
Two Novel Species and Two New Records within the Genus Pluteus (Agaricomycetes, Agaricales) from Thailand




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Morphological Study
2.2. Molecular Procedures and Phylogenetic Analyses
3. Results
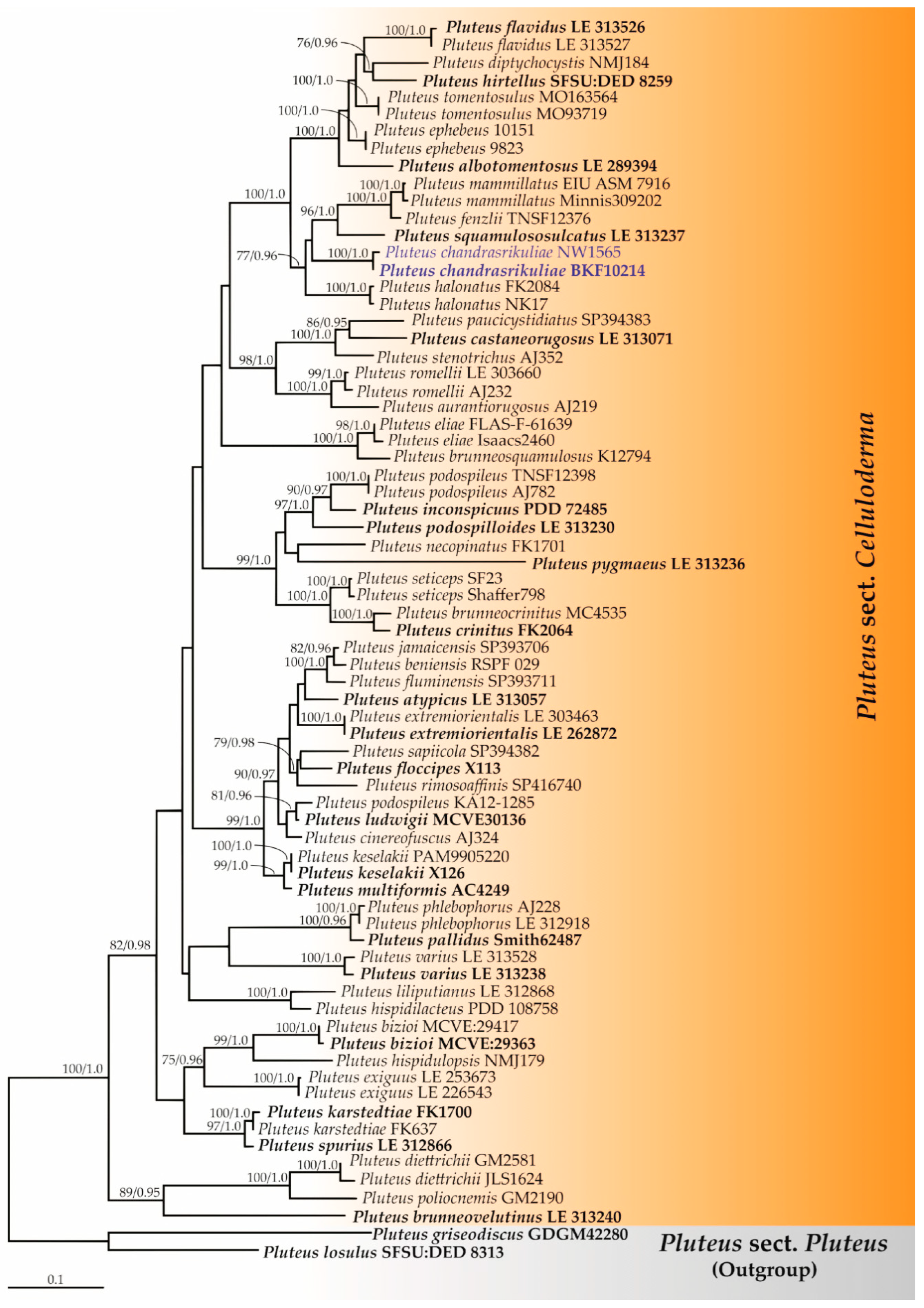
3.1. Phylogenetic Relationships
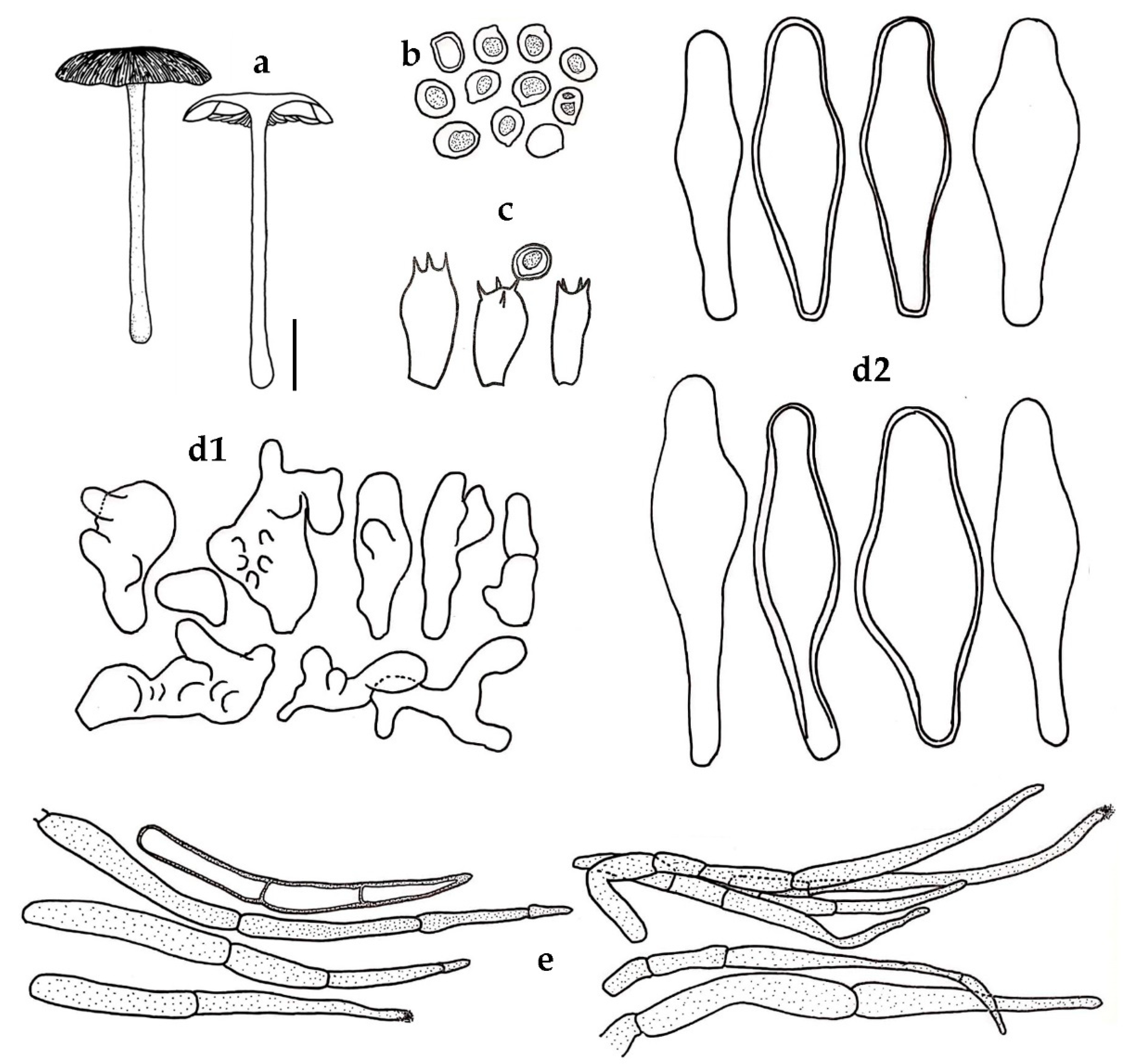
3.2. Taxonomic Description of New Species
3.3. Taxonomic Description of New Record
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, M.Q.; Zhao, R.L.; Hyde, K.D.; Begerow, D.; Kemler, M.; Yurkov, A.; McKenzie, E.H.C.; Raspé, O.; Kakishima, M.; SánchezRamírez, S.; et al. Notes, outline and divergence times of Basidiomycota. Fungal Divers. 2019, 99, 105–367. [Google Scholar]
- Singer, R. The Agaricales in Modern Taxonomy, 4th ed.; Koeltz Scientific Books: Koenigstein, Germany, 1986; pp. 454–459. [Google Scholar]
- Justo, A.; Vizzini, A.; Minnis, A.M.; Menolli, N., Jr.; Capelari, M.; Rodríguez, O.; Malysheva, E.; Contu, M.; Ghinone, S.; Hibbett, D.S. Phylogeny of Pluteaceae (Agaricales, Basidiomycota): Taxonomy and character evolution. Fungal Biol.-UK 2011, 115, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.H. Speciation in higher fungi in relation to modern generic concepts. Mycologia 1968, 60, 742–755. [Google Scholar] [CrossRef]
- Vellinga, E.C. Pluteus . In Flora Agaricina Neerlandica; Bas, C., Kuyper, T.W., Noordeloos, M.E., Vellinga, E.C., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1990; Volume 2, pp. 31–55. [Google Scholar]
- Justo, A.; Minnis, A.M.; Ghignone, S.; Menolli, N.; Capelari, M.; Rodríguez, O.; Malysheva, E.; Contu, M.; Vizzini, A. Species recognition in Pluteus and Volvopluteus (Pluteaceae, Agaricales): Morphology, geography and phylogeny. Mycol. Prog. 2011, 10, 453–479. [Google Scholar] [CrossRef]
- Justo, A.; Malysheva, E.; Bulyonkova, T.; Vellinga, E.C.; Cobian, G.; Nguyen, N.; Minnis, A.M.; Hibbett, D.S. Molecular phylogeny and phylogeography of Holarctic species of Pluteus section Pluteus (Agaricales: Pluteaceae), with description of twelve new species. Phytotaxa 2014, 180, 1–85. [Google Scholar] [CrossRef]
- Vellinga, E.C.; Schreurs, J. Pluteus Fr. in Western Europe. Persoonia 1985, 12, 337–373. [Google Scholar]
