Inter-Habitat Variability in Parrotfish Bioerosion Rates and Grazing Pressure on an Indian Ocean Reef Platform


Abstract
1. Introduction
2. Materials and Methods
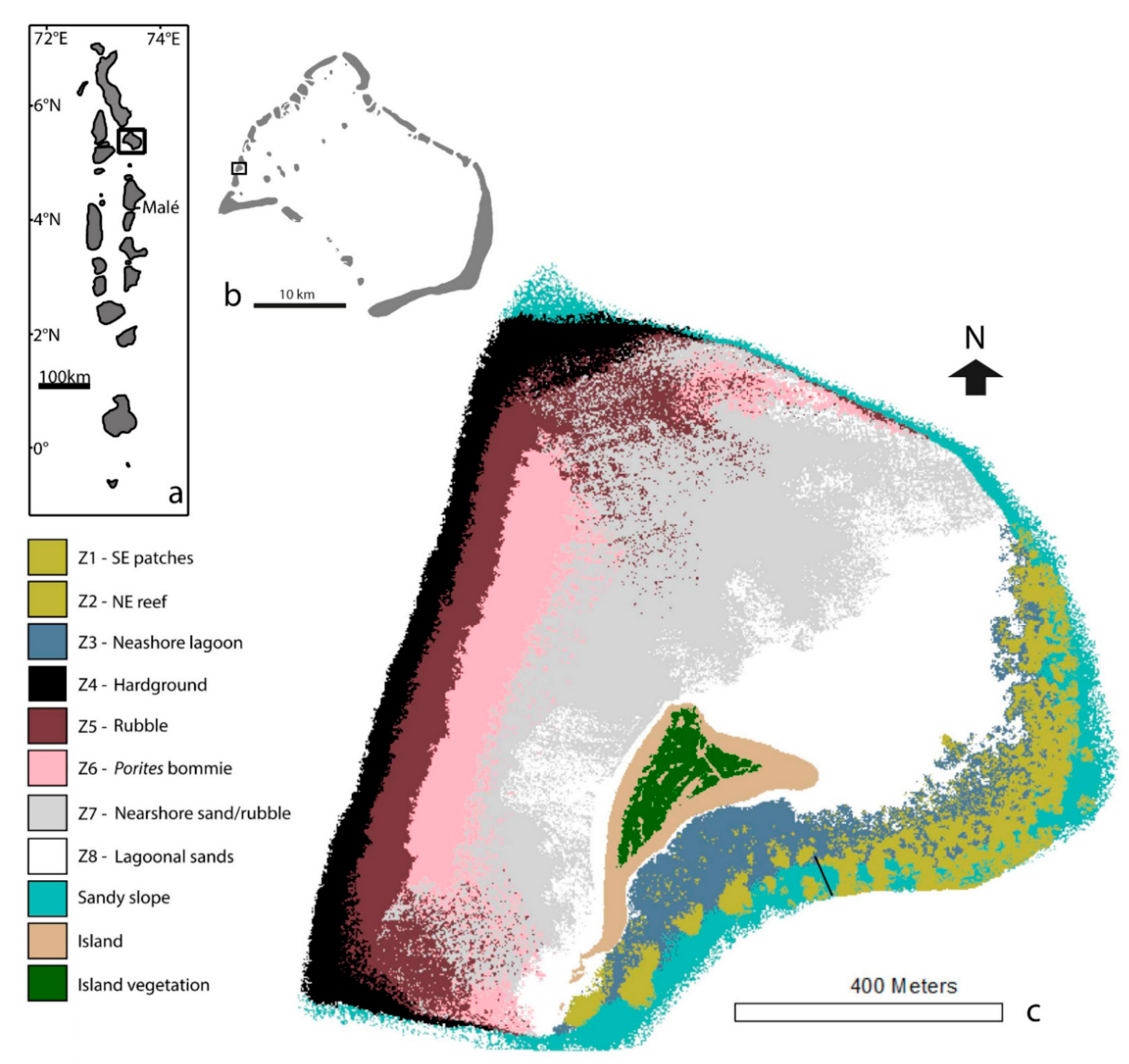
2.1. Study Site
2.2. Remote Underwater Video (RUV) Surveys
2.3. Inter-Habitat Variability in Total Parrotfish Bioerosion Rates
2.4. Inter-Habitat Variability in Total Parrotfish Grazing Pressure
3. Results
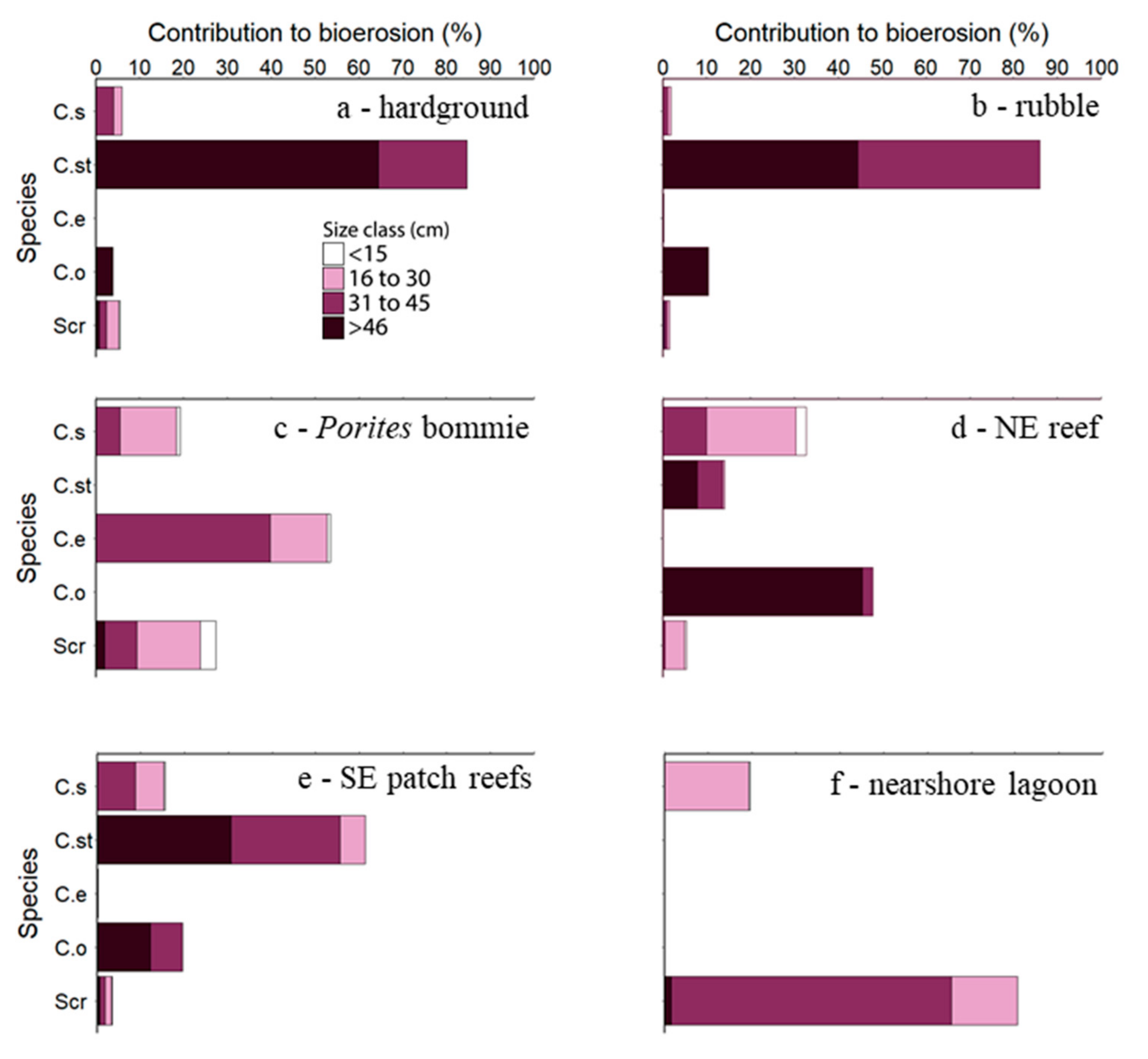
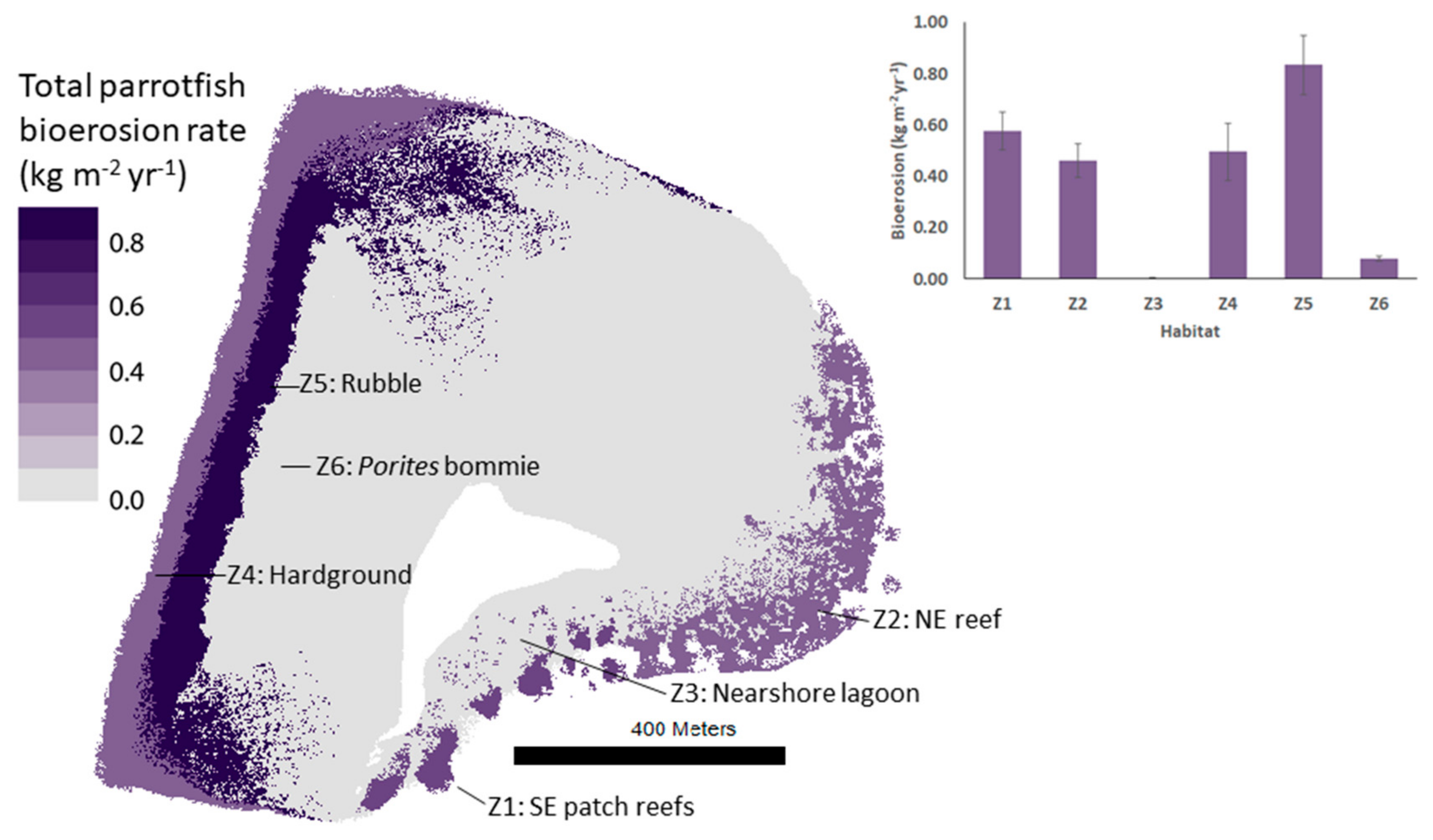
3.1. Species Contributions to Bioerosion and Inter-Habitat Variability in Bioerosion Rates
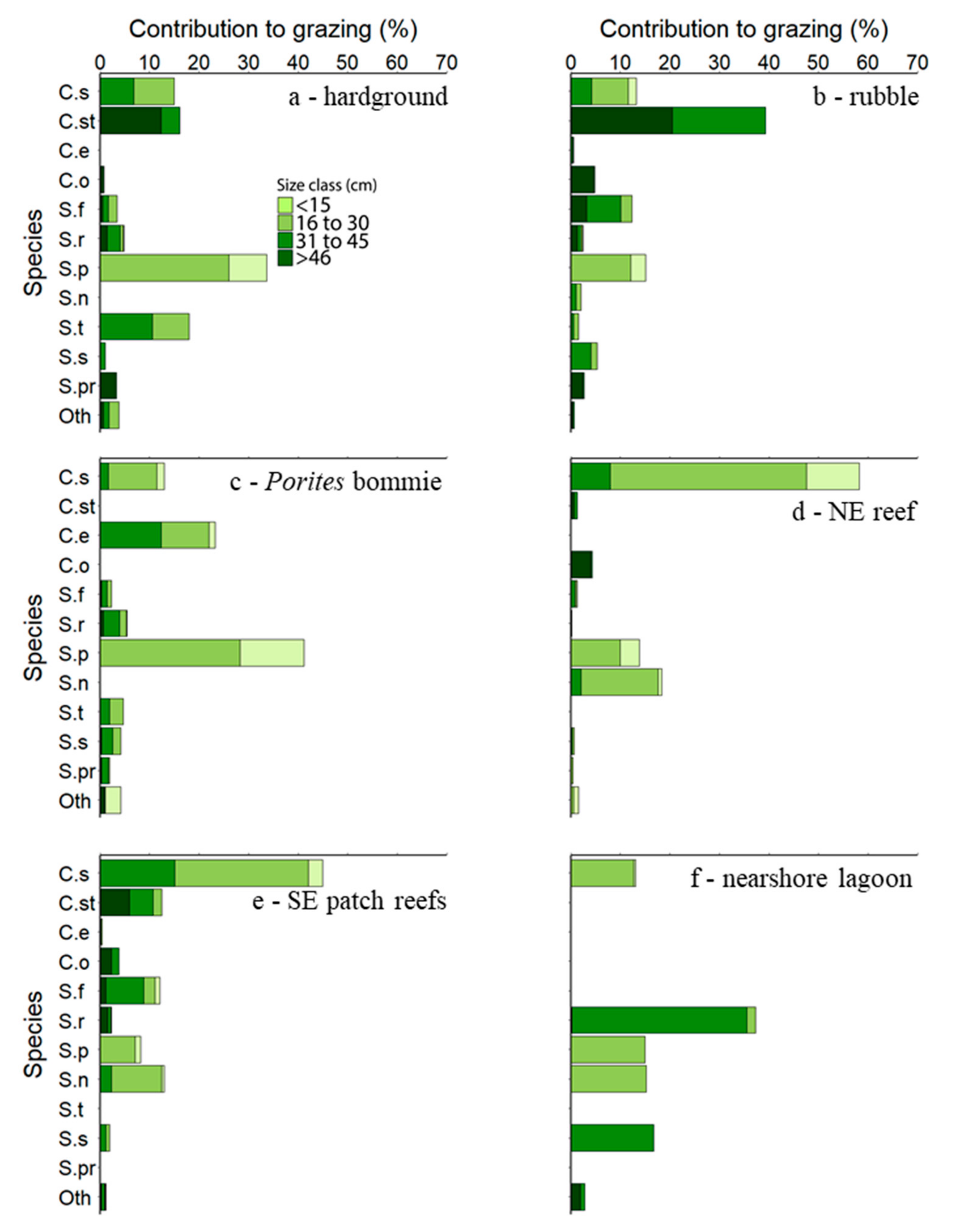
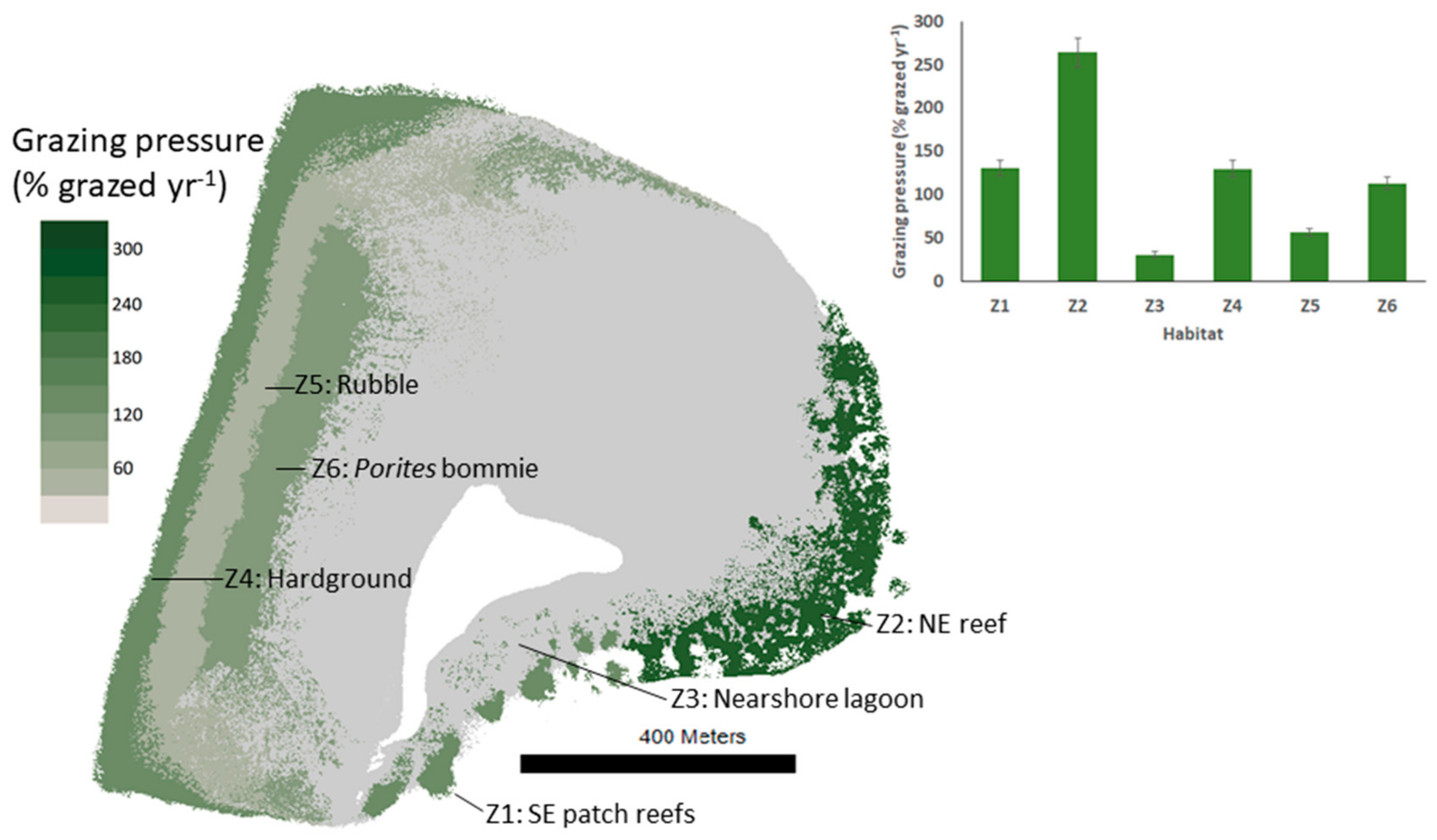
3.2. Species Contributions to Grazing and Interhabitat Variability in Grazing Pressure
4. Discussion
4.1. Spatial Patterns of Parrotfish Bioerosion Rate and Grazing Pressure
4.2. Factors Influencing the Observed Spatial Patterns
4.3. Implications
Data Access
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wild, C.; Hoegh-Guldberg, O.; Naumann, M.S.; Colombo-Pallotta, M.F.; Ateweberhan, M.; Fitt, W.K.; Iglesias-Prieto, R.; Palmer, C.; Bythell, J.C.; Ortiz, J.C.; et al. Climate change impedes scleractinian corals as primary reef ecosystem engineers. Mar. Freshw. Res. 2011, 62, 205–215. [Google Scholar] [CrossRef]
- Lugo-Fernández, A.; Roberts, H.H.; Suhayda, J.N. Wave transformations across a caribbean fringing-barrier Coral Reef. Cont. Shelf Res. 1998, 18, 1099–1124. [Google Scholar] [CrossRef]
- Moberg, F.F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Ruckelshaus, M.; Doney, S.C.; Galindo, H.M.; Barry, J.P.; Chan, F.; Duffy, J.E.; English, C.A.; Gaines, S.D.; Grebmeier, J.M.; Hollowed, A.B.; et al. Securing ocean benefits for society in the face of climate change. Mar. Policy 2013, 40, 154–159. [Google Scholar] [CrossRef]
- Ferrario, F.; Beck, M.W.; Storlazzi, C.D.; Micheli, F.; Shepard, C.C.; Airoldi, L. The effectiveness of coral reefs for coastal hazard risk reduction and adaptation. Nat. Commun. 2014, 5, 3794. [Google Scholar] [CrossRef] [PubMed]
- Williams, I.; Polunin, N. Large-scale associations between macroalgal cover and grazer biomass on mid-depth reefs in the Caribbean. Coral Reefs 2001, 19, 358–366. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Goatley, C.H.R.; Bellwood, D.R. Algal turf sediments across the Great Barrier Reef: Putting coastal reefs in perspective. Mar. Pollut. Bull. 2018, 137, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Box, S.J.; Mumby, P.J. Effect of macroalgal competition on growth and survival of juenile Caribbean corals. Mar. Ecol. Prog. Ser. 2007, 342, 139–149. [Google Scholar] [CrossRef]
