
Vitamin E in Plants: Biosynthesis Pathways, Biofortification Strategies, and Regulatory Dynamics


Abstract
1. Introduction
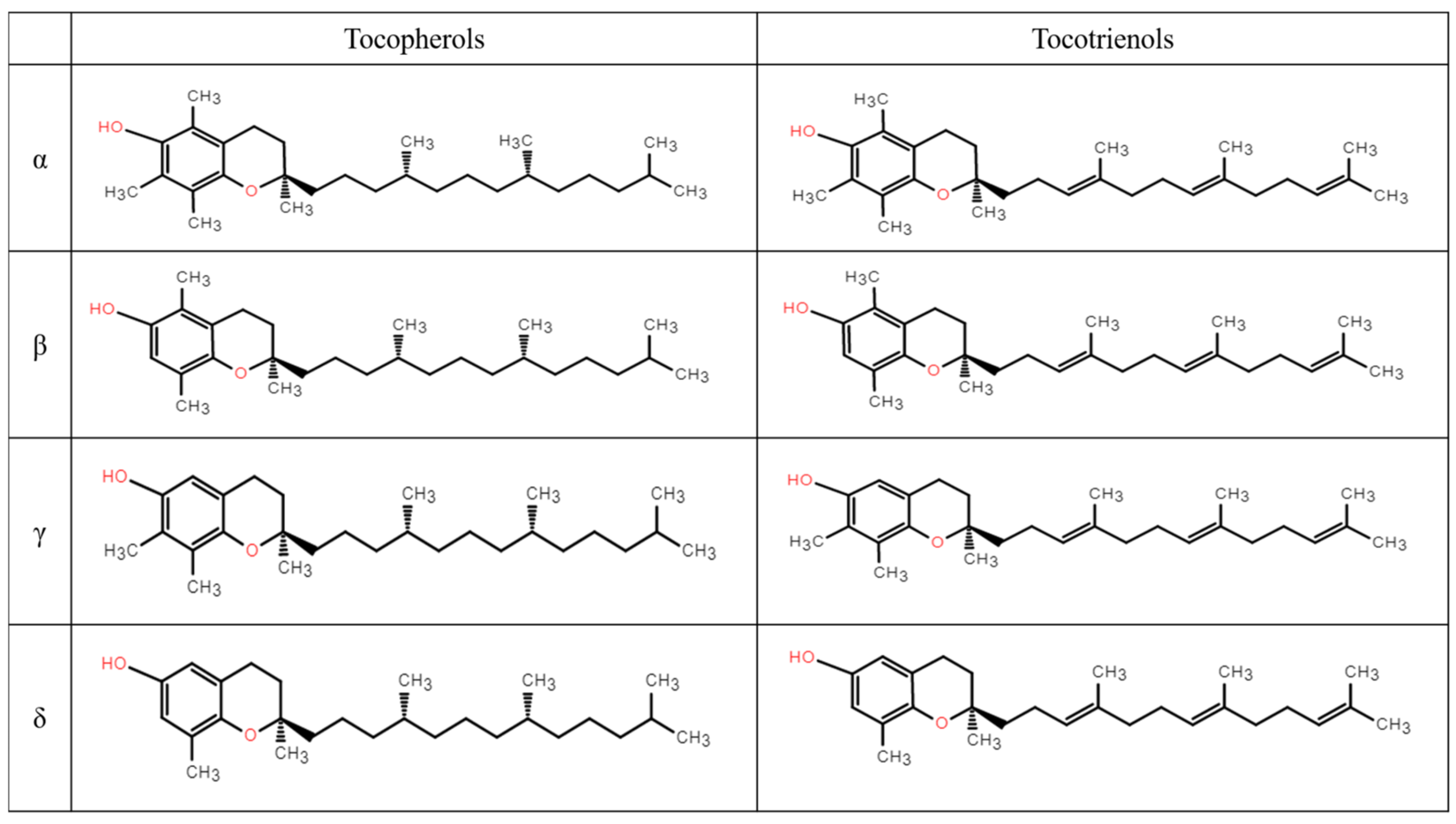
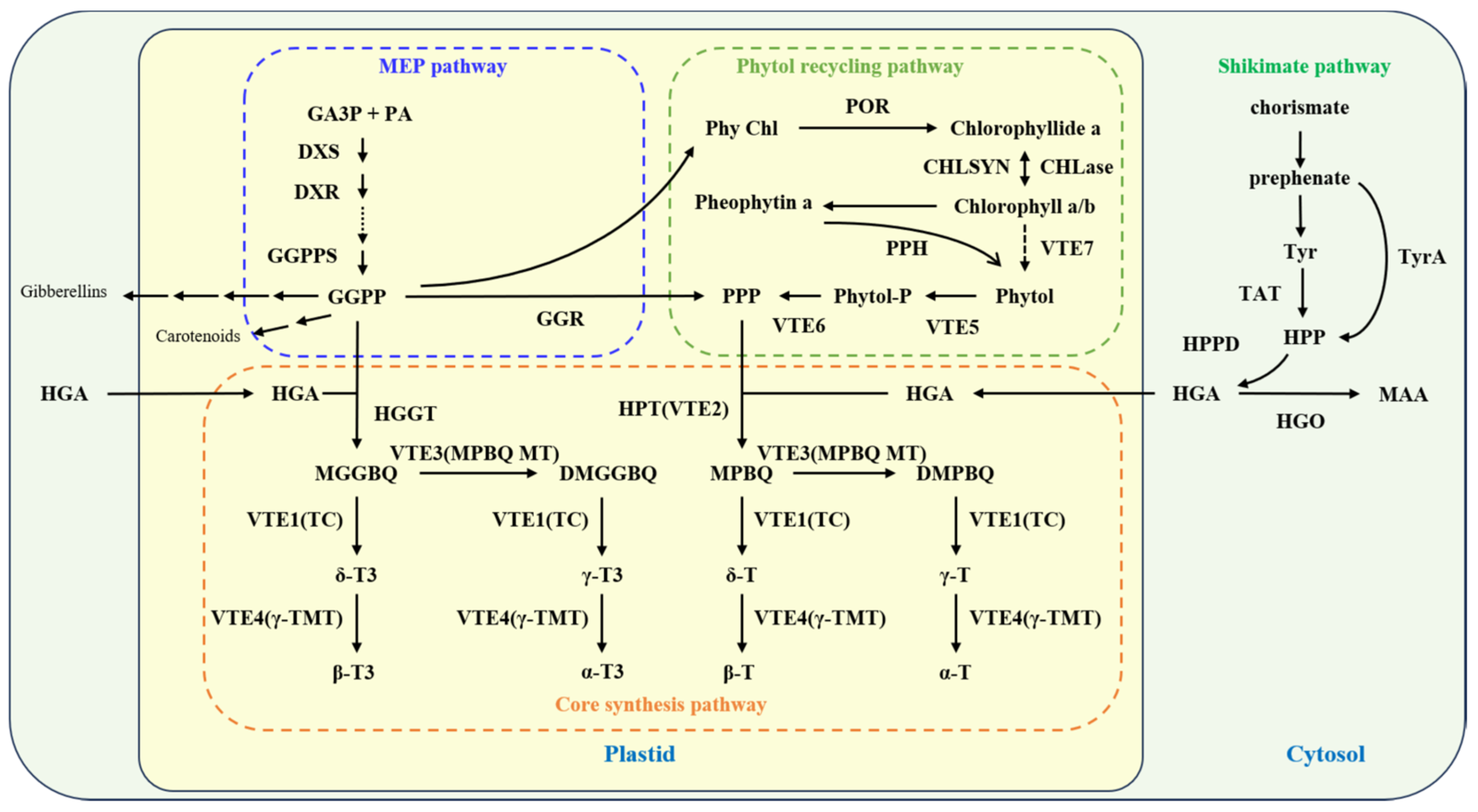
2. The Synthetic Pathway of Vitamin E
3. Genetic and Regulatory Analysis of Vitamin E Synthesis
4. Biofortification of Vitamin E
4.1. Addition of Vitamin E Precursors to Cell Cultures
4.2. Overexpression of Genes Involved in Vitamin E Biosynthetic Pathway in Plants
4.2.1. Boosting the Synthesis of HGA
4.2.2. Genetic Enhancement of GGPP and PPP Contents
4.2.3. Overexpression of Rate-Limiting Enzymes HPT and HGGT
4.2.4. Increase the Content of Highly Active α-Tocopherol
5. Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Evans, H.M.; Bishop, K.S. On the existence of a hitherto unrecognized dietary factor essential for reproduction. Science 1922, 56, 650–651. [Google Scholar] [CrossRef] [PubMed]
- Sussmann, R.A.; Angeli, C.B.; Peres, V.J.; Kimura, E.A.; Katzin, A.M. Intraerythrocytic stages of Plasmodium falciparum biosynthesize vitamin E. FEBS Lett. 2011, 585, 3985–3991. [Google Scholar] [CrossRef] [PubMed]
- Mène-Saffrané, L.; Pellaud, S. Current strategies for vitamin E biofortification of crops. Curr. Opin. Biotechnol. 2017, 44, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Kyndt, T.; Hancock, R.D. Vitamin C in Plants: Novel Concepts, New Perspectives, and Outstanding Issues. Antioxid. Redox Signal. 2020, 32, 463–485. [Google Scholar] [CrossRef]
- Che, P.; Zhao, Z.Y.; Glassman, K.; Dolde, D.; Hu, T.X.; Jones, T.J.; Gruis, D.F.; Obukosia, S.; Wambugu, F.; Albertsen, M.C. Elevated vitamin E content improves all-trans beta-carotene accumulation and stability in biofortified sorghum. Proc. Natl. Acad. Sci. USA 2016, 113, 11040–11045. [Google Scholar] [CrossRef]
- FRYER, M.J. The antioxidant effects of thylakoid Vitamin E (α-tocopherol). Plant Cell Environ. 1992, 15, 381–392. [Google Scholar] [CrossRef]
- Salimath, S.S.; Romsdahl, T.B.; Konda, A.R.; Zhang, W.; Cahoon, E.B.; Dowd, M.K.; Wedegaertner, T.C.; Hake, K.D.; Chapman, K.D. Production of tocotrienols in seeds of cotton (Gossypium hirsutum L.) enhances oxidative stability and offers nutraceutical potential. Plant Biotechnol. J. 2021, 19, 1268–1282. [Google Scholar] [CrossRef]
- Elstner, E.F. Oxygen radicals—biochemical basis for their efficacy. Klin. Wochenschr. 1991, 69, 949–956. [Google Scholar] [CrossRef]
- Kamal-Eldin, A.; Appelqvist, L.A. The chemistry and antioxidant properties of tocopherols and tocotrienols. Lipids 1996, 31, 671–701. [Google Scholar] [CrossRef]
- Matringe, M.; Ksas, B.; Rey, P.; Havaux, M. Tocotrienols, the unsaturated forms of vitamin E, can function as antioxidants and lipid protectors in tobacco leaves. Plant Physiol. 2008, 147, 764–778. [Google Scholar] [CrossRef]
- Muñoz, P.; Munné-Bosch, S. Vitamin E in Plants: Biosynthesis, Transport, and Function. Trends Plant Sci. 2019, 24, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Ali, E.; Hussain, S.; Hussain, N.; Kakar, K.U.; Shah, J.M.; Zaidi, S.H.R.; Jan, M.; Zhang, K.; Khan, M.A.; Imtiaz, M. Tocopherol as plant protector: An overview of Tocopherol biosynthesis enzymes and their role as antioxidant and signaling molecules. Acta Physiol. Plant. 2022, 44, 1–11. [Google Scholar] [CrossRef]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef]
- Lan, D.; Yao, C.; Li, X.; Liu, H.; Wang, D.; Wang, Y.; Qi, S. Tocopherol attenuates the oxidative stress of BMSCs by inhibiting ferroptosis through the PI3k/AKT/mTOR pathway. Front. Bioeng. Biotechnol. 2022, 10, 938520. [Google Scholar] [CrossRef]
- Eitsuka, T.; Nakagawa, K.; Miyazawa, T. Down-regulation of telomerase activity in DLD-1 human colorectal adenocarcinoma cells by tocotrienol. Biochem. Biophys. Res. Commun. 2006, 348, 170–175. [Google Scholar]
- He, L.; Mo, H.; Hadisusilo, S.; Qureshi, A.A.; Elson, C.E. Isoprenoids suppress the growth of murine B16 melanomas in vitro and in vivo. J. Nutr. 1997, 127, 668–674. [Google Scholar]
- Khanna, S.; Roy, S.; Ryu, H.; Bahadduri, P.; Swaan, P.W.; Ratan, R.R.; Sen, C.K. Molecular basis of vitamin E action: Tocotrienol modulates 12-lipoxygenase, a key mediator of glutamate-induced neurodegeneration. J. Biol. Chem. 2003, 278, 43508–43515. [Google Scholar]