- Kaygusuz, O.; Türkekul, I.; Knudsen, H.; Çolak, Ö.F. New records of Pluteus section Hispidoderma in Turkey based on morphological characteristics and molecular data. Phytotaxa 2019, 413, 175–206. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Perry, B.A. The genus Pluteus (Basidiomycota, Agaricales) from Republic of São Tomé and Príncipe, West Africa. Mycosphere 2018, 8, 1317–1391. [Google Scholar] [CrossRef]
- Menolli, N., Jr.; Justo, A.; Arrillaga, P.; Pradeep, C.K.; Minnis, A.M.; Capelari, M. Taxonomy and phylogeny of Pluteus glaucotinctus sensu lato (Agaricales, Basidiomycota), a multicontinental species complex. Phytotaxa 2014, 188, 78–90. [Google Scholar] [CrossRef]
- Malysheva, E.F.; Malysheva, V.F.; Justo, A. Observation on Pluteus (Pluteaceae) diversity in South Siberia, Russia: Morphological and molecular data. Mycol. Prog. 2016, 15, 861–882. [Google Scholar] [CrossRef]
- Pegler, D.N. A preliminary Agaric Flora of East Africa; Kew Bulletin Additional Series 6; Her Majesty’s Stationery Office: Richmond, UK, 1977; pp. 1–615. [Google Scholar]
- Ferisin1, G.; Dovana, F.; Justo, A. Pluteus bizioi (Agaricales, Pluteaceae), a new species from Italy. Phytotaxa 2019, 408, 099–108. [Google Scholar] [CrossRef]
- Kaygusuz, O.; Gezer, K.; Şeker, M. Four new records of Pluteus Fr. from interesting habitats in the Aegean region of Turkey. Bot. Lett. 2016, 163, 251–259. [Google Scholar] [CrossRef]
- Kaygusuz, O.; Knudsen, H.; Menolli, N., Jr.; Türkekul, I. Pluteus anatolicus (Pluteaceae, Agaricales): A new species of Pluteus sect. Celluloderma from Turkey based on both morphological and molecular evidence. Phytotaxa 2021, 482, 240–250. [Google Scholar] [CrossRef]
- Malysheva, E.F.; Morozova, O.V.; Zvyagina, E.A. New records of the annulate Pluteus in European and Asian Russia. Acta Mycol. 2007, 42, 153–160. [Google Scholar] [CrossRef]
- Ševčíková, H.; Malysheva, E.F.; Justo, A.; Heilmamm-Clausen, J.; Tomšovský, M. Pluteus dianae and P. punctatus resurrected, with first records from eastern and northern Europe. Mycotaxon 2020, 135, 245–274. [Google Scholar] [CrossRef]
- Vizzini, A.; Ercole, E. A new annulate Pluteus variety from Italy. Mycologia 2011, 103, 904–911. [Google Scholar] [CrossRef][Green Version]
- Radu, M.-I.; Şesan, T.-E. Pluteus aurantiorugosus (Trog) Sacc. and Coprinus patouillardii Quél.—New records of macrofungi for Romania. Acta Mycol. 2013, 48, 189–196. [Google Scholar] [CrossRef]
