
Integrative Taxonomy of Two Peruvian Strains of Brachionus plicatilis Complex with Potential in Aquaculture

Abstract

1. Introduction
2. Material and Methods
2.1. Sample Collection
2.2. Culture Conditions, Morphometry, and Parameter Evaluation
2.3. Molecular Analysis
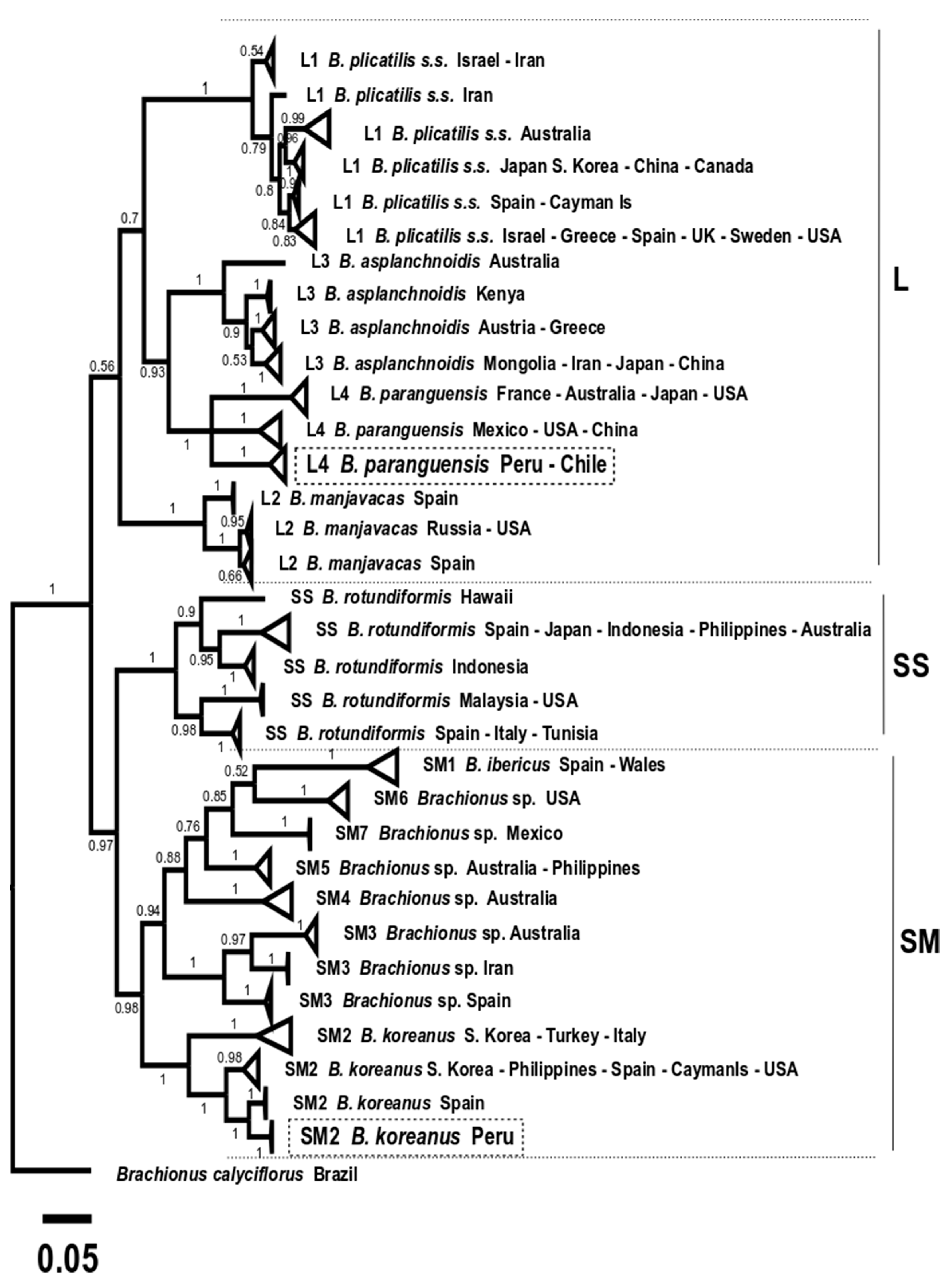
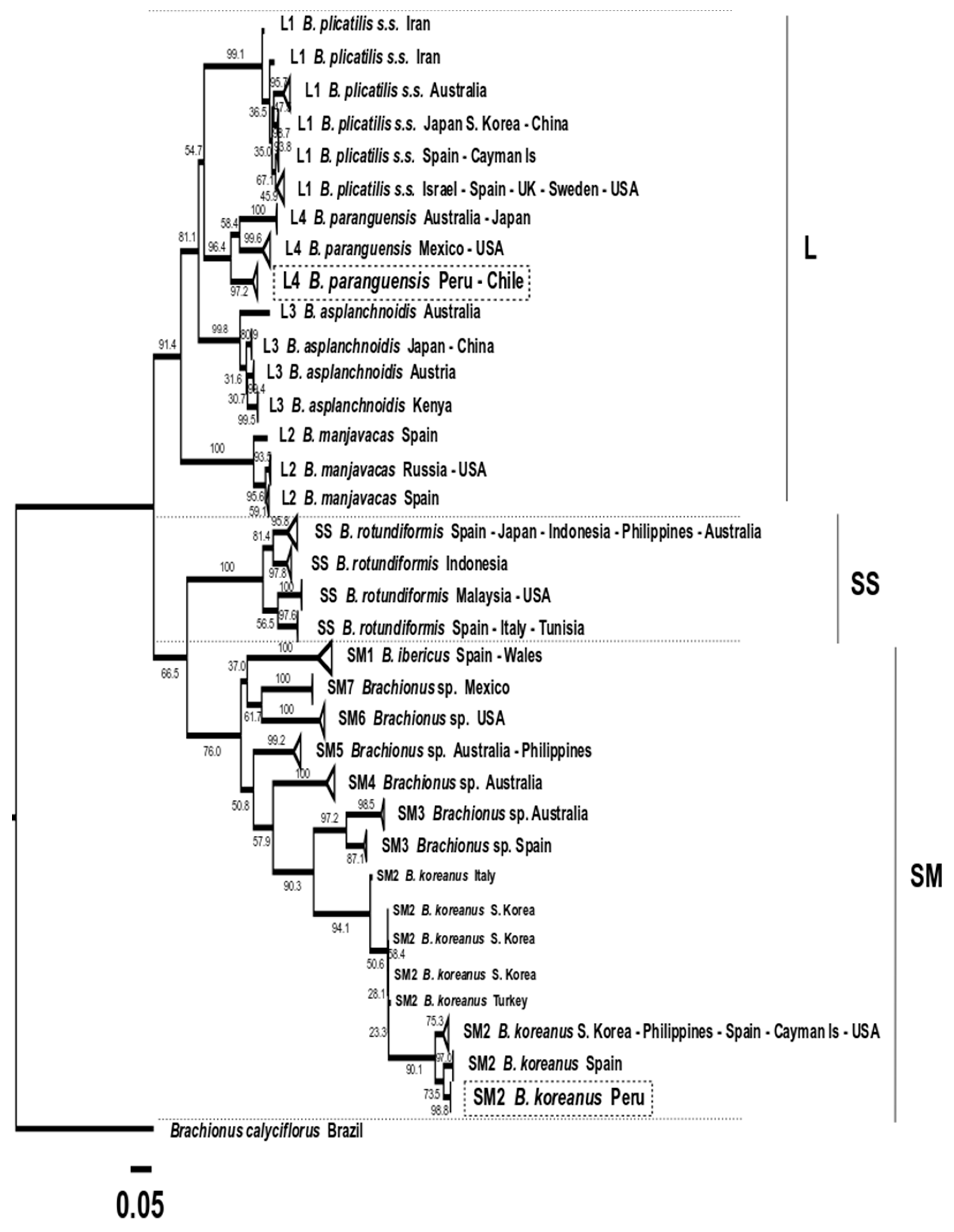
3. Results
3.1. Taxonomy
- Strain IMP-BG Z010-VL
- Class Eurotatoria De Ridder, 1957
- Subclass Monogononta Plate, 1889
- Superorder Pseudotrocha Kutikova, 1970
- Order Ploima Hudson & Gosse, 1886
- Family Brachionidae Ehrenberg, 1838
- Genus Brachionus Pallas, 1766
- Species Brachionus paranguensis Guerrero-Jiménez, 2019
- Sub species Brachionus paranguensis ventanillensis subsp. nov.
- Strain IMP-BG Z018-SD
- Class Eurotatoria De Ridder, 1957
- Subclass Monogononta Plate, 1889
- Superorder Pseudotrocha Kutikova, 1970
- Order Ploima Hudson & Gosse, 1886
- Family Brachionidae Ehrenberg, 1838
- Genus Brachionus Pallas, 1766
- Species Brachionus koreanus Hwang, 2013
- Sub species Brachionus koreanus santodomingensis subsp. nov.
3.2. Etymology
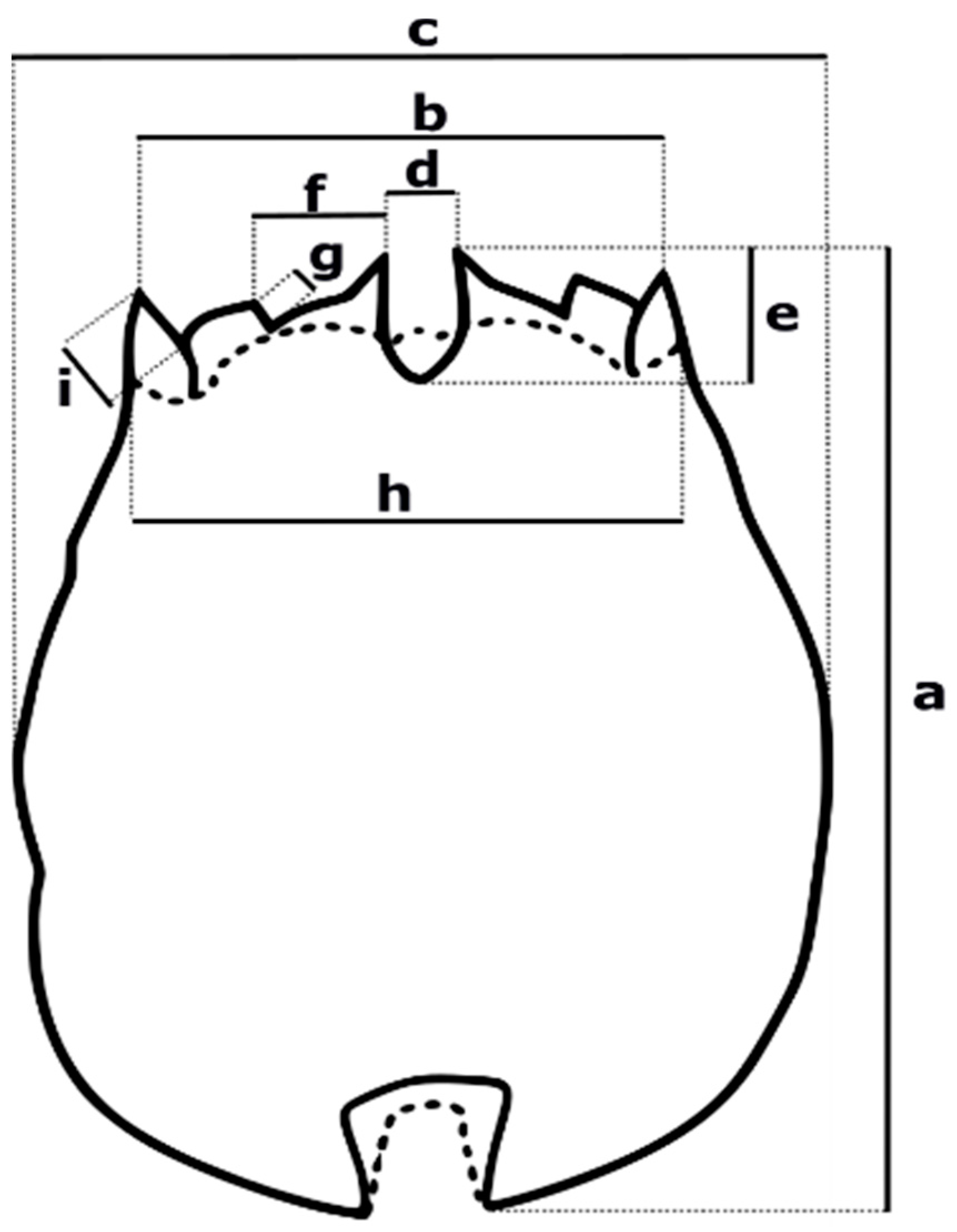
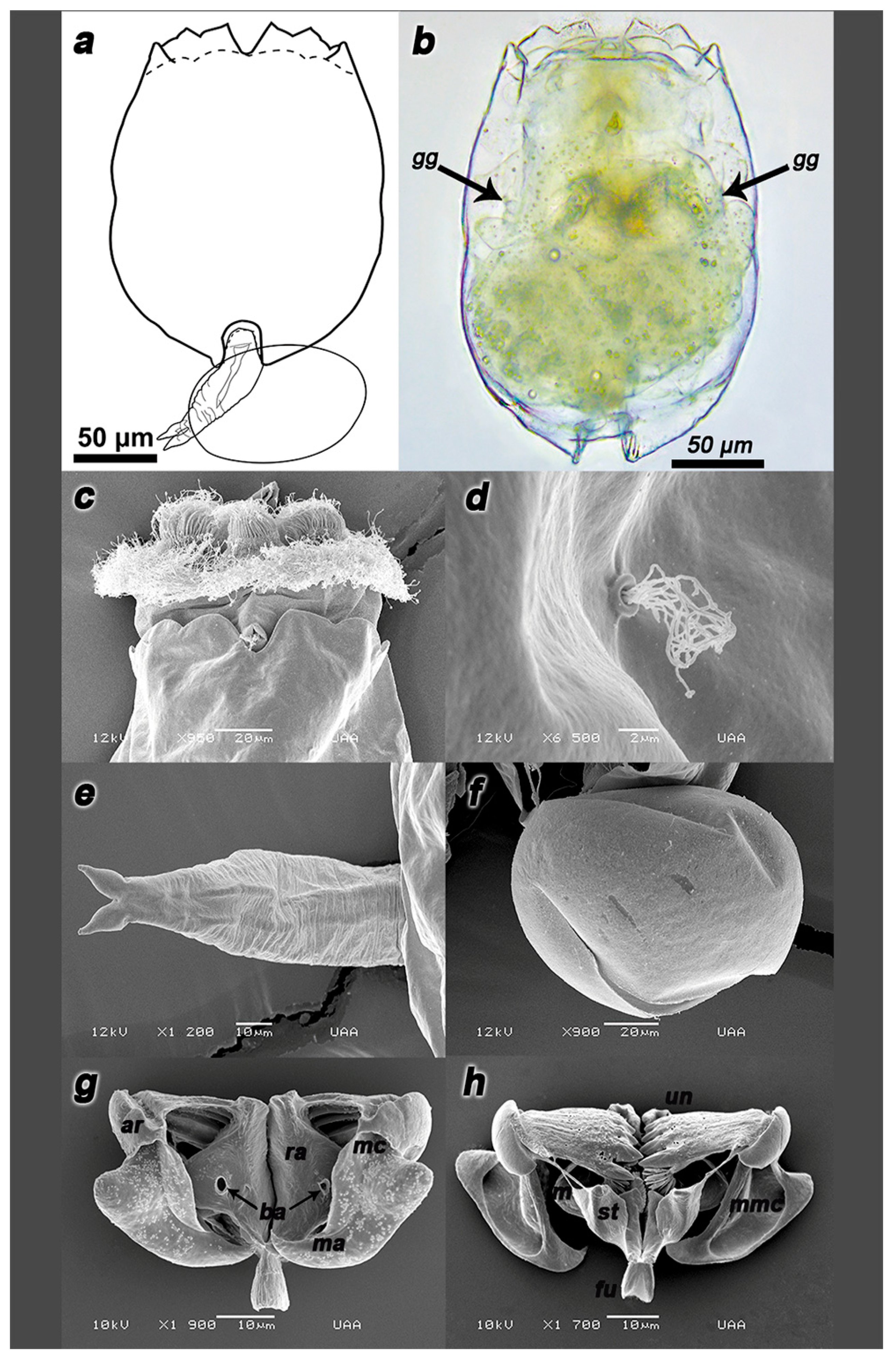
3.3. Morphological Differences of Peruvian Strains
3.3.1. Strain IMP-BG Z010-VL
Description
Type Locality
Differential Diagnosis
Trophi
3.3.2. Strain IMP-BG Z018-SD
Description
Type Locality
Differential Diagnosis
Trophi
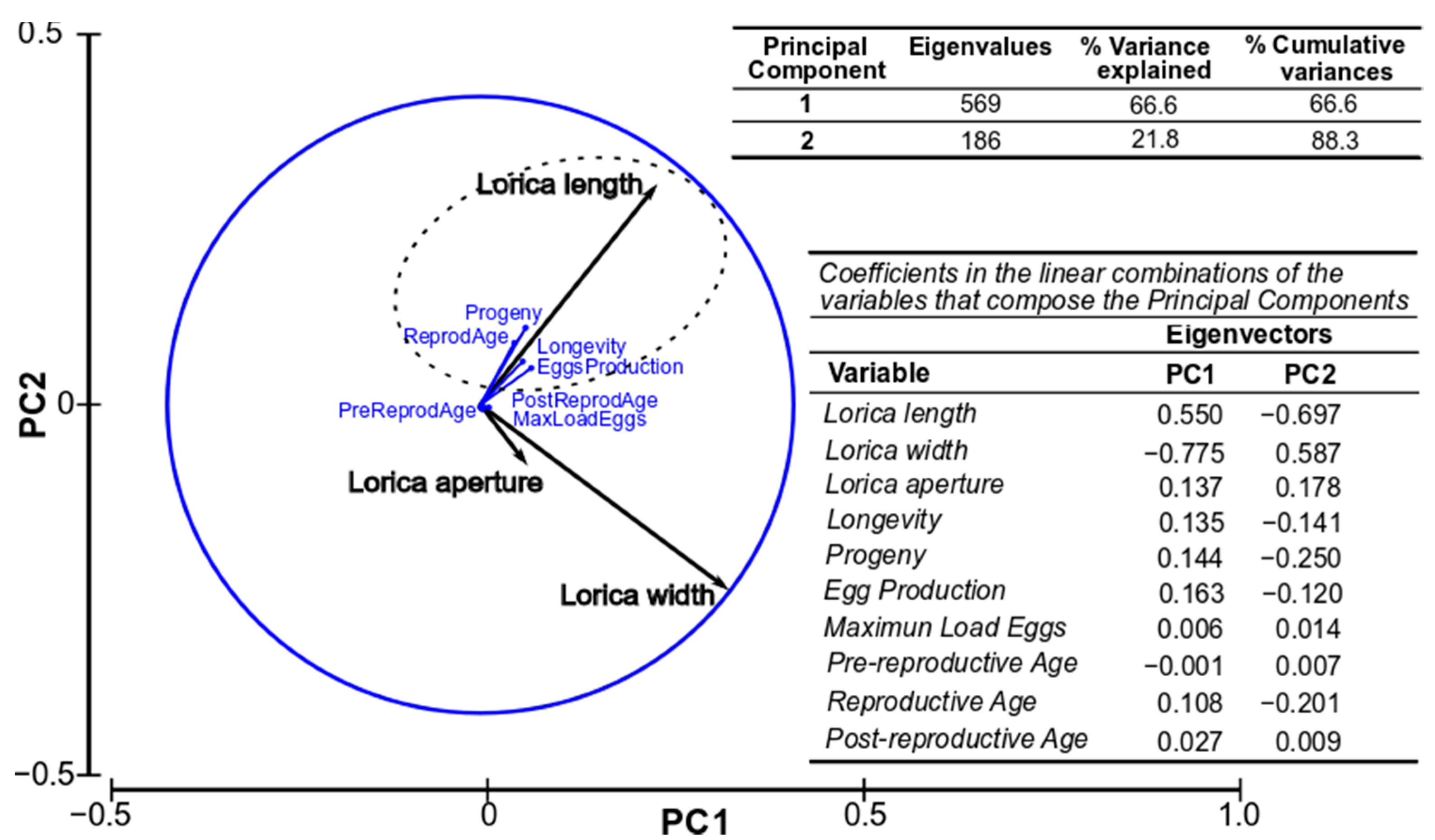
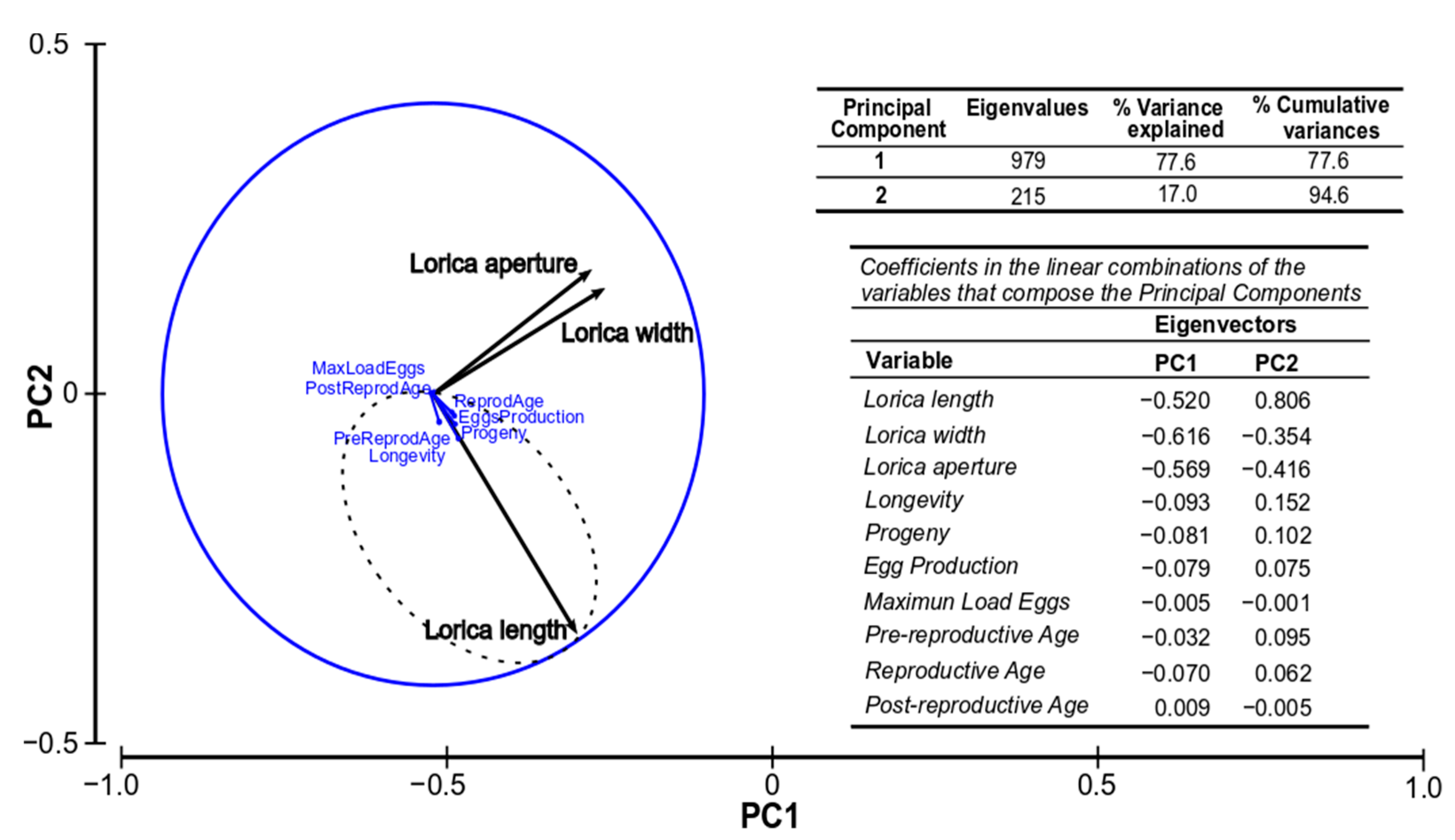
3.4. Relationship between Biometric and Production Parameters
3.5. Canonical Discriminant Analysis
3.6. Molecular Taxonomy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jennings, H.S. Rotatoria of the United States, with especial reference to those of the great lakes. Bull. US Fish Comm. 1900, 19, 14–22. [Google Scholar]
- Harring, H.K.; Myers, F.J. The rotifer fauna of Wisconsin. IV. The Dicranophorinae. Trans. Wis. Acad. Sci. Arts Lett. 1928, 23, 667–808. [Google Scholar]
- Serra, M.; Miracle, M.R. Biometric analysis of Brachionus plicatilis ecotypes from Spanish lagoons. Hydrobiologia 1983, 104, 279–291. [Google Scholar] [CrossRef]
- Mills, S.; Alcántara-Rodríguez, J.A.; Ciros-Pérez, J.; Gómez, A.; Hagiwara, A.; Galindo, K.H.; Jersabek, C.D.; Malekzadeh-Viayeh, R.; Leasi, F.; Lee, J.-S.; et al. Fifteen species in one: Deciphering the Brachionus plicatilis species complex (Rotifera, Monogononta) through DNA taxonomy. Hydrobiologia 2017, 796, 39–58. [Google Scholar] [CrossRef]
