
New Records of Marine Tardigrades (Arthotardigrada) from the Iberian Peninsula: Biogeographical Implications




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
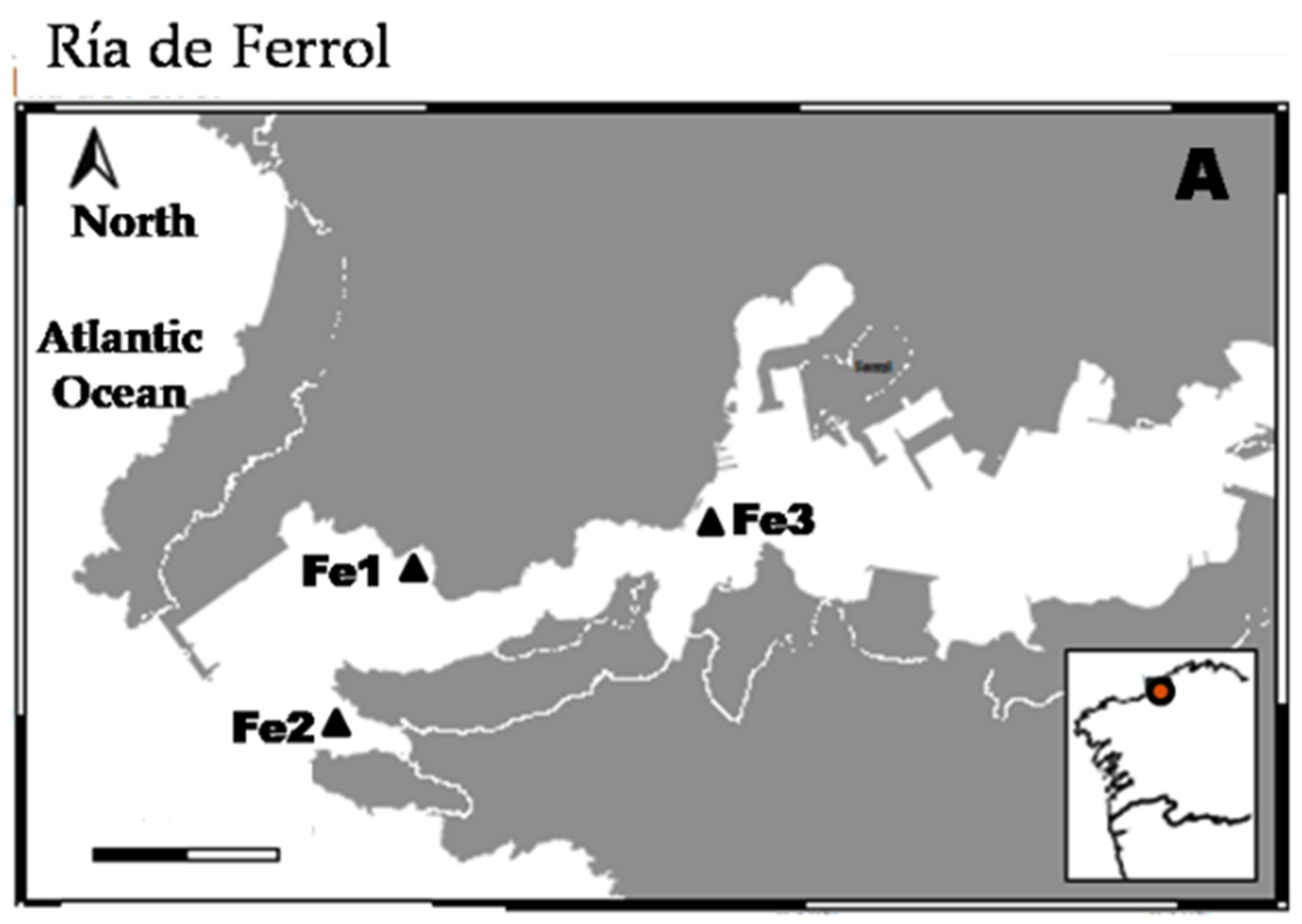
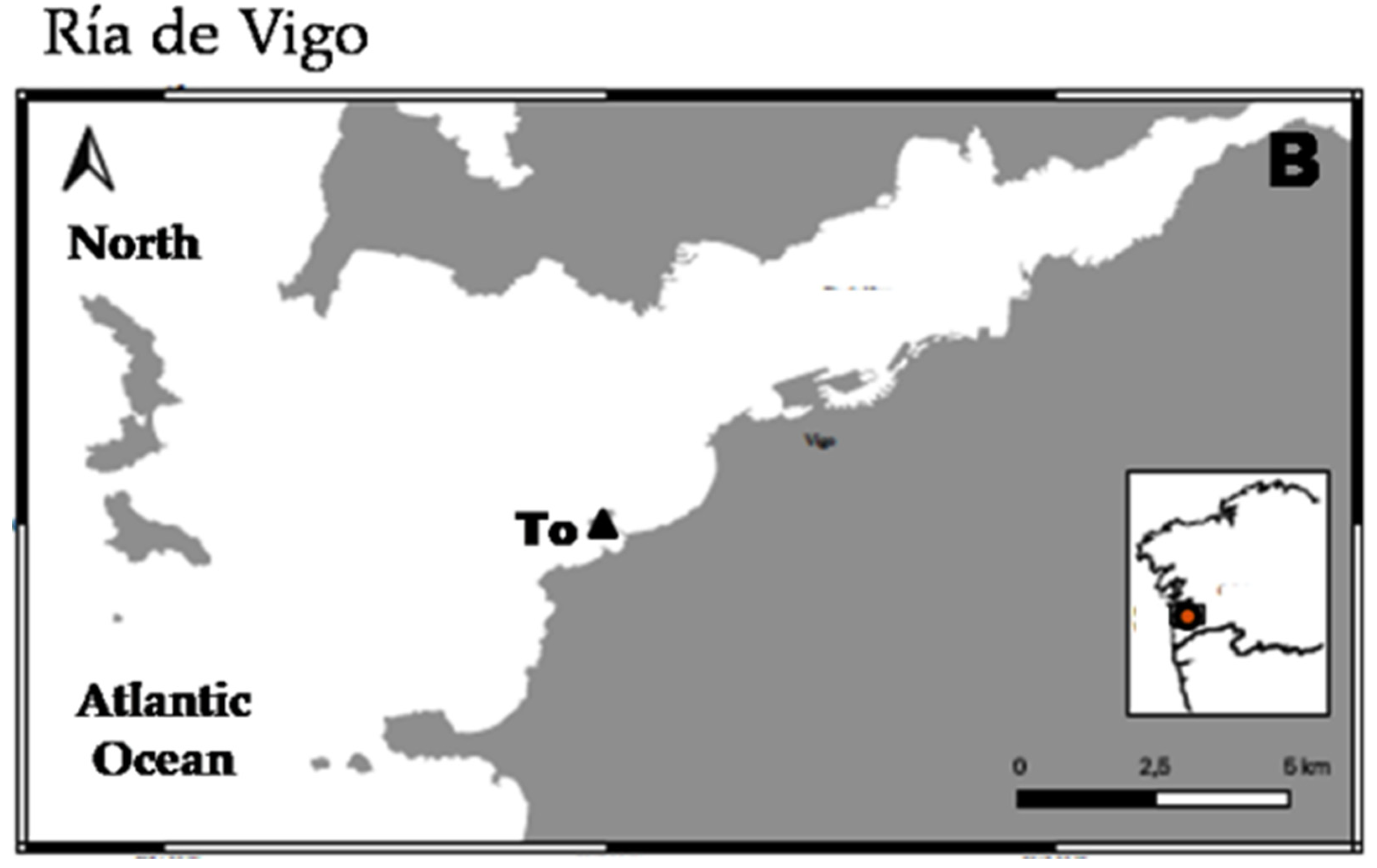
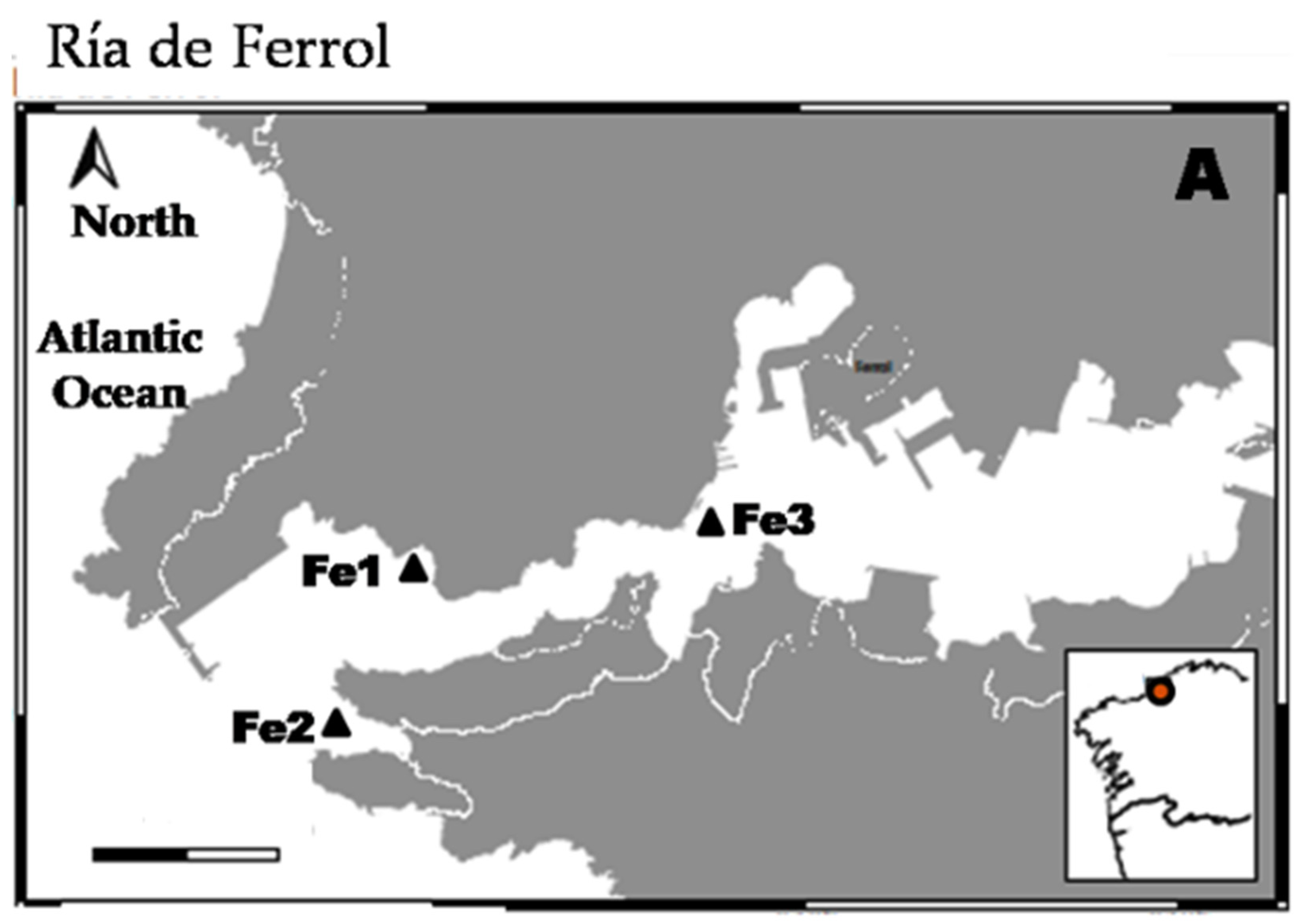
2.1. Studied Area and Sampling
2.2. Laboratory Methodology and Systematics
3. Results
- Phylum Tardigrada Doyère, 1840 [16]
- Class Heterotardigrada Marcus, 1927 [17]
- Order Arthrotardigrada Marcus, 1927 [17]
- Genus Batillipes Richters, 1909 [6]
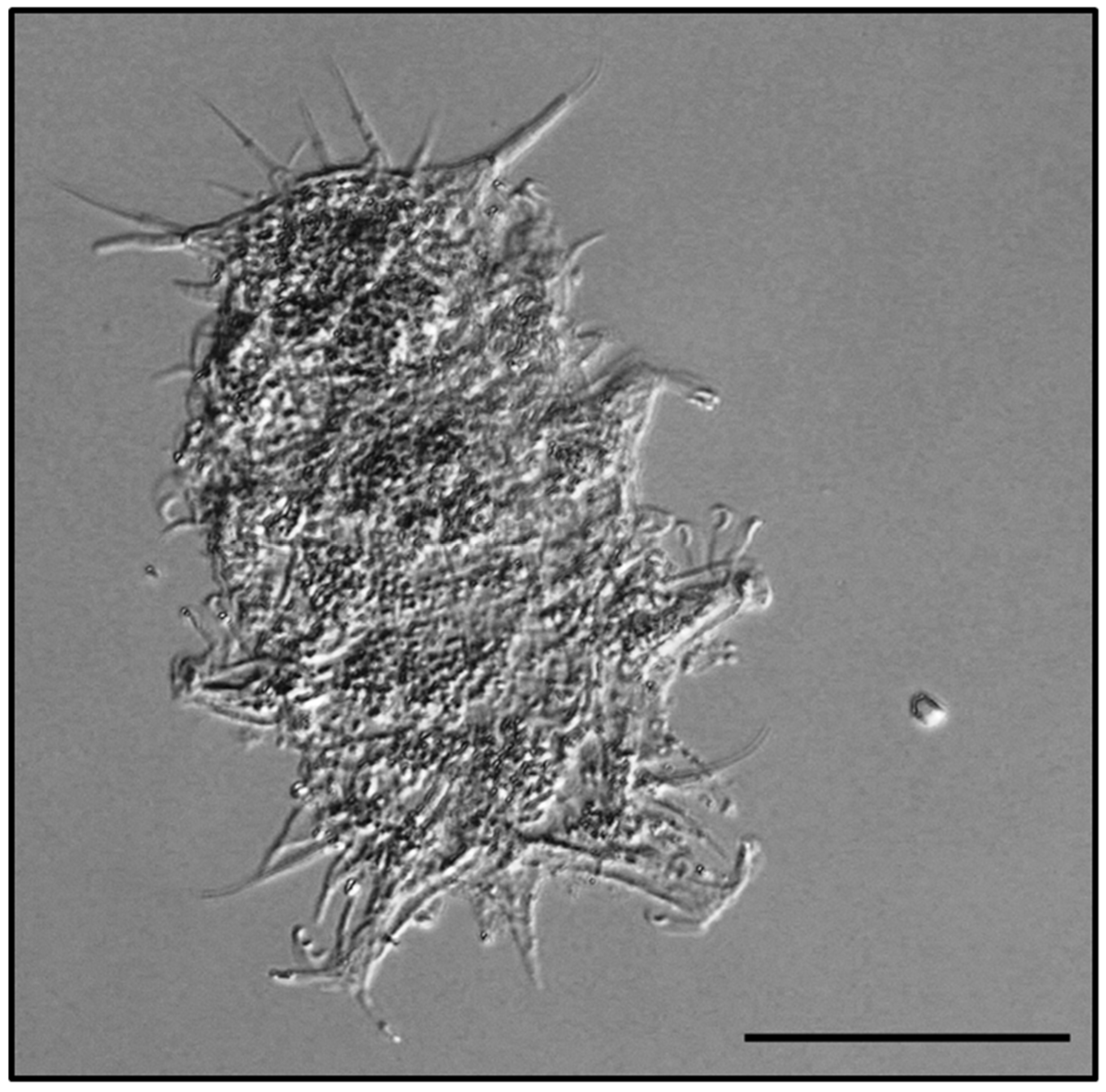
3.1. Batillipes Marcelli Morone De Lucia, D’Addabbo Gallo and Grimaldi de Zio, 1988 [20]
3.1.1. Material Examined
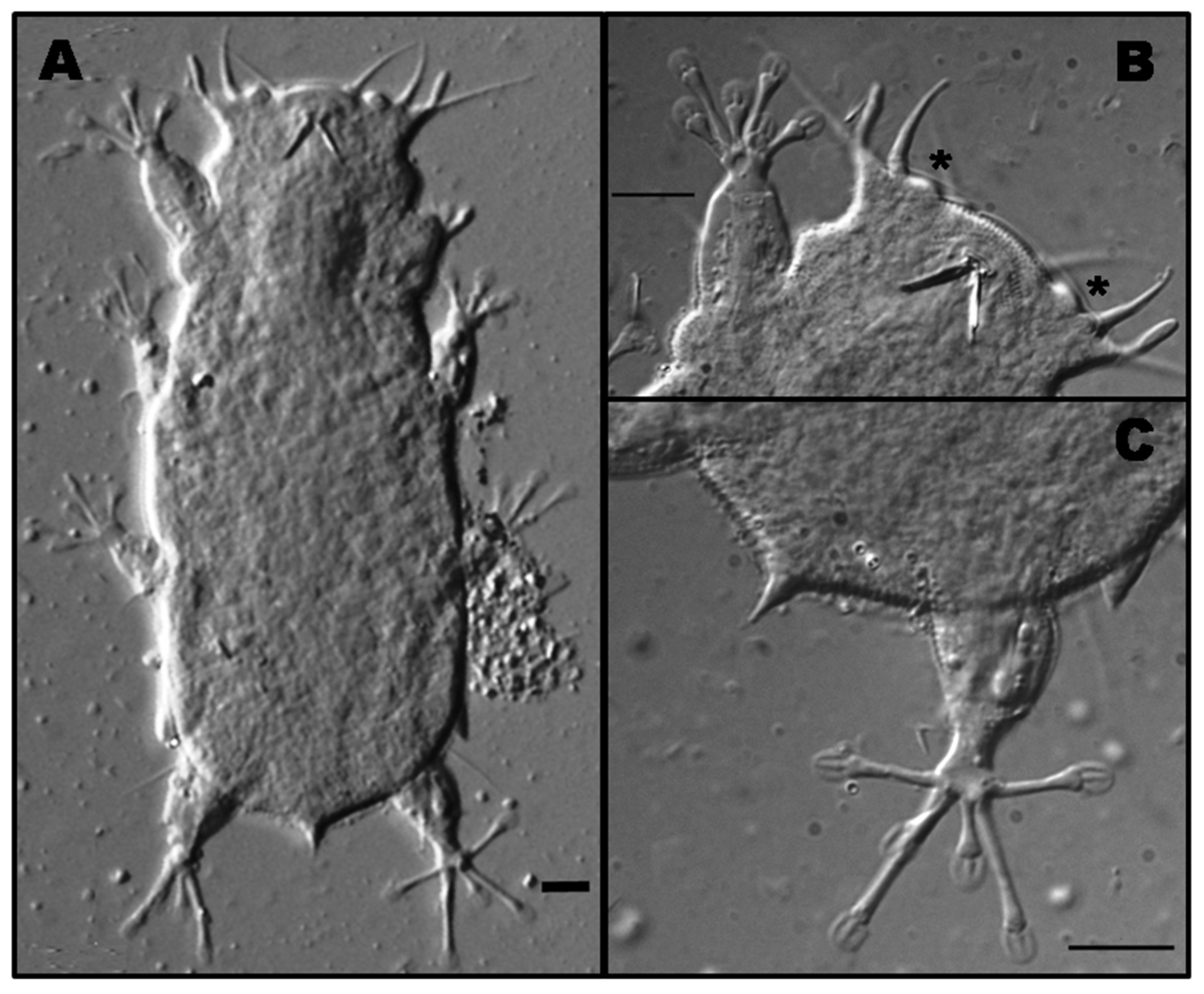
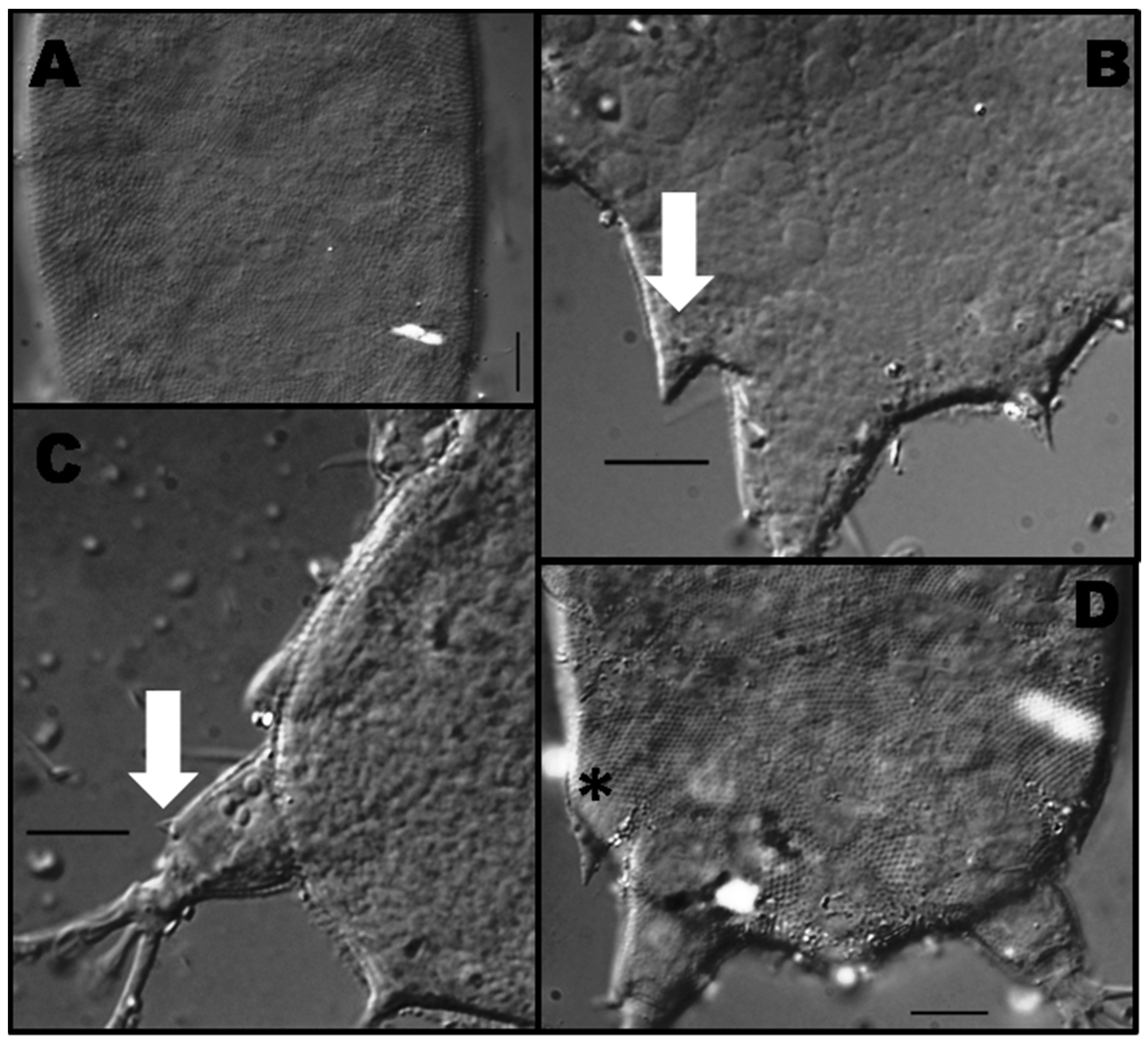
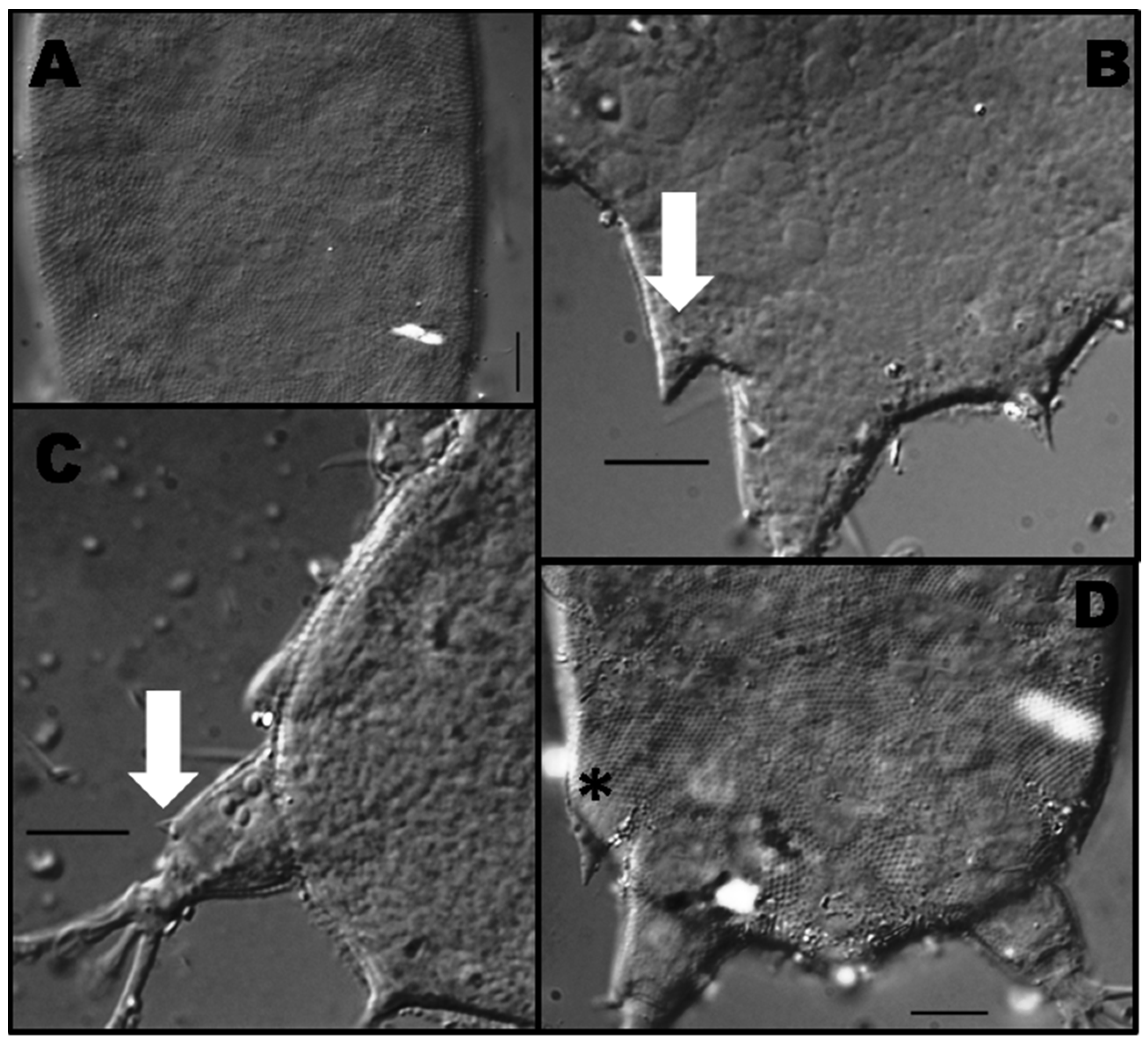
3.1.2. Morphological and Ecological Remarks
3.1.3. Distribution
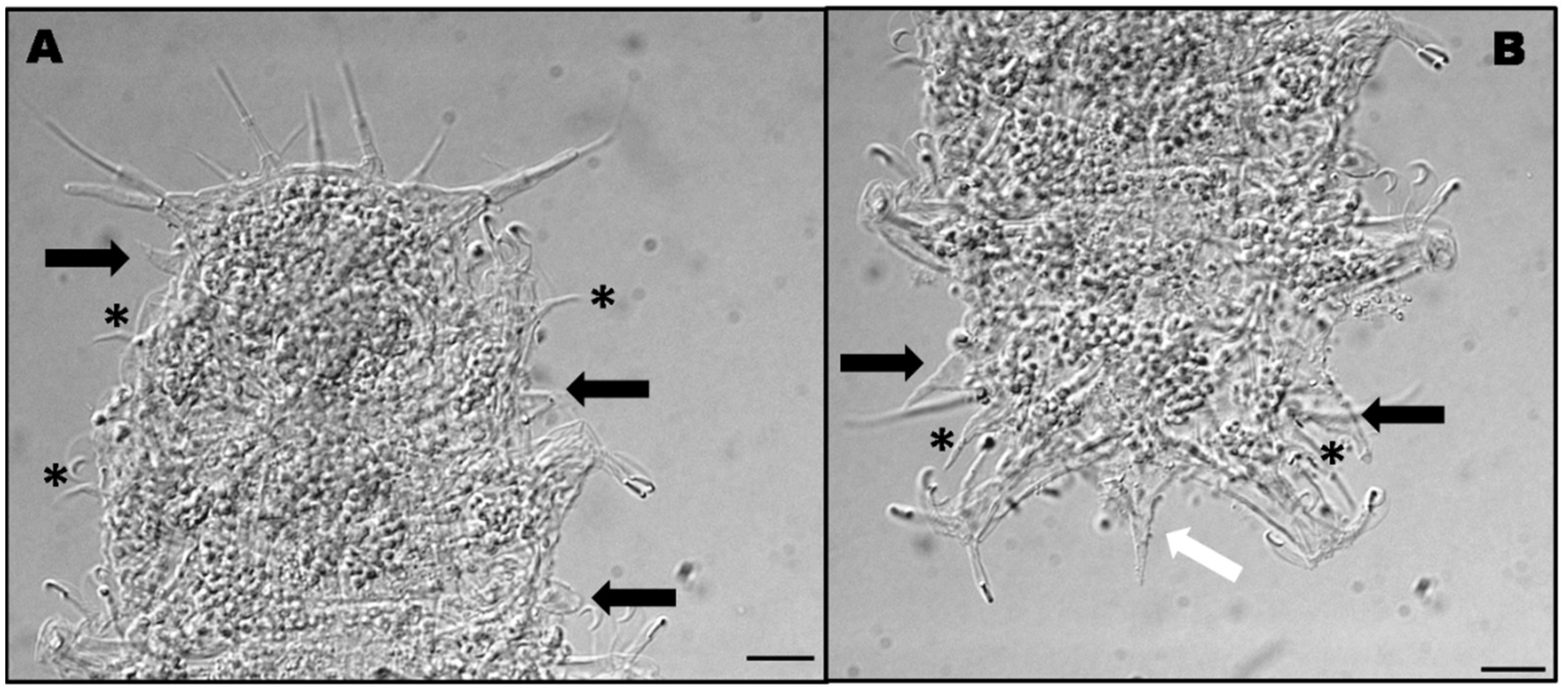
3.2. Halechiniscus Remanei Remanei Schulz, 1955 [8]
3.2.1. Material Examined
3.2.2. Morphological and Ecological Remarks
3.2.3. Distribution
- Genus: Actinarctus Schulz, 1935 [31]
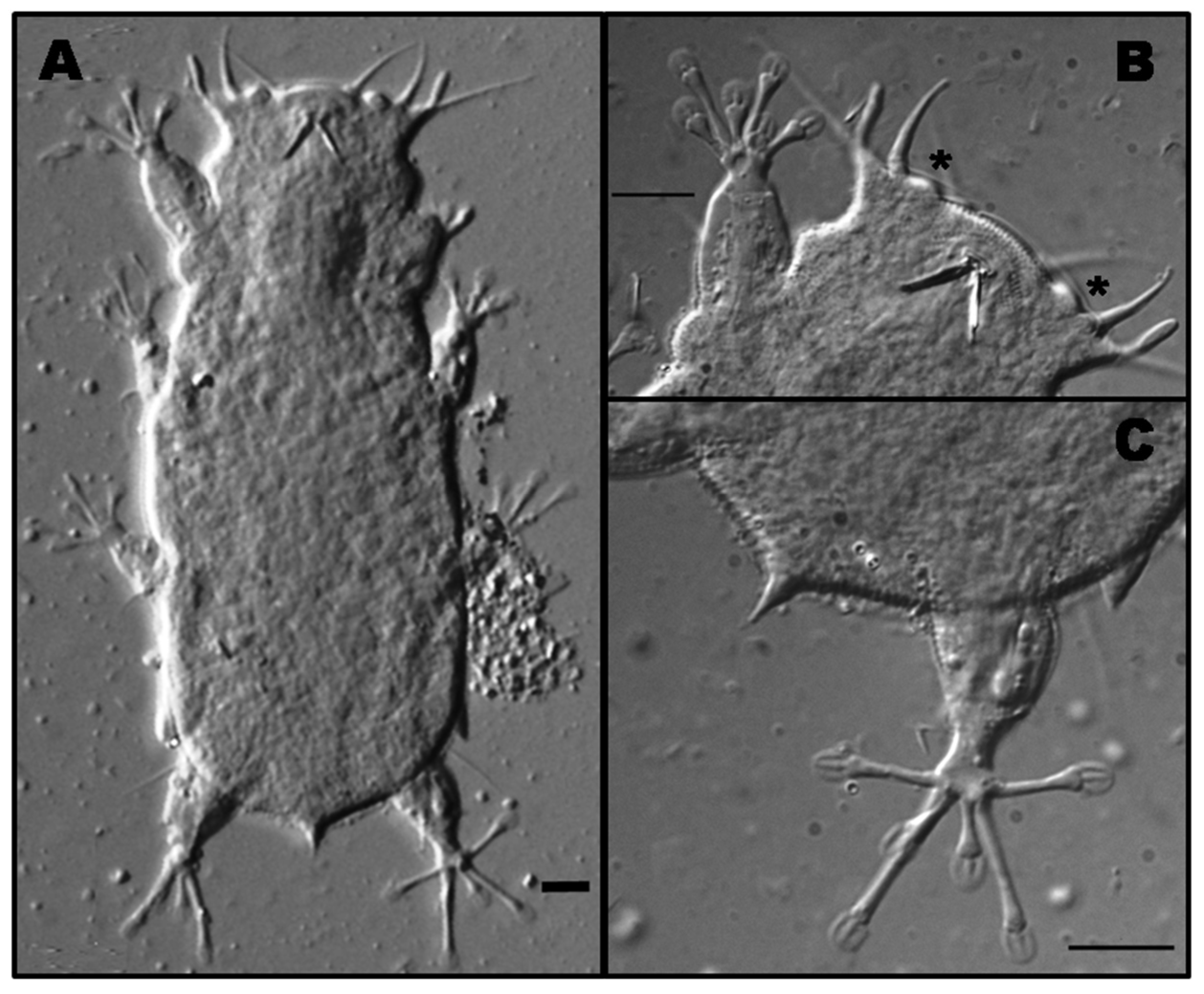
3.3. Actinarctus Doryphorus Doryphorus Schulz, 1935 [31]
3.3.1. Material Examined
3.3.2. Morphological and Ecological Remarks
3.3.3. Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Measurements in μm of Different Structures in the Studied Specimens of Batillipes marcelli
| Specimen | Bm1 Female | Bm2 Female | Bm3 Indet. | Bm4 Indet. |
| Body length | 123.7 | 144.0 | 92 | 177 |
| Body width | 52.3 | 56.7 | 34.5 | 78 |