- Qureshi, A.A.; Peterson, D.M.; Hasler-Rapacz, J.O.; Rapacz, J. Novel tocotrienols of rice bran suppress cholesterogenesis in hereditary hypercholesterolemic swine. J. Nutr. 2001, 131, 223–230. [Google Scholar]
- Qureshi, A.A.; Sami, S.A.; Salser, W.A.; Khan, F.A. Synergistic effect of tocotrienol-rich fraction (TRF25) of rice bran and lovastatin on lipid parameters in hypercholesterolemic humans. J. Nutr. Biochem. 2001, 12, 318–329. [Google Scholar]
- Sen, C.K.; Khanna, S.; Roy, S.; Packer, L. Molecular basis of vitamin E action. Tocotrienol potently inhibits glutamate-induced pp60c-Src kinase activation and death of HT4 neuronal cells. J. Biol. Chem. 2000, 275, 13049–13055. [Google Scholar]
- Serbinova, E.; Kagan, V.; Han, D.; Packer, L. Free radical recycling and intramembrane mobility in the antioxidant properties of alpha-tocopherol and alpha-tocotrienol. Free Radic. Biol. Med. 1991, 10, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Serbinova, E.; Khwaja, S.; Reznick, A.Z.; Packer, L. Thioctic acid protects against ischemia-reperfusion injury in the isolated perfused langendorff heart. Free Radic. Res. Commun. 1992, 17, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.M.; Bhuvanendran, S.; Nair, R.S.; Radhakrishnan, A.K. Exploring the anti-inflammatory activities, mechanism of action and prospective drug delivery systems of tocotrienol to target neurodegenerative diseases. F1000 Res. 2023, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Raptova, R.; Alomar, S.Y.; Alwasel, S.H.; Nepovimova, E.; Kuca, K.; Valko, M. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: Chronic diseases and aging. Arch. Toxicol. 2023, 97, 2499–2574. [Google Scholar] [CrossRef]
- National Feed Work Office CFIA. China Feed Industry Yearbook 2006/2007; China Commercial Press: Beijing, China, 2008. [Google Scholar]
- National Feed Work Office CFIA. China Feed Industry Yearbook 2008; China Commercial Press: Beijing, China, 2009. [Google Scholar]
- Hunter, S.C.; Cahoon, E.B. Enhancing vitamin E in oilseeds: Unraveling tocopherol and tocotrienol biosynthesis. Lipids 2007, 42, 97–108. [Google Scholar] [CrossRef]
- Kim, Y.N.; Cho, Y.O. Vitamin E status of 20-to 59-year-old adults living in the Seoul metropolitan area of South Korea. Nutr. Res. Pract. 2015, 9, 192–198. [Google Scholar] [CrossRef]
- Polito, A.; Intorre, F.; Andriollo-Sanchez, M.; Azzini, E.; Raguzzini, A.; Meunier, N.; Ducros, V.; O’Connor, J.M.; Coudray, C.; Roussel, A.M.; et al. Estimation of intake and status of vitamin A, vitamin E and folate in older European adults: The ZENITH. Eur. J. Clin. Nutr. 2005, 59, S42–S47. [Google Scholar] [CrossRef]
- Chaudhary, R.; Chaturvedi, S.; Sharma, R.; Tiwari, S. Global Scenario of Vitamin Deficiency and Human Health. In Advances in Agri-Food Biotechnology; Sharma, T.R., Deshmukh, R., Sonah, H., Eds.; Springer Singapore: Singapore, 2020; pp. 199–220. [Google Scholar]
- Passarelli, S.; Free, C.M.; Shepon, A.; Beal, T.; Batis, C.; Golden, C.D. Global estimation of dietary micronutrient inadequacies: A modelling analysis. Lancet Glob. Health 2024, 12, e1590–e1599. [Google Scholar] [CrossRef]
- Peter, S.; Friedel, A.; Roos, F.F.; Wyss, A.; Eggersdorfer, M.; Hoffmann, K.; Weber, P. A Systematic Review of Global Alpha-Tocopherol Status as Assessed by Nutritional Intake Levels and Blood Serum Concentrations. J. Int. Vitaminol. Nutr. 2015, 85, 261–281. [Google Scholar] [CrossRef]
- Society, C.N. Reference Intake of Dietary Nutrients for Chinese Residents in 2023. J. Nutr. 2023, 45, 414. [Google Scholar] [CrossRef]
- Borel, P.; Preveraud, D.; Desmarchelier, C. Bioavailability of vitamin E in humans: An update. Nutr. Rev. 2013, 71, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q. Metabolism of natural forms of vitamin E and biological actions of vitamin E metabolites. Free. Radic. Biol. Med. 2022, 179, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Albert, E.; Kim, S.; Magallanes-Lundback, M.; Bao, Y.; Deason, N.; Danilo, B.; Wu, D.; Li, X.; Wood, J.C.; Bornowski, N.; et al. Genome-wide association identifies a missing hydrolase for tocopherol synthesis in plants. Proc. Natl. Acad. Sci. USA 2022, 119, e2113488119. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhang, Q.; Wang, J.; Li, Y.; Wang, X.; Bao, Y. Vitamin E synthesis and response in plants. Front. Plant Sci. 2022, 13, 994058. [Google Scholar] [CrossRef]