- Guerrero-Jiménez, G.; Vannucchi, P.E.; Silva-Briano, M.; Ortiz, A.A.; Rico-Martínez, R.; Roberts, D.; Neilson, R.; Elías-Gutiérrez, M. Brachionus paranguensis sp. nov. (Rotifera, Monogononta), a member of the L group of the Brachionus plicatilis complex. ZooKeys 2019, 880, 1–23. [Google Scholar] [CrossRef]
- Romero, G.L. Caracterización Morfométrica y Aspectos Filogenéticos de Cepas de Rotíferos del Grupo Brachionus plicatilis (Rotifera: Brachionidae) Utilizados en la Acuicultura Peruana. Master’s Dissertation, Universidad Nacional Mayor de San Marcos, Lima, Peru, 2008. [Google Scholar]
- Toscano, E.; Severino, R. Brachionidae (Rotifera: Monogononta) de la albufera El Paraíso y el reporte de Brachionus ibericus en el Perú. Rev. Peru. Biol. 2013, 20, 177–180. [Google Scholar] [CrossRef]
- Chinchayán, M. Cultivo de la Microalga Nannochloropsis oculata y su Consumo por el Rotífero Brachionus plicalitis (línea S). Master’s Dissertation, Universidad Nacional Agraria La Molina, Lima, Peru, 1996. [Google Scholar]
- Alayo, M.; Iannacone, J. Population growth of the rotífer eurihalin Brachionus plicatilis hepatotomus fed on the microalga nannochloris sp. Bol. Lima 2001, 23, 87–93. [Google Scholar]
- Alayo, M.; Iannacone, J. Ensayos ecotoxicológicos con petróleo crudo, diesel 2 y diesel 6 con dos subespecies de Brachionus plicatilis müller 1786 (rotifera: Monogononta). Gayana 2002, 66, 45–58. [Google Scholar] [CrossRef]
- Cisneros, R. Rendimiento poblacional del rotífero nativo Brachionus sp. “Cayman”, utilizando diferentes enriquecedores. Ecol. Apl. 2011, 10, 99–105. [Google Scholar] [CrossRef][Green Version]
- Rosales-Barrantes, R. Efecto de la Temperatura, la Salinidad y sus Interacciones Sobre el Crecimiento Poblacional del Rotífero Nativo Brachionus sp. Cayman, Cepa Chilca, Perú. Master’s Dissertation, Universidad Ricardo Palma, Lima, Peru, 2012. [Google Scholar]
- Alcedo-Durán, N.; Córdova-Calle, J.C. Efecto de Tres Concentraciones de Harina de Ensilado de Vísceras de Gallus gallus Domesticus “pollo” en el Crecimiento Poblacional y Contenido de Proteínas y Lípidos de Brachionus sp. en Laboratorio. Master’s Dissertation, Universidad Nacional del Santa, Nuevo Chimbote, Peru, 2014. [Google Scholar]
- Murrieta-Morey, G.A.; Nájar, J.; Alcantara-Bocanegra, F. Producción experimental de rotíferos en bolsas de plástico utilizando harina de pescado como fuente de nutrientes. Folia Amaz. 2015, 24, 15. [Google Scholar] [CrossRef]
