Abstract
Western Tianshan National Nature Reserve in Xinjiang, China stands out for its uniqueness and high biodiversity, including lichens. This study aims to characterize lichen diversity and compare distribution patterns of different life forms, substratum affinities and photobiont types. Surveys were conducted from June to August 2024 using stratified sampling methods at elevation ranging from 1100 m to 3400 m in the study area. Morphological, anatomical and chemical studies revealed 173 lichen species from 24 families and 58 genera, of which 100 species were identified as crustose, 46 as foliose and 27 as fruticose. Among the different habitat groups, strictly saxicolous lichens were dominant with 89 species, followed by corticolous lichens with 44 species and terricolous lichens with 40 species. The total species richness of lichens has a bimodal pattern: one peak appears at a low altitude (1701–2000 m) and the other at a high altitude (2901–3200 m). Among the three substratum categories studied, the species richness of terricolous lichens showed a unimodal relationship with elevation, and the saxicolous lichen had a bimodal pattern. The species richness of corticolous lichens was highest at lower and medium elevations and decreased at higher elevations. With respect to photobiont type, the species richness of cyanolichens showed a unimodal relationship with elevation. Maximum richness occurred at 2700 m, contrary to the chlorolichens, which had a bimodal pattern. Species richness of all three growth forms of lichens showed a bimodal pattern related to elevation. Among the three morphological types, crustose and foliose species richness had their highest values of 38 and 19, respectively, at 1701–2000 m, and fruticose lichens peaked with a maximum of 13 species at 2301–2600 m. The species richness of crustose lichens is lowest between altitudes 2300 and 2600 m, while the lowest species richness of fruticose and foliose lichens occurs at 2001–2300 m and elevations above 2900 m.
1. Introduction
Protecting biodiversity has always been a hot topic in the world. At present, more than one million species of animals and plants in the world are on the verge of extinction [1,2]. Climate change and biodiversity loss are becoming two major challenges facing the world today, which are closely related to human development [1]. China is a country with an extremely rich biodiversity which ranks eighth in the world. Protecting biodiversity has become a national strategy to safeguard China’s ecological security [3]. In recent years, with the COP15 meeting of the Conference of the Parties to the United Nations Convention on Biological Diversity, the ICG-16 Forum on Biodiversity and Global Emergencies was held in China, and the publication of the White Paper on Biodiversity Conservation in China and the concept and practice of biodiversity conservation in China have been introduced to the world, which has enhanced the international community’s understanding of biodiversity conservation in China [3,4].
A lichen is fundamentally composed of a fungal species which exists symbiotically with photosynthetic organisms such as blue-green algae and/or cyanobacteria [5]. Lichens can grow almost anywhere, although the type of species may differ based on the environmental conditions [6]. An estimated 13,500 to 30,000 species of lichens are distributed worldwide in a wide range of different terrestrial habitats [7,8], and c. 8% of the terrestrial surface of the earth is covered by lichen-dominated vegetation [9]. China is undoubtedly a lichen-rich country, but currently only 3014 species have been documented [10,11]. An assessment of the threatened categories of lichens in China by Wei et al. (2020) showed that more than half the species, 1473 (68.07%), were Data Deficient (DD) and could not be further evaluated due to lack of data [12]. This highlights the severe lack of lichen research and the urgent need for lichen taxonomists in China [12].
Xinjiang (Xinjiang Uygur Autonomous Region of China) is the largest of China’s five autonomous regions and 23 provinces. It occupies an area of 1.6 million km2 (0.64 million square miles). The earliest list of 67 species, 19 varieties and 17 forms of lichens for Xinjiang was published by Wei (1991) [13] and was extended by Tumur et al. (2021) increased the list to 596 taxa (580 species, 4 subspecies and 12 varieties) in 160 genera [14].
Tian Shan Mountains, also called Tien Shan, Celestial Mountains or Heavenly Mountains, is a large system of mountain ranges situated in the Eurasia hinterland. The Tian Shan Mountains’ location makes it the furthest mountain system from the sea with widespread drought regions. As one of the seven mountain systems in the world, Tian Shan Mountains stretch through four countries (China, Kazakhstan, Kyrgyzstan and Uzbekistan) with a length of 2500 km (about 1553 miles) from east to west [15]. It has become the most typical representative of the large-scale mountain ecosystem in the temperate arid region of the world because of its geographical location, temperate continental arid climate, landform pattern of alternating mountains and basins, numerous glacial rivers, wonderful natural scenery, special biota and ecological processes [15]. The Western Tianshan National Nature Reserve (WTNNR) is situated on the northern slope of the Nalati Mountains, a branch of the western Tianshan Mountains and located in the eastern part of Gongliu County, Ili Kazak Autonomous Prefecture, Xinjiang Uygur Autonomous Region, China. The protected area is 28 km long from north to south and 14 km wide from east to west, with a total area of 31,217 hm2. The lowest altitude is 1100 m, and the highest peak, Kabanbayi Peak, is 4257 m above sea level. The average annual temperature is 5–7 °C, with extreme maximum and minimum temperatures of 37.4 °C and −36.7 °C, respectively, with a frost-free period of 120 days and an annual precipitation of 800–1000 mm, reaching the subtropical precipitation level. The Reserve is rich in water resources, with a mild and humid climate and good natural conditions. It is the region with the highest precipitation in Xinjiang and is known as the “wet island” in arid deserts. It belongs to the priority protection forest ecosystem in Xinjiang due to the high biodiversity. It is a “natural gene pool” of wild animal and plant species, which is highly typical and representative, and has great value in ecological protection [15]. In April 1983, the People’s Government of Xinjiang Uygur Autonomous Region, China approved the establishment of the “Gongliu Spruce Nature Reserve” in Gongliu County, Ili Kazak Autonomous Prefecture, and in April 2000 it was renamed “Western Tianshan National Nature Reserve”. This Reserve preserves the largest primitive coniferous forest in the west of China, forming four altitudinal temperate-to-cold landscape zones [15]. The basic vegetation types of the protected area are cold temperate coniferous forest, temperate deciduous forest, mixed coniferous and deciduous forest, and mountain meadow. There are 847 species of vascular plants, belonging to 390 genera and 93 families [15].
However, in recent years, due to overgrazing, grassland degradation has become severe, and many herdsmen have chosen grazing sites within protected areas, leading to a reduction in biodiversity within these areas. In addition, the nature reserve attracts large numbers of tourists due to its unique scenery, but many tourism facilities within the nature reserve are still incomplete due to a lack of funds. Overtourism development has caused environmental pollution within the protected area, resulting in serious impacts on the ecological system of the protected area [15].
Lichen changes at the community or population level are used as sensitive indicators of the biological effect of pollutants [16]. The earliest lichen records from the western part of the Tianshan Mountains were listed by Elenkin [17,18] and Vainio [19], who studied the lichens collected from deserts and steppes. Yang et al. [20], who reported 78 lichen species belonging to 31 genera and 15 families, analyzed the characteristics of the flora and compared them with neighboring regions [20]. Subsequent research mainly focused on the distribution and community structure of saprophytic and corticolous lichen species [21,22].