- Herbers, K. Vitamin production in transgenic plants. J. Plant Physiol. 2003, 160, 821–829. [Google Scholar]
- Wang, H.; Xu, S.; Fan, Y.; Liu, N.; Zhan, W.; Liu, H.; Xiao, Y.; Li, K.; Pan, Q.; Li, W.; et al. Beyond pathways: Genetic dissection of tocopherol content in maize kernels by combining linkage and association analyses. Plant Biotechnol. J. 2018, 16, 1464–1475. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, W.; Ren, G.; Li, D.; Cahoon, R.E.; Chen, M.; Zhou, Y.; Yu, B.; Cahoon, E.B. Chlorophyll Synthase under Epigenetic Surveillance Is Critical for Vitamin E Synthesis, and Altered Expression Affects Tocopherol Levels in Arabidopsis. Plant Physiol. 2015, 168, 1503–1511. [Google Scholar] [CrossRef]
- Ruiz-Sola, M.Á.; Coman, D.; Beck, G.; Barja, M.V.; Colinas, M.; Graf, A.; Welsch, R.; Rütimann, P.; Bühlmann, P.; Bigler, L.; et al. Arabidopsis GERANYLGERANYL DIPHOSPHATE SYNTHASE 11 is a hub isozyme required for the production of most photosynthesis-related isoprenoids. New Phytol. 2016, 209, 252–264. [Google Scholar] [CrossRef]
- Tanaka, R.; Rothbart, M.; Oka, S.; Takabayashi, A.; Takahashi, K.; Shibata, M.; Myouga, F.; Motohashi, R.; Shinozaki, K.; Grimm, B.; et al. LIL3, a light-harvesting-like protein, plays an essential role in chlorophyll and tocopherol biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 16721–16725. [Google Scholar] [CrossRef]
- Qin, P.; Chen, P.; Zhou, Y.; Zhang, W.; Zhang, Y.; Xu, J.; Gan, L.; Liu, Y.; Romer, J.; Dörmann, P.; et al. Vitamin E biofortification: Enhancement of seed tocopherol concentrations by altered chlorophyll metabolism. Front. Plant Sci. 2024, 15, 1344095. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, T.; Ren, G.; Hörtensteiner, S.; Zhou, Y.; Cahoon, E.B.; Zhang, C. Chlorophyll degradation: The tocopherol biosynthesis-related phytol hydrolase in Arabidopsis seeds is still missing. Plant Physiol. 2014, 166, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Georgiadou, E.C.; Ntourou, T.; Goulas, V.; Manganaris, G.A.; Kalaitzis, P.; Fotopoulos, V. Temporal analysis reveals a key role for VTE5 in vitamin E biosynthesis in olive fruit during on-tree development. Front. Plant Sci. 2015, 6, 871. [Google Scholar] [CrossRef]
- Almeida, J.; Azevedo, S.; Spicher, L.; Glauser, G.; vom Dorp, K.; Guyer, L.; del Valle Carranza, A.; Asis, R.; de Souza, A.P.; Buckeridge, M.; et al. Down-regulation of tomato PHYTOL KINASE strongly impairs tocopherol biosynthesis and affects prenyllipid metabolism in an organ-specific manner. J. Exp. Bot. 2016, 67, 919–934. [Google Scholar] [CrossRef] [PubMed]
- Vom Dorp, K.; Hölzl, G.; Plohmann, C.; Eisenhut, M.; Abraham, M.; Weber, A.P.; Hanson, A.D.; Dörmann, P. Remobilization of phytol from chlorophyll degradation is essential for tocopherol synthesis and growth of Arabidopsis. Plant Cell 2015, 27, 2846–2859. [Google Scholar] [CrossRef]
- Diepenbrock, C.H.; Kandianis, C.B.; Lipka, A.E.; Magallanes-Lundback, M.; Vaillancourt, B.; Gongora-Castillo, E.; Wallace, J.G.; Cepela, J.; Mesberg, A.; Bradbury, P.J.; et al. Novel loci underlie natural variation in Vitamin E levels in maize grain. Plant Cell 2017, 29, 2374–2392. [Google Scholar] [CrossRef]
- Liu, N.; Du, Y.; Yan, S.; Chen, W.; Deng, M.; Xu, S.; Wang, H.; Zhan, W.; Huang, W.; Yin, Y.; et al. The light and hypoxia induced gene ZmPORB1 determines tocopherol content in the maize kernel. Sci. China Life Sci. 2024, 67, 435–448. [Google Scholar] [CrossRef]
- Zhan, W.; Liu, J.; Pan, Q.; Wang, H.; Yan, S.; Li, K.; Deng, M.; Li, W.; Liu, N.; Kong, Q.; et al. An allele of ZmPORB2 encoding a protochlorophyllide oxidoreductase promotes tocopherol accumulation in both leaves and kernels of maize. Plant J. Cell Mol. Biol. 2019, 100, 114–127. [Google Scholar] [CrossRef]
- Muthulakshmi, M.; Srinivasan, A.; Srivastava, S. Antioxidant Green Factories: Toward Sustainable Production of Vitamin E in Plant In Vitro Cultures. ACS Omega 2023, 8, 3586–3605. [Google Scholar] [CrossRef]
- Caretto, S.; Bray, S.; Fachechi, C.; Gala, R.; Zacheo, G.; Giovinazzo, G. Enhancement of vitamin E production in sunflower cell cultures. Plant Cell Rep. 2004, 23, 174–179. [Google Scholar] [CrossRef]