- Nugues, M.M.; Bak, R.P.M. Differential competitive abilities between Caribbean coral species and a brown alga: A year of experiments and a long-term perspective. Mar. Ecol. Prog. Ser. 2006, 315, 75–86. [Google Scholar] [CrossRef]
- Nugues, M.M.; Delvoye, L.; Bak, R.P.M. Coral defence against macroalgae: Differential effects of mesenterial filaments on the green alga Halimeda opuntia. Mar. Ecol. Prog. Ser. 2004, 278, 103–114. [Google Scholar] [CrossRef]
- Mumby, P.J.; Harborne, A.R.; Williams, J.; Kappel, C.V.; Brumbaugh, D.R.; Micheli, F.; Holmes, K.E.; Dahlgren, C.P.; Paris, C.B.; Blackwell, P.G. Trophic cascade facilitates coral recruitment in a marine reserve. Proc. Natl. Acad. Sci. USA 2007, 104, 20. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, E.V.; Perry, C.T.; Halloran, P.R.; Iglesias-Prieto, R.; Schönberg, C.H.L.L.; Wisshak, M.; Form, A.U.; Carricart-Ganivet, J.P.; Fine, M.; Eakin, C.M.; et al. Avoiding coral reef functional collapse requires local and global action. Curr. Biol. 2013, 23, 912–918. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.T.; Murphy, G.N.; Kench, P.S.; Smithers, S.G.; Edinger, E.N.; Steneck, R.S.; Mumby, P.J. Caribbean-wide decline in carbonate production threatens coral reef growth. Nat. Commun. 2013, 4, 1402. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R. Patterns of bioerosion and sediment release by parrotfishes (family Scaridae) on the Great Barrier. Mar. Ecol. Prog. Ser. 1995, 117, 127–136. [Google Scholar] [CrossRef]
- Lokrantz, J.; Nyström, M.; Thyresson, M.; Johansson, C. The non-linear relationship between body size and function in parrotfishes. Coral Reefs 2008, 27, 967–974. [Google Scholar] [CrossRef]
- Perry, C.T.; Murphy, G.N.; Kench, P.S.; Edinger, E.N.; Smithers, S.G.; Steneck, R.S.; Mumby, P.J. Changing dynamics of Caribbean reef carbonate budgets: Emergence of reef bioeroders as critical controls on present and future reef growth potential. Proc. R. Soc. B Biol. Sci. 2014, 281, 20142018. [Google Scholar] [CrossRef]
- Clements, K.D.; German, D.P.; Piché, J.; Tribollet, A.D.; Choat, J.H. Integrating ecological roles and trophic resources on coral reefs: Multiple lines of evidence identify parrotfishes as microphages. Biol. J. Linn. Soc. 2017, 120, 729–751. [Google Scholar] [CrossRef]
- Nicholson, G.M.; Clements, K.D. Resolving resource partitioning in parrotfishes (Scarini) using microhistology of feeding substrata. Coral Reefs 2020. [Google Scholar] [CrossRef]