Although lichens are an important component of the forest ecosystem in the Western Tianshan National Nature Reserve, research on lichen species diversity and species distribution characteristics is limited. The aims of the present study are to characterize lichen diversity and compare distribution patterns of different life forms, substratum affinities and photobiont types in the western Tianshan Mountain National Nature Reserve, and to provide a scientific basis for effective protection of lichen resources.
2. Materials and Methods
2.1. Study Area
The study sites in the Western Tianshan National Nature Reserve (WTNNR) are located within latitudes 43°03′–43°15′ N and longitudes 82°51′–83°06′ E. The conservation area covers an area of 31,217 hm2 and encompasses a wide range of elevations, from 1100 to 4257 m [15,23]. The diverse topography and climate make WTNNR an ideal location for studying lichen diversity and patterns of species distribution (Figure 1).
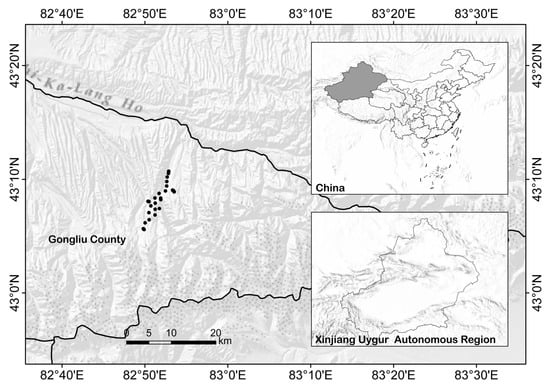
Figure 1.
Location of Western Tianshan National Nature Reserve.
2.2. Lichen Collection and Identification
We conducted the lichen survey of WTNNR between June and August in 2024 using the stratified sampling method at elevation of 1100 m to 3400 m. We visited woody debris, rocks, soil, and stumps, trunks and branches of trees, and where we observed any lichen species of interest we made a collection, recording the collection site with a Garmin GPS (Figure 2).

Figure 2.
Sampling zones: (A,B) Mountain meadow, (C,D) Cold temperate coniferous forest, (E) Temperate deciduous forest, (F) Mixed coniferous and deciduous forest.
In total, 472 lichen specimens were gathered within an elevation range of 1100 to 3400 m. The collected lichen specimens were identified at the Laboratory of the Arid Land Lichens Research Center of the College of Life Sciences and Technology, Xinjiang University, China, with the help of the following keys and checklists [7,24], and the nomenclature generally follows Index Fungorum. The identification process involved studying the morphology, anatomy and chemistry of the specimens. Lichen samples were morpho-anatomically examined and identified up to species level using a stereomicroscope (Nikon Eclipse E200, Otawara, Japan) and a light microscope (Zeiss Axioskop 2 plus, Oberkochen, Baden-Württemberg, Germany), and chemically with the help of spot tests, UV light and standardized thin-layer chromatography [7,25]. In addition, data about growth forms, photobiont types, substrate and elevation of each collected sample were recorded. Voucher specimens are deposited in the Lichen Herbarium of College of Life Science and Technology, Xinjiang University (XJU), China.
2.3. Data Analysis
The elevation range of lichens in WTNNR, from 1100 to 3400 m, was divided into 23 elevational bands each of 100 m, and a complete set of a presence/absence of lichen species data matrix through elevation was prepared [26]. This is a macro-scale study where the definition of species richness is the total number of lichen species occurring in each 100 m elevation band covering the entire elevational range of WTNNR [27].
3. Results
3.1. Lichen Species Composition
The study revealed the occurrence of 173 lichen species belonging to 58 genera and 24 families in the area (Appendix A). Among these lichens, 100 species (57.8% of total species) were crustose (including squamulose and placodioid), 46 species (26.5% of total species) were foliose, and 27 species (15.6% of total species) were fruticose (including dimorphic species); of these, 8.1% had a cyanobacterial photobiont and 91.8% had a green algae photobiont. Among the different habitat groups, strictly saxicolous lichens were dominant with 89 species, followed by corticolous lichens with 44 species, and terricolous lichens with 40 species (Appendix A).
The species richness at family level shows that there are four dominant families containing 87 species (50% of total species). The polytypic families, such as Lecideaceae, Teloschistaceae, Peltigeracea, Acarosporaceae and Megasporaceae, have 51 species (29.5% of total species). There are 10 oligotypic families including 30 species (7.4% of species), and five monotypic families (2.8% of total species) (Table 1). At the generic level, four are dominant, namely Cladonia Nyl. (15 species), Lecanora Ach. (11 species), Lecidea Nyl. (11 species) and Peltigera Willd. (11 species). Lecidella Körb. (8 species) and Xanthoparmelia (Vain.) Hale (6 species) belong to polytypic genera. There are 28 oligotypic and 24 monotypic genera, which are widely distributed over the different elevations (Table 2).

Table 1.
Statistical results of genera and species in the lichen family in the research area.
Table 2.
Statistics of species richness of genera.
3.2. The Relationship Between Species Richness and Elevation of Lichens
The total of species-rich lichens has a bimodal pattern: one peak with the highest species richness appears at low elevation (1701–2000 m) and the other at high elevation (2901–32,000 m) (Figure 3).
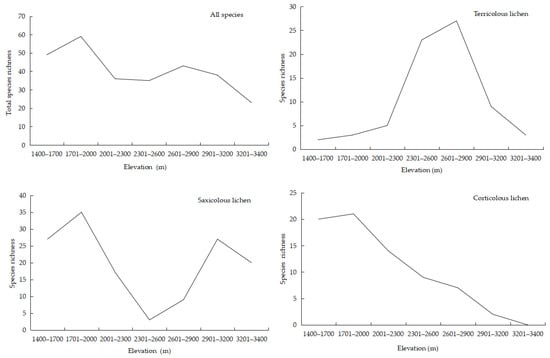
Figure 3.
Relationship between elevation and lichen species richness.
The observed maximum richness (59 species), however, was at c. 1850 m. Among the three substratum categories studied, the species richness of terricolous lichens showed a unimodal relationship with elevation; the maximum richness occurred at 2601–2900 m, rising to 27 species at c. 2780 m. The saxicolous lichens have a bimodal pattern for species richness, with significant increases in number between 1701 and 2000 m, and between 2901 and 3000 m. For corticolous lichens, maximum species richness occurred at 1701–2000 m. Then, the species richness decreases with increasing elevation and becomes zero at 3201–3400 m.
Regarding photobionts, the species richness of cyanolichens showed a unimodal relationship with elevation; maximum richness occurred at 2700 m and the lowest levels between 1400 and 2000 m. However, species richness of chlorolichens had a bimodal pattern, reaching a maximum at 1701–2000 m, and maintained a relatively high level at all of the other elevations (Figure 4).
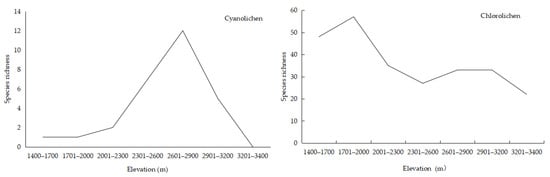
Figure 4.