- Furuya, T.; Yoshikawa, T.; Kimura, T.; Kaneko, H. Production of tocopherols by cell culture of safflower. Phytochemistry 1987, 26, 2741–2747. [Google Scholar] [CrossRef]
- Karunanandaa, B.; Qi, Q.; Hao, M.; Baszis, S.R.; Jensen, P.K.; Wong, Y.-H.H.; Jiang, J.; Venkatramesh, M.; Gruys, K.J.; Moshiri, F.; et al. Metabolically engineered oilseed crops with enhanced seed tocopherol. Metab. Eng. 2005, 7, 384–400. [Google Scholar] [CrossRef] [PubMed]
- Tsegaye, Y.; Shintani, D.K.; DellaPenna, D. Overexpression of the enzyme p-hydroxyphenolpyruvate dioxygenase in Arabidopsis and its relation to tocopherol biosynthesis. Plant Physiol. Biochem. 2002, 40, 913–920. [Google Scholar] [CrossRef]
- Zhang, C.; Cahoon, R.E.; Hunter, S.C.; Chen, M.; Han, J.; Cahoon, E.B. Genetic and biochemical basis for alternative routes of tocotrienol biosynthesis for enhanced vitamin E antioxidant production. Plant J. Cell Mol. Biol. 2013, 73, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Falk, J.; Andersen, G.; Kernebeck, B.; Krupinska, K. Constitutive overexpression of barley 4-hydroxyphenylpyruvate dioxygenase in tobacco results in elevation of the vitamin E content in seeds but not in leaves. FEBS Lett. 2003, 540, 35–40. [Google Scholar] [CrossRef]
- Rippert, P.; Scimemi, C.; Dubald, M.; Matringe, M. Engineering plant shikimate pathway for production of tocotrienol and improving herbicide resistance. Plant Physiol. 2004, 134, 92–100. [Google Scholar] [CrossRef]
- Tzin, V.; Galili, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Stacey, M.G.; Cahoon, R.E.; Nguyen, H.T.; Cui, Y.; Sato, S.; Nguyen, C.T.; Phoka, N.; Clark, K.M.; Liang, Y.; Forrester, J.; et al. Identification of Homogentisate Dioxygenase as a Target for Vitamin E Biofortification in Oilseeds. Plant Physiol. 2016, 172, 1506–1518. [Google Scholar] [CrossRef]
- Konda, A.R.; Gelli, M.; Pedersen, C.; Cahoon, R.E.; Zhang, C.; Obata, T.; Cahoon, E.B. Vitamin E biofortification: Maximizing oilseed tocotrienol and total vitamin E tocochromanol production by use of metabolic bypass combinations. Metab. Eng. 2023, 79, 66–77. [Google Scholar] [CrossRef]
- Estévez, J.M.; Cantero, A.; Reindl, A.; Reichler, S.; León, P. 1-Deoxy-d-xylulose-5-phosphate synthase, a limiting enzyme for plastidic isoprenoid biosynthesis in plants. J. Biol. Chem. 2001, 276, 22901–22909. [Google Scholar] [CrossRef]
- Wright, L.P.; Rohwer, J.M.; Ghirardo, A.; Hammerbacher, A.; Ortiz-Alcaide, M.; Raguschke, B.; Schnitzler, J.-P.; Gershenzon, J.; Phillips, M.A. Deoxyxylulose 5-phosphate synthase controls flux through the methylerythritol 4-phosphate pathway in Arabidopsis thaliana. Plant Physiol. 2014, 165, 1488–1504. [Google Scholar] [CrossRef]
- Carretero-Paulet, L.; Cairó, A.; Botella-Pavía, P.; Besumbes, O.; Campos, N.; Boronat, A.; Rodríguez-Concepción, M. Enhanced flux through the methylerythritol 4-phosphate pathway in Arabidopsis plants overexpressing deoxyxylulose 5-phosphate reductoisomerase. Plant Mol. Biol. 2006, 62, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Kimura, E.; Abe, T.; Murata, K.; Kimura, T.; Otoki, Y.; Yoshida, T.; Miyazawa, T.; Nakagawa, K. Identification of OsGGR2, a second geranylgeranyl reductase involved in α-tocopherol synthesis in rice. Sci. Rep. 2018, 8, 1870. [Google Scholar] [CrossRef] [PubMed]
- Valentin, H.E.; Lincoln, K.; Moshiri, F.; Jensen, P.K.; Qi, Q.; Venkatesh, T.V.; Karunanandaa, B.; Baszis, S.R.; Norris, S.R.; Savidge, B.; et al. The Arabidopsis vitamin E pathway gene5-1 mutant reveals a critical role for phytol kinase in seed tocopherol biosynthesis. Plant Cell 2006, 18, 212–224. [Google Scholar] [PubMed]
- Harish, M.C.; Dachinamoorthy, P.; Balamurugan, S.; Bala Murugan, S.; Sathishkumar, R. Overexpression of homogentisate phytyltransferase (HPT) and tocopherol cyclase (TC) enhances α-tocopherol content in transgenic tobacco. Biol. Plant. 2013, 57, 395–400. [Google Scholar] [CrossRef]
- Collakova, E.; DellaPenna, D. The role of homogentisate phytyltransferase and other tocopherol pathway enzymes in the regulation of tocopherol synthesis during abiotic stress. Plant Physiol. 2003, 133, 930–940. [Google Scholar] [CrossRef]
- Lee, K.; Lee, S.M.; Park, S.R.; Jung, J.; Moon, J.K.; Cheong, J.J.; Kim, M. Overexpression of Arabidopsis homogentisate phytyltransferase or tocopherol cyclase elevates vitamin E content by increasing gamma-tocopherol level in lettuce (Lactuca sativa L.). Mol. Cells 2007, 24, 301–306. [Google Scholar]
