Strategy for the Removal of Satellite Bacteria from the Cultivated Diatom

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
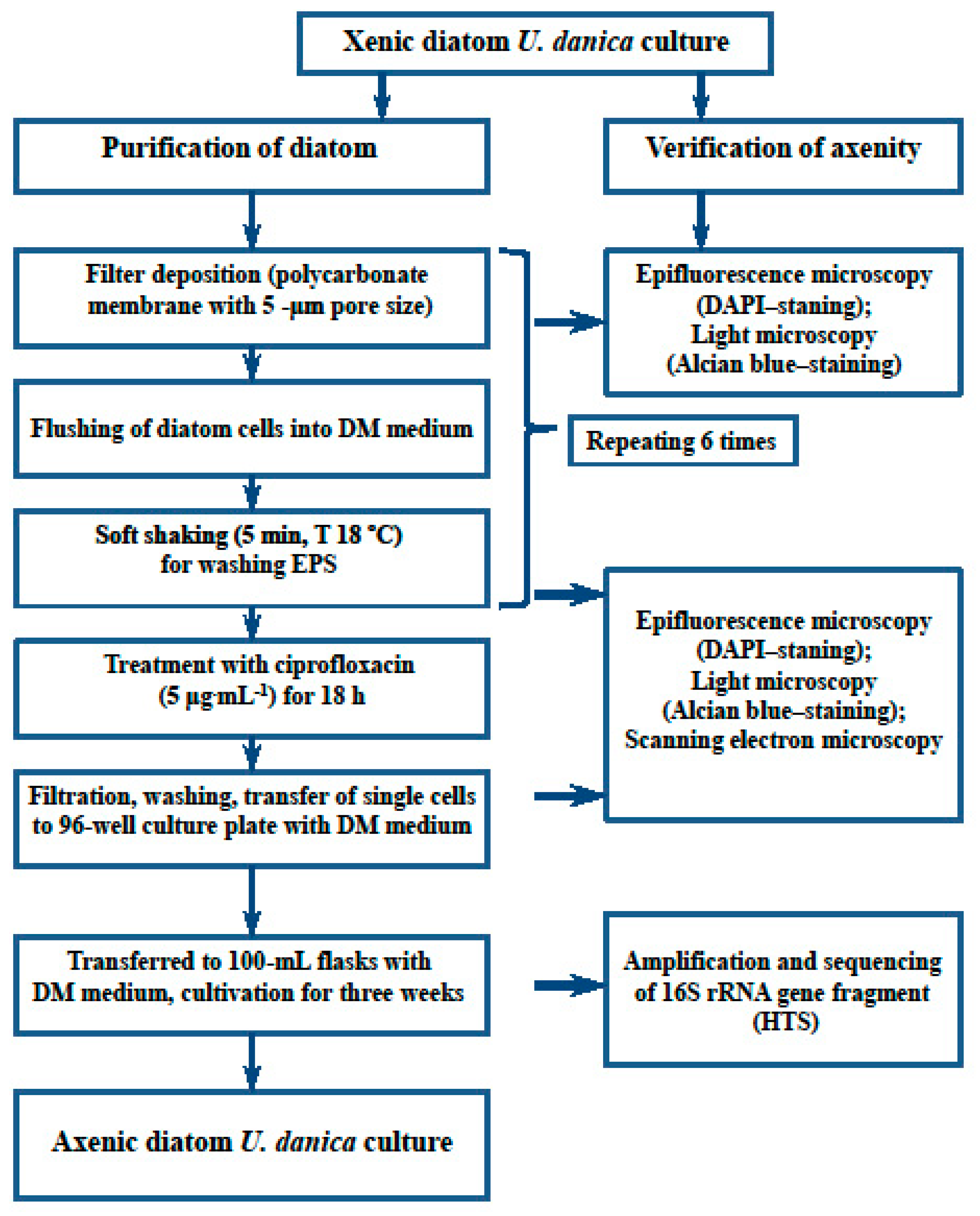
2.1. Isolation and Cultivation of Diatom Strains
2.2. Purification of Planktonic Diatom Ulnaria danica
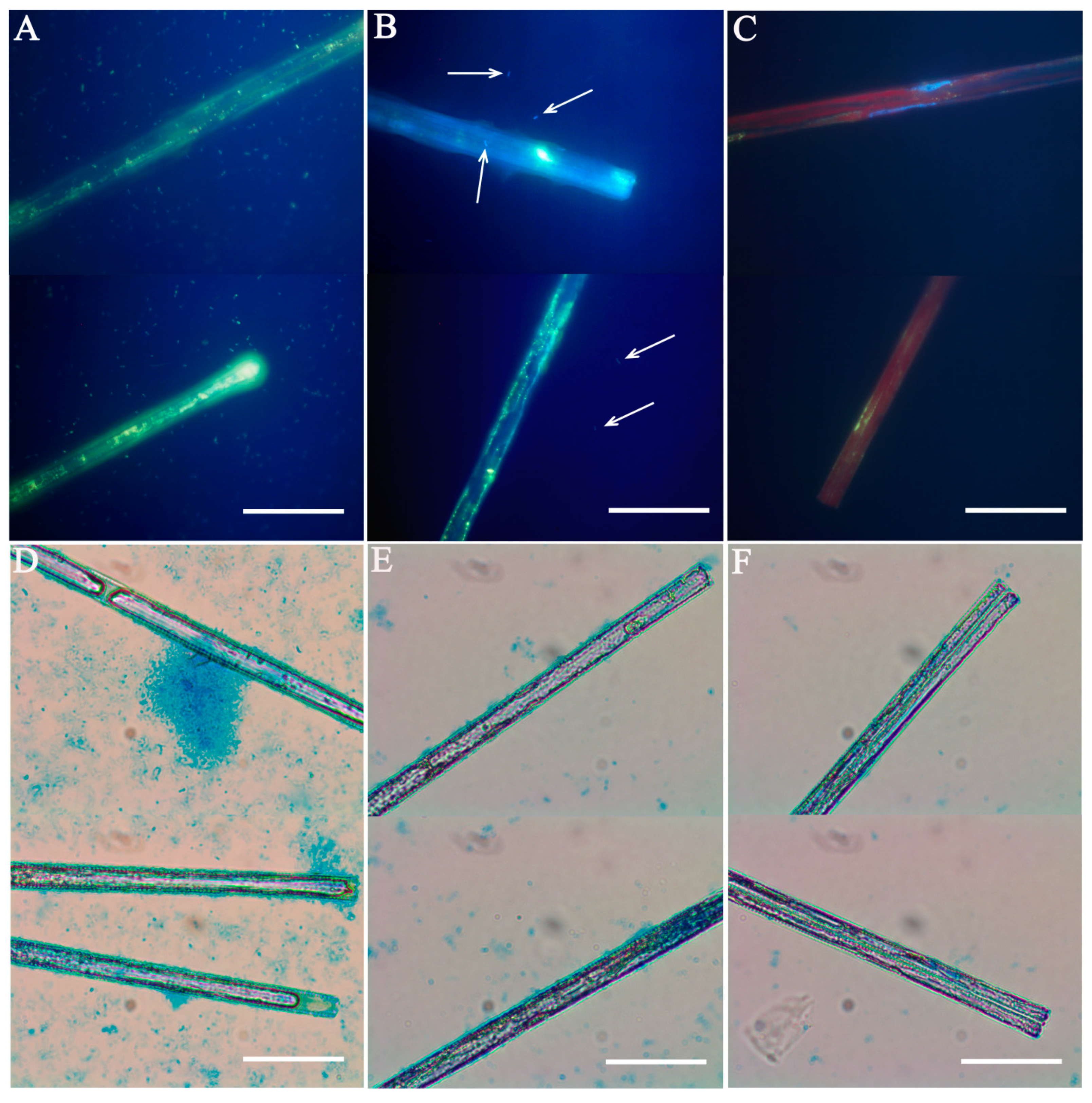
2.3. Verification of Diatom Culture Axenity
3. Results and Discussion
3.1. Removal of Associated Bacteria from the Diatom Monoclonal Culture
3.2. Verification of Axenity
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P.G. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (EPS) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Myklestad, S.M. Release of extracellular products by phytoplankton with special emphasis on polysaccharides. Sci. Total Environ. 1995, 165, 155–164. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Paterson, D.M. The importance of extracellular carbohydrate production by marine epipelic diatoms. Adv. Bot. Res. 2003, 40, 184–240. [Google Scholar]
- Bahulikar, R.A.; Kroth, P.G. Localization of EPS components secreted by freshwater diatoms using differential staining with fluorophore-conjugated lectins and other fluorochromes. Eur. J. Phycol. 2007, 42, 199–208. [Google Scholar] [CrossRef]
- Amin, S.A.; Parker, M.S.; Armbrust, E.V. Interactions between Diatoms and Bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 667. [Google Scholar] [CrossRef]
- Keeling, P.J.; Burki, F.; Wilcox, H.M.; Allam, B.; Allen, E.E.; Amaral-Zettler, L.A.; Armbrust, E.V.; Archibald, J.M.; Bharti, A.K.; Bell, C.J.; et al. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): Illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Biol. 2014, 12, e1001889. [Google Scholar] [CrossRef]
- Bowler, C.; Allen, A.E.; Badger, J.H.; Grimwood, J.; Jabbari, K.; Kuo, A.; Maheswari, U.; Martens, C.; Maumus, F.; Otillar, R.P.; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar] [CrossRef]
- Vancaester, E.; Depuydt, T.; Osuna-Cruz, C.M.; Vandepoele, K. Systematic and functional analysis of horizontal gene transfer events in diatoms. bioRxiv 2020. [Google Scholar] [CrossRef]
- Armbrust, E.V.; Berges, J.A.; Bowler, C.; Green, B.R.; Martinez, D.S.T.; Putnam, N.H.; Zhou, S.; E Allen, A.; Apt, K.E.; Bechner, M.; et al. The Genome of the Diatom Thalassiosira Pseudonana: Ecology, Evolution, and Metabolism. Science 2004, 306, 79–86. [Google Scholar] [CrossRef]