- Huanacuni-Pilco, J.I.; Espinoza-Ramos, L.A. Producción de alimento vivo para la investigación en acuicultura de peces marinos en la UNJBG, Tacna. Cienc. Desarro. 2019, 17, 82–86. [Google Scholar] [CrossRef]
- Aguilar-Samanamud, C.P.; Gaspar, W.; Inga, G.; Flores, L.; Sanchez, P.; Hernandez-Acevedo, H.; Borda-Soares, R.; Olivera-Galvez, A. Variables that intervene in the weight of rotifer biomass and fatty acids. J. World Aquac. Soc. 2021, 1–13. [Google Scholar] [CrossRef]
- Lubzens, E. Raising rotifers for use in aquaculture. In Rotifer Symposium IV; May, L., Wallace, R., Herzig, A., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 245–255. ISBN 9789401083027/9789400940598. [Google Scholar]
- Andersen, R.A. Algal Culturing Techniques; Elsevier: Amsterdam, The Netherlands, 2005; p. 589. [Google Scholar]
- Segers, H.; Murugan, G.; Dumont, H.J. On the taxonomy of the Brachionidae: Description of Plationus n. gen. (Rotifera, Monogononta). Hydrobiologia 1993, 268, 1–8. [Google Scholar] [CrossRef]
- Silva-Briano, M.; Adabache-Ortiz, A.; Guerrero-Jiménez, G.; Rico-Martínez, R.; Zavala-Padilla, G. Ultrastructural and Morphological Description of the Three Major Groups of Freshwater Zooplankton (Rotifera, Cladocera, and Copepoda) from the State of Aguascalientes, Mexico. In The Transmission Electron Microscope: Theory and Applications; INTECH Open Science/Open Minds: Rijeka, Croatia, 2015; pp. 307–325. [Google Scholar]
- Fu, Y.; Hirayama, K.; Natsukari, Y. Morphological differences between two types of the rotifer Brachionus plicatilis O.F. Müller. J. Exp. Mar. Biol. Ecol. 1991, 151, 29–41. [Google Scholar] [CrossRef]
- Ciros-Pérez, J.; Gómez, A.; Serra, M. On the taxonomy of three sympatric sibling species of the Brachionus plicatilis (Rotifera) complex from Spain, with the description of B. ibericus n. sp. J. Plankton Res. 2001, 23, 1311–1328. [Google Scholar] [CrossRef]
- Montero-Pau, J.; Gomez, A.; Munoz, J. Application of an inexpensive and high-throughput genomic DNA extraction method for the molecular ecology of zooplanktonic diapausing eggs. Limnol. Oceanogr. Methods ASLO 2008, 6, 218–222. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Prosser, S.; Martínez-Arce, A.; Elías-Gutiérrez, M. A new set of primers for COI amplification from freshwater microcrustaceans. Mol. Ecol. Resour. 2013, 13, 1151–1155. [Google Scholar] [CrossRef]
- Messing, J. New M13 vectors for cloning. In Methods in Enzymology; Recombinant DNA Part C; Academic Press: Cambridge, MA, USA, 1983; Volume 101, pp. 20–78. [Google Scholar]