- Orton, P.D. Bristish Fungus Flora Agarics and Boleti 4—Pluteaceae: Pluteus & Volvariella; Royal Botanic Garden: Edinburgh, UK, 1986; pp. 1–99. [Google Scholar]
- Orton, P.D. New check list of British Agarics and Boleti, part III (Note on genera and species in the list). Trans. Brit. Mycol. Soc. 1960, 43, 159–439. [Google Scholar] [CrossRef]
- Citérin, M.; Eyssartier, G. Clé analytique du genre Pluteus Fr. Doc. Mycol. 1998, 28, 47–67. [Google Scholar]
- Kaygusuz, O.; Justo, A.; Knudsen, H.; Ševčíková, H.; Türkekul, I. Pluteus lauracearum (Agaricales, Basidiomycota), a new species of Pluteus sect. Hispidoderma from thermophilic Laurus forests. Phytotaxa 2021, 523, 126–140. [Google Scholar] [CrossRef]
- Kaygusuz, O.; Turkekul, I.; Knudsen, H.; Menolli, N., Jr. Volvopluteus and Pluteus section Pluteus (Agaricales: Pluteaceae) in Turkey based on morphological and molecular data. Turk. J. Bot. 2021, 45, 224–242. [Google Scholar] [CrossRef]
- Ševčíková, H.; Bororvička, J. Pluteus rugosidiscus (Basidiomycota, Pluteaceae), first record of this North American species in Europe. Nova Hedwigia 2019, 108, 227–241. [Google Scholar] [CrossRef]
- Bonnard, J. Pluteus lidipocystis sp. nov. Mycol. Helv. 1986, 2, 35–42. [Google Scholar]
- Bonnard, J. Pluteus brunneoradiatus sp. nov. Mycol. Helv. 1987, 2, 141–154. [Google Scholar]
- Bonnard, J. Les cystides de la section Pluteus (Agaricales). Mycol. Helv. 1988, 3, 53–72. [Google Scholar]
- Bonnard, J. Pluteus primus sp. nov. (Agaricales, Basidiomycetes). Mycol. Helv. 1991, 4, 169–178. [Google Scholar]
- Bonnard, J. Clé provisoire des Plutées européens à boucles. Mycol. Helv. 1993, 6, 203–205. [Google Scholar]
- Bonnard, J. Pluteus pellitus désignation d’un néotype (Section Pluteus, Agaricales, Basidiomycetes). Mycol. Helv. 1995, 7, 97–103. [Google Scholar]
- Bonnard, J. Pluteus albineus sp. nov. (Agaricales, Basidiomycetes). Mycol. Helv. 2001, 11, 131–136. [Google Scholar]
- Justo, A.; Castro, M.L. Observations in Pluteus section Pluteus in Spain: Two new records for Europe. Mycotaxon 2007, 102, 209–220. [Google Scholar]
- Justo, A.; Castro, M.L. An annotated checklist of Pluteus in the Iberian Peninsula and Balearic Islands. Mycotaxon 2007, 102, 231–234. [Google Scholar]
- Campi, M.G.; Maubet, Y.; Cristaldo, E.; Grassi, E.; Menolli, N., Jr. Pluteus Fr. (Pluteaceae, Agaricales) in Paraguay: Morphological studies and new records. Check List 2019, 15, 313–322. [Google Scholar] [CrossRef]