- Bruggemann, J.; van Oppen, M.; Breeman, A. Foraging by the stoplight parrotfish Sparisoma viride. I. Food selection in different, socially determined habitats. Mar. Ecol. Prog. Ser. 1994, 106, 41–55. [Google Scholar] [CrossRef]
- Bellwood, D.R. Direct estimate of bioerosion by two parrotfish species, Chlorurus gibbus and C. sordidus, on the Great Barrier Reef, Australia. Mar. Biol. 1995, 121, 419–429. [Google Scholar] [CrossRef]
- Afeworki, Y.; Bruggemann, J.H.; Videler, J.J. Limited flexibility in resource use in a coral reef grazer foraging on seasonally changing algal communities. Coral Reefs 2011, 30, 109–122. [Google Scholar] [CrossRef][Green Version]
- Bellwood, D.R.; Choat, J.H. A functional analysis of grazing in parrotfishes (family Sacaridae): The ecological implications. Environ. Biol. Fishes 1990, 28, 189–214. [Google Scholar] [CrossRef]
- Carr, A.; Tibbetts, I.R.; Kemp, A.; Truss, R.; Drennan, J. Inferring Parrotfish (Teleostei: Scaridae) Pharyngeal Mill Function From Dental Morphology, Wear, and Microstructure. J. Morphol. 2006, 267, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Bellwood, D.R. Production and reworking of sediment by parrotfishes (family Scaridae) on the Great Barrier Reef, Australia. Mar. Biol. 1996, 125, 795–800. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2008, 27, 37–47. [Google Scholar] [CrossRef]
- Perry, C.T.; Kench, P.S.; O’Leary, M.J.; Morgan, K.M.; Januchowski-Hartley, F. Linking reef ecology to island building: Parrotfish identified as major producers of island-building sediment in the Maldives. Geology 2015, 43, 503–506. [Google Scholar] [CrossRef]
- Morgan, K.M.; Kench, P.S. Parrotfish erosion underpins reef growth, sand talus development and island building in the Maldives. Sediment. Geol. 2016, 341, 50–57. [Google Scholar] [CrossRef]
- Perry, C.T.; Morgan, K.M.; Yarlett, R.T. Reef Habitat Type and Spatial Extent as Interacting Controls on Platform-Scale Carbonate Budgets. Front. Mar. Sci. 2017, 4, 185. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nystrom, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Mumby, P.J. The Impact of Exploiting Grazers (Scaridae) on the Dynamics of Caribbean Coral Reefs. Ecol. Appl. 2006, 16, 747–769. [Google Scholar] [CrossRef]
- Bruggemann, J.H.; van Kessel, A.M.; van Rooij, J.M.; Breeman, A.M. Bioerosion and sediment ingestion by the caribbean parrotfish Scarus vetula and Sparisoma viride: Implications of fish size, feeding mode and habitat use. Mar. Ecol. Prog. Ser. 1996, 134, 59–71. [Google Scholar] [CrossRef]
