Iron Supplementation in Suckling Piglets: An Ostensibly Easy Therapy of Neonatal Iron Deficiency Anemia

 , ,
, , {kind=link}
{kind=link}
Abstract
1. Iron Deficiency Anemia (IDA) in Suckling Piglets
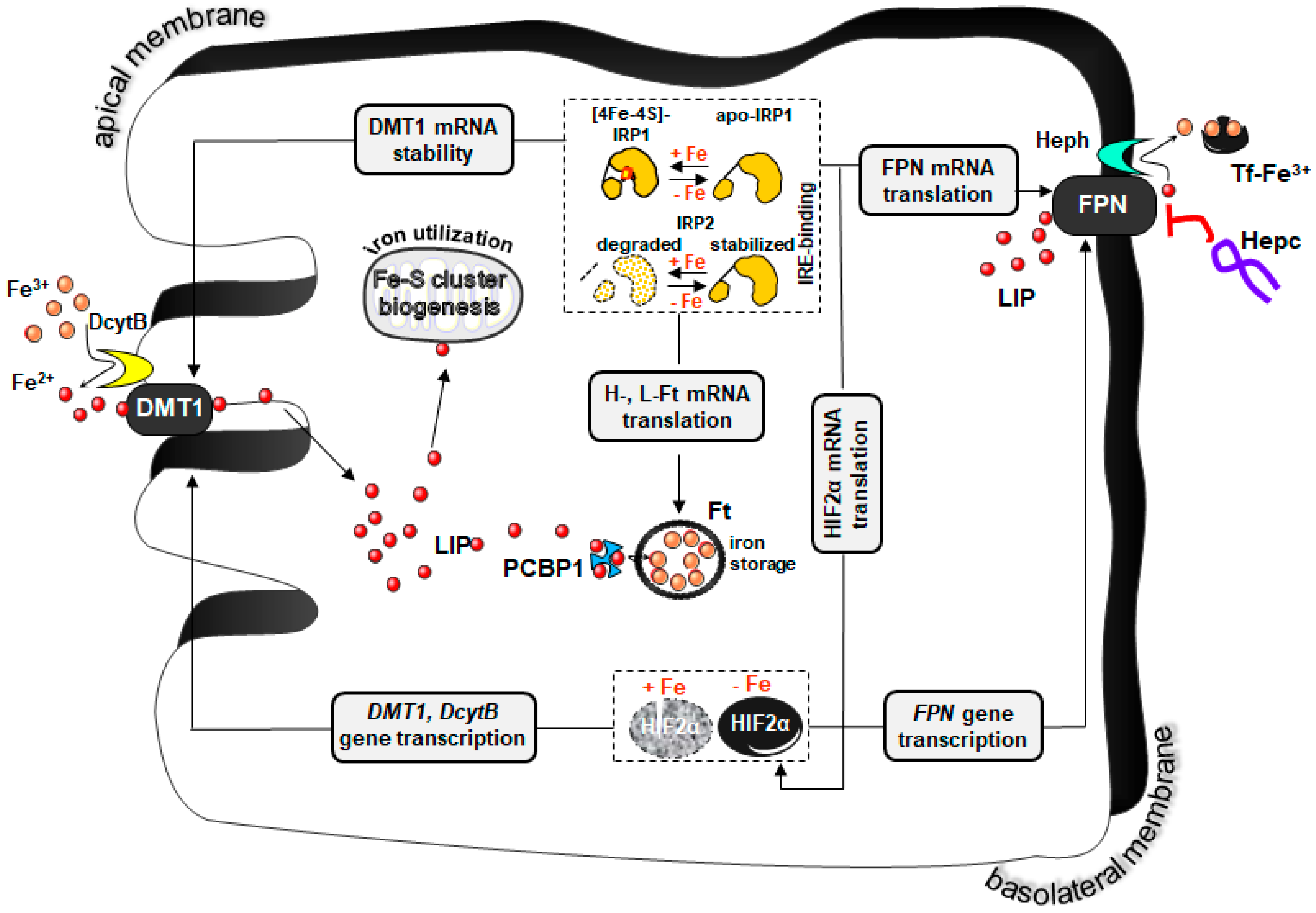
Etiology of IDA in Suckling Piglets
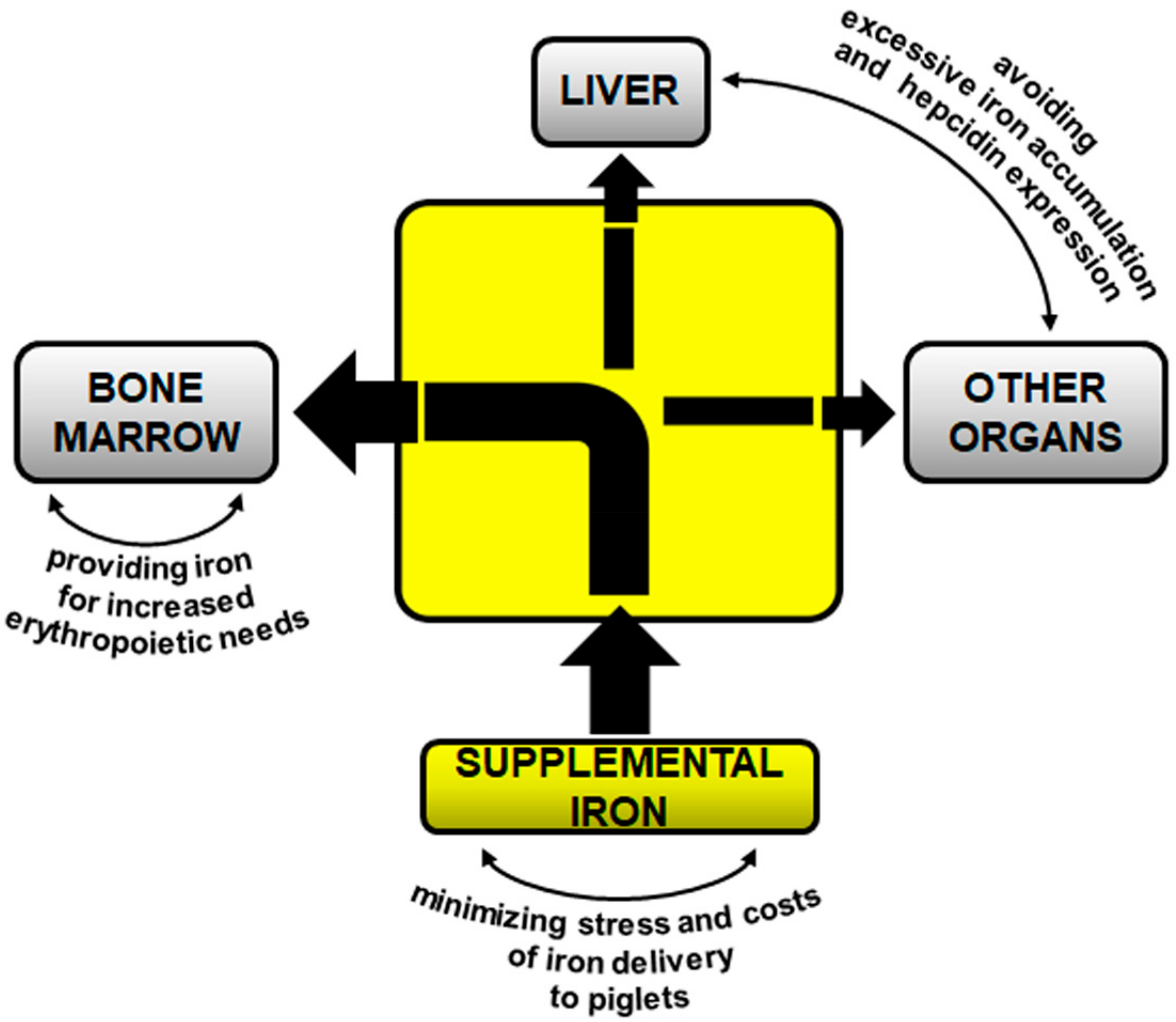
2. Iron Supplementation of Piglets
2.1. Intramuscular Supplementation with Iron Dextran (FeDex)—A Gold Standard?
2.2. Oral Supplementation
3. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Camaschella, C. New insights into iron deficiency and iron deficiency anemia. Blood Rev. 2016, 31, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Strauss, R.G. Anaemia of prematurity: Pathophysiology and treatment. Blood Rev. 2010, 24, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Venn, J.A.J.; McCance, R.A.; Widdowson, E.M. Iron metabolism in piglet anemia. J. Comp. Pathol. 1947, 5, 314–325. [Google Scholar] [CrossRef]
- Svoboda, M.; Drabek, J. Iron deficiency in suckling piglets; ethiology, clinical aspects and diagnosis. Folia Vet. 2005, 49, 104–111. [Google Scholar]
- Kim, J.C.; Wilcock, P.; Bedford, M.R. Iron status of piglets and impact of phytase superdosing on iron physiology: A review. Anim. Feed Sci. Technol. 2018, 235, 8–14. [Google Scholar] [CrossRef]
- Boussingault, J.B. Du ironr contenu dans le sang et dans les aliments. Acad. Sci. Parts Rend. Acad. Sci. 1872, 74, 1353. [Google Scholar]
- McGovan, J.P.; Crichton, A. Iron deficiency in pigs. Biochem. J. 1924, 18, 265–272. [Google Scholar] [CrossRef]
- Hart, E.B.C.; Elvehjem, C.A.; Steenbock, H.; Bohstedt, G.; Fargo, J.M. Anemia in suckling pigs. J. Anim. Sci. 1929, 409. [Google Scholar] [CrossRef]
- Svoboda, M.; Vaňhara, J.; Berlinská, J. Parenteral iron administration in suckling piglets—A review. Acta Vet. Brno 2017, 86, 249–261. [Google Scholar] [CrossRef]
- Svoboda, M.; Pískova, K. Oral iron administration in suckling piglets—A review. Acta Vet. Brno 2018, 87, 77–83. [Google Scholar] [CrossRef]
- Godyń, D.; Pieszka, M.; Lipiński, P.; Starzyński, R.R. Diagnostics of iron deficiency anaemia in piglets in the early postnatal period—A review. Anim. Sci. Pap. Rep. 2016, 34, 1–12. [Google Scholar]
- Egeli, A.K.; Framstad, T.; Morberg, H. Clinical biochemistry, haematology and body weight in piglets. Acta Vet. Scand. 1998, 39, 381–393. [Google Scholar] [PubMed]
- Kegley, E.B.; Spears, J.W.; Flowers, W.L.; Schoenherr, W.D. Iron methionine as a source of iron for the neonatal pig. Nutr. Res. 2002, 22, 1209–1217. [Google Scholar] [CrossRef]