- Justo, A.; Battistin, E.; Angelini, C. Two new species of Pluteus section Celluloderma from the Dominican Republic. Mycotaxon 2012, 120, 11–21. [Google Scholar] [CrossRef]
- Homola, R.L. Section Celluloderma of the Genus Pluteus in North America. Mycologia 1972, 64, 1211–1257. [Google Scholar] [CrossRef]
- Menolli, N., Jr.; Asai, T.; Capelari, M. Records and new species of Pluteus from Brazil based on morphological and moleculardata. Mycology 2010, 1, 130–153. [Google Scholar] [CrossRef]
- Menolli, N., Jr.; Justo, A.; Capelari, M. Phylogeny of Pluteus section Celluloderma including eight new species from Brazil. Mycologia 2015, 107, 1205–1220. [Google Scholar] [CrossRef]
- Menolli, N., Jr.; Justo, A.; Capelari, M. Pluteus section Hispidoderma in Brazil with new records based on morphological and molecular Data. Cryptogam. Mycol. 2015, 36, 331–354. [Google Scholar] [CrossRef]
- Minnis, A.M.; Sundberg, W.J. Pluteus section Celluloderma in the U.S.A. N. Am. Fungi 2010, 5, 1–107. [Google Scholar] [CrossRef][Green Version]
- Murrill, W.A. The Agaricaceae of Tropical North America—IV. Mycologia 1911, 3, 271–282. [Google Scholar]
- Minnis, A.M.; Sundberg, W.J.; Methven, A.S.; Sipes, S.D.; Nickrent, D.L. Annulate Pluteus species: A study of the genus Chamaeota in the United States. Mycotaxon 2006, 96, 31–39. [Google Scholar]
- Brandrud, T.E.; Krisai-Greilhuber, I.; Kunca, V. Pluteus fenzlii . In The IUCN Red List of Threatened Species 2019; GBIF: Copenhagen, Denmark, 2019; pp. 1–8. [Google Scholar]
- Leonard, P.L. Pluteus bressollensis . In The IUCN Red List of Threatened Species 2019; GBIF: Copenhagen, Denmark, 2019; pp. 1–6. [Google Scholar]
- Hosen, M.I.; Liang, X.; Xu, J.; Li, T.H. Pluteus squarrosus sp. nov. (Pluteus sect. Celluloderma, Pluteaceae) from northeast China. Nord. J. Bot. 2019, 37, e02427. [Google Scholar] [CrossRef]
- Xu, J.; Li, T.H.; Justo, A.; Ge, Z.W. Two new species of Pluteus (Agaricales, Pluteaceae) from China. Phtyotaxa 2015, 233, 61–68. [Google Scholar] [CrossRef][Green Version]
- Hosaka1, K.; Kobayashi, T.; Castellano, M.A.; Orihara, T. The status of voucher specimens of mushroom species thought to be extinct from Japan. Bull. Natl. Mus. Nat. Sci. Ser. B 2018, 44, 53–66. [Google Scholar]
- Lezzi, T.; Vizzin, A.; Ercole, E.; Magliozzi, V.; Justo, A. Phylogenetic and morphological comparison of Pluteus variabilicolor and P. castri (Basidiomycota, Agaricales). IMA Fungus 2014, 5, 415–423. [Google Scholar] [CrossRef]