- Ong, L.; Holland, K.N. Bioerosion of coral reefs by two Hawaiian parrotfishes: Species, size differences and fishery implications. Mar. Biol. 2010, 157, 1313–1323. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Arthur, R. The Effect of Marine Reserves and Habitat on Populations of East African Coral Reef Fishes. Ecol. Appl. 2001, 11, 559–569. [Google Scholar] [CrossRef]
- Darling, E.S.; Graham, N.A.J.; Januchowski-Hartley, F.A.; Nash, K.L.; Pratchett, M.S.; Wilson, S.K. Relationships between structural complexity, coral traits, and reef fish assemblages. Coral Reefs 2017, 36, 561–575. [Google Scholar] [CrossRef]
- Ruttenberg, B.I.; Adam, T.C.; Duran, A.; Burkepile, D.E. Identity of coral reef herbivores drives variation in ecological processes over multiple spatial scales. Ecol. Appl. 2019, 29, e01893. [Google Scholar] [CrossRef] [PubMed]
- Hoey, A.S.; Feary, D.A.; Burt, J.A.; Vaughan, G.; Pratchett, M.S.; Berumen, M.L. Regional variation in the structure and function of parrotfishes on Arabian reefs. Mar. Pollut. Bull. 2016, 105, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, C.A.M.M.; Mendes, T.C.; Harborne, A.R.; Ferreira, C.E.L. Spatial distribution of nominally herbivorous fishes across environmental gradients on Brazilian rocky reefs. J. Fish Biol. 2015, 89, 939–958. [Google Scholar] [CrossRef]
- Yadav, S.; Abdulla, A.; Bertz, N.; Mawyer, A. King tuna: Indian Ocean trade, offshore fishing, and coral reef resilience in the Maldives archipelago. ICES J. Mar. Sci. 2019, 77, 398–407. [Google Scholar] [CrossRef]
- Green, A.L.; Bellwood, D.R. Monitoring Functional Groups of Herbivorous Reef Fishes as Indicators of Coral Reef Resilience A Practical Guide for Coral Reef Managers in the Asia Pacific Region; IUCN Working Group on Climate Change and Coral Reefs; IUCN: Gland, Switzerland, 2009. [Google Scholar]
- Streit, R.P.; Cumming, G.S.; Bellwood, D.R. Patchy delivery of functions undermines functional redundancy in a high diversity system. Funct. Ecol. 2019, 33, 1144–1155. [Google Scholar] [CrossRef]
- Harvey, E.; Fletcher, D.; Shortis, M.; Kendrick, G.A. A comparison of underwater visual distance estimates made by scuba divers and a stereo-video system: Implications for underwater visual census of reef fish abundance. Mar. Freshw. Res. 2004, 55, 573–580. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Streit, R.P.; Brandl, S.J.; Tebbett, S.B. The meaning of the term ‘function’ in ecology: A coral reef perspective. Funct. Ecol. 2019, 33, 948–961. [Google Scholar]
- Watson, D.L.; Harvey, E.S.; Fitzpatrick, B.M.; Langlois, T.J.; Shedrawi, G. Assessing reef fish assemblage structure: How do different stereo-video techniques compare? Mar. Biol. 2010, 157, 1237–1250. [Google Scholar] [CrossRef]