- Steinhardt, M.; Bünger, U.; Furcht, G.; Schönfelder, E. Determination of normal red blood picture values in piglets. Arch. Exp. Veterinarmed. 1982, 36, 707–719. [Google Scholar] [PubMed]
- Lipiński, P.; Starzyński, R.R.; Canonne-Hergaux, B.; Tudek, R.; Oliński, P.; Kowalczyk, P.; Dziamam, T.; Thibaudeau, O.; Gralak, M.A.; Smuda, E.; et al. Benefits and risks of iron supplementation in anemic neonatal pigs. Am. J. Pathol. 2010, 177, 1233–1243. [Google Scholar] [CrossRef] [PubMed]
- Staroń, R.; Van Swelm, R.P.L.; Lipiński, P.; Gajowiak, A.; Lenartowicz, M.; Bednarz, A.; Gajewska, M.; Pieszka, M.; Laarakkers, C.M.; Swinkels, D.W.; Starzyński, R.R. Urinary hepcidin levels in iron-deficient and iron-supplemented piglets correlate with hepcidin hepatic mRNA and serum levels and with body iron status. PLoS ONE 2015, 10, e0136695. [Google Scholar] [CrossRef] [PubMed]
- Staroń, R.; Lipiński, P.; Lenartowicz, M.; Bednarz, A.; Gajowiak, A.; Smuda, E.; Krzeptowski, W.; Pieszka, M.; Korolonek, T.; Hamza, I.; et al. Dietary hemoglobin rescues young piglets from severe. iron deficiency anemia: Duodenal expression profile of genes involved in heme iron absorption. PLoS ONE 2017, 12, e0181117. [Google Scholar]
- Egeli, A.K.; Framstad, T. An evaluation of iron-dextran supplementation in piglets administered by injection on the first, third or fourth day after birth. Res. Vet. Sci. 1999, 66, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Rincker, M.J.; Clarke, S.L.; Eisenstein, R.S.; Link, J.E.; Hill, G.M. Effects of iron supplementation on binding activity of iron regulatory proteins and the subsequent effect on growth performance and indices of hematological and mineral status of young pigs. J. Anim. Sci 2005, 83, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Jolliff, J.S.; Mahan, D.C. Effect of injected and dietary iron in young pigs on blood hematology and postnatal pig growth performance. J. Anim. Sci. 2011, 89, 4068–4080. [Google Scholar] [CrossRef] [PubMed]
- Szabo, P.; Bilkei, G. Iron deficiency in outdoor pig production. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2002, 49, 390–391. [Google Scholar] [CrossRef] [PubMed]
- Rao, R.; Georgieff, M.K. Iron in fetal and neonatal nutrition. Semin. Fetal Neonatal Med. 2007, 12, 54–63. [Google Scholar] [CrossRef] [PubMed]
- McPherson, R.L.; Ji, F.; Wu, G.; Blanton, J.R.; Kim, S.W. Growth and compositional changes of fetal tissues in pigs. J. Anim. Sci 2004, 82, 2534–2540. [Google Scholar] [CrossRef] [PubMed]