- Ashworth, J.; Coesel, S.; Lee, A.; Armbrust, E.V.; Orellana, M.V.; Baliga, N.S. Genome-wide diel growth state transitions in the diatom Thalassiosira pseudonana. Proc. Natl. Acad. Sci. USA 2013, 110, 7518–7523. [Google Scholar] [CrossRef] [PubMed]
- Galachyants, Y.P.; Zakharova, Y.R.; Petrova, D.P.; Morozov, A.A.; Sidorov, I.A.; Marchenkov, A.M.; Logacheva, M.D.; Markelov, M.L.; Khabudaev, K.V.; Likhoshway, Y.V.; et al. Sequencing of the complete genome of an araphid pennate diatom Synedra acus subsp. radians from Lake Baikal. Dokl. Biochem. Biophys. 2015, 461, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, R.P.; Tesson, B.; Norden-Krichmar, T.; Federowicz, S.; Hildebrand, M.; Allen, A.E. Whole transcriptome analysis of the silicon response of the diatom Thalassiosira pseudonana. BMC Genomics 2012, 13, 499. [Google Scholar] [CrossRef] [PubMed]
- Dyhrman, S.T.; Jenkins, B.D.; Rynearson, T.A.; Saito, M.A.; Mercier, M.L.; Alexander, H.; Whitney, L.P.; Drzewianowski, A.; Bulygin, V.A.; Bertrand, E.M.; et al. The transcriptome and proteome of the diatom Thalassiosira pseudonana reveal a diverse phosphorus stress response. PLoS ONE 2012, 7, e33768. [Google Scholar] [CrossRef]
- Remmers, I.M.; D’Adamo, S.; Martens, D.E.; de Vos, R.C.; Mumm, R.; America, T.A.; Cordewener, J.H.; Bakker, L.V.; Peters, S.A.; Wijffels, R.H.; et al. Orchestration of transcriptome, proteome and metabolome in the diatom Phaeodactylum tricornutum during nitrogen limitation. Algal Res. 2018, 35, 33–49. [Google Scholar] [CrossRef]
- Rastogi, A.; Maheswari, U.; Dorrell, R.G.; Vieira, F.R.J.; Maumus, F.; Kustka, A.; McCarthy, J.; E Allen, A.; Kersey, P.; Bowler, C.; et al. Integrative analysis of large scale transcriptome data draws a comprehensive landscape of Phaeodactylum tricornutum genome and evolutionary origin of diatoms. Sci. Rep. 2018, 8, 4834. [Google Scholar] [CrossRef]
- Galachyants, Y.P.; Zakharova, Y.R.; Volokitina, N.A.; Morozov, A.A.; Likhoshway, Y.V.; Grachev, M.A. De novo transcriptome assembly and analysis of the freshwater araphid diatom Fragilaria radians, Lake Baikal. Sci. Data 2019, 6, 183. [Google Scholar] [CrossRef]