- Yarlett, R.T.; Perry, C.T.; Wilson, R.W.; Philpot, K.E. Constraining species-size class variability in rates of parrotfish bioerosion on Maldivian coral reefs: Implications for regional-scale bioerosion estimates. Mar. Ecol. Prog. Ser. 2018, 590, 155–169. [Google Scholar] [CrossRef]
- Choat, J.H.; klanten, O.S.; van Herwerden, L.; Robertson, D.R.; Clements, K.D. Patterns and processes in the evolutionary history of parrotfishes (Family Labridae). Biol. J. Linn. Soc. 2012, 107, 529–557. [Google Scholar] [CrossRef]
- Carlson, P.M.; Davis, K.; Warner, R.R.; Caselle, J.E. Fine-scale spatial patterns of parrotfish herbivory are shaped by resource availability. Mar. Ecol. Prog. Ser. 2017, 577, 165–176. [Google Scholar] [CrossRef]
- Johnson, G.B.; Taylor, B.M.; Robbins, W.D.; Franklin, E.C.; Toonen, R.; Bowen, B.; Choat, J.H. Diversity and Structure of Parrotfish Assemblages across the Northern Great Barrier Reef. Diversity 2019, 11, 14. [Google Scholar] [CrossRef]
- Adam, T.C.; Kelley, M.; Ruttenberg, B.I.; Burkepile, D.E. Resource partitioning along multiple niche axes drives functional diversity in parrotfishes on Caribbean coral reefs. Oecologia 2015, 179, 1173–1185. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Wilson, S.K.; Carr, P.; Hoey, A.S.; Jennings, S.; MacNeil, M.A. Seabirds enhance coral reef productivity an functioning in the absence of invasive rates. Nature 2018, 559, 250–253. [Google Scholar] [CrossRef]
- Alwany, M.A.; Thaler, E.; Stachowitsch, M. Parrotfish bioerosion on Egyptian Red Sea reefs. J. Exp. Mar. Bio. Ecol. 2009, 371, 170–176. [Google Scholar] [CrossRef]
- Brandl, S.J.; Bellwood, D.R. Individual-based analyses reveal limited functional overlap in a coral reef fish community. J. Anim. Ecol. 2014, 83, 661–670. [Google Scholar] [CrossRef]
- McClanahan, T.R. Coral Reefs effects of fishing, substrate complexity, and sea urchins. Coral Reefs 1994, 13, 231–241. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Nash, K.L. The importance of structural complexity in coral reef ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- Komyakova, V.; Munday, P.L.; Jones, G.P. Relative importance of coral cover, habitat complexity and diversity in determining the structure of reef fish communities. PLoS ONE 2013, 8, e83178. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Parrish, J.D. Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. J. Exp. Mar. Biol. Ecol. 1998, 224, 1–30. [Google Scholar] [CrossRef]