| Buccal tube | 15.1 | 19 | ||
| Median cirrus | 11.3 | 11.5 | ||
| Internal cirri | 16.5 | 17.4 | 14.1 | 17.4 |
| External cirri | 13.5 | 15.7 | 10.6 | 17.5 |
| Lateral cirri | 21.8 | 21.7 | 18.8 | 23.6 |
| Primary clavae | 8.5 | 9.7 | 7.4 | 11.7 |
| Cirri E | 15.7 | 21.6 | 17.5 | 17.3 |
| Leg IV base | 5.9 | 6 | 4.7 | 5.8 |
| Leg IV spine | 7.8 | 5.3 | 4.4 | 7.1 |
| Leg III spine | 8.0 | 10.6 | ||
| Leg II spine | 8.9 | 10.7 | ||
| Leg I spine | 7.7 | 8.7 | 7.8 | |
| Projection 1 | 3.1 | 4.9 | 3.5 | |
| Projection 2 | 2.9 | 2.7 | ||
| Projection3 | 4.4 | 3.9 | 3.1 | 3.8 |
| Projection4 | 4.5 | 3.6 | 6.1 | 7.1 |
| Caudal appendage (L) | 6.6 | 6.5 | 3.8 | 5.6 |
| Leg I (toe 1) | 8.6 | 5.4 | 7.0 | |
| (toe 2) | 5.7 | 3.8 | 4.8 | |
| (toe 3) | 12.8 | 9.5 | 13.1 | |
| (toe 4) | 9.1 | 5 | 7.9 | |
| (toe 5) | 9.4 | 10 | 15.0 | |
| (toe 6) | 14.5 | 5.8 | 8.9 | |
| Leg IV (toe 1) | 12.2 | 11.7 | 9.4 | 12.6 |
| (toe 2) | 16.7 | 15.0 | 13.7 | 18.2 |
| (toe 3) | 8.2 | 7.1 | 7.6 | 11.3 |
| (toe 4) | 9.3 | 8.1 | 7.4 | 11.6 |
| (toe 5) | 16.7 | 17.4 | 13.7 | 21.0 |
| (toe 6) | 11.2 | 10 | 6.8 | 12.6 |
Appendix B. Measurements in μm of Different Structures in the Studied Specimens of Halechiniscus remanei remanei
| Specimen | Hr1 Male | Hr2 Indet. | Hr3 Indet. | Hr4 Indet. |
| Body length | 102.0 | 103.0 | 114.0 | 113.4 |
| Median cirrus | 21.9 | 13.7 | 15.7 | 16.4 |
| Internal cirri | 26.6 | 22.4 | 24.5 | 25.8 |
| External cirri | 22.5 | 21.7 | 15.2 | 16.3 |
| Lateral cirri | 29.2 | 27.2 | 30.3 | 32.0 |
| Primary clavae | 13.2 | 14.5 | 14.1 | 14.8 |