- Daste, P.; Neuville, D.; Baptiste, B.V. A simple procedure for obtaining clonal isolation of diatoms. Eur. J. Phycol. 1983, 18, 1–3. [Google Scholar] [CrossRef]
- Brown, R.; Bischoff, H. A new and useful method for obtaining axenic cultures of algae. Phycol. Soc. Am. News Bull. 1962, 15, 43–44. [Google Scholar]
- Wigglesworth-Cooksey, B.; Cooksey, K.E. Use of fluorophore-conjugated lectins to study cell-cell interactions in model marine biofilms. Appl. Environ. Microbiol. 2005, 71, 428–435. [Google Scholar] [CrossRef]
- Windler, M.; Gruber, A.; Kroth, P.G. Technical note: Purification of benthic diatoms from associated bacteria using the antibiotic imipenem. J. Endocytobiosis Cell Res. 2012, 22, 62–65. [Google Scholar]
- Bruckner, C.G.; Kroth, P.G. Protocols for the removal of bacteria from freshwater benthic diatom cultures. J. Phycol. 2009, 45, 981–986. [Google Scholar] [CrossRef]
- Hunt, B.E.; Mandoli, D.F. Axenic cultures of Acetabularia (Chlorophyta): A decontamination protocol with potential application to other algae. J. Phycol. 1992, 28, 407–414. [Google Scholar] [CrossRef]
- Jones, A.K.; Rhodes, M.E.; Evans, S.C. The use of antibiotics to obtain axenic cultures of algae. Eur. J. Phycol. 1973, 8, 185–196. [Google Scholar] [CrossRef]
- Nagai, S.; Imai, I.; Manabe, T.A. simple and quick technique for establishing axenic cultures of the centric diatom Coscinodiscus wailesii Gran. J. Plankt. Res. 1998, 20, 1417–1420. [Google Scholar] [CrossRef]
- Staats, N.; de Winder, B.; Stal, L.J.; Mur, L.R. Isolation and characterization of extracellular polysaccharides from the epipelic diatoms Cylindrotheca closterium and Navicula salinarum. Eur. J. Phycol. 1999, 34, 161–169. [Google Scholar] [CrossRef]
- Kobayashi, H.; Oethinger, M.; Tuohy, M.J.; Procop, G.W.; Bauer, T.W. Improved detection of biofilm-formative bacteria by vortexing and sonication: A pilot study. Clin. Orthop. Rel. Res. 2009, 467, 1360–1364. [Google Scholar] [CrossRef]
- Shishlyannikov, S.M.; Zakharova, Y.R.; Volokitina, N.A.; Mikhailov, I.S.; Petrova, D.P.; Likhoshway, Y.V. A procedure for establishing an axenic culture of the diatom Synedra acus subsp. radians (Kutz.) Skabibitsch. from Lake Baikal. Limnol. Oceanogr. Methods 2011, 9, 478–484. [Google Scholar] [CrossRef]