Relationship between elevation and Cyanolichen and Chlorolichen species richness.
Species richness of all three growth forms of lichens showed a bimodal pattern with elevation. Among the three morphological types, crustose and foliose species richness had the highest values of 38 and 19, respectively, at 1701–2000 m, and fruticose species richness peaked at a maximum of 13 species at 2301–2600 m (Figure 5).
Figure 5.
Relationship between elevation and lichen growth form.
3.3. The Relationship Between Dominant Families and Elevation
Among the dominant families found on WTNNR, Lecanoraceae, Parmeliaceae, Cladoniaceae and Physciaceae showed a significantly curvilinear relationship (Figure 6). Lecanoraceae peaked with a maximum species richness of 17 species at 1701–2000 m and a minimum species richness of 2 species at 2901–3200 m. A maximum richness of 14 species for Parmeliaceae occurs at 1401–1700 m, 11 species for Parmeliaceae at 2301–2650 m, and 9 species for Physciaceae at 1701–2000 m.
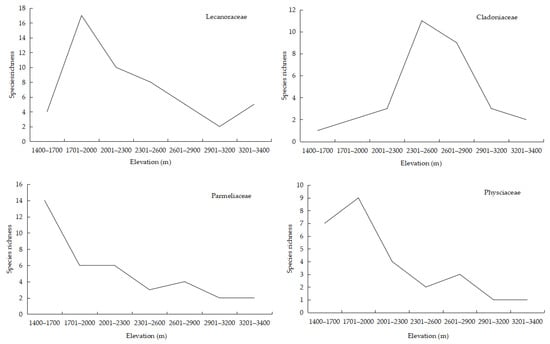
Figure 6.
Elevational richness pattern shown by dominant lichen families.
4. Discussion
Lichens, which grow on trees, rocks, and various substrates, exhibit resilience to water scarcity and do not require complex living conditions [28]. Their widespread distribution in arid and semi-arid areas in mountainous regions in Xinjiang highlights their ability to inhabit diverse ecological niches, and since forests also provide the most suitable habitats for lichens, the WTNNR supports an exceptionally high biodiversity of lichens [24]. Our study determined the presence of 173 lichen species, spanning 24 families and 58 genera, with considerable differences in species composition and abundance between the various elevations. This study showed that crustose lichens (57.8% of species) were predominant, followed by foliose (26.5%) and fruticose (15.6%). Studies from other protected areas in Xinjiang have also shown crustose species to be more numerous than other growth forms [24]. Crustose lichens demonstrate a greater resilience to rising temperatures than foliose and fruticose–filamentose species. This resilience is attributed to their lower surface-to-volume ratio, their less exposed surface area resulting in a higher tolerance to drying out [29]. In well-lit and open forests at high elevations and in the upper canopies of most trees, fruticose lichens can utilize light from all directions, whereas many foliose and crustose lichens utilize more-or-less unidirectional light in shady positions within dense forests and on low canopy branches [30]. In our research, the highest species richness of fruticose lichens occurred in relatively open canopy forest, and foliose lichens were distributed in shady habitats in dense forest and on lower canopy branches.
Ecological factors play an important role in the growth, development, distribution and diversity of lichen species [31,32]. Variations in microclimatic conditions, particularly light, water and nutrients, affected by local sources of disturbance, such as roads or farms, different land uses or habitat fragmentation, can influence lichen diversity [33,34]. In WTNNR, the distribution of lichen species lacks uniformity and reflects variability in environmental conditions at different elevations. Saxicolous species showed a bimodal pattern at lower and higher elevations because of the higher ability to adapt to extreme harsh environmental conditions and artificial disturbances, such as tourism and the interference of grazing at lower altitudes, and at higher elevations many exposed rocks and boulders provide ideal habitat for saxicolous lichens. The high atmospheric moisture and cooler summer temperatures at mid-elevations favor terricolous lichens in India [35,36]. Phytosociological factors, such as a decrease in competition from vascular plants, also contribute to the mid-elevational peak species richness of terricolous lichens [26,37]. The decrease in terricolous species richness beyond mid-elevations can be attributed to a decrease in overall soil cover, as the landscape at higher elevations is dominated by exposed rocks, and soil crusts are limited to rock crevices and some flat faces of rocks [35,36]. In WTNNR, species richness of terricolous lichens showed a unimodal relationship with elevation, and the maximum richness occurred at 2601–2900 m, where shrubs such as Sabina vulgaris and S. pseudosabina are dominant, and coverage of vascular plants is less than 40% [15]. Because of the large number of temperate broad-leaved and coniferous species with bark differing in roughness, moisture retention capacity and pH present a wide variety of habitats for lichens [26]. In WTNNR, between 1700 and 2200 m, the species richness of corticolous lichens is highest, with 24 species. However, lichen diversity can be greatly limited by lack of long ecological continuity in forests and may be less prevalent at low altitudes because of dense forest canopies and intensive land use [38]. Our study showed that the strong reduction in lichen richness at low elevations is a result of various types of human interference.
In conclusion, the lichen flora of the WTNNR is remarkably rich and diverse, with a unimodal and bimodal pattern of species diversity related to elevation. There are distinct species assemblages at different elevational sites, showing restrictive species distribution and signifying a need for the protection of large areas for lichen conservation.
Author Contributions
A.T.; investigation, methodology, writing—original draft preparation, R.M.; project administration, formal analysis, M.R.D.S.; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants from the Central Forestry and Grassland Ecological Protection and Restoration Project in Xinjiang Western Tianshan National Nature Reserve in 2024 (grant number 202406140025).
Institutional Review Board Statement
This study did not require ethical approval.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We are thanks to Administration office of Western Tianshan National Nature Reserve for allowing us to field investigation and grateful to Wuernisha Shayiding (Xinjiang University, China) for help the species identification, Zhu Yong Qi (Xinjiang University, China), Guldiyar Adil and Jinsiguli Bahanur (graduate students of Xinjiang University, China) for their help with the field work. David H.S Richardson (Saint Mary’s University, Canada) for editorial help and valuable comments.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| WTNNR | Western Tianshan National Nature Reserve |
Appendix A
Table A1.
Elevational ranges of lichens of Western Tianshan National Nature Reserve, their families and growth forms.
Table A1.