- Sundararajan, S.; Rajendran, V.; Sivakumar, H.P.; Nayeem, S.; Mani Chandra, H.; Sharma, A.; Ramalingam, S. Enhanced vitamin E content in an Indica rice cultivar harbouring two transgenes from Arabidopsis thaliana involved in tocopherol biosynthesis pathway. Plant Biotechnol. J. 2021, 19, 1083–1085. [Google Scholar] [CrossRef]
- Upadhyaya, D.C.; Bagri, D.S.; Upadhyaya, C.P.; Kumar, A.; Thiruvengadam, M.; Jain, S.K. Genetic engineering of potato (Solanum tuberosum L.) for enhanced α-tocopherols and abiotic stress tolerance. Physiol. Plant. 2021, 173, 116–128. [Google Scholar] [CrossRef]
- Ren, W.; Zhao, L.; Zhang, L.; Wang, Y.; Cui, L.; Tang, Y.; Sun, X.; Tang, K. Molecular analysis of a homogentisate phytyltransferase gene from Lactuca sativa L. Mol. Biol. Rep. 2011, 38, 1813–1819. [Google Scholar] [CrossRef]
- Seo, Y.S.; Kim, S.J.; Harn, C.H.; Kim, W.T. Ectopic expression of apple fruit homogentisate phytyltransferase gene (MdHPT1) increases tocopherol in transgenic tomato (Solanum lycopersicum cv. Micro-Tom) leaves and fruits. Phytochemistry 2011, 72, 321–329. [Google Scholar] [CrossRef]
- Chen, J.; Liu, C.; Shi, B.; Chai, Y.; Han, N.; Zhu, M.; Bian, H. Overexpression of HvHGGT enhances tocotrienol levels and antioxidant activity in barley. J. Agric. Food Chem. 2017, 65, 5181–5187. [Google Scholar] [CrossRef] [PubMed]
- Cahoon, E.B.; Hall, S.E.; Ripp, K.G.; Ganzke, T.S.; Hitz, W.D.; Coughlan, S.J. Metabolic redesign of vitamin E biosynthesis in plants for tocotrienol production and increased antioxidant content. Nat. Biotechnol. 2003, 21, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Konda, A.R.; Nazarenus, T.J.; Nguyen, H.; Yang, J.; Gelli, M.; Swenson, S.; Shipp, J.M.; Schmidt, M.A.; Cahoon, R.E.; Ciftci, O.N.; et al. Metabolic engineering of soybean seeds for enhanced vitamin E tocochromanol content and effects on oil antioxidant properties in polyunsaturated fatty acid-rich germplasm. Metab. Eng. Commun. 2020, 57, 63–73. [Google Scholar] [CrossRef]
- Tanaka, H.; Yabuta, Y.; Tamoi, M.; Tanabe, N.; Shigeoka, S. Generation of transgenic tobacco plants with enhanced tocotrienol levels through the ectopic expression of rice homogentisate geranylgeranyl transferase. Plant Biotechnol. 2015, 32, 233–238. [Google Scholar] [CrossRef]
- Dolde, D.; Wang, T. Oxidation of Crude Corn Oil with and without Elevated Tocotrienols. J. Am. Oil Chem. Soc. 2011, 88, 1367–1372. [Google Scholar] [CrossRef]
- Shintani, D.; DellaPenna, D. Elevating the vitamin E content of plants through metabolic engineering. Science 1998, 282, 2098–2100. [Google Scholar] [CrossRef]
- Cho, E.A.; Lee, C.A.; Kim, Y.S.; Baek, S.H.; de los Reyes, B.G.; Yun, S.J. Expression of gamma-tocopherol methyltransferase transgene improves tocopherol composition in lettuce (Latuca sativa L.). Mol. Cells 2005, 19, 16–22. [Google Scholar] [CrossRef]
- Jin, S.; Daniell, H. Expression of γ-tocopherol methyltransferase in chloroplasts results in massive proliferation of the inner envelope membrane and decreases susceptibility to salt and metal-induced oxidative stresses by reducing reactive oxygen species. Plant Biotechnol. J. 2014, 12, 1274–1285. [Google Scholar] [CrossRef]
- Yusuf, M.A.; Sarin, N.B. Antioxidant value addition in human diets: Genetic transformation of Brassica juncea with gamma-TMT gene for increased alpha-tocopherol content. Transgenic Res. 2007, 16, 109–113. [Google Scholar] [CrossRef]
- Chen, D.F.; Zhang, M.; Wang, Y.Q.; Chen, X.W. Expression of γ-tocopherol methyltransferase gene from Brassica napus increased α-tocopherol content in soybean seed. Biol. Plant. 2012, 56, 131–134. [Google Scholar] [CrossRef]
- Youngjin, K.; HongYul, S.; TaeIl, P.; SoHyeon, B.; WoonChul, S.; Hyunsoon, K.; Junggon, K.; YongEui, C.; SongJoong, Y. Enhanced biosynthesis of α-tocopherol in transgenic soybean by introducing γ-TMT gene. Plant Biotechnol. J. 2005, 7, 1–7. [Google Scholar]