- Mumby, P.J. Stratifying herbivore fisheries by habitat to avoid ecosystem overfishing of coral reefs. Fish Fish. 2016, 17, 266–278. [Google Scholar] [CrossRef]
- Fulton, C.J.; Bellwood, D.R. Wave exposure, swimming performance, and the structure of tropical and temperate reef fish assemblages. Mar. Biol. 2004, 144, 429–437. [Google Scholar] [CrossRef]
- Harborne, A.R. The ecology, behaviour and physiology of fishes on coral reef flats, and the potential impacts of climate change. J. Fish Biol. 2013, 83, 417–447. [Google Scholar] [CrossRef] [PubMed]
- Bejarano, S.; Jouffray, J.B.; Chollett, I.; Allen, R.; Roff, G.; Marshell, A.; Steneck, R.; Ferse, S.C.A.; Mumby, P.J. The shape of success in a turbulent world: Wave exposure filtering of coral reef herbivory. Funct. Ecol. 2017, 31, 1312–1324. [Google Scholar] [CrossRef]
- O’Farrell, S.; Harborne, A.R.; Bozec, Y.M.; Luckhurst, B.E.; Mumby, P.J. Protection of functionally important parrotfishes increases their biomass but fails to deliver enhanced recruitment. Mar. Ecol. Prog. Ser. 2015, 522, 245–254. [Google Scholar] [CrossRef]
- Mumby, P.J.; Wabnitz, C.C.C. Spatial patterns of aggression, territory size, and harem size in five sympatric Caribbean parrotfish species. Environ. Biol. Fishes 2002, 63, 265–279. [Google Scholar] [CrossRef]
- Afeworki, Y.; Zekeria, Z.A.; Videler, J.J.; Bruggemann, J.H. Food intake by the parrotfish Scarus ferrugineus varies seasonally and is determined by temperature, size and territoriality. Mar. Ecol. Prog. Ser. 2013, 489, 213–224. [Google Scholar] [CrossRef]
- Graham, N.A.J. Habitat complexity: Coral structural loss leads to fisheries declines. Curr. Biol. 2014, 24, R359–R361. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.; Blanchard, J.L.; Mumby, P.J. Vulnerability of coral reef fisheries to a loss of structural complexity. Curr. Biol. 2014, 24, 1000–1005. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.A.J.; Robinson, J.P.W.; Smith, S.E.; Govinden, R.; Gendron, G.; Wilson, S.K. Changing role of coral reef marine reserves in a warming climate. Nat. Commun. 2020, 11, 2000. [Google Scholar] [CrossRef]
- Perry, C.T.; Morgan, K.M. Bleaching drives collapse in reef carbonate budgets and reef growth potential on southern Maldives reefs. Sci. Rep. 2017, 7, 40581. [Google Scholar] [CrossRef]
- Perry, C.T.; Morgan, K.M. Post-bleaching coral community change on southern Maldivian reefs: Is there potential for rapid recovery? Coral Reefs 2017, 36, 1189–1194. [Google Scholar] [CrossRef]
- Leggat, W.P.; Camp, E.F.; Suggett, D.J.; Heron, S.F.; Fordyce, A.J.; Gardner, S.; Deakin, L.; Turner, M.; Beeching, L.J.; Kuzhiumparambil, U.; et al. Rapid Coral Decay is Associated with Marine Heatwave Mortality Events on Reefs. Curr. Biol. 2019, 29, 2723–2730. [Google Scholar] [CrossRef]