- Polish Pig Breeders and Producers Association “POLSUS”. Performance Results Pigs. 2017, pp. 17–21. Available online: https://polsus.pl/images/photos/Wydawnictwa/Wyniki%20Oceny/2017/WynikiOcenyTCh2017.pdf (accessed on 22 November 2018).
- Fernández-Llario, P.; Mateos-Quesada, P. Body size and reproductive parameters in the wild boar Sus scrofa. Acta Theriol. 1998, 43, 439–444. [Google Scholar] [CrossRef]
- Larson, G.; Dobney, K.; Albarella, U.; Fang, M.; Matisoo-Smith, E.; Robins, J.; Lowden, S.; Finlayson, H.; Brand, T.; Willerslev, E.; et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication. Science. 2005, 307, 1618–1621. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Fleming, M.D. The placenta: The forgotten essential organ of iron transport. Nutr. Rev. 2016, 74, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Brady, P.S.; Ku, P.K.; Ullrey, D.E.; Miller, E.R. Evaluation of an amino acid-iron chelate hematinic for the baby pig. J. Anim. Sci. 1978, 47, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Pfau, A.; Rudoplhi, K. Modelluntersuchungen zur oralen Eisensupplementierung beim Saugferkel. Zuchtungskd. 1978, 50, 227–233. [Google Scholar]
- Braude, R.; Chamberlain, A.G.; Kotarbińska, M.; Mitchell, K.G. The metabolism of iron in piglets given labelled iron either orally or by injection. Br. J. Nutr. 1962, 16, 427–449. [Google Scholar] [CrossRef] [PubMed]
- Pond, W.G.; Lowery, R. S.; Maner, J.H.; Looslu, J.K. Parental iron administration to sows during gestation and lactation. J. Anim. Sci. 1961, 20, 747–750. [Google Scholar] [CrossRef]
- Leong, W.I.; Bowlus, C.L.; Tallkvist, J.; Lönnerdal, B. DMT1 and FPN1 expression during infancy: Developmental regulationof iron absorption. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G1153–G1161. [Google Scholar] [CrossRef] [PubMed]
- Leong, W.I.; Bowlus, C.L.; Tallkvist, J.; Lönnerdal, B. Iron supplementation during infancy–effects on expression of iron transporters, iron absorption, and iron utilization in rat pups. Am. J. Clin. Nutr. 2003, 8, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Lopez, V.; Suzuki, Y.A.; Lönnerdal, B. Ontogenic changes in lactoferrin receptor and DMT1 in mouse small intestine: Implications for iron absorption during early life. Biochem. Cell Biol. 2006, 84, 337–344. [Google Scholar]
- Gunshin, H.; MacKenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef] [PubMed]