- Agostini, V.O.; Borges, L.V.; Maxagata, E.; Abreu, P.C. Antibiotics choice: Procedure for establishment of Conticribra weissflogii and Isochrysis galbana cultures. Pan-Amer. J. Aquat. Sci. 2019, 14, 1–7. [Google Scholar]
- Agostini, V.O.; Lopes, L.F.P.; Macedo, A.J.; Muxagata, E. Review on the effects of antimicrobials use in cultures of planktonic organisms: A procedure for ecological experiments. Latin. Amer. J. Aquat. Res. 2019, 47, 394–415. [Google Scholar] [CrossRef]
- Ishii, K.I.; Imai, I.; Natsuike, M.; Sawayama, S.; Ishino, R.; Liu, W.; Fukusaki, K.; Ishikawaa, A. Simple technique for establishing axenic cultures of centric diatoms from resting stage cells in bottom sediments. Phycologia 2018, 57, 674–679. [Google Scholar] [CrossRef]
- Goto, N.; Kawamura, T.; Mitamura, O.; Terai, H. Importance of extracellular organic carbon production in the total primary production by tidal-flat diatoms in comparison to phytoplankton. Mar. Ecol. Prog. Ser. 1999, 190, 289–295. [Google Scholar] [CrossRef]
- Bruckner, C.G.; Bahulikar, R.; Rahalkar, M.; Schink, B.; Kroth, P.G. Bacteria associated with benthic diatoms from Lake Constance: Phylogeny and influences on diatom growth and EPS secretion. Appl. Environ. Microbiol. 2008, 74, 7740–7749. [Google Scholar] [CrossRef] [PubMed]
- Debenest, T.; Silvestre, J.; Coste, M.; Delmas, F.; Pinelli, E. A new cell primo-culture method for freshwater benthic diatom communities. J. Appl. Phycol. 2009, 21, 65–73. [Google Scholar] [CrossRef]
- Ceri, H.; Olson, M.E.; Stremick, C.; Read, R.R.; Morck, D.; Buret, A. The calgary biofilm device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 1999, 37, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Zakharova, Y.R.; Adelshin, R.V.; Parfenova, V.V.; Bedoshvili, Y.D.; Likhoshway, Y.V. Taxonomic characterization of the microorganisms associated with the cultivable diatom Synedra acus from Lake Baikal. Microbiology 2010, 79, 679–687. [Google Scholar] [CrossRef]
- Zakharova, Y.R.; Bedoshvili, Y.D.; Petrova, D.P.; Marchenkov, A.M.; Volokitina, N.A.; Bashenkhaeva, M.V.; Kopyrina, L.I.; Grachev, M.A.; Likhoshway, Y.V. Morphological description and molecular phylogeny of two diatom clones from the genus Ulnaria (Kützing) Compère isolated from an ultraoligotrophic lake at the Pole of Cold in the Northern Hemisphere, Republic of Sakha (Yakutia), Russia. Cryptogam. Algol. 2020, 41, 37–45. [Google Scholar] [CrossRef]