| Cirri E | 26.6 | 29.5 | 23.4 | |
| Leg IV base | 9.1 | 7.6 | 9.0 | |
| Leg IV spine | 4.6 | 5.3 | 6.1 | |
| Leg III spine | 10.0 | |||
| Leg II spine | 9.7 | 10.4 | ||
| Leg I spine | 6.7 | 9.4 | 9.8 | |
| Projection 1 | 5.4 | 5.1 | 5.9 | 9 |
| Projection 2 | 6.6 | 6.3 | ||
| Projection3 | 7.8 | 13.3 | 9.1 | |
| Projection4 | 9.9 | 14.4 | 13.1 | 15.9 |
| Caudal appendage (L) | 13.8 | 16.9 | 18.3 | |
| Claw | 3.6 | 3.9 | ||
| Digit and claw | 11.8 | 11.3 | 11.8 |
Appendix C. Measurements in μm of Different Structures in the Studied Specimens of Actinarctus doryphorus doryphorus
| Specimen | Ad1 Female | Ad2 Male | Ad3 Male | Ad4 Male |
| Body length | 119.1 | 76.1 | 98.8 | 114.7 |
| Body width | 58.7 | 38.1 | 52.1 | 71.9 |
| Median cirrus | 35.0 | |||
| Internal cirri | 82.8 | 58.9 | 67.2 | 78.4 |
| External cirri | 55.0 | 43.2 | 48.8 | 51.6 |
| Lateral cirri | 16.3 | 29.1 | 41.8 | 43.2 |
| Primary clavae | 12.1 | 92.2 | 150.1 | 170.5 |
| Buccal clavae | 24.8 | 24.9 | ||
| Cirri E | 48.4 | |||
| Leg IV | 96.3 | 95.7 | 1.5.5 | |
| Leg III spine | ||||
| Leg II spine | ||||
| Leg I spine | 11.2 | |||
| Frontal ala | 35.4 | 22.2 | 34.3 | 38.9 |
| Lateral ala | 71.3 | 49.7 | 62.2 | 64.8 |
| Caudal ala | 47.4 | 32.7 | 39.1 | 43.5 |
| Leg I | ||||
| External digit | 14.1 | 10.4 | 13.2 | 16.1 |
| Internal digit | 18.4 | 14.5 | 17.2 | 20.4 |
| Leg IV | ||||
| External digit | 18.5 | 12.4 | 17.3 | 16.0 |
| Internal digit | 22.7 | 16.4 | 22.1 | 22.3 |
References