- Takahashi, H. Notes on new Agaricales of Japan 2. Mycoscience 2001, 42, 347–353. [Google Scholar] [CrossRef]
- Takahashi, H. Pluteus romellii (Agaricales, Basidiomycetes), new to Japan, found in Odawara (in Japanese with English summary). Nat Hist. Rep. Kanagawa 2001, 22, 21–23. [Google Scholar]
- Takehashi, S.; Kasuya, T. First record of Pluteus chrysophaeus and reexamination of Pluteus leoninus from Japan. Mycoscience 2007, 48, 321–325. [Google Scholar] [CrossRef]
- Lee, J.N.; Lee, H.K.; Min, K.H.; Park, W.H.; Kim, Y.S. Studies on genus Pluteus of Korea. Korean Mycol. 1992, 20, 296–301. [Google Scholar]
- Cho, S.; Eun, K.Y.; Jo, J.; Won, H.S.; Oh, S.H.; Kim, C.S. Macrofungal diversity of urbanized areas in southern part of Korea. J. Asia Pac. Biodivers. 2020, 13, 189–197. [Google Scholar] [CrossRef]
- Park, M.S.; Cho, H.J.; Kim, N.K.; Park, J.Y.; Lee, H.; Park, K.H.; Kim, M.J.; Kim, J.J.; Kim, C.; Lim, Y.W. Ten new recorded species of macrofungi on Ulleung Island, Korea. Mycobiology 2017, 45, 286–296. [Google Scholar] [CrossRef]
- Lee, T.S. The full list of recorded mushrooms in Korea. Kor. J. Mycol. 1990, 18, 233–259. [Google Scholar]
- Ševčíková, H.; Antonín, V.; Borovička, J. Pluteus septocystidiatus, a new species with unique pleurocystidia. Sydowia 2014, 66, 229–239. [Google Scholar]
- Pegler, D.N. Agaric Flora of Sri Lanka; Kew Bulletin Additional Series 12; Her Majesty’s Stationery Office: Richmond, UK, 1986; pp. 1–519. [Google Scholar]
- Berkeley, M.J.; Broome, C.E. The fungi of Ceylon. J. Linn. Soc., Bot. 1871, 11, 494–567. [Google Scholar] [CrossRef]
- Petch, T. Ceylon pink-spored agarics. Roy. Bot. Gard. (Peradeniya) 1924, 9, 201–216. [Google Scholar]
- Pradeep, C.K.; Vrinda, K.B. New and noteworthy species of Pluteus (Pluteaceae, Agaricales) from Kerala State, India. Persoonia 2006, 19, 95–99. [Google Scholar]
- Pradeep, C.K.; Justo, A.; Vrinda, K.B.; Shibu, V.P. Two new species of Pluteus (Pluteaceae, Agaricales) from India and additional observations on Pluteus chrysaegis. Mycol. Prog. 2012, 11, 869–878. [Google Scholar] [CrossRef]
- Malysheva, E.; Malysheva, V.; Alexandrova, A.; Morozova, O. Observations on Pluteaceae in Vietnam. 2. One new record and ten new species of Pluteus. Phytotaxa 2020, 461, 79–107. [Google Scholar] [CrossRef]