- McKie, A.T. The role of Dcytb in iron metabolism: An update. Biochem. Soc. Trans. 2008, 36, 1239–1241. [Google Scholar] [CrossRef] [PubMed]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Vulpe, C.D.; Kuo, Y.M.; Murphy, T.L.; Cowley, L.; Askwith, C.; Libina, N.; Gitschier, J.; Anderson, G.J. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat. Genet. 1999, 21, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Gunshin, H.; Starr, C.N.; Direnzo, C.; Fleming, M.D.; Jin, J.; Greer, E.L.; Sellers, V.M.; Galica, S.M.; Andrews, N.C. Cybrd1 (duodenal cytochrome b) is not necessary for dietary iron absorption in mice. Blood 2005, 106, 2879–2883. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochim. Biophys. Acta. 2012, 1823, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science. 2004, 306, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Vanoaica, L.; Darshan, D.; Richman, L.; Schümann, K.; Kühn, L.C. Intestinal ferritin H is required for an accurate control of iron absorption. Cell Metab. 2010, 12, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Galy, B.; Ferring-Appel, D.; Becker, C.; Gretz, N.; Gröne, H.J.; Schümann, K.; Hentze, M.W. Iron regulatory proteins control a mucosal block to intestinal ironabsorption. Cell Rep. 2013, 3, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Granick, S. Ferritin; increase of the protein apoferritin in the gastrointestinal mucosa as a direct response to iron feeding; the function of ferritin in the regulation of iron absorption. J. Biol. Chem. 1946, 164, 737–746. [Google Scholar] [PubMed]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Biophys. Acta 1996, 1275, 161–203. [Google Scholar] [CrossRef]
- Mastrogiannaki, M.; Matak, P.; Keith, B.; Simon, M.C.; Vaulont, S.; Peyssonnaux, C. HIF-2alpha, but not HIF-1alpha, promotes iron absorption in mice. J. Clin. Investig. 2009, 119, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Mastrogiannaki, M.; Matak, P.; Peyssonnaux, C. The gut in iron homeostasis: Role of HIF-2 under normal and pathological conditions. Blood 2013, 122, 885–892. [Google Scholar] [CrossRef] [PubMed]
- Manzoni, P.; Dall’Agnola, A.; Tomé, D.; Kaufman, D.A.; Tavella, E.; Pieretto, M.; Messina, A.; De Luca, D.; Bellaiche, M.; Mosca, A.; et al. Role of lactoferrin in neonates and infants: An update. Am. J. Perinatol. 2018, 35, 561–565. [Google Scholar] [PubMed]
- Suzuki, Y.A.; Shin, K.; Lönnerdal, B. Molecular cloning and functional expression of a human intestinal lactoferrin receptor. Biochemistry. 2001, 40, 15771–15779. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Lopez, V.; Shafizadeh, T.B.; Halsted, C.H.; Lönnerdal, B. Cloning of a pig homologue of the human lactoferrinreceptor: Expression and localization during intestinal maturationin piglets. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2007, 148, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Yu, I.T.; Lin, J.; Wu, J.F.; Yen, H.T.; Lee, S.L.; Yang, T.S. Reevaluation of the necessity of iron injection to newborn piglets. Asian-Australas. Anim. Sci. 2002, 15, 79–83. [Google Scholar] [CrossRef]