- Nelson, D.R.; Bartels, P.J.; Guil, N. Tardigrade Ecology. In Water Bears: The Biology of Tardigrades; Schill, R.O., Ed.; Zoological Monographs 2; Springer Nature Switzerland AG: Cham, Switzerland, 2018. [Google Scholar]
- Kaczmarek, Ł.; Bartels, P.J.; Roszkowska, M.; Nelson, D.R. The Zoogeography of Marine Tardigrada. Zootaxa 2015, 4037, 1–189. [Google Scholar] [CrossRef]
- de Zio Grimaldi, S.; Gallo D’Addabbo, M.; Sandulli, R.; D’Addabbo, R. Checklist of the Italian marine Tardigrada. Meiofauna Mar. 2003, 12, 97–135. [Google Scholar]
- Rubal, M.; Veiga, P.; Fontoura, P.; Santos, E.; Sousa-Pinto, I. Biodiversity of marine tardigrades from the northern coast of Portugal (Iberian Peninsula). Zool. J. Linn. Soc. 2016, 178, 747–754. [Google Scholar] [CrossRef]
- Santos, E.; Rubal, M.; Veiga, P.; da Rocha, C.M.C.; Fontoura, P. Batillipes (Tardigrada, Arthrotardigrada) from the Portuguese coast with the description of two new species and a new dichotomous key for all species. Eur. J. Taxon. 2018, 425, 1–32. [Google Scholar] [CrossRef]
- Richters, F. Marine Tardigraden. Verh. Der Dtsch. Zool. Ges. 1909, 19, 84–95. [Google Scholar]
- Marcus, E. Batillipes pennaki, a new marine tardigrade from the North and South American Atlantic coast. Comun. Zoológicas Del Mus. Hist. Nat. Montev. 1946, 2, 1–3. [Google Scholar]
- Schulz, E. Studien an marinen tardigraden. Kiel. Meeresforsch. 1955, 11, 74–79. [Google Scholar]
- Du Bois-Reymond Marcus, E. On South American Malacopoda. Bol. Da Fac. De Filos. Ciências E Let. Da Univ. De São Paulo Ser. Zool. 1952, 17, 189–209. [Google Scholar]