- Van Tol, H.M.; Amin, S.A.; Armbrust, E.V. Ubiquitous marine bacterium inhibits diatom cell division. ISME J. 2016, 1–12. [Google Scholar] [CrossRef]
- Behringer, G.; Ochsenkühn, M.A.; Fei, C.; Fanning, J.; Koester, J.A.; Amin, S.A. Bacterial communities of diatoms display strong conservation across strains and time. Front. Microbiol. 2018, 9, 659. [Google Scholar] [CrossRef]
- Thompson, A.S.; Rhodes, J.C.; Pettman, I. Culture Collection of Algae and Protozoa, Catalogue of Strains; Freshwater Biological Association: Ambleside, UK, 1988. [Google Scholar]
- Safonova, T.A.; Aslamov, I.A.; Basharina, T.N.; Chenski, A.G.; Vereschagin, A.L.; Glyzina, O.Y.; Grachev, M.A. Cultivation and automatic counting of diatom algae cells in multi-well plastic plates. Diatom Res. 2007, 22, 189–195. [Google Scholar] [CrossRef]
- Marchenkov, A.M.; Petrova, D.P.; Morozov, A.A.; Zakharova, Y.R.; Grachev, M.A.; Bondar, A. A family of silicon transporter structural genes in a pennate diatom Synedra ulna subsp. danica (Kütz.) Skabitsch. PLoS ONE 2018, 13, e0203161. [Google Scholar] [CrossRef]
- Bates, S.T.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. ISME J. 2011, 5, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Opensource, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Galachyants, Y.P.; Morozov, A.A.; Mardanov, A.V.; Beletsky, A.V.; Ravin, N.V.; Petrova, D.P.; Likhoshway, E.V. Complete Chloroplast Genome Sequence of Freshwater Araphid Pennate Diatom Alga Synedra acus from Lake Baikal. Int. J. Biol. 2012, 4, 27–35. [Google Scholar] [CrossRef][Green Version]
- Marchenkov, A.M.; Bondar, A.A.; Petrova, D.P.; Habudaev, K.V.; Galachyants, Y.P.; Zakharova, Y.R.; Volokitina, N.A.; Grachev, M.A. Unique configuration of genes of silicon transporter in the freshwater pennate diatom Synedra acus subsp. radians. Dokl. Biochem. Biophys. 2016, 471, 407–409. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.A.; Hmelo, L.R.; van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef]
- Doghri, I.; Lavaud, J.; Dufour, A.; Bazire, A.; Lanneluc, I.; Sable, S. Cell-bound exopolysaccharides from an axenic culture of the intertidal mudflat Navicula phyllepta diatom affect biofilm formation by benthic bacteria. J. Appl. Phycol. 2017, 29, 165–177. [Google Scholar] [CrossRef]