- Geisser, P.; Baer, M.; Schaub, E. Structure/histotoxicity relationship of parenteral iron preparations. Arzneimittelforschung 1992, 42, 1439–1452. [Google Scholar] [PubMed]
- Katkiewicz, M.; Malicka, E.; Preibisch, J. Effect of iron overload on the morphology of some organs in piglets. Pol. Arch. Vet. 1986, 25, 75. [Google Scholar]
- Kruszewski, M.; Iwaneńko, T.; Bartłomiejczyk, T.; Woliński, J.; Starzyński, R.R.; Gralak, M.A.; Zabielski, R.; Lipiński, P. Hepatic iron content corresponds with the susceptibility of lymphocytes to oxidative stress in neonatal pigs. Mutat. Res. 2008, 657, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Starzyński, R.; Szudzik, M.; Staroń, R.; Jonczy, A.; Smuda, E.; Pieszka, M.; Kamyczek, M.; Lipiński, P. Comparison of the therapeutical potential of oral sucrosomial iron and parenteral iron dextran supplementations in neonatal iron deficiency anemia in pigs. Am. J. Hematol. 2017, 92, E286–E286. [Google Scholar]
- Velásquez, J.I.; Aranzazu, D. An acute case of iron toxicity on newborn piglets from vitamin E/Se deficient sows. Rev. Colomb. Cienc. Pecu. 2004, 17, 60–62. [Google Scholar]
- Ueberschär, S. Sudden death in suckling piglets following administration of iron-dextran. Dtsch. Tierarztl. Wochenschr. 1966, 73, 145–50. [Google Scholar] [PubMed]
- Moretti, D.; Goede, J.S.; Zeder, C.; Jiskra, M.; Chatzinakou, V.; Tjalsma, H.; Melse-Boonstra, A.; Brittenham, G.; Swinkels, D.W.; Zimmermann, M.B. Oral iron supplements increase hepcidin and decrease iron absorption from daily or twice-daily doses in iron-depleted young women. Blood 2015, 126, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Starzyński, R.R.; Laarakkers, C.M.; Tjalsma, H.; Swinkels, DW.; Pieszka, M.; Styś, A.; Mickiewicz, M.; Lipiński, P. Iron supplementation in suckling piglets: How to correct iron deficiency anemia without affecting plasma hepcidin levels. PLoS ONE 2013, 8, e64022. [Google Scholar] [CrossRef] [PubMed]
- Colombo, C.; Palumbo, G.; He, J.-Z.; Pinton, R.; Cesco, S. Review on iron availability in soil: Interaction of Fe minerals, plants, and microbes. J. Soils Sediments 2014, 14, 538. [Google Scholar] [CrossRef]