- Palumbi, S.R. Nucleic acids II: The polymerase chain reaction. In Molecular Systematics, 2nd ed.; Hillis, D., Moritz, C., Mable, B., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 205–247. ISBN 0878932828 9780878932825. [Google Scholar]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New Algorithms and Methods to Estimate Maximum-Likelihood Phylogenies: Assessing the Performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.-S.; Dahms, H.-U.; Park, H.G.; Lee, J.-S. A new intertidal Brachionus and intrageneric phylogenetic relationships among Brachionus as revealed by allometry and CO1-ITS1 gene analysis. Zool. Stud. 2013, 52, 13. [Google Scholar] [CrossRef]
- Campaña-Torres, A.; Martinez-Cordova, L.R.; Villarreal-Colmenares, H.; Hernández-López, J.; Ezquerra-Brauer, J.M.; Cortés-Jacinto, E. Efecto de la adición del rotífero Brachionus rotundiformis (Tschugunoff, 1921) sobre la calidad del agua y la producción, en cultivos super-intensivos de camarón blanco del Pacífico Litopenaeus vannamei (Boone, 1931). Rev. Biol. Mar. Oceanogr. 2009, 44, 335–342. [Google Scholar] [CrossRef]
- Dumont, H.J. Biogeography of rotifers. In Proceedings of the Biology of Rotifers; Pejler, B., Starkweather, R., Nogrady, T., Eds.; Springer: Dordrecht, The Netherlands, 1983; pp. 19–30. [Google Scholar]
- Castellanos Páez, M.E.; Garza Mouriño, G.; Marañón Herrera, S. Aislamiento, Caracterización, Biología y Cultivo del Rotífero Brachionus plicatilis (O.F. Muller); Universidad Autónoma Metropolitana, Unidad Xochimilco: Mexico City, Mexico, 1999. [Google Scholar]
- Gribble, K.; Welch, D.B.M. Life-Span Extension by Caloric Restriction Is Determined by Type and Level of Food Reduction and by Reproductive Mode in Brachionus manjavacas (Rotifera). J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2013, 68, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Fontaneto, D.; Kaya, M.; Herniou, E.A.; Barraclough, T.G. Extreme levels of hidden diversity in microscopic animals (Rotifera) revealed by DNA taxonomy. Mol. Phylogenet. Evol. 2009, 53, 182–189. [Google Scholar] [CrossRef]
- Malekzadeh-Viayeh, R.; Pak-Tarmani, R.; Rostamkhani, N.; Fontaneto, D. Diversity of the rotifer Brachionus plicatilis species complex (Rotifera: Monogononta) in Iran through integrative taxonomy. Zool. J. Linn. Soc. 2014, 170, 233–244. [Google Scholar] [CrossRef]