- Mönnich, J.; Tebben, J.; Bergemann, J.; Case, R.; Wohlrab, S.; Harder, T. Niche-based assembly of bacterial consortia on the diatom Thalassiosira rotula is stable and reproducible. ISME J. 2020, 14, 1614–1625. [Google Scholar] [CrossRef]
- Khandeparker, L.; D’Costa, P.M.; Anil, A.C.; Sawant, S.S. Interactions of bacteria with diatoms: Influence on natural marine biofilms. Mar. Ecol. 2014, 35, 233–248. [Google Scholar] [CrossRef]
- Scholz, B. Purification and culture characteristics of 36 benthic marine diatoms isolated from the Solthörn tidal flat (southern North Sea). J. Phycol. 2014, 50, 685–697. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Orvain, F.; de Crignis, M.; Guizien, K.; Lefebvre, S.; Mallet, C.; Takahashi, E.; Dupuy, C. Tidal and seasonal effects on the short-term temporalpatterns of bacteria, microphytobenthos and exopolymers in natural intertidal biofilms (Brouage, France). J. Sea Res. 2014, 92, 6–18. [Google Scholar] [CrossRef]
- Schallenberg, M.; Kalff, J.; Rasmussen, J. Solutions to problems in enumerating sediment bacteria by direct counts. Appl. Environ. Microb. 1989, 55, 1214–1219. [Google Scholar] [CrossRef] [PubMed]
- Steedman, H.F. Alcian blue 8GS: A new stain for mucin. Q. J. Microsc. Sci. 1950, 91, 477–479. [Google Scholar]
- Ramus, J. Alcian blue: A quantitative aqueous assay for algal acid and sulfated polysaccharides. J. Phycol. 1977, 13, 345–348. [Google Scholar] [CrossRef]
- Passow, U.; Alldredge, A.L. A dye-binding assay for the spectrophotometric measurement of transparent exopolymer particles (TEP). Limnol. Oceanogr. 1995, 40, 1326–1335. [Google Scholar] [CrossRef]
- Claquin, P.; Probert, I.; Lefebvre, S.; Veron, B. Effects of temperature on photosynthetic parameters and TEP production in eight species of marine microalgae. Aquat. Microb. Ecol. 2008, 51, 1–11. [Google Scholar] [CrossRef]
- Passow, U. Transparent exopolymer particles (TEP) in aquatic environments. Prog. Oceanogr. 2002, 55, 287–333. [Google Scholar] [CrossRef]
- Krembs, C.; Eicken, H.; Deming, J.W. Exopolymer alteration of physical properties of sea ice and implications for ice habitability and biogeochemistry in a warmer Arctic. Proc. Natl. Acad. Sci. USA 2011, 108, 3653–3658. [Google Scholar] [CrossRef]
