
Novel Single Inhibitor of HDAC6/8 and Dual Inhibitor of PI3K/HDAC6 as Potential Alternative Treatments for Prostate Cancer

 ,
,  and
and 
Abstract
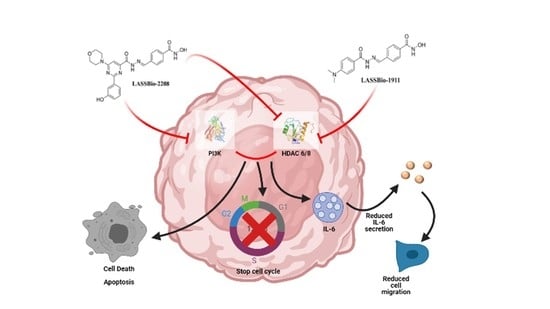
1. Introduction
2. Results
2.1. LASSBio-1911 and LASSBio-2208 Reduced the Viability of PC3 Prostate Cancer Cells
2.2. LASSBio-1911 and LASSBio-2208 Induced the Apoptosis of PC3 Cells
2.3. Effects of LASSBio-1911 and LASSBio-2208 on Cell Cycle Distribution
2.4. LASSBio-1911 and LASSBio-2208 Inhibited PC3 Cell Proliferation via the JNK Pathway
2.5. LASSBio-2208 Inhibited JAK/STAT3 Phosphorylation in PC3 Cells
2.6. LASSBio-1911 and LASSBio-2208 Inhibited IL-6 Secretion from PC3 Cells
2.7. Effects of LASSBio-1911 and LASSBio-2208 on the Motility of Prostate Cancer Cells
3. Discussion
4. Materials and Methods
4.1. LASSBio-1911 and LASSBio-2208 Synthesis
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Cell Cycle Analysis
4.5. Apoptosis Assay
4.6. Cell Scratch Assay
4.7. Western Blot Analysis
4.8. IL-6 Quantification
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rawla, P. Epidemiology of prostate cancer. World J. Oncol. 2019, 10, 63–89. [Google Scholar] [CrossRef] [PubMed]
- Scher, H.I.; Sawyers, C.L. Biology of progressive, castration-resistant prostate cancer: Directed therapies targeting the androgen-receptor signaling axis. J. Clin. Oncol. 2005, 23, 8253–8261. [Google Scholar] [CrossRef] [PubMed]
- Kaliks, R.A.; Del Giglio, A. Management of advanced prostate cancer. Rev. Assoc. Med. Bras. 2008, 54, 178–182. [Google Scholar] [CrossRef][Green Version]
- Vara, J.A.F.; Casado, E.; de Castro, J.; Cejas, P.; Belda-Iniesta, C.; González-Barón, M. PI3K/Akt signalling pathway and cancer. Cancer Treat. Rev. 2004, 30, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Hanker, A.B.; Kaklamani, V.; Arteaga, C.L. Challenges for the clinical development of pi3k inhibitors: Strategies to improve their impact in solid tumors. Cancer Disc. 2019, 9, 482–491. [Google Scholar] [CrossRef]
- Shaik, B.; Zafar, T.; Gupta, S.P. Recent studies on HDAC inhibitors acting against prostate cancer. Curr. Enz. Inhibition 2002, 16, 74–89. [Google Scholar] [CrossRef]
- Aldana-Masangkay, G.I.; Sakamoto, K.M. The role of HDAC6 in cancer. J. Biomed. Biotechnol. 2011, 2011, 875824. [Google Scholar] [CrossRef]
- Robey, R.W.; Chakraborty, A.R.; Basseville, A.; Luchenko, V.; Bahr, J.; Zhan, Z.; Bates, S.E. Histone deacetylase inhibitors: Emerging mechanisms of resistance. Mol. Pharm. 2011, 8, 2021–2031. [Google Scholar] [CrossRef]
- Seidel, C.M.; Schnekenburger, M.; Dicato, M.; Diederich, M. Histone deacetylase 6 in health and disease. Epigenomics 2015, 7, 103–118. [Google Scholar] [CrossRef]
- Gupta, M.; Ansell, S.M.; Novak, A.J.; Kumar, S.; Kaufmann, S.H.; Witzig, T.E. Inhibition of histone deacetylase overcomes rapamycin-mediated resistance in diffuse large B-cell lymphoma by inhibiting Akt signaling through mTORC2. Blood 2009, 114, 2926–2935. [Google Scholar] [CrossRef]
- Saijo, K.; Imamura, J.; Narita, K.; Oda, A.; Shimodaira, H.; Katoh, T.; Ishioka, C. Biochemical, biological and structural properties of romidepsin (FK228) and its analogs as novel HDAC/PI3K dual inhibitors. Cancer Sci. 2015, 106, 208–215. [Google Scholar] [CrossRef]