- Bilkei, G. The management of veterinary extension activities and the practical experiences of pig breeding advisory bureaus in western Europe. In Proceedings of the Pig Days, Budapest, Hungary, 3–4 April 1996; Bilkei, G., Ed.; Self Publishing: Budapest, Hungary; pp. 1–3. [Google Scholar]
- Kleinbeck, S.N.; McGlone, J.J. Intensive indoor versus outdoor swine production systems: Genotype and supplemental iron effects on blood hemoglobin and selected immune measures in young pigs. J. Anim. Sci. 1999, 77, 2384–2390. [Google Scholar] [CrossRef] [PubMed]
- Rincker, M.J.; Hill, G.M.; Link, J.E.; Rowntree, J.E. Effects of dietary iron supplementation on growth performance, hematological status, and whole-body mineral concentrations of nursery pigs. J. Anim. Sci. 2004, 82, 3189–3197. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, M.; Ficek, R.; Synková, B.; Drábek, J. Efficiency of carbonyl iron in prevention of anaemia in piglets. Acta Vet. Brno 2007, 76, 179–185. [Google Scholar] [CrossRef]
- Svoboda, M.; Ficek, R.; Drábek, J. Evaluation of the efficacy of iron polymaltose complex in the prevention of anaemia in piglets. Bull. Vet. Inst. Pulawy 2008, 52, 119–123. [Google Scholar]
- Antileo, R.; Figueroa, J.; Valenzuela, C. Characterization of a novel encapsulated oral iron supplement to prevent iron deficiency anemia in neonatal piglets. J. Anim. Sci. 2016, 94, 157–160. [Google Scholar] [CrossRef]
- Maes, D.; Steyaert, M.; Vanderhaeghe, C.; López Rodríguez, A.; de Jong, E.; Del Pozo Sacristán, R.; Vangroenweghe, F.; Dewulf, J. Comparison of oral versus parenteral iron supplementation on the health and productivity of piglets. Vet. Rec. 2011, 168, 188. [Google Scholar] [CrossRef] [PubMed]
- Zödl, B.; Sargazi, M.; Zeiner, M.; Roberts, N.B.; Steffan, I.; Marktl, W.; Ekmekcioglu, C. Toxicological effects of iron on intestinal cells. Cell Biochem. Funct. 2004, 22, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Loh, T.; Leong, K.; Too, H.; Mah, C.; Choo, P. The effects of iron supplementation in preweaning piglets. Malays. J. Nutr. 2001, 7, 41–49. [Google Scholar]
- Marchant, J.N.; Broom, D.M.; Corning, S. The influence of sow behaviour on piglet mortality due to crushing in an open farrowing system. Anim. Sci. 2001, 72, 19–28. [Google Scholar] [CrossRef]
- Valenzuela, C.; Lagos, G.; Figueroa, J.; Tadich, T. Behavior of suckling pigs supplemented with an encapsulated iron oral formula. J. Vet. Behav. 2016, 13, 6–9. [Google Scholar] [CrossRef]