- Bosak, S.; Pletikapić, G.; Hozić, A.; Svetličić, V.; Sarno, D.; Viličić, D. A Novel Type of Colony Formation in Marine Planktonic Diatoms Revealed by Atomic Force Microscopy. PLoS ONE 2012, 7, e44851. [Google Scholar] [CrossRef] [PubMed]
- Winand, R.; Bogaerts, B.; Hoffman, S.; Lefevre, L.; Delvoye, M.; van Braekel, J.; Fu, Q.; Roosens, N.H.C.; de Keersmaecker, S.C.J.; Vanneste, K. Targeting the 16S rRNA Gene for Bacterial Identification in Complex Mixed Samples: Comparative Evaluation of Second (Illumina) and Third (Oxford Nanopore Technologies) Generation Sequencing Technologies. Int. J. Mol. Sci. 2019, 21, 298. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Exponential Phase of Growth (cells/ mL) | Latent Phase of Growth (cells/ mL) |
|---|---|---|
| Initial algal suspension before treatment | 1.0 × 107 ± 2.9 × 105 | 5.0 × 105 ± 4.8 × 104 |
| Physico-chemical treatment | ||
| Filtering | 9.0 × 105 ± 7.0 × 104 | 4.3 × 104 ± 2.2 × 103 |
| Triton X-100 *+ filtering | 2.1×105 ± 4.3 × 103 | not used |
| Washing EPS | not used | 1.0 × 104 ± 103 |
| Antibiotic treatment after filtering | ||
| Ciprofloxacin | 4.7 × 104 ± 4.4 × 103 | no bacteria detected |
| Kanamycin * | 1.5 × 105 ± 2.2 × 103 | not used |
| Ciprofloxacin + Kanamycin * | 1.0 × 105 ± 2.2 × 103 | not used |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakharova, Y.; Marchenkov, A.; Volokitina, N.; Morozov, A.; Likhoshway, Y.; Grachev, M. Strategy for the Removal of Satellite Bacteria from the Cultivated Diatom. Diversity 2020, 12, 382. https://doi.org/10.3390/d12100382
Zakharova Y, Marchenkov A, Volokitina N, Morozov A, Likhoshway Y, Grachev M. Strategy for the Removal of Satellite Bacteria from the Cultivated Diatom. Diversity. 2020; 12(10):382. https://doi.org/10.3390/d12100382
Chicago/Turabian StyleZakharova, Yulia, Artem Marchenkov, Nadezhda Volokitina, Aleksey Morozov, Yelena Likhoshway, and Mikhail Grachev. 2020. "Strategy for the Removal of Satellite Bacteria from the Cultivated Diatom" Diversity 12, no. 10: 382. https://doi.org/10.3390/d12100382
APA StyleZakharova, Y., Marchenkov, A., Volokitina, N., Morozov, A., Likhoshway, Y., & Grachev, M. (2020). Strategy for the Removal of Satellite Bacteria from the Cultivated Diatom. Diversity, 12(10), 382. https://doi.org/10.3390/d12100382