- Schultze, M. 1865. Echiniscus sigismundi, ein Arctiscoide der Nordsee. Arch. Mikr. Anat. 1865, 1, 428–436. [Google Scholar] [CrossRef]
- Santos, E.; Veiga, P.; Rubal, M.; Bartels, P.J.; Da Rocha, C.M.C.; Fontoura, P. Batillipes pennaki Marcus, 1946 (Arthrotardigrada: Batillipedidae): Deciphering a species complex. Zootaxa 2019, 4648, 549–567. [Google Scholar] [CrossRef]
- Faurby, S.; Jørgensen, A.; Kristensen, R.M.; Funch, P. Phylogeography of North Atlantic intertidal tardigrades: Refugia, cryptic speciation and the history of the Mid-Atlantic Islands. J. Biogeogr. 2011, 38, 1613–1624. [Google Scholar] [CrossRef]
- Faurby, S.; Jørgensen, A.; Kristensen, R.M.; Funch, P. Distribution and speciation in marine intertidal tardigrades: Testing the roles of climatic and geographical isolation. J. Biogeogr. 2012, 39, 1596–1607. [Google Scholar] [CrossRef]
- Fontoura, P.; Bartels, P.J.; Jørgensen, A.; Kristensen, R.M.; Hansen, J.G. A dichotomous key to the genera of the Marine Heterotardigrades (Tardigrada). Zootaxa 2017, 4294, 1–45. [Google Scholar] [CrossRef]
- Perry, E.; Miller, W.R.; Kaczmarek, L. Recommended abbreviations for the names of genera of the phylum Tardigrada. Zootaxa 2019, 4608, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Doyère, L.M. Mémoire sur les Tardigrades. I. Ann. Sci. Nat. Série 2 1840, 14, 269–361. [Google Scholar]
- Marcus, E. Zur Anatomie und Ökologie mariner Tardigraden. Zool. Jahrbücher. Abt. Für Syst. Ökologie Und Geogr. Der Tiere 1927, 53, 487–588. [Google Scholar]
- Ramazzotti, G. Il Phylum Tardigrada. Mem. Dell’istituto Ital. Idrobiol. 1962, 14, 1–595. [Google Scholar]