- Lin, Y.W.; Wang, J. Structure and function of heme proteins in non-native states: A mini-review. J. Inorg. Biochem. 2013, 129, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Anderson, G.J.; Frazer, D.M.; McKie, A.T.; Vulpe, C.D.; Smith, A. Mechanisms of haem and non-haem iron absorption: Lessons from inherited disorders of iron metabolism. Biometals 2005, 18, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Fillebeen, C.; Gkouvatsos, K.; Fragoso, G.; Calvé, A.; Garcia-Santos, D.; Buffle, R.M.; Becker, C.; Schümann, K.; Ponka, P.; Santos, M.M.; Pantopoulos, K. Mice are poor heme absorbers and do not require intestinal Hmox1 for dietary heme iron assimilation. Haematologica 2015, 100, e334–e337. [Google Scholar] [CrossRef] [PubMed]
- González-Rosendo, G.; Polo, J.; Rodríguez-Jerez, J.J.; Puga-Díaz, R.; Reyes-Navarrete, E.G.; Quintero-Gutiérrez, A.G. Bioavailability of a heme-iron concentrate product added to chocolate biscuit filling in adolescent girls living in a rural area of Mexico. J. Food Sci. 2010, 75, H73–H78. [Google Scholar] [CrossRef] [PubMed]
- Young, M.F.; Griffin, I.; Pressman, E.; McIntyre, A.W.; Cooper, E.; McNanley, T.; Harris, Z.L.; Westerman, M.; O’Brien, K.O. Utilization of iron from an animal-based iron source is greater than that of ferrous sulfate in pregnant and nonpregnant women. J. Nutr. 2010, 140, 2162–2166. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Gutiérrez, A.G.; González-Rosendo, G.; Sánchez-Muñoz, J.; Polo-Pozo, J.; Rodríguez-Jerez, J.J. Bioavailability of heme iron in biscuit filling using piglets as an animal model for humans. Int. J. Biol. Sci. 2008, 4, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M. The potential of encapsulated iron compounds in food fortification: A review. Int. J. Vitam. Nutr. Res. 2004, 74, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Bryszewska, M.A.; Laghi, L.; Zannoni, A.; Gianotti, A.; Barone, F.; Taneyo Saa, D.L.; Bacci, M.L.; Ventrella, D.; Forni, M. Bioavailability of microencapsulated iron from fortified bread assessed using piglet model. Nutrients. 2017, 9, 272. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Liposomal drug delivery systems: From concept to clinical applications. Adv. Drug Deliv. Rev. 2013, 65, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Riccio, E.; Sabbatini, M.; Andreucci, M.; Del Rio, A.; Visciano, B. Effect of oral liposomal iron versus intravenous iron for treatment of iron deficiency anaemia in CKD patients: A randomized trial. Nephrol. Dial. Transplant. 2015, 30, 645–652. [Google Scholar] [CrossRef] [PubMed]