- García-Morales, A.E.; Elías-Gutiérrez, M. DNA barcoding of freshwater Rotifera in Mexico: Evidence of cryptic speciation in common rotifers. Mol. Ecol. Resour. 2013, 13, 1097–1107. [Google Scholar] [CrossRef]
- Obertegger, U.; Cieplinski, A.; Fontaneto, D.; Papakostas, S. Mitonuclear discordance as a confounding factor in the DNA taxonomy of monogonont rotifers. Zool. Scr. 2018, 47, 122–132. [Google Scholar] [CrossRef]
- Rokas, A.; Williams, B.L.; King, N.; Carroll, S.B. Genome-scale approaches to resolving incongruence in molecular phylogenies. Nature 2003, 425, 798–804. [Google Scholar] [CrossRef]
- Toews, D.P.L.; Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907–3930. [Google Scholar] [CrossRef]
- Després, L. One, two or more species? Mitonuclear discordance and species delimitation. Mol. Ecol. 2019, 28, 3845–3847. [Google Scholar] [CrossRef]
- Snell, T.W.; Shearer, T.L.; Smith, H.A.; Kubanek, J.; Gribble, K.E.; Welch, D.B.M. Genetic determinants of mate recognition in Brachionus manjavacas (Rotifera). BMC Biol. 2009, 7, 60. [Google Scholar] [CrossRef]
- Wendel, J.F.; Doyle, J.J. Phylogenetic Incongruence: Window into Genome History and Molecular Evolution. In Molecular Systematics of Plants II: DNA Sequencing; Soltis, D.E., Soltis, P.S., Doyle, J.J., Eds.; Springer: Boston, MA, USA, 1998; pp. 265–296. [Google Scholar]
- Álvarez, I.; Wendel, J.F. Ribosomal ITS sequences and plant phylogenetic inference. Mol. Phylogenet. Evol. 2003, 29, 417–434. [Google Scholar] [CrossRef]
- Aránguiz-Acuña, A.; Pérez-Portilla, P.; De La Fuente, A.; Fontaneto, D. Life-history strategies in zooplankton promote coexistence of competitors in extreme environments with high metal content. Sci. Rep. 2018, 8, 11060. [Google Scholar] [CrossRef]
- Suatoni, E.; Vicario, S.; Rice, S.; Snell, T.; Caccone, A. An analysis of species boundaries and biogeographic patterns in a cryptic species complex: The rotifer—Brachionus plicatilis. Mol. Phylogenet. Evol. 2006, 41, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Stelzer, C.-P.; Riss, S.; Stadler, P. Genome size evolution at the speciation level: The cryptic species complex Brachionus plicatilis (Rotifera). BMC Evol. Biol. 2011, 11, 90. [Google Scholar] [CrossRef] [PubMed]
- Michaloudi, E.; Mills, S.; Papakostas, S.; Stelzer, C.-P.; Triantafyllidis, A.; Kappas, I.; Vasileiadou, K.; Proios, K.; Abatzopoulos, T.J. Morphological and taxonomic demarcation of Brachionus asplanchnoidis Charin within the Brachionus plicatilis cryptic species complex (Rotifera, Monogononta). Hydrobiologia 2017, 796, 19–37. [Google Scholar] [CrossRef]
