

