- Perry, C.T.; Morgan, K.M.; Lange, I.D.; Yarlett, R.T. Bleaching-driven reef community shifts drive pulses of increased reef sediment generation. R. Soc. Open Sci. 2020, 7, 192153. [Google Scholar] [CrossRef]
- Newton, K.; Côté, I.M.; Pilling, G.M.; Jennings, S.; Dulvy, N.K. Current and Future Sustainability of Island Coral Reef Fisheries. Curr. Biol. 2007, 17, 655–658. [Google Scholar] [CrossRef]
- Aswani, S.; Sabetian, A. Implications of Urbanization for Artisanal Parrotfish Fisheries in the Western Solomon Islands. Conserv. Biol. 2010, 24, 520–530. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hoey, A.; Hughes, T.P. Human activity selectively impacts the ecosystem roles of parrotfishes on coral reefs. Proc. R. Soc. B 2012, 279, 1621–1629. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, R.J.; Almany, G.R.; Stevens, D.; Bode, M.; Pita, J.; Peterson, N.A.; Choat, J.H. Hyperstability masks declines in bumphead parrotfish (Bolbometopon muricatum) populations. Coral Reefs 2016, 35, 751–763. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species with Missing Data | Data Assumption |
|---|---|
| Chlorurus enneacanthus | Chlorurus sordidus |
| Scarus tricolor | Scarus niger |
| Scarus scaber | Scarus frenatus |
| Scarus prasiognathos | Scarus frenatus |
| Scarus viridifucatus | Scarus frenatus |
| Scarus russelii | Scarus frenatus |
| Hipposcarus harid | Scarus frenatus |
| Cetoscarus ocellatus | Chlorurus strongylocephalus |
| Juveniles | Lowest measured bioerosion rate at <15 cm |
| Species | Size Class | N | Mean | SE |
|---|---|---|---|---|
| Chlorurus sordidus | <15 cm | 13 | 0.03 | 0.01 |
| 16 to 30 cm | 22 | 0.05 | 0.01 | |
| 31 to 45 cm | 7 | 0.19 | 0.05 | |
| >46 cm | N/A | N/A | N/A | |
| Chlorurus strongylocephalus | <15 cm | 13 | 0.03 | 0.01 |
| 16 to 30 cm | 19 | 0.17 | 0.03 | |
| 31 to 45 cm | 12 | 0.57 | 0.11 | |
| >46 cm | 12 | 0.88 | 0.17 | |
| Scarus frenatus | <15 cm | 6 | 0.02 | 0.01 |
| 16 to 30 cm | 11 | 0.04 | 0.01 | |
| 31 to 45 cm | 10 | 0.10 | 0.02 | |
| >46 cm | N/A | N/A | N/A | |
| Scarus niger | <15 cm | 14 | 0.01 | 0.003 |
| 16 to 30 cm | 12 | 0.05 | 0.01 | |
| 31 to 45 cm | 9 | 0.08 | 0.02 | |
| >46 cm | N/A | N/A | N/A | |
| Scarus rubroviolaceus | <15 cm | 4 | 0.01 | 0.003 |
| 16 to 30 cm | 7 | 0.02 | 0.003 | |
| 31 to 45 cm | 12 | 0.08 | 0.03 | |
| >46 cm | 7 | 0.15 | 0.04 |
| Z1 | Z2 | Z3 | Z4 | Z5 | Z6 | Z7 | Z8 | |
|---|---|---|---|---|---|---|---|---|