- Xu, R.; Hu, J. The role of JNK in prostate cancer progression and therapeutic strategies. Biomed. Pharmacother. 2020, 121, 109679. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Jiang, C.; Pei, H.; Wang, L.; Zhang, J.; Hu, H.; Lü, J. In vivo molecular mediators of cancer growth suppression and apoptosis by selenium in mammary and prostate models: Lack of involvement of gadd genes. Mol. Cancer Ther. 2009, 8, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Vacas, J.M.; Herrero-Aguayo, V.; Gómez-Gómez, E.; León-González, A.J.; Sáez-Martínez, P.; Alors-Pérez, E.; Fuentes-Fayos, A.C.; Martínez-López, A.; Sánchez-Sánchez, R.; González-Serrano, T.; et al. Spliceosome component SF3B1 as novel prognostic biomarker and therapeutic target for prostate cancer. Transl. Res. 2019, 212, 89–103. [Google Scholar] [CrossRef]
- Hodge, D.R.; Hurt, E.M.; Farrar, W.L. The role of IL-6 and STAT3 in inflammation and cancer. Eur. J. Cancer. 2005, 41, 2502–2512. [Google Scholar] [CrossRef]
- Wang, S.W.; Sun, Y.M. The IL-6/JAK/STAT3 pathway: Potential therapeutic strategies in treating colorectal cancer. Int. J. Oncol. 2014, 44, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.A.; Guerra, F.S.; Sagrillo, F.S.; de Sena, M.; Pinheiro, P.; Alves, M.A.; Thota, S.; Chaves, L.S.; Sant’Anna, C.M.R.; Fernandes, P.D.; et al. Design, synthesis, and pharmacological evaluation of first-in-class multitarget n-acylhydrazone derivatives as selective HDAC6/8 and PI3Kα inhibitors. ChemMedChem 2020, 15, 539–551. [Google Scholar] [CrossRef]
- Kaliszczak, M.; van Hechanova, E.; Li, Y.; Alsadah, H.; Parzych, K.; Auner, H.W.; Aboagye, E.O. The HDAC6 inhibitor C1A modulates autophagy substrates in diverse cancer cells and induces cell death. Br. J. Cancer 2018, 119, 1278–1287. [Google Scholar] [CrossRef] [PubMed]
- Deskin, B.; Yin, Q.; Zhuang, Y.; Saito, S.; Shan, B.; Lasky, J.A. Inhibition of HDAC6 attenuates tumor growth of non-small cell lung cancer. Transl. Oncol. 2020, 13, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Foukas, L.C.; Berenjeno, I.M.; Gray, A.; Khwaja, A.; Vanhaesebroeck, B. Activity of any class IA PI3K isoform can sustain cell proliferation and survival. Proc. Nat. Acad. Sci. USA 2010, 107, 11381–11386. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; SONG, L. Applicability of the MTT assay for measuring viability of cyanobacteria and algae, specifically for Microcystis aeruginosa (Chroococcales, Cyanobacteria). Phycologia 2007, 46, 593–599. [Google Scholar] [CrossRef]
- Rai, Y.; Pathak, R.; Kumari, N.; Sah, D.K.; Pandey, S.; Kalra, N.; Soni, R.; Dwarakanath, B.S.; Bhatt, A.N. Mitochondrial biogenesis and metabolic hyperactivation limits the application of MTT assay in the estimation of radiation induced growth inhibition. Sci. Rep. 2018, 8, 1531. [Google Scholar] [CrossRef] [PubMed]
- Jabbar, S.A.; Twentyman, P.R.; Watson, J.V. The MTT assay underestimates the growth inhibitory effects of interferons. Br. J. Cancer. 1989, 60, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, S.; Liu, Z.; Ma, C.; Jiang, Y.; Sun, C.; Li, K.; Cao, G.; Lin, Z.; Wang, P.; et al. Polyphyllin I induces cell cycle arrest in prostate cancer cells via the upregulation of IL6 and P21 expression. Medicine 2019, 98, e17743. [Google Scholar] [CrossRef]
- Cao, J.; Lv, W.; Wang, L.; Xu, J.; Yuan, P.; Huang, S.; He, Z.; Hu, J. Ricolinostat (ACY-1215) suppresses proliferation and promotes apoptosis in esophageal squamous cell carcinoma via miR-30d/PI3K/AKT/mTOR and ERK pathways. Cell Death Dis. 2018, 9, 817. [Google Scholar] [CrossRef]
- To, K.K.W.; Fu, L.W. CUDC-907, a dual HDAC and PI3K inhibitor, reverses platinum drug resistance. Investig. New Drugs 2018, 36, 10–19. [Google Scholar] [CrossRef]
