.Y.; Chao, Y.S.; Lee, S.J. Novel small-molecule inhibitors of transmissible gastroenteritis virus. Antimicrob. Agents Chemother. 2007, 51, 3924–3931. [Google Scholar] [CrossRef]
- Yang, C.W.; Lee, Y.Z.; Hsu, H.Y.; Wu, C.M.; Chang, H.Y.; Chao, Y.S.; Lee, S.J. c-Jun-mediated anticancer mechanisms of tylophorine. Carcinogenesis 2013, 34, 1304–1314. [Google Scholar] [CrossRef]
- Yang, C.W.; Lee, Y.Z.; Hsu, H.Y.; Shih, C.; Chao, Y.S.; Chang, H.Y.; Lee, S.J. Targeting Coronaviral Replication and Cellular JAK2 Mediated Dominant NF-kappaB Activation for Comprehensive and Ultimate Inhibition of Coronaviral Activity. Sci. Rep. 2017, 7, 4105. [Google Scholar] [CrossRef]
- Yang, C.W.; Chang, H.Y.; Hsu, H.Y.; Lee, Y.Z.; Chang, H.S.; Chen, I.S.; Lee, S.J. Identification of anti-viral activity of the cardenolides, Na+/K+-ATPase inhibitors, against porcine transmissible gastroenteritis virus. Toxicol. Appl. Pharmacol. 2017, 332, 129–137. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ouabain (µM) | |||||
|---|---|---|---|---|---|
| Inhibitor | µM | 0.3 | 0.2 | 0.1 | 0 |
 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-W.; Lee, Y.-Z.; Hsu, H.-Y.; Zhao, G.-H.; Lee, S.-J. Tyrphostin AG1024 Suppresses Coronaviral Replication by Downregulating JAK1 via an IR/IGF-1R Independent Proteolysis Mediated by Ndfip1/2_NEDD4-like E3 Ligase Itch. Pharmaceuticals 2022, 15, 241. https://doi.org/10.3390/ph15020241
Yang C-W, Lee Y-Z, Hsu H-Y, Zhao G-H, Lee S-J. Tyrphostin AG1024 Suppresses Coronaviral Replication by Downregulating JAK1 via an IR/IGF-1R Independent Proteolysis Mediated by Ndfip1/2_NEDD4-like E3 Ligase Itch. Pharmaceuticals. 2022; 15(2):241. https://doi.org/10.3390/ph15020241
Chicago/Turabian StyleYang, Cheng-Wei, Yue-Zhi Lee, Hsing-Yu Hsu, Guan-Hao Zhao, and Shiow-Ju Lee. 2022. "Tyrphostin AG1024 Suppresses Coronaviral Replication by Downregulating JAK1 via an IR/IGF-1R Independent Proteolysis Mediated by Ndfip1/2_NEDD4-like E3 Ligase Itch" Pharmaceuticals 15, no. 2: 241. https://doi.org/10.3390/ph15020241
APA StyleYang, C.-W., Lee, Y.-Z., Hsu, H.-Y., Zhao, G.-H., & Lee, S.-J. (2022). Tyrphostin AG1024 Suppresses Coronaviral Replication by Downregulating JAK1 via an IR/IGF-1R Independent Proteolysis Mediated by Ndfip1/2_NEDD4-like E3 Ligase Itch. Pharmaceuticals, 15(2), 241. https://doi.org/10.3390/ph15020241