Elevational ranges of lichens of Western Tianshan National Nature Reserve, their families and growth forms.
| S.No. | Name of Lichen Species | Family | Algal Photobiont | Growth Form | Substrate Types | Altitudinal Range (m) | Number of Specimens Examined | |
|---|---|---|---|---|---|---|---|---|
| 1 | Acarospora aeginaica H. Magn. | Acarosporaceae | Gr | Cr | Saxicolous | 1453 | 3253 | 2 |
| 2 | A. atrata Hue. | Acarosporaceae | Gr | Cr | Saxicolous | 2021 | 3121 | 2 |
| 3 | A.bohlinii H. Magn. | Acarosporaceae | Gr | Cr | Saxicolous | 2510 | 3265 | 4 |
| 4 | A.invadens H.Magn. | Acarosporaceae | Gr | Cr | Saxicolous | 2048 | - | 1 |
| 5 | A.nitrophila H. Magn. | Acarosporaceae | Gr | Cr | Saxicolous | 2035 | - | 1 |
| 6 | Anaptychia ciliaris (L.) Flot. | Physciaceae | Gr | Fr | Corticolous | 1457 | - | 1 |
| 7 | A. setifera (Mereschk.) Räsänen | Physciaceae | Gr | Fr | Corticolous | 1684 | - | 1 |
| 8 | Aspicilia calcitrapa Cl. Roux & A. Nordin | Megasporaceae | Gr | Cr | Saxicolous | 1807 | - | 1 |
| 9 | A. phaea Owe-Larss. & A. Nordin | Megasporaceae | Gr | Cr | Saxicolous | 1732 | - | 1 |
| 10 | Athallia pyracea (Ach.) Arup, Frödén & Søchting | Teloschistaceae | Gr | Cr | Saxicolous | 1798 | - | 1 |
| 11 | Biatora vernalis (L.) Fr. | Ramalinaceae | Gr | Cr | Terricolous | 1774 | - | 1 |
| 12 | Calogaya biatorina (A. Massal.) Arup, Frödén & Søchting | Teloschistaceae | Gr | Cr | Saxicolous | 1684 | 3000 | 2 |
| 13 | Caloplaca atroflava (Turner) Mong. | Teloschistaceae | Gr | Cr | Saxicolous | 1732 | - | 1 |
| 14 | C. cerina (Hedw.) Th. Fr. | Teloschistaceae | Gr | Cr | Corticolous | 1460 | - | 1 |
| 15 | Calicium abietinum Pers. | Caliciaceae | Gr | Cr | Corticolous | 1800 | 2048 | 5 |
| 16 | Candelariella aurella (Hoffm.) Zahlbr. | Candelariaceae | Gr | Cr | Corticolous | 1756 | 2540 | 7 |
| 17 | C. efflorescens R.C. Harris & W.R. Buck | Candelariaceae | Gr | Cr | Corticolous | 1561 | 2641 | 2 |
| 18 | Canomaculina subtinctoria Zahlbr. (Elix) | Parmeliaceae | Gr | Fl | Corticolous | 1594 | - | 1 |
| 19 | Circinarea maculata (H. Magn.) Q. Ren | Megasporaceae | Gr | Cr | Saxicolous | 1457 | 2800 | 7 |
| 20 | C. tortuosa (H. Magn.) Q. Ren | Megasporaceae | Gr | Cr | Saxicolous | 1807 | 2700 | 5 |
| 21 | Cladonia acuminata (Ach.) Norrl. | Cladoniaceae | Gr | Dm | Terricolous | 1685 | - | 1 |
| 22 | C. cariosa (Ach.) Spreng | Cladoniaceae | Gr | Dm | Terricolous | 2021 | 3150 | 2 |
| 23 | C. chlorophaea (Flörke ex Sommerf.) Spreng. | Cladoniaceae | Gr | Dm | Terricolous | 1405 | 2600 | 9 |
| 24 | C.coniocraea (Flörke) Spreng. | Cladoniaceae | Gr | Dm | Terricolous | 2453 | 2600 | 4 |
| 25 | C. fimbriata (L.) Fr. | Cladoniaceae | Gr | Dm | Terricolous | 2460 | 2600 | 7 |
| 26 | C.floerkeana (Fr.) Flörke | Cladoniaceae | Gr | Dm | Terricolous | 2584 | 2600 | 2 |
| 27 | C. furcata (Huds.) Baumg. | Cladoniaceae | Gr | Dm | Terricolous | 2820 | 2900 | 2 |
| 28 | C. macilenta Hoffm. | Cladoniaceae | Gr | Dm | Terricolous | 2866 | 2900 | 7 |
| 29 | C. multiformis G. Merr. | Cladoniaceae | Gr | Dm | Terricolous | 2798 | 3350 | 2 |
| 30 | C. pocillum (Ach.) Grognot | Cladoniaceae | Gr | Dm | Terricolous | 2548 | 3200 | 2 |
| 31 | C. polycarpoides Nyl. | Cladoniaceae | Gr | Dm | Terricolous | 2798 | 2900 | 3 |
| 32 | C. pyxidata (L.) Hoffm. | Cladoniaceae | Gr | Dm | Terricolous | 2594 | 3400 | 8 |
| 33 | C. scabriuscula (Delise) Leight. | Cladoniaceae | Gr | Dm | Terricolous | 2416 | 2500 | 2 |
| 34 | C. squamosa Hoffm. | Cladoniaceae | Gr | Dm | Terricolous | 2807 | 2910 | 6 |
| 35 | C. subulata (L.) F.H. Wigg. | Cladoniaceae | Gr | Dm | Terricolous | 1756 | - | 1 |
| 36 | Collema flaccidum (Ach.) Ach. | Collemataceae | Bg | Fl | Terricolous | 2439 | 2700 | 2 |
| 37 | Dermatocarpon miniatum (L.) W. Mann | Verrucariaceae | Gr | Fl | Saxicolous | 1807 | 3400 | 4 |
| 38 | Dimelaena oreina (Ach.) Norman | Caliciaceae | Gr | Cr | Saxicolous | 1798 | 3000 | 5 |
| 39 | Diploschistes diacapsis (Ach.) Lumbsch. | Graphidaceae | Gr | Cr | Terricolous | 1869 | - | 1 |
| 40 | D. muscorum (Scop.) R. Sant. | Graphidaceae | Gr | Cr | Corticolous | 1561 | 2650 | 3 |
| 41 | D. scruposus (Schreb.) Norman. | Graphidaceae | Gr | Cr | Saxicolous | 1756 | 3200 | 2 |
| 42 | D. tianshaniensis A. Abbas, S.Y. Guo & Ababaikeli | Graphidaceae | Gr | Cr | Saxicolous | 2035 | - | 1 |