- Wasim, L.; Chopra, M. Synergistic anticancer effect of panobinostat and topoisomerase inhibitors through ROS generation and intrinsic apoptotic pathway induction in cervical cancer cells. Cell. Oncol. 2018, 41, 201–212. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, G.W.; Kwon, S.H. The HDAC6-selective inhibitor is effective against non-Hodgkin lymphoma and synergizes with ibrutinib in follicular lymphoma. Mol. Carcinog. 2019, 58, 944–956. [Google Scholar] [CrossRef]
- Kotian, S.L.; Zhang, M.; Boufraqech, M.; Gaskins, K.; Gara, S.K.; Quezado, M.; Nilubol, N.; Kebebew, E. Dual inhibition of HDAC and tyrosine kinase signaling pathways with CUDC-907 inhibits thyroid cancer growth and metastases. Clin. Cancer Res. 2017, 23, 5044–5054. [Google Scholar] [CrossRef]
- Bubici, C.; Papa, S. JNK signalling in cancer: In need of new, smarter therapeutic targets. Br. J. Pharmacol. 2014, 171, 24–37. [Google Scholar] [CrossRef]
- Zhao, H.F.; Wang, J.; Jiang, H.R.; Chen, Z.P.; To, S.S.T. PI3K p110β isoform synergizes with JNK in the regulation of glioblastoma cell proliferation and migration through Akt and FAK inhibition. J. Exp. Clin. Cancer Res. 2016, 35, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.F.; Wang, J.; Jiang, H.R.; Chen, Z.P.; To, S.S. Tanshinone IIA induces apoptosis of ovarian cancer cells in vitro and in vivo through attenuation of PI3K/AKT/JNK signaling pathways. Oncol. Lett. 2019, 17, 1896–1902. [Google Scholar]
- Bousoik, E.; Montazeri Aliabadi, H. “Do we know jack” about JAK? A closer look at JAK/STAT signalling pathway. Front. Oncol. 2018, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Huang, W.; Chen, B.; Bai, P.D.; Wang, X.G.; Xing, J.C. Up-regulation of miR-124inhibits invasion and proliferation of prostate cancer cells through mediating JAK-STAT3 signalling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 2338–2345. [Google Scholar]
- Abdulghani, J.; Gu, L.; Dagvadorj, A.; Lutz, J.; Leiby, B.; Bonuccelli, G.; Lisanti, M.P.; Zellweger, T.; Alanen, K.; Mirtti, T.; et al. Stat3 promotes metastatic progression of prostate cancer. Am. J. Pathol. 2008, 172, 1717–1728. [Google Scholar] [CrossRef] [PubMed]
- Fantin, V.R.; Loboda, A.; Paweletz, C.P.; Hendrickson, R.C.; Pierce, J.W.; Roth, J.A.; Li, L.; Gooden, F.; Korenchuk, S.; Hou, X.S.; et al. Constitutive activation of signal transducers and activators of transcription predicts vorinostat resistance in cutaneous T-cell lymphoma. Cancer Res. 2008, 68, 3785–3794. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, P.C.; Behrmann, I.; Haan, S.; Hermanns, H.M.; Müller-Newen, G.; Schaper, F. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem. J. 2003, 374, 1–20. [Google Scholar] [CrossRef]
- Chang, Q.; Bournazou, E.; Sansone, P.; Berishaj, M.; Gao, S.P.; Daly, L.; Wels, J.; Theilen, T.; Granitto, S.; Zhang, X.; et al. The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and metastasis. Neoplasia 2013, 15, 848–862. [Google Scholar] [CrossRef]
- Tsukamoto, H.; Kubota, K.; Shichiku, A.; Maekawa, M.; Mano, N.; Yagita, H.; Ohta, S.; Tomioka, Y. An agonistic anti-Toll-like receptor 4 monoclonal antibody as an effective adjuvant for cancer immunotherapy. Immunology 2019, 158, 136–149. [Google Scholar] [CrossRef]
- Pei, Z.; Lin, D.; Song, X.; Li, H.; Yao, H. TLR4 signaling promotes the expression of VEGF and TGFbeta1 in human prostate epithelial PC3 cells induced by lipopolysaccharide. Cell Immunol. 2008, 254, 20–27. [Google Scholar] [CrossRef]