- Tavva, V.S.; Kim, Y.H.; Kagan, I.A.; Dinkins, R.D.; Kim, K.H.; Collins, G.B. Increased alpha-tocopherol content in soybean seed overexpressing the Perilla frutescens gamma-tocopherol methyltransferase gene. Plant Cell Rep. 2007, 26, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Jia, H.; Feng, G.; Wang, Z.; Li, J.; Gao, H.; Wang, X. Overexpression of Medicago sativa TMT elevates the α-tocopherol content in Arabidopsis seeds, alfalfa leaves, and delays dark-induced leaf senescence. Plant Sci. 2016, 249, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Qiu, D.; Gao, H.; Wen, H.; Wu, Y.; Pang, Y.; Wang, X.; Qin, Y. Over-expression of a γ-tocopherol methyltransferase gene in vitamin E pathway confers PEG-simulated drought tolerance in alfalfa. BMC Plant Biol. 2020, 20, 226. [Google Scholar] [CrossRef]
- Zhang, L.; Luo, Y.; Liu, B.; Zhang, L.; Zhang, W.; Chen, R.; Wang, L. Overexpression of the maize γ-tocopherol methyltransferase gene (ZmTMT) increases α-tocopherol content in transgenic Arabidopsis and maize seeds. Transgenic Res. 2020, 29, 95–104. [Google Scholar] [CrossRef]
- Zhang, G.Y.; Liu, R.R.; Xu, G.; Zhang, P.; Li, Y.; Tang, K.X.; Liang, G.H.; Liu, Q.Q. Increased α-tocotrienol content in seeds of transgenic rice overexpressing Arabidopsis γ-tocopherol methyltransferase. Transgenic res 2013, 22, 89–99. [Google Scholar] [CrossRef]
- YAO, X.; Yang, W.; LUO, Y.; CHEN, R.; WANG, L.; ZHANG, L. Acquisition and Characteristic Analysis of Transgenic Maize with phyA2, ZmTMT, and Bar. Sci. Agric. Sin. 2020, 53, 4982–4991. [Google Scholar]
- Van Eenennaam, A.L.; Lincoln, K.; Durrett, T.P.; Valentin, H.E.; Shewmaker, C.K.; Thorne, G.M.; Jiang, J.; Baszis, S.R.; Levering, C.K.; Aasen, E.D.; et al. Engineering vitamin e content: From Arabidopsis mutant to soy oil. Plant Cell 2003, 15, 3007–3019. [Google Scholar] [CrossRef]
- Collakova, E.; DellaPenna, D. Homogentisate phytyltransferase activity is limiting for tocopherol biosynthesis in Arabidopsis. Plant Physiol. 2003, 131, 632–642. [Google Scholar] [CrossRef]
- Meena, V.K.; Chand, S.; Shekhawat, H.V.S.; Choudhary, K.; Sharma, J.K.; Sheera, A.; Lekha; Yadava, D.K. Advances in plant tocopherol biosynthesis: From pathway elucidation to crop biofortification strategies. Discover Plants 2025, 2, 9. [Google Scholar] [CrossRef]
- Lipka, A.E.; Gore, M.A.; Magallanes-Lundback, M.; Mesberg, A.; Lin, H.; Tiede, T.; Chen, C.; Buell, C.R.; Buckler, E.S.; Rocheford, T.; et al. Genome-wide association study and pathway-level analysis of tocochromanol levels in maize grain. G3 Genes Genomes Genet. 2013, 3, 1287–1299. [Google Scholar] [CrossRef]
- Deng, M.; Chen, H.; Zhang, W.; Cahoon, E.B.; Zhou, Y.; Zhang, C. Genetic improvement of tocotrienol content enhances the oxidative stability of canola oil. Front. Plant Sci. 2023, 14, 1247781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Luo, Y.; Zhu, Y.; Zhang, L.; Zhang, W.; Chen, R.; Xu, M.; Fan, Y.; Wang, L. GmTMT2a from soybean elevates the α-tocopherol content in corn and Arabidopsis. Transgenic Res. 2013, 22, 1021–1028. [Google Scholar] [CrossRef] [PubMed]
- DellaPenna, D.; Mène-Saffrané, L. Vitamin E. In Biosynthesis of Vitamins in Plants Part B; Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2011; pp. 179–227. [Google Scholar]
- Li, Q.; Yang, X.; Xu, S.; Cai, Y.; Zhang, D.; Han, Y.; Li, L.; Zhang, Z.; Gao, S.; Li, J.; et al. Genome-wide association studies identified three independent polymorphisms associated with α-tocopherol content in maize kernels. Public Libr. Sci. One 2012, 7, e36807. [Google Scholar] [CrossRef]
- Chu, D.; Zhang, Z.; Hu, Y.; Fang, C.; Xu, X.; Yuan, J.; Zhang, J.; Tian, Z.; Wang, G. Genome-wide scan for oil quality reveals a coregulation mechanism of tocopherols and fatty acids in soybean seeds. Plant Commun. 2023, 4, 100598. [Google Scholar] [CrossRef]
- Maeda, H.; Sage, T.L.; Isaac, G.; Welti, R.; Dellapenna, D. Tocopherols modulate extraplastidic polyunsaturated fatty acid metabolism in Arabidopsis at low temperature. Plant Cell 2008, 20, 452–470. [Google Scholar] [CrossRef]
- Austin, J.R.; Frost, E.; Vidi, P.A.; Kessler, F.; Staehelin, L.A. Plastoglobules are lipoprotein subcompartments of the chloroplast that are permanently coupled to thylakoid membranes and contain biosynthetic enzymes. Plant Cell 2006, 18, 1693–1703. [Google Scholar] [CrossRef]
- Bermúdez, L.; del Pozo, T.; Lira, B.S.; de Godoy, F.; Boos, I.; Romanó, C.; Previtali, V.; Almeida, J.; Bréhélin, C.; Asis, R.; et al. A Tomato Tocopherol Binding Protein Sheds Light on Intracellular α-tocopherol Metabolism in Plants. Plant Cell Physiol. 2018, 59, 2188–2203. [Google Scholar] [CrossRef]