| 43 | D. viridis R. Mamut, J.J. Fang & T. Payzulla | Graphidaceae | Gr | Cr | Saxicolous | 2274 | - | 1 |
| 44 | Evernia divaricata (L.) Ach. | Parmeliaceae | Gr | Fr | Corticolous | 2260 | 2307 | 3 |
| 45 | Flavoparmelia baltimorensis (Gyeln. & Fóriss) Hale. | Parmeliaceae | Gr | Fr | Corticolous | 1570 | - | 1 |
| 46 | F. caperata (L.) Hale | Parmeliaceae | Gr | Fr | Corticolous | 1500 | 1690 | 8 |
| 47 | Flavopunctelia sorediata (Nyl.) Hale | Parmeliaceae | Gr | Fl | Corticolous | 1684 | - | 1 |
| 48 | Gyalolechia epiphyta (Lynge) Vondrák | Teloschistaceae | Gr | Cr | Saxicolous | 1467 | - | 1 |
| 49 | G. marmorata (Bagl.) Nimis & Arup | Teloschistaceae | Gr | Cr | Saxicolous | 1582 | - | 1 |
| 50 | Gypsoplaca albimarginata R. Mamut | Gypsoplacaceae | Gr | Sq | Terricolous | 1460 | - | 1 |
| 51 | Lecanora argopholis (Ach.) Ach. | Lecanoraceae | Gr | Cr | Saxicolous | 1869 | 3350 | 5 |
| 52 | L. coniferarum Printzen | Lecanoraceae | Gr | Cr | Saxicolous | 2016 | - | 1 |
| 53 | L. horiza (Ach.) Nyl. | Lecanoraceae | Gr | Cr | Saxicolous | 2031 | - | 1 |
| 54 | L. intricata (Ach.) Ach. | Lecanoraceae | Gr | Cr | Saxicolous | 2031 | - | 1 |
| 55 | L. meridionalis H. Magn | Lecanoraceae | Gr | Cr | Corticolous | 1869 | 2300 | 2 |
| 56 | L. quercicola Coppins & P. James | Lecanoraceae | Gr | Cr | Saxicolous | 1900 | 3016 | 2 |
| 57 | L. rupicola (L.) Zahlbr. | Lecanoraceae | Gr | Cr | Saxicolous | 1869 | - | 1 |
| 58 | L. saligna (Schrad.) Zahlbr. | Lecanoraceae | Gr | Cr | Saxicolous | 1869 | - | 1 |
| 59 | L. subravida Nyl. | Lecanoraceae | Gr | Cr | Saxicolous | 2031 | 3350 | 4 |
| 60 | L. symmicta (Ach.) Ach. | Lecanoraceae | Gr | Cr | Corticolous | 2031 | - | 1 |
| 61 | L. xylophila Hue | Lecanoraceae | Gr | Cr | Corticolous | 2016 | - | 1 |
| 62 | Lecidea atrobrunnea (DC.) Schaer | Lecideaceae | Gr | Cr | Saxicolous | 2021 | 3000 | 2 |
| 63 | L. auriculata Th. Fr | Lecideaceae | Gr | Cr | Saxicolous | 1820 | 3150 | 4 |
| 64 | L. confluens (Weber) Ach. | Lecideaceae | Gr | Cr | Saxicolous | 2150 | - | 1 |
| 65 | L. elabens Fr. | Lecideaceae | Gr | Cr | Saxicolous | 2230 | 3400 | 3 |
| 66 | L. fuscoatra (L.) Ach | Lecideaceae | Gr | Cr | Saxicolous | 1808 | - | 1 |
| 67 | L. laboriosa Müll. Arg | Lecideaceae | Gr | Cr | Saxicolous | 1798 | 3100 | 2 |
| 68 | L. lapicida (Ach.) Ach. | Lecideaceae | Gr | Cr | Saxicolous | 2021 | 3300 | 2 |
| 69 | L. paratropoides Müll. Arg | Lecideaceae | Gr | Cr | Saxicolous | 1798 | 3300 | 2 |
| 70 | L. perlatolica Hertel & Leuckert | Lecideaceae | Gr | Cr | Saxicolous | 2230 | - | 1 |
| 71 | L. promiscens Nyl. | Lecideaceae | Gr | Cr | Saxicolous | 1820 | - | 1 |
| 72 | L. tessellata Flörke | Lecideaceae | Gr | Cr | Saxicolous | 2150 | 3400 | 8 |
| 73 | Lecidella carpathica Koerb | Lecanoraceae | Gr | Cr | Saxicolous | 1798 | - | 1 |
| 74 | L. effugiens (Nilson) Knoph & Hertel | Lecanoraceae | Gr | Cr | Saxicolous | 1500 | 2798 | 6 |
| 75 | L. elaeochroma (Ach.) M. Choisy | Lecanoraceae | Gr | Cr | Saxicolous | 1808 | - | 1 |
| 76 | L. enteroleucella (Nyl.) Hertel | Lecanoraceae | Gr | Cr | Saxicolous | 2021 | - | 1 |
| 77 | L. euphorea (Flörke) Hertel | Lecanoraceae | Gr | Cr | Saxicolous | 1820 | - | 1 |
| 78 | L. patavina (A. Massal.) Knoph & Leuckert | Lecanoraceae | Gr | Cr | Saxicolous | 2230 | - | 1 |
| 79 | L. stigmatea (Ach.) Hertel & Leuckert | Lecanoraceae | Gr | Cr | Saxicolous | 1808 | - | 1 |
| 80 | L. tumidula (A.Massal.) Knoph & Leuckert | Lecanoraceae | Gr | Cr | Saxicolous | 1850 | 3150 | 2 |
| 81 | Lecaimmeria botryoides C.M. Xie & Xin Y. | Lecideaceae | Gr | Cr | Saxicolous | 3001 | - | 1 |
| 82 | L. mongolica C.M. Xie & Lu L. Zhang | Lecideaceae | Gr | Cr | Saxicolous | 3001 | - | 1 |
| 83 | L. orbicularis C.M. Xie & Lu L. | Lecideaceae | Gr | Cr | Saxicolous | 3001 | - | 1 |
| 84 | Leptogium saturninum (Dicks.) Nyl. | Collemataceae | Bg | Fl | Corticolous | 1798 | 2950 | 8 |
| 85 | Lepraria incana (L.) Ach. | Stereocaulaceae | Gr | Cr | Terricolous | 2048 | 2848 | 2 |
| 86 | L. lobificans Nyl. | Stereocaulaceae | Gr | Cr | Terricolous | 2500 | 2807 | 2 |
| 87 | Leproplaca cirrochroa (Ach.) Arup, Frödén & Søchting | Teloschistaceae | Gr | Cr | Saxicolous | 1774 | - | 1 |
| 88 | Lichinella nigritella (Lettau) P.P. Moreno & Egea. | Lichinaceae | Bg | Fl | Saxicolous | 1808 | 2400 | 7 |
| 89 | Lobothallia alphoplaca (Wahlenb.) Hafellner | Megasporaceae | Gr | Cr | Saxicolous | 1570 | 3100 | 4 |