- Chandrasrikul, A.; Suwanarit, P.; Sangwanit, U.; Lunyong, S.; Payapanon, A.; Sanoamuang, N.; Pukahuta, C.; Petcharat, V.; Sardsud, U.; Duengkae, K.; et al. Checklist of Mushrooms (Basidiomycetes) in Thailand; Office of Natural Resources and Environmental Policy and Planning: Bangkok, Thailand, 2011; pp. 112–113. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978; pp. 1–252. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Stamatakis, A. Raxml-vi-hpc: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Hoehna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree Tree Figure Drawing Tool Version 131, Institute of Evolutionary 623 Biology, University of Edinburgh. Available online: http://treebioedacuk/software/figtree/ (accessed on 25 December 2021).
- Menolli, N., Jr.; Meijer, A.A.R.; Capelari, M. The genus Pluteus (Pluteaceae, Agaricales) from the state of Paraná, Brazil. Nova Hedwigia 2015, 100, 101–157. [Google Scholar] [CrossRef]
- Takahashi, H.; Kasuya, T. Type study of Pluteus atrofuscens (Agaricales, Pluteaeceae). Mycoscience 2010, 51, 81–83. [Google Scholar] [CrossRef]
- Singer, R. Contributions towards a monograph of the genus Pluteus. Trans. Brit. Mycol. Soc. 1956, 39, 145–232. [Google Scholar] [CrossRef]
- Imazeki, R.; Otani, Y.; Hongo, T. Fungi of Japan, 2nd ed.; Yama-Kei Publishers: Tokyo, Japan, 1989; pp. 179–180. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wannathes, N.; Suwannarach, N.; Khuna, S.; Kumla, J. Two Novel Species and Two New Records within the Genus Pluteus (Agaricomycetes, Agaricales) from Thailand. Diversity 2022, 14, 156. https://doi.org/10.3390/d14030156
Wannathes N, Suwannarach N, Khuna S, Kumla J. Two Novel Species and Two New Records within the Genus Pluteus (Agaricomycetes, Agaricales) from Thailand. Diversity. 2022; 14(3):156. https://doi.org/10.3390/d14030156
Chicago/Turabian StyleWannathes, Nopparat, Nakarin Suwannarach, Surapong Khuna, and Jaturong Kumla. 2022. "Two Novel Species and Two New Records within the Genus Pluteus (Agaricomycetes, Agaricales) from Thailand" Diversity 14, no. 3: 156. https://doi.org/10.3390/d14030156
APA StyleWannathes, N., Suwannarach, N., Khuna, S., & Kumla, J. (2022). Two Novel Species and Two New Records within the Genus Pluteus (Agaricomycetes, Agaricales) from Thailand. Diversity, 14(3), 156. https://doi.org/10.3390/d14030156