- Gallo D’Addabbo, M.; Sandulli, R.; Grimaldi de Zio, S. New Batillipedidae (Tardigrada, Heterotardigrada) from the Orosei Gulf, Sardinia, Tyrrhenian Sea. Zool. Anz. 2005, 243, 219–225. [Google Scholar] [CrossRef]
- Morone De Lucia, R.M.; D’Addabo Gallo, M.; Grimaldi de Zio, S. Descrizione di due nuove specie di Batillipedidae (Tardigrada: Heterotardigrada). Cah. Biol. Mar. 1988, 29, 361–373. [Google Scholar]
- Thulin, G. Über die Phylogenie und das System der Tardigraden. Hered. Genet. Arkiv 1928, 11, 207–266. [Google Scholar] [CrossRef]
- Fujimoto, S.; Jørgensen, A.; Hansen, J.G. A molecular approach to Arthrotardigrade phylogeny (Heterotardigrada, Tardigrada). Zool. Scr. 2017, 46, 496–505. [Google Scholar] [CrossRef]
- Grimaldi de Zio, S.; D’Addabbo Gallo, M.; Morone De Lucia, R.M. Revision of the genus Halechiniscus (Halechiniscidae, Arthrotardigrada). Cah. Biol. Mar. 1990, 31, 271–279. [Google Scholar]
- Richters, F. Marine Tardigraden. Zool. Anz. 1908, 33, 77–85. [Google Scholar]
- Bartels, P.J.; Fontoura, P.; Nelson, D.R.; Orozco-Cubero, S.; Mioduchowska, M.; Gawlak, M.; Kaczmarek, Ł.; Cortés, J. A trans-isthmus survey of marine tardigrades from Costa Rica (Central America) with descriptions of seven new species. Mar. Biol. Res. 2021, 17, 120–166. [Google Scholar] [CrossRef]
- Miller, W.R.; Perry, E.S. The coastal marine Tardigrada of the Americas. Zootaxa 2016, 4126, 375–396. [Google Scholar] [CrossRef] [PubMed]
- Renaud-Mornant, J. Nouveaux Arthrotardigrades des Antilles. Bull. Muséum Natl. d’Histoire Nat. Série 4e 1984, 6, 975–988. [Google Scholar]