- Wang, M.; Toda, K.; Maeda, H.A. Biochemical properties and subcellular localization of tyrosine aminotransferases in Arabidopsis thaliana. Phytochemistry 2016, 132, 16–25. [Google Scholar] [CrossRef]
- Garcia, I.; Rodgers, M.; Lenne, C.; Rolland, A.; Sailland, A.; Matringe, M. Subcellular localization and purification of a p-hydroxyphenylpyruvate dioxygenase from cultured carrot cells and characterization of the corresponding cDNA. Biochem. J. 1997, 325 Pt 3, 761–769. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Genes Involved in Biofortification | Promoters Type | Target Plant | Biofortification Effect | References |
| Arabidopsis HPPD | constitutive CaMV 35S | Arabidopsis | Tocopherol in leaves increased to 1.15~1.37-fold. Tocopherol in seeds increased to 1.10~1.11-fold. | [59] |
| Arabidopsis HPPD | seed-specific pDC3 | Tocopherol in seeds increased to 1.24~1.28-fold. | ||
| E. coli TyrA | constitutive CaMV 35S | Tocopherol in leaves increased to 2~3-fold. | [98] | |
| Arabidopsis HPPD + E. coli TyrA | Tocopherol in leaves increased to 1.5-fold. | |||
| Hordeum vulgare HPPD | Tobacco | Tocopherol in leaves increased to 2-fold. | [61] | |
| Arabidopsis HPPD | seed-specific napin | Arabidopsis | Tocopherol in seeds increased to 1.09-fold. | [58] |
| E. coli TyrA | Tocopherol in seeds increased to 1.2-fold. | |||
| Arabidopsis VTE2 | Tocopherol in seeds increased to 1.41-fold. | |||
| Arabidopsis HPPD + Arabidopsis VTE2 | Tocopherol in seeds increased to 1.46-fold. | |||
| Arabidopsis HPPD + E. coli TyrA | Tocopherol in seeds increased to 1.8-fold. | |||
| E. coli TyrA + Arabidopsis VTE2 | Tocopherol in seeds increased to 1.8-fold. | |||
| Arabidopsis HPPD + E. coli TyrA + Arabidopsis VTE2 | Vitamin E in seeds increased to 2.88-fold. | |||
| E. coli TyrA | Canola | Tocopherol in seeds increased to 1.6-fold. | ||
| Arabidopsis HPPD + E. coli TyrA | Vitamin E in seeds increased to 2.4-fold. | |||
| Arabidopsis HPPD + E. coli TyrA + Glycine max. VTE2 | Vitamin E in seeds increased to 2-fold. | |||
| E. coli TyrA | Soybean | Tocopherol in seeds increased to 1.1-fold. | ||
| Arabidopsis VTE2 | Tocopherol in seeds increased to 1.16-fold. | |||
| Arabidopsis HPPD + E. coli TyrA | Vitamin E in seeds increased to 2.6-fold. | |||
| Arabidopsis HPPD + E. coli TyrA + Glycine max. VTE2 | Vitamin E in seeds increased to 3.3-fold. | |||
| Arabidopsis HPPD + E. coli TyrA + Arabidopsis VTE2 | Vitamin E in seeds increased to 3-fold. | |||
| Arabidopsis DXS | constitutive CaMV 35S | Arabidopsis | Vitamin E in seeds increased to 2-fold. | [66] |
| Arabidopsis DXS | Vitamin E in leaves of mature plants showed no significant increase. | [67] | ||
| Arabidopsis DXR | Vitamin E showed no significant change. | [68] | ||
| Zea mays PORB2 | Ubi | Maize | Tocopherol in leaves and seeds increased by 1.28~1.50 and 1.18~1.19-fold. | [50] |
| Arabidopsis VTE7 | constitutive CaMV 35S | Arabidopsis Atvte7 | Tocopherol in leaves increased to 3.6-fold. | [36] |
| Zea mays VTE7 | Tocopherol in leaves increased to 6.9-fold. | |||
| Arabidopsis VTE5 | seed-specific napin | Arabidopsis Atvte5 | Vitamin E in seeds increased to 3.2-fold. | [70] |
| Arabidopsis VTE6 | constitutive CaMV 35S | Arabidopsis | Vitamin E in seeds increased to 1.15-fold. | [47] |
| Arabidopsis VTE2 | Tocopherol in leaves increased to 3~4.4-fold. | [72] | ||
| Arabidopsis VTE2 + Arabidopsis VTE4 | Tocopherol in leaves increased by 1.8~2.1-fold. | |||
| Arabidopsis VTE7 + Arabidopsis VTE2 | Tocopherol in leaves increased by 5.9~6.4-fold. | [36] | ||
| Arabidopsis VTE2 | Tobacco | α-Tocopherol in leaves increased by 5.4-fold. | [71] | |
| Arabidopsis VTE1 | α-Tocopherol in leaves increased by 4-fold. | |||
| Arabidopsis VTE1 + Arabidopsis VTE2 | α-Tocopherol in leaves increased by 7.1-fold. | |||
| Arabidopsis VTE1 | Lettuce | Tocopherol in leaves increased to 2~2.7-fold. | [73] | |
| Arabidopsis VTE2 | Tocopherol in leaves increased to 2-fold. | |||
| Arabidopsis VTE1 | Indica rice ASD16 | Tocopherol in leaves increased to 2.2~3.26-fold. | [74] | |
| Arabidopsis VTE2 | Tocopherol in leaves increased to 2.8~3.5-fold. | |||
| Arabidopsis VTE1 + Arabidopsis VTE2 | Tocopherol in leaves increased to 4.3~5.3-fold. | |||