| Total parrotfish bioerosion rate (kg m−2 yr−1) | 0.58 ± 0.07 | 0.46 ± 0.07 | 0.01 ± 0.00 | 0.50 ± 0.11 | 0.84 ± 0.12 | 0.08 ± 0.01 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Morning bioerosion (% of total rate) | 41 | 37 | 62 | 31 | 77 | 53 | N/A | N/A |
| Midday bioerosion (% of total rate) | 6 | 16 | 12 | 4 | 21 | 4 | N/A | N/A |
| Afternoon bioerosion (% of total rate) | 53 | 47 | 27 | 65 | 2 | 43 | N/A | N/A |
| Total parrotfish habitat bioerosion (kg yr−1) | 8413 ± 1069 | 23,872 ± 3400 | 280 ± 50 | 34,143 ± 7634 | 80,503 ± 11122 | 6351 ± 789 | 0 ± 0 | 0 ± 0 |
| % of total platform parrotfish bioerosion | 5.48 | 15.55 | 0.18 | 22.23 | 52.42 | 4.14 | 0.00 | 0.00 |
| % platform area | 1.74 | 6.19 | 6.53 | 8.25 | 11.56 | 9.68 | 28.93 | 22.12 |
| Z1 | Z2 | Z3 | Z4 | Z5 | Z6 | Z7 | Z8 | |
|---|---|---|---|---|---|---|---|---|
| Habitat Surface Area (m2) (Area × Rugosity) | 32,449 | 102,233 | 59,367 | 92,150 | 159,990 | 107,405 | 255,725 | 191,750 |
| Substrate available for feeding % | 42.84 | 40.87 | 21.31 | 61.89 | 80.02 | 62.38 | N/A | N/A |
| Total area grazed by parrotfish (m2 yr−1) | 18,112 ± 1190 | 110,218 ± 6864 | 3826 ± 465 | 74,054 ± 5283 | 72,722 ± 5531 | 75,696 ± 4689 | 0 ± 0 | 0 ± 0 |
| Morning grazing (% of total rate) | 25 | 38 | 67 | 43 | 57 | 52 | N/A | N/A |
| Midday grazing (% of total rate) | 18 | 17 | 11 | 15 | 12 | 8 | N/A | N/A |
| Afternoon grazing (% of total rate) | 57 | 45 | 22 | 42 | 31 | 40 | N/A | N/A |
| % of habitat area grazed by parrotfish per year | 130 ± 9 | 264 ± 16 | 30 ± 4 | 130 ± 9 | 57 ± 4 | 113 ± 7 | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yarlett, R.T.; Perry, C.T.; Wilson, R.W.; Harborne, A.R. Inter-Habitat Variability in Parrotfish Bioerosion Rates and Grazing Pressure on an Indian Ocean Reef Platform. Diversity 2020, 12, 381. https://doi.org/10.3390/d12100381
Yarlett RT, Perry CT, Wilson RW, Harborne AR. Inter-Habitat Variability in Parrotfish Bioerosion Rates and Grazing Pressure on an Indian Ocean Reef Platform. Diversity. 2020; 12(10):381. https://doi.org/10.3390/d12100381
Chicago/Turabian StyleYarlett, Robert T., Chris T. Perry, Rod W. Wilson, and Alastair R. Harborne. 2020. "Inter-Habitat Variability in Parrotfish Bioerosion Rates and Grazing Pressure on an Indian Ocean Reef Platform" Diversity 12, no. 10: 381. https://doi.org/10.3390/d12100381
APA StyleYarlett, R. T., Perry, C. T., Wilson, R. W., & Harborne, A. R. (2020). Inter-Habitat Variability in Parrotfish Bioerosion Rates and Grazing Pressure on an Indian Ocean Reef Platform. Diversity, 12(10), 381. https://doi.org/10.3390/d12100381