- Shukla, S.; Maclennan, G.T.; Hartman, D.J.; Fu, P.; Resnick, M.I.; Gupta, S. Activation of PI3K-Akt signaling pathway promotes prostate cancer cell invasion. Int. J. Cancer 2007, 121, 1424–1432. [Google Scholar] [CrossRef]
- Pan, Y.R.; Chen, C.C.; Chan, Y.T.; Wang, H.J.; Chien, F.T.; Chen, Y.L.; Liu, J.L.; Yang, M.H. STAT3-coordinated migration facilitates the dissemination of diffuse large B-cell lymphomas. Nat. Commun. 2018, 9, 3696. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Z.; Luo, Y.R.; Du, J.; Yu, Y.; Yang, X.Z.; Cui, Y.J.; Jin, X.F. MiR-296-5p inhibits cell invasion and migration of esophageal squamous cell carcinoma by downregulating STAT3 signaling. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5206–5214. [Google Scholar]
- Denizot, F.; Lang, R. Rapid colorimetric assay for cell growth and survival. Modifications to the tetrazolium dye procedure giving improved sensitivity and reliability. J. Immunol. Methods 1986, 89, 271–277. [Google Scholar] [CrossRef]
- Guerra, F.S.; Sampaio, L.D.S.; Konig, S.; Bonamino, M.; Rossi, M.I.D.; Costa, M.L.; Fernandes, P.D.; Mermelstein, C.S. Membrane cholesterol depletion reduces breast tumor cell migration by a mechanism that involves non-canonical Wnt signaling and IL-10 secretion. Transl. Med. Commun. 2016, 1, 3–13. [Google Scholar] [CrossRef]
- Guerra, F.S.; Oliveira, R.G.; Fraga, C.A.M.; Mermelstein, C.S.; Fernandes, P.D. ROCK inhibition with Fasudil induces beta-catenin nuclear translocation and inhibits cell migration of MDA-MB 231 human breast cancer cells. Sci. Rep. 2017, 7, 13723. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.I.; Fernandes, P.D.; Amarantes-Mendes, S.G.P.; Jancar, S. Phagocytosis of apoptotic and necrotic thymocytes is inhibited by PAF-receptor antagonists and affects LPS-induced COX-2 expression in murine macrophages. Prostaglandins Lipid Med. 2006, 80, 62–73. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Cycle Phases (%) | Concentration | G0/G1 | S | G2 + M |
|---|---|---|---|---|
| Control | - | 13.2 ± 2.49 | 43.8 ± 5.57 | 31.09 ±9.0 |
| Vehicle | - | 12.6 ± 2.8 | 38.6 ± 2.8 | 49.6 ± 9.7 |
| LASSBio-1911 | 0.1 µM | 2.86 ± 0.8 | 3.2 ± 1.1 | 82.1 ± 3.9 * |
| 1 µM | 3.46 ± 0.9 | 3.1 ± 0.8 | 81.8 ± 4.0 * | |
| 5 µM | 2.3 ± 0.7 | 2.9 ± 0.87 | 82.1 ± 4.8 * | |
| LASSBio-2208 | 0.1 µM | 3.2 ± 0.1 | 3.4 ± 0.3 | 81.0 ± 2.4 * |
| 1 µM | 2.9 ± 0.75 | 2.9 ± 0.8 | 81.0 ± 5.5 * | |
| 5 µM | 2.63 ± 0.2 | 3.9 ± 0.7 | 81.0 ± 4.0 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, F.S.; Rodrigues, D.A.; Fraga, C.A.M.; Fernandes, P.D. Novel Single Inhibitor of HDAC6/8 and Dual Inhibitor of PI3K/HDAC6 as Potential Alternative Treatments for Prostate Cancer. Pharmaceuticals 2021, 14, 387. https://doi.org/10.3390/ph14050387
Guerra FS, Rodrigues DA, Fraga CAM, Fernandes PD. Novel Single Inhibitor of HDAC6/8 and Dual Inhibitor of PI3K/HDAC6 as Potential Alternative Treatments for Prostate Cancer. Pharmaceuticals. 2021; 14(5):387. https://doi.org/10.3390/ph14050387
Chicago/Turabian StyleGuerra, Fabiana Sélos, Daniel Alencar Rodrigues, Carlos Alberto Manssour Fraga, and Patricia Dias Fernandes. 2021. "Novel Single Inhibitor of HDAC6/8 and Dual Inhibitor of PI3K/HDAC6 as Potential Alternative Treatments for Prostate Cancer" Pharmaceuticals 14, no. 5: 387. https://doi.org/10.3390/ph14050387
APA StyleGuerra, F. S., Rodrigues, D. A., Fraga, C. A. M., & Fernandes, P. D. (2021). Novel Single Inhibitor of HDAC6/8 and Dual Inhibitor of PI3K/HDAC6 as Potential Alternative Treatments for Prostate Cancer. Pharmaceuticals, 14(5), 387. https://doi.org/10.3390/ph14050387