| 90 | L. praeradiosa (Nyl.) Hafellner. | Megasporaceae | Gr | Cr | Saxicolous | 1570 | 2850 | 6 |
| 91 | Melanelia exasperatula (Nyl.) Essl. | Parmeliaceae | Gr | Fl | Corticolous | 1774 | 2950 | 4 |
| 92 | M. subargentifera (Nyl.) Essl. | Parmeliaceae | Gr | Fl | Corticolous | 1678 | 2800 | 5 |
| 93 | Melanohalea elegantula (Zahlbr.) O. Blanco et al. | Parmeliaceae | Gr | Fl | Corticolous | 2100 | 2865 | 4 |
| 94 | M. exasperatula (Nyl.) O. Blanco et al. | Parmeliaceae | Gr | Fl | Corticolous | 2100 | 2865 | 5 |
| 95 | M. subelegantula (Essl.) O. Blanco et al. | Parmeliaceae | Gr | Fl | Corticolous | 2100 | 2865 | 3 |
| 96 | Montanelia panniformis (Nyl.) Divakar | Parmeliaceae | Gr | Fl | Saxicolous | 1807 | - | 1 |
| 97 | Myriolecis crenulata (Ach.) Śliwa, Zhao Xin & Lumbsch. | Lecanoraceae | Gr | Cr | Saxicolous | 1678 | - | 1 |
| 98 | M. dispersa (Pers.) Śliwa, Zhao Xin & Lumbsch | Lecanoraceae | Gr | Cr | Saxicolous | 1712 | 3400 | 3 |
| 99 | M. hagenii (Ach.) Ach. | Lecanoraceae | Gr | Cr | Saxicolous | 1678 | 3400 | 4 |
| 100 | M. invadens (H. Magn.) Śliwa, Zhao Xin & Lumbsch. | Lecanoraceae | Gr | Cr | Saxicolous | 1712 | - | 1 |
| 101 | M. semipallida (H. Magn.) Śliwa, Zhao Xin & Lumbsch. | Lecanoraceae | Gr | Cr | Saxicolous | 1678 | - | 1 |
| 102 | Parmelia sulcata Taylor | Parmeliaceae | Gr | Fl | Corticolous | 1684 | 2100 | 9 |
| 103 | Peltigera canina (L.) Willd. | Peltigeraceae | Bg | Fl | Terricolous | 1711 | 2800 | 2 |
| 104 | P. elisabethae Gyeln. | Peltigeraceae | Bg | Fl | Terricolous | 1822 | 2950 | 5 |
| 105 | P. didactyla (With.) J.R. Laundon | Peltigeraceae | Bg | Fl | Terricolous | 1711 | 2970 | 2 |
| 106 | P. horizontalis (Huds.) Baumg. | Peltigeraceae | Bg | Fl | Terricolous | 1679 | - | 1 |
| 107 | P. leucophlebia (Nyl.) Gyeln. | Peltigeraceae | Bg | Fl | Terricolous | 1822 | 2900 | 2 |
| 108 | P. neopolydactyla (Gyeln.) Gyeln. | Peltigeraceae | Bg | Fl | Terricolous | 1579 | 3200 | 2 |
| 109 | P. pacifica Vitic. | Peltigeraceae | Bg | Fl | Terricolous | 2579 | - | 1 |
| 110 | P. polydactylon (Neck.) Hoffm. | Peltigeraceae | Bg | Fl | Terricolous | 2411 | 2750 | 8 |
| 111 | P. praetextata (Flörke ex Sommerf.) Zopf | Peltigeraceae | Bg | Fl | Terricolous | 2579 | 2800 | 7 |
| 112 | P. rufescens (Weiss) Humb. | Peltigeraceae | Bg | Fl | Terricolous | 2822 | 3100 | 5 |
| 113 | P. venosa (L.) Hoffm. | Peltigeraceae | Bg | Fl | Terricolous | 2579 | 2700 | 5 |
| 114 | Pertusaria australis Vain. | Pertusariaceae | Gr | Cr | Terricolous | 2494 | - | 1 |
| 115 | P. multipuncta (Turner) Nyl. | Pertusariaceae | Gr | Cr | Corticolous | 1798 | - | 1 |
| 116 | P. oculata (Dicks.) Th. Fr. | Pertusariaceae | Gr | Cr | Terricolous | 2450 | 2000 | 2 |
| 117 | P. qilianensis Q. Ren & Z.T. Zhao | Pertusariaceae | Gr | Cr | Terricolous | 2201 | 2400 | 2 |
| 118 | Phaeophyscia ciliata (Hoffm.) Moberg. | Physciaceae | Gr | Fl | Corticolous | 1760 | 2700 | 2 |
| 119 | P. sciastra (Ach.) Moberg. | Physciaceae | Gr | Fl | Terricolous | 2774 | 3000 | 3 |
| 120 | Physconia grisea (Lam.) Poelt. | Physciaceae | Gr | Fl | Corticolous | 2000 | 2274 | 2 |
| 121 | P. muscigena (Ach.) Poelt. | Physciaceae | Gr | Fl | Terricolous | 2274 | 3400 | 4 |
| 122 | P. perisidiosa (Erichsen) Moberg. | Physciaceae | Gr | Fl | Corticolous | 1780 | 2274 | 2 |
| 123 | Physcia aipolia (Ehrh. ex Humb.) Fürnr. | Physciaceae | Gr | Fl | Corticolous | 1774 | 2850 | 2 |
| 124 | P. caesia (Hoffm.) Hampe ex Fürnr. | Physciaceae | Gr | Fl | Corticolous | 1774 | 2300 | 6 |
| 125 | P. dubia (Hoffm.) Lettau | Physciaceae | Gr | Fl | Corticolous | 1869 | 2350 | 7 |
| 126 | P. stellaris (L.) Nyl. | Physciaceae | Gr | Fl | Corticolous | 1711 | 1950 | 2 |
| 127 | P. tenella (Scop.) DC. | Physciaceae | Gr | Fl | Corticolous | 1563 | 1800 | 3 |
| 128 | Protoparmelia badia (Hoffm.) Hafellner | Parmeliaceae | Gr | Fl | Saxicolous | 1690 | - | 1 |
| 129 | Protoblastenia rupestris (Scop.) J. Steiner | Psoraceae | Gr | Cr | Saxicolous | 1406 | - | 1 |
| 130 | Protoparmeliopsis baicalensis (Zahlbr.) S.Y. Kondr. | Lecanoraceae | Gr | Pl | Saxicolous | 1711 | 2700 | 4 |
| 131 | P. garovaglii (Körb.) Arup, Zhao Xin & Lumbsch | Lecanoraceae | Gr | Pl | Saxicolous | 1860 | 2700 | 4 |
| 132 | P. muralis (Schreb.) M. Choisy | Lecanoraceae | Gr | Pl | Saxicolous | 1711 | 2900 | 8 |