| Arabidopsis VTE2 + Arabidopsis VTE4 | Potato | α-Tocopherol content in tubers increased to 2.6~2.8-fold, while that in leaves rose to 4.8~5.1-fold. | [75] | |
| Lactuca sativa L. VTE2 | Arabidopsis | α-Tocopherol in leaves increased by 18-fold. | [76] | |
| Malus domestica Borkh. VTE2 | Tomato | α-Tocopherol in leaves and fruits increased to 3.6 and 1.7-fold. | [77] | |
| Hordeum vulgare L. HGGT | endogenous D-Hordein | Barley | Tocotrienol in seeds increased to 1.10~1.15-fold. | [78] |
| Hordeum vulgare L. HGGT | constitutive CaMV 35S | Arabidopsis | Vitamin E in leaves increased to 10~15-fold. | [79] |
| Hordeum vulgare L. HGGT | strong embryo-specific | Maize | Tocotrienol in seeds increased to 6-fold. | |
| Hordeum vulgare L. HGGT | strong seed specific | Soybean | Tocotrienol in seeds increased to 8~10-fold. | [80] |
| Hordeum vulgare L. HGGT | seed-specific napin | Cotton | Cottonseed vitamin E content exhibited a 2~3-fold enhancement. | [7] |
| Hordeum vulgare L. HGGT + Glycine max VTE4 | strong seed specific | Soybean | Tocotrienol in seeds increased to 7-fold. | [80] |
| Hordeum vulgare L. HGGT | seed-specific napin | Canola | Vitamin E in seeds increased to 4.19-fold. | [99] |
| Oryza sativa L. HGGT | constitutive CaMV 35S | Tobacco | Vitamin E in leaves increased to 3.4~4.7-fold. | [81] |
| Hordeum vulgare L. HGGT | —— | Maize | Tocotrienol in kernels and crude oil increased by 18-fold. | [82] |
| Arabidopsis VTE4 | seed-specific carrot DC3 | Arabidopsis | α-Tocopherol in seeds increased by more than 80-fold. | [83] |
| Arabidopsis VTE4 | constitutive CaMV 35S | Lettuce | α-/γ-Tocopherol ratio in T0 plants reached a maximum of 320. | [84] |
| Arabidopsis VTE4 | 16s rRNA | Tobacco chloroplasts | α-Tocopherol in seeds increased to 9.6-fold. | [85] |
| Arabidopsis VTE4 | constitutive CaMV 35S | Brassica juncea | α-Tocopherol in seeds increased by more than 6-fold. | [86] |
| Brassica napus VTE4 | seed-specific | Soybean | α- and β-Tocopherol in T2 seeds increased to 11.1 and 18.9-fold. | [87] |
| Arabidopsis VTE4 | constitutive CaMV 35S | α-Tocopherol in seeds increased to 4-fold. | [88] | |
| Perilla frutescens VTE4 | seed-specific | α-Tocopherol in T2 seeds increased to 10.4-fold. | [89] | |
| Medicago sativa VTE4 | constitutive CaMV 35S | Medicago sativa | α-Tocotrienol in leaves increased by 0.6~2.4-fold. | [90] |
| Medicago sativa VTE4 | Arabidopsis | α-Tocopherol in seeds was at least 10-fold. | ||
| Medicago sativa VTE4 | Medicago sativa | α-Tocopherol and total tocopherol in leaves increased to 1.36 and 1.31-fold. | [91] | |
| Glycine max. VTE4 | Ubi | Maize | α-Tocopherol in seeds increased by 3~4.5-fold. | [100] |
| Glycine max. VTE4 | Arabidopsis | α-Tocopherol in seeds increased by 4~6-fold. | ||
| Zea mays VTE4 | embryo-preferred Glb1 | Maize | α-Tocopherol in seeds increased by 6.5-fold. | [92] |
| Zea mays VTE4 | Arabidopsis | α-Tocopherol in seeds increased by 4~5-fold. | ||
| Arabidopsis VTE3 | seed-specific | Soybean | γ-Tocopherol increased from 50% to 80%. | [95] |
| Arabidopsis VTE3 + Arabidopsis VTE4 | α-Tocopherol increased by more than 8-fold. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, D.; Ren, Y.; Luo, Y.; Zheng, H.; Liu, Y.; Wang, L.; Zhang, L. Vitamin E in Plants: Biosynthesis Pathways, Biofortification Strategies, and Regulatory Dynamics. Int. J. Mol. Sci. 2025, 26, 3380. https://doi.org/10.3390/ijms26073380
Li Y, Yang D, Ren Y, Luo Y, Zheng H, Liu Y, Wang L, Zhang L. Vitamin E in Plants: Biosynthesis Pathways, Biofortification Strategies, and Regulatory Dynamics. International Journal of Molecular Sciences. 2025; 26(7):3380. https://doi.org/10.3390/ijms26073380
Chicago/Turabian StyleLi, Yanjiao, Di Yang, Yuqing Ren, Yanzhong Luo, Hongyan Zheng, Yuan Liu, Lei Wang, and Lan Zhang. 2025. "Vitamin E in Plants: Biosynthesis Pathways, Biofortification Strategies, and Regulatory Dynamics" International Journal of Molecular Sciences 26, no. 7: 3380. https://doi.org/10.3390/ijms26073380
APA StyleLi, Y., Yang, D., Ren, Y., Luo, Y., Zheng, H., Liu, Y., Wang, L., & Zhang, L. (2025). Vitamin E in Plants: Biosynthesis Pathways, Biofortification Strategies, and Regulatory Dynamics. International Journal of Molecular Sciences, 26(7), 3380. https://doi.org/10.3390/ijms26073380