- Yoshinaga, T.; Minegishi, Y.; Rumengan, I.; Kaneko, G.; Furukawa, S.; Yanagawa, Y.; Tsukamoto, K.; Watabe, S. Molecular phylogeny of the rotifers with two Indonesian Brachionus lineages. Coast. Mar. Sci. 2004, 29, 45–56. [Google Scholar]
- Yasuike, M.; Tezuka, N.; Sekino, M.; Katoh, M. Species identification of the four rotifer genetic resources preserved at the FRA Genebank Project based on ITS1 and COI sequences. Unpublished work. 2017. [Google Scholar]
- Papakostas, S.; Dooms, S.; Triantafyllidis, A.; Deloof, D.; Kappas, I.; Dierckens, K.; De Wolf, T.; Bossier, P.; Vadstein, O.; Kui, S.; et al. Evaluation of DNA methodologies in identifying Brachionus species used in European hatcheries. Aquaculture 2006, 255, 557–564. [Google Scholar] [CrossRef]
- Kostopoulou, V.; Miliou, H.; Katis, G.; Verriopoulos, G. Changes in the Population Structure of the Lineage ‘Nevada’ Belonging to the Brachionus plicatilis Species Complex, Batch-Cultured under Different Feeding Regimes. Aquac. Int. 2006, 14, 451–466. [Google Scholar] [CrossRef]
- Kostopoulou, V.; Miliou, H.; Krontira, Y.; Verriopoulos, G. Mixis in rotifers of the lineage ‘Nevada’, belonging to the Brachionus plicatilis species complex, under different feeding regimes. Aquac. Res. 2007, 38, 1093–1105. [Google Scholar] [CrossRef]
- Pourriot, R.; Rougier, C. Taux de reproduction en fonction de la concentration en nourriture et de la température chez trois espèces du genre Brachionus (Rotifères). Ann. Limnol. Int. J. Limnol. 1997, 33, 23–31. [Google Scholar] [CrossRef][Green Version]
- Blanchot, J.; Pourriot, R. Influence de trois facteurs de l’environnement, lumiere, temperature et salinite sur l’eclosion des oeufs de duree d’un clone de Brachionus plicatilis (O. F. Müller) rotifere. CR Acad. Sci. Paris 1982, 295, 243–246. [Google Scholar]
- Gómez, A.; Serra, M.; Carvalho, G.R.; Lunt, D.H. Speciation in ancient cryptic species complexes: Evidence from the molecular phylogeny of Brachionus plicatilis (Rotifera). Evolution 2002, 56, 1431–1444. [Google Scholar] [CrossRef]
- Mills, S.; Lunt, D.H.; Gómez, A. Global isolation by distance despite strong regional phylogeography in a small metazoan. BMC Evol. Biol. 2007, 7, 225. [Google Scholar] [CrossRef] [PubMed]
- Anitha, P.; George, R. The taxonomy of Brachionus plicatilis species complex (Rotifera: Monogononta) from the Southern Kerala (India) with a note on their repro- ductive preferences. J. Mar. Biol. Assoc. India 2006, 48, 6–13. [Google Scholar]