| 133 | P. peltata Ramond ex Arup, Zhao Xin & Lumbsch | Lecanoraceae | Gr | Pl | Saxicolous | 1860 | 2900 | 4 |
| 134 | P. phaedrophthalma (Poelt) S.Y. Kondr. | Lecanoraceae | Gr | Pl | Saxicolous | 1711 | 3400 | 4 |
| 135 | Psora decipiens (Hedw.) Hoffm. | Psoraceae | Gr | Sq | Terricolous | 2822 | 3100 | 5 |
| 136 | Ramalina farinacea (L.) Ach. | Ramalinaceae | Gr | Fr | Saxicolous | 1548 | - | 1 |
| 137 | R. pollinaria (Westr.) Ach. | Ramalinaceae | Gr | Fr | Corticolous | 1548 | 2670 | 2 |
| 138 | R. sinensis Jatta | Ramalinaceae | Gr | Fr | Corticolous | 1579 | 2500 | 6 |
| 139 | Rhizocarpon disporum (Nägeli ex Hepp) Müll. Arg. | Ramalinaceae | Gr | Cr | Saxicolous | 2969 | 3000 | 2 |
| 140 | R. effguratum (Anzi) Th. Fr. | Ramalinaceae | Gr | Cr | Saxicolous | 2969 | 3400 | 2 |
| 141 | R. geographicum (L.) DC. | Ramalinaceae | Gr | Cr | Saxicolous | 2969 | 3400 | 4 |
| 142 | R. macrosporum Räsänen | Ramalinaceae | Gr | Cr | Saxicolous | 2969 | - | 1 |
| 143 | Rhizoplaca chrysoleuca (Sm.) Zopf | Ramalinaceae | Gr | Sq | Saxicolous | 2969 | 3400 | 2 |
| 144 | R. melanophthalma (DC.) Leuckert & Poelt | Ramalinaceae | Gr | Sq | Saxicolous | 2969 | 3400 | 4 |
| 145 | R. sundescrepans (Nyl.) R. Sant. | Ramalinaceae | Gr | Sq | Saxicolous | 1420 | 2969 | 3 |
| 146 | Rinodina cf. freyi H. Magn. | Physciaceae | Gr | Cr | Corticolous | 1438 | - | 1 |
| 147 | R. aff. oleae Bagl. | Physciaceae | Gr | Cr | Corticolous | 1438 | - | 1 |
| 148 | R. sarcogynoides H. Magn. | Physciaceae | Gr | Cr | Saxicolous | 1438 | - | 1 |
| 149 | R. sophodes (Ach.) A. Massal. | Physciaceae | Gr | Cr | Corticolous | 1438 | - | 1 |
| 150 | R. terrestris Tomin. | Physciaceae | Gr | Cr | Terricolous | 2438 | - | 1 |
| 151 | Rusavskia elegans (Link) S.Y. Kondr. & Kärnefelt | Teloschistaceae | Gr | Fl | Saxicolous | 1678 | 3400 | 5 |
| 152 | Sarcogyne picea H. Magn. | Acarosporaceae | Gr | Cr | Saxicolous | 1712 | 3000 | 2 |
| 153 | S. pruinosa (Ach.) A. Massal. | Acarosporaceae | Gr | Cr | Saxicolous | 1664 | - | 1 |
| 154 | Sporastatia testudinea (Ach.) A. Massal. | Sporastatiaceae | Gr | Cr | Saxicolous | 2400 | 2865 | 2 |
| 155 | Umbilicaria virginis Schrad. | Umbilicariaceae | Gr | Fl | Saxicolous | 1822 | 3200 | 8 |
| 156 | Usnea baileyi (Stirt.) Zahlbr. | Parmeliaceae | Gr | Fr | Corticolous | 1808 | - | 1 |
| 157 | U. ceratina Ach. | Parmeliaceae | Gr | Fr | Corticolous | 2230 | - | 1 |
| 158 | U. lapponica Vain. | Parmeliaceae | Gr | Fr | Corticolous | 1530 | 1664 | 1 |
| 159 | U. subfloridana Stirt. | Parmeliaceae | Gr | Fr | Corticolous | 2230 | 2700 | 2 |
| 160 | Verrucaria fuscella (Turner) Winch | Verrucariaceae | Gr | Cr | Saxicolous | 1684 | - | 1 |
| 161 | V. nigrescens Pers. | Verrucariaceae | Gr | Cr | Saxicolous | 1684 | 2400 | 2 |
| 162 | Xanthomendoza allfiridii (S.Y. Kondr. & Poelt) Søchting, Kärnefelt & S.Y. Kondr. | Teloschistaceae | Gr | Sq | Corticolous | 2274 | - | 1 |
| 163 | X. ullophylodes (Räsänen) Søchting, Kärnefelt & S.Y. Kondr. | Teloschistaceae | Gr | Sq | Corticolous | 1711 | - | 1 |
| 164 | Xanthoparmelia camtschadalis (Ach.) Hale. | Parmeliaceae | Gr | Fl | Saxicolous | 1570 | 2300 | 4 |
| 165 | X. chlorochroa (Tuck.) Hale | Parmeliaceae | Gr | Fl | Terricolous | 2711 | - | 1 |
| 166 | X. coreana (Gyeln.) Hale. | Parmeliaceae | Gr | Fl | Saxicolous | 1684 | 3300 | 3 |
| 167 | X. somloensis (Gyeln.) Hale | Parmeliaceae | Gr | Fl | Saxicolous | 1684 | 3400 | 5 |
| 168 | X. stenophylla (Ach.) Ahti & D. Hawksw. | Parmeliaceae | Gr | Fl | Saxicolous | 1690 | - | 1 |
| 169 | X. wyomingica (Gyelnic) Hale | Parmeliaceae | Gr | Fl | Saxicolous | 1711 | 3100 | 2 |
| 170 | Xanthoria fallax Arnold | Teloschistaceae | Gr | Fl | Corticolous | 1711 | 2450 | 2 |
| 171 | X. fulva (Hoffm.) Poelt & Petut. | Teloschistaceae | Gr | Fl | Saxicolous | 1579 | 3200 | 4 |
| 172 | X. parietina (L.) Th. Fr. | Teloschistaceae | Gr | Fl | Saxicolous | 1579 | 2980 | 3 |
| 173 | Xylographa parallela (Ach.) Fr. | Xylographaceae | Gr | Cr | Corticolous | 1822 | 2846 | 2 |
Note: Gr: green algae; Bg: blue-green algae; Cr: crustose; Fl: foliose; Fr: fruticose; Sq: squamulose; Pl: placodioi; Dm: dimorphic.
References
- IPBES. Global Assessment Report on Biodiversity and Ecosystem Services, IPBES Secretariat, Bonn, Germany. 2019. Available online: https://www.ipbes.net/global-assessment (accessed on 3 June 2019).