- Grimaldi de Zio, S.; D’Addabbo Gallo, M.; Morone De Lucia, M.R.; D’Addabbo, L. Marine Arthrotardigrada and Echiniscoidea (Tardigrada, Heterotardigrada) from the Indian Ocean. Boll. Zool. 1987, 4, 347–357. [Google Scholar] [CrossRef]
- McGinty, M. Batillipes gilmartini, a new marine tardigrade from a California beach. Pac. Sci. 1969, 23, 394–396. [Google Scholar]
- Renaud-Mornant, J. Description de trois espécies nouvelles du genre Tanarctus Renaud-Debyser, 1959, et création de la sous-famille des Tanarctinae subfam. nov. (Tardigrada, Heterotardigrada). Bull. Muséum Natl. d’Histoire Nat. Sér 4e 1980, 2, 129–141. [Google Scholar]
- Schulz, E. Actinarctus doryphorus, nov. gen. nov. spec. ein merkwürdiger Tardigrad aus der Nordsee. Zool. Anz. 1935, 111, 285–288. [Google Scholar]
- D’Addabbo Gallo, M.; Pietanza, R.; D’Addabbo, R.; Morone De Lucia, M.R.; Grimaldi de Zio, S. A redescription of Actinarctus doryphorus (Tardigrada, Heterotardigrada). Cah. Biol. Mar. 1999, 40, 21–27. [Google Scholar]
- Grimaldi de Zio, S.; Morone De Lucia, M.R.; D’Addabbo Gallo, M.; Grimaldi, P. Osservazioni su alcuni tardigradi di una spiaggia pugliese e descrizione di Batillipes adriaticus sp. nov. (Heterotardigrada). Thalass. Salentina 1979, 9, 39–50. [Google Scholar]
- D’Addabbo Gallo, M.; Morone De Lucia, M.R.; Grimaldi de Zio, S. Two new species of the genus Styraconyx (Tardigrada: Heterotardigrada). Cah. Biol. Mar. 1989, 30, 17–33. [Google Scholar]