- Derry, A.M.; Hebert, P.D.N.; Prepas, E.E. Evolution of rotifers in saline and subsaline lakes: A molecular phylogenetic approach. Limnol. Oceanogr. 2003, 48, 675–685. [Google Scholar] [CrossRef]
- Kutikova, L.A. Parallelism in the evolution of rotifers. Hydrobiologia 1983, 104, 3–7. [Google Scholar] [CrossRef]
- Snell, T.W. Rotifers as models for the biology of aging. Int. Rev. Hydrobiol. 2014, 99, 84–95. [Google Scholar] [CrossRef]
- Korstad, J.; Neyts, A.; Danielsen, T.; Overrein, I.; Olsen, Y. Use of swimming speed and egg ratio as predictors of the status of rotifer cultures in aquaculture. Hydrobiologia 1995, 313–314, 395–398. [Google Scholar] [CrossRef]
- Gilbert, J.J. Monoxenic cultivation of the rotifer Brachionus calyciflorus in a defined medium. Oecologia 1970, 4, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Vizcardo, C.; Gil-Kodaka, P. Estructura de las comunidades Macrozoobentónicas de los Humedales de Ventanilla, Callao, Perú. An. Cient. 2015, 76, 1–11. [Google Scholar] [CrossRef]
- Aponte, H.; Ramírez, D.W.; Vargas, R. First stages of the post-fire natural regeneration of vegetation in the ventanilla wetlands (Lima-Peru). Ecol. Apl. 2017, 16, 23–30. [Google Scholar] [CrossRef]
- Rodríguez, R.; Retamozo-Chavez, R.; Aponte, H.; Valdivia, E. Evaluación microbiológica de un cuerpo de agua del ACR Humedales de Ventanilla (Callao, Perú) y su importancia para la salud pública local. Ecol. Apl. 2017, 16, 15–21. [Google Scholar] [CrossRef][Green Version]
- Gilbert, J.J. Dormancy in Rotifers. Trans. Am. Microsc. Soc. 1974, 93, 490–513. [Google Scholar] [CrossRef]
- Ricci, C. Dormancy patterns in rotifers. Hydrobiologia 2001, 446, 1–11. [Google Scholar] [CrossRef]
- WWT Consulting. Good Practices Handbook for Integrating Urban Development and Wetland Conservation; WWT Consulting: Slimbridge, UK, 2018; pp. 1–49. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Primer | Sequence (5′-3′) | Size (pb) | Reference |
|---|---|---|---|---|
| COI | ZplankF1_t1 | TGTAAAACGACGGCCAGTTCTASWAATCATAARGATATTGG | ~700 | [26] |
| ZplankR1_t1 | CAGGAAACAGCTATGACTTCAGGRTGRCCRAARAATCA | |||
| * M13F | TGTAAAACGACGGCCAGT | [27] | ||
| * M13R | CAGGAAACAGCTATGAC | |||
| ITS1 | III | CACACCGCCCGTCGCTACTACCGATTG | ~560 | [28] |
| VIII | GTGCGTTCGAAGTGTCGATGATCAA |
| Function | Eigenvalues | Variance (%) | Accumulated Variance (%) |
|---|---|---|---|
| 1 | 28.259 | 90.6 | 90.6 |