- Wei, Q.; Yang, H.L. A Brief Discussion on the Challenges and Countermeasures of Global Biodiversity Conservation. Biodivers. Conserv. Green Dev. 2024, 1, 11–16. [Google Scholar]
- Ma, K.P.; Ren, H.; Long, C.L. Biodiversity conservation needs more research. Guihaia 2023, 43, 1347–1349. (In Chinese) [Google Scholar]
- Mi, X.C.; Feng, G.; Hu, Y.B.; Zhang, J.; Chen, L.; Corlett, R.T.; Hughes, A.C.; Pimm, S.; Schmid, B.; Shi, S.H.; et al. The global significance of biodiversity science in China: An overview. Natl. Sci. Rev. 2021, 8, 2–25. [Google Scholar] [CrossRef] [PubMed]
- Hawksworth, D.L. The fungal partner. In CRC Handbook of Lichenology; Galun, M., Ed.; CRC Press: Boca Raton, FL, USA, 1988; Volume 1, pp. 35–38. [Google Scholar]
- Abas, A.; Khalid, R.M.; Rosandy, A.R.; Sulaiman, N. Lichens of Pulau Pangkor, Perak, Malaysia. Malays. For. 2019, 82, 59–66. [Google Scholar]
- Irwin, M.B.; Sylvia, D.S.; Stephen, S. Lichens of North America; Yale Univeristy Press: New Haven, CT, USA, 2001; p. 828. [Google Scholar]
- Feuerer, T.; Hawksworth, D.L. Biodiversity of lichens, including a world-wide analysis of checklist data based on Takhtajan’s floristic regions. Biodivers. Conserv. 2007, 16, 85–98. [Google Scholar] [CrossRef]
- Chen, J.; Blume, H.; Beyer, L. Weathering of rocks induced by lichen colonization—A review. Catena 2000, 39, 121–146. [Google Scholar] [CrossRef]
- Chen, J.B. The particularity and biological diversity of lichens, and their importance to science and application. Biodivers. Sci. 1995, 3, 113–117. (In Chinese) [Google Scholar]
- Wei, J.C. A review on the present situation of lichenology in China. Mycosystema 2018, 38, 812–818. (In Chinese) [Google Scholar]
- Wei, X.L.; Deng, H.; Wei, J.C. Threatened categories assessment of lichens in China. Biodivers. Sci. 2020, 28, 54–65. (In Chinese) [Google Scholar]
- Wei, J.C. An Enumeration of Lichens in China; International Academic Publishers: Beijing, China, 1991; pp. 1–278. [Google Scholar]
- Tumur, A.; Mamut, R.; Seaward, M.R.D. An updated checklist of lichens of Xinjiang Province, China. Herzogia 2021, 34, 62–93. [Google Scholar] [CrossRef]
- Xiao, W.F.; Yasheng, A.; Qian, F.W.; Xu, H.X.; Cheng, K.W.; Liu, W.C. Biodiversity and Nature Conservation Management of Western Tianshan National Nature Reserve, Xinjiang; China Forestry Publishing House: Beijing, China, 2015; pp. 1–6. [Google Scholar]
- Richardson, D.H.S. Lichens and man. In Frontiers in Mycology; Hawksworth, D.L., Ed.; International Mycological Institute: London, UK, 1991; pp. 187–210. [Google Scholar]
- Elenkin, A.A. Migratory lichens of the deserts and steppes. In Bulletin du Jardin Imperial Botanique de St. Petersburg; University of Michigan Library: Ann Arbor, MI, USA, 1901; pp. 16–38. [Google Scholar]
- Elenkin, A.A. Lichenes Florae Rossiae et Regionum Confinium Orientalium. Fasciculus I, No. 1–50. Acta Horti Petropolitani 1901, 19, 1–52. [Google Scholar]
- Vainio, E.A. Lichenes ab Ove Paulsen praecipue in provencia Ferghana (Asia Media) et a Boris Fedtschenko in Tjanschan anno 1898 et 1899 collecti. Bot. Tidsskr. 1905, 26, 241–250. [Google Scholar]
- Yang, X.R.; Song, X.H.; Peng, L.; Shi, X.G.; Akemu, S.; Ye, C.X. Lichens from Qiaxi National Conservation for Picea schrenkiana of Gongliu County in Ili, Xinjiang, China. J. Zhongshan Univ. 2008, 47, 85–90. (In Chinese) [Google Scholar]
- Tumur, A.; Abbas, A. DCA ordination of ecological distribution of saprophytic lichens in forest ecosystem of Tianshan. J. Plant Resour. Environ. 2005, 11, 41–45. (In Chinese) [Google Scholar]
- Tumur, A.; Abdulla, A.; Abbas, A. Numerical classification and species diversity of corticolous lichens communities in forest ecosystem of the Tianshan Mountains. Acta Phytoecol. Sin. 2005, 29, 615–622. (In Chinese) [Google Scholar]
- Yue, X. Protecting the biodiversity of western Tianshan National Nature Reserve. China For. 2009, 13, 60. (In Chinese) [Google Scholar]
- Abbas, A.; Wu, J.R. Lichens of Xinjiang; Science Technology & Hygiene Publishing House of Xinjiang: Urumqi, China, 1998; pp. 1–16. [Google Scholar]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens, 2nd ed.; British Lichen Society: London, UK, 2010; pp. 1–101. [Google Scholar]
- Baniya, C.B.; Solhoy, T.; Gauslaa, Y.; Palmer, M.W. The elevation gradient of lichen species richness in Nepal. Lichenologist 2010, 42, 83–96. [Google Scholar] [CrossRef]
- Rai, H.; Khare, R.; Baniya, C.B.; Upreti, D.K.; Gupta, R.K. Elevational gradients of terricolous lichen species richness in the Western Himalaya. Biodivers. Conserv. 2015, 24, 1155–1174. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; El-Ghwas, D.E.; Daba, G.M. Lichens uses: Surprising uses of lichens that improve human life. J. Biomed. Res. Environ. Sci. 2022, 3, 189–194. [Google Scholar]
- Nascimbene, J.; Marini, L. Epiphytic lichen diversity along elevational gradients: Biological traits reveal a complex response to water and energy. J. Biogeogr. 2015, 42, 1222–1232. [Google Scholar] [CrossRef]
- Gauslaa, Y.; Palmqvist, K.; Solhaug, K.A.; Hilmo, O.; Holien, H.; Nybakken, L.; Ohlson, M. Size dependent growth in two old-growth associated macrolichen species. New Phytol. 2009, 18, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Brunialti, G.; Giordani, P. Variability of lichen diversity in a climatically heterogeneous area (Ligaria, NW Italy). Lichenologist 2003, 35, 55–69. [Google Scholar] [CrossRef]
- Pinokiyo, A.; Singh, K.P.; Sigh, J.S. Diversity and distribution of lichens in relation to altitude within a protected biodiversity hot spot, north-east India. Lichenologist 2008, 40, 47–62. [Google Scholar] [CrossRef]
- Jonsson, B.G.; Jonsell, M. Exploring potential biodiversity indicators in boreal forests. Biodivers. Conserv. 1999, 8, 1417–1433. [Google Scholar] [CrossRef]
- Moen, J.; Jonsson, B.G. Edge effects on liverworts and lichens in forest patches in a mosaic of boreal forest and wetland. Conserv. Biol. 2003, 17, 380–388. [Google Scholar] [CrossRef]
- Rai, H.; Khare, R.; Gupta, R.K.; Upreti, D.K. Terricolous lichens as indicator of anthropogenic disturbances in a high altitude grassland in Garhwal (Western Himalaya), India. Bot. Orient. J. Plant Sci. 2012, 8, 16–23. [Google Scholar] [CrossRef]
- Rai, H.; Upreti, D.K.; Gupta, R.K. Diversity and distribution of terricolous lichens as an indicator of habitat heterogeneity and grazing induced trampling in a temperate-alpine shrub and meadow. Biodivers. Conserv. 2012, 21, 97–113. [Google Scholar] [CrossRef]
- Rawat, D.S. Elevational reduction of plant species diversity in high altitudes of Garhwal Himalaya, India. Curr. Sci. 2011, 100, 833–836. [Google Scholar]
- Rose, F. Lichenological indicators of age and environmental continuity in woodlands. In Lichenology: Progress and Problems; Brown, D.H., Hawksworth, D.L., Bailey, R.H., Eds.; Academic Press: London, UK, 1976; pp. 279–307. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).