- Bartels, P.J.; Kaczmarek, Ł.; Roszkowska, M.; Nelson, D.R. Interactive Map of Marine Tardigrades of the World. 2015. Available online: https://paul-bartels.shinyapps.io/marine-tardigrades/ (accessed on 19 January 2023).
- Thulin, G. Ein neuer mariner Tardigrad. Medd. Fran Göteborgs Musei Zool. Avd. 1942, 99, 1–10. [Google Scholar]
- Pollock, L.W. On some British marine Tardigrada, including two new species of Batillipes. J. Mar. Biol. Assoc. UK. 1971, 51, 93–103. [Google Scholar] [CrossRef]
- Santos, E.; Rubal, M.; Veiga, P.; Bartels, P.J.; da Rocha, C.M.C.; Fontoura, P. On the distribution of Batillipes tubernatis Pollock, 1971 (Arthrotardigrada: Batillipedidae) in the Atlantic Basin. Mar. Biodivers. 2019, 49, 621–631. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubal, M.; Fontoura, P.; Veiga, P. New Records of Marine Tardigrades (Arthotardigrada) from the Iberian Peninsula: Biogeographical Implications. Diversity 2023, 15, 210. https://doi.org/10.3390/d15020210
Rubal M, Fontoura P, Veiga P. New Records of Marine Tardigrades (Arthotardigrada) from the Iberian Peninsula: Biogeographical Implications. Diversity. 2023; 15(2):210. https://doi.org/10.3390/d15020210
Chicago/Turabian StyleRubal, Marcos, Paulo Fontoura, and Puri Veiga. 2023. "New Records of Marine Tardigrades (Arthotardigrada) from the Iberian Peninsula: Biogeographical Implications" Diversity 15, no. 2: 210. https://doi.org/10.3390/d15020210
APA StyleRubal, M., Fontoura, P., & Veiga, P. (2023). New Records of Marine Tardigrades (Arthotardigrada) from the Iberian Peninsula: Biogeographical Implications. Diversity, 15(2), 210. https://doi.org/10.3390/d15020210