- Giordano, G.; Mondello, P.; Tambaro, R.; Perrotta, N.; D’Amico, F.; D’Aveta, A.; Berardi, G.; Carabellese, B.; Patriarca, A.; Corbi, G.M.; et al. Biosimilar epoetin α is as effective as originator epoetin-α plus liposomal iron (Sideral®), vitamin B12 and folates in patients with refractory anemia: A retrospective real-life approach. Mol. Clin. Oncol. 2015, 3, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Berti, C.; Mandò, C.; Martinelli, A.; Mazzali, C.; Cetin, I. Effects of different regimens of iron prophylaxis on maternal iron status and pregnancy outcome: A randomized control trial. J. Matern. Fetal Neonatal Med. 2017, 30, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- Muckenthaler, M.U.; Rivella, S.; Hentze, M.W.; Galy, B. A red carpet for iron metabolism. Cell 2017, 168, 344–361. [Google Scholar] [CrossRef] [PubMed]
- Lipiński, P.; Styś, A.; Starzyński, R.R. Molecular insights into the regulation of iron metabolism during the prenatal and early postnatal periods. Cell. Mol. Life Sci. 2013, 70, 23–38. [Google Scholar] [CrossRef] [PubMed]
- Collard, K.J. Iron homeostasis in the neonate. Pediatrics 2009, 123, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.R.; Ullrey, D.E. The pig as a model for human nutrition. Annu. Rev. Nutr. 1987, 7, 361–382. [Google Scholar] [CrossRef] [PubMed]
- Schook, L.; Beattie, C.; Beever, J.; Donovan, S.; Jamison, R.; Zuckermann, F.; Niemi, S.; Rothschild, M.; Rutherford, M.; Smith, D. Swine in biomedical research: Creating the building blocks of animal models. Anim. Biotechnol. 2005, 16, 183–190. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szudzik, M.; Starzyński, R.R.; Jończy, A.; Mazgaj, R.; Lenartowicz, M.; Lipiński, P. Iron Supplementation in Suckling Piglets: An Ostensibly Easy Therapy of Neonatal Iron Deficiency Anemia. Pharmaceuticals 2018, 11, 128. https://doi.org/10.3390/ph11040128
Szudzik M, Starzyński RR, Jończy A, Mazgaj R, Lenartowicz M, Lipiński P. Iron Supplementation in Suckling Piglets: An Ostensibly Easy Therapy of Neonatal Iron Deficiency Anemia. Pharmaceuticals. 2018; 11(4):128. https://doi.org/10.3390/ph11040128
Chicago/Turabian StyleSzudzik, Mateusz, Rafał R. Starzyński, Aneta Jończy, Rafał Mazgaj, Małgorzata Lenartowicz, and Paweł Lipiński. 2018. "Iron Supplementation in Suckling Piglets: An Ostensibly Easy Therapy of Neonatal Iron Deficiency Anemia" Pharmaceuticals 11, no. 4: 128. https://doi.org/10.3390/ph11040128
APA StyleSzudzik, M., Starzyński, R. R., Jończy, A., Mazgaj, R., Lenartowicz, M., & Lipiński, P. (2018). Iron Supplementation in Suckling Piglets: An Ostensibly Easy Therapy of Neonatal Iron Deficiency Anemia. Pharmaceuticals, 11(4), 128. https://doi.org/10.3390/ph11040128