| 2 | 1.916 | 6.1 | 96.8 |
| 3 | 1.008 | 3.2 | 100.0 |
| Measurement (in Ln) | Function 1 | Function 2 | ||
|---|---|---|---|---|
| Coefficient | Correlation | Coefficient | Correlation | |
| (a) Lorica length | 1.236 | 0.857 | 0.092 | 0.195 |
| (b) Lorica aperture | −0.054 | 0.43 | 0.01 | −0.08 |
| (c) Lorica width | −0.64 | 0.252 | −0.452 | 0.079 |
| (d) Distance between central spines | 0.054 | 0.022 | 0.564 | 0.69 |
| (e) Dorsal sinus depth | −0.131 | 0.266 | 0.255 | 0.542 |
| (f) Distance between central and medial spines | 0.002 | −0.015 | −0.232 | 0.498 |
| (g) Medial spine length | −0.26 | 0.112 | 0.58 | 0.46 |
| (h) Head aperture | −0.122 | 0.432 | 0.28 | −0.034 |
| (i) Lateral spine length | 0.539 | 0.237 | −0.038 | 0.059 |
| Marker | Origin of Haplotypes | |||||
|---|---|---|---|---|---|---|
| (This Study) | Chile | * Mexico | China–USA | Australia–Japan–USA | France | |
| COI | Peru (Z010-VL) | 0.009–0.011 | 0.103 | 0.103 | 0.131 | 0.131 |
| ITS1 | Peru (Z010-VL) | 0 | 0 | 0 | 0 | 0 |
| Marker | Origin of Haplotypes | ||||||
|---|---|---|---|---|---|---|---|
| (This Study) | Spain | USA | Philippines–* Korea | Cayman Islands | Turkey | Italy | |
| COI | Peru (Z018-SD) | 0.045–0.056 | 0.056–0.060 | 0.062–0.122 | 0.060–0.124 | 0.124 | 0.146 |
| ITS1 | Peru (Z018-SD) | 0.003–0.009 | 0.009 | 0–0.003 | 0.009 | 0.003–0.009 | 0.012 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez-Dávila, P.P.A.; Sotil, G.; Adabache-Ortiz, A.; Cueva, D.; Silva-Briano, M. Integrative Taxonomy of Two Peruvian Strains of Brachionus plicatilis Complex with Potential in Aquaculture. Diversity 2021, 13, 671. https://doi.org/10.3390/d13120671
Sánchez-Dávila PPA, Sotil G, Adabache-Ortiz A, Cueva D, Silva-Briano M. Integrative Taxonomy of Two Peruvian Strains of Brachionus plicatilis Complex with Potential in Aquaculture. Diversity. 2021; 13(12):671. https://doi.org/10.3390/d13120671
Chicago/Turabian StyleSánchez-Dávila, Pedro Pablo Alonso, Giovanna Sotil, Araceli Adabache-Ortiz, Deivis Cueva, and Marcelo Silva-Briano. 2021. "Integrative Taxonomy of Two Peruvian Strains of Brachionus plicatilis Complex with Potential in Aquaculture" Diversity 13, no. 12: 671. https://doi.org/10.3390/d13120671
APA StyleSánchez-Dávila, P. P. A., Sotil, G., Adabache-Ortiz, A., Cueva, D., & Silva-Briano, M. (2021). Integrative Taxonomy of Two Peruvian Strains of Brachionus plicatilis Complex with Potential in Aquaculture. Diversity, 13(12), 671. https://doi.org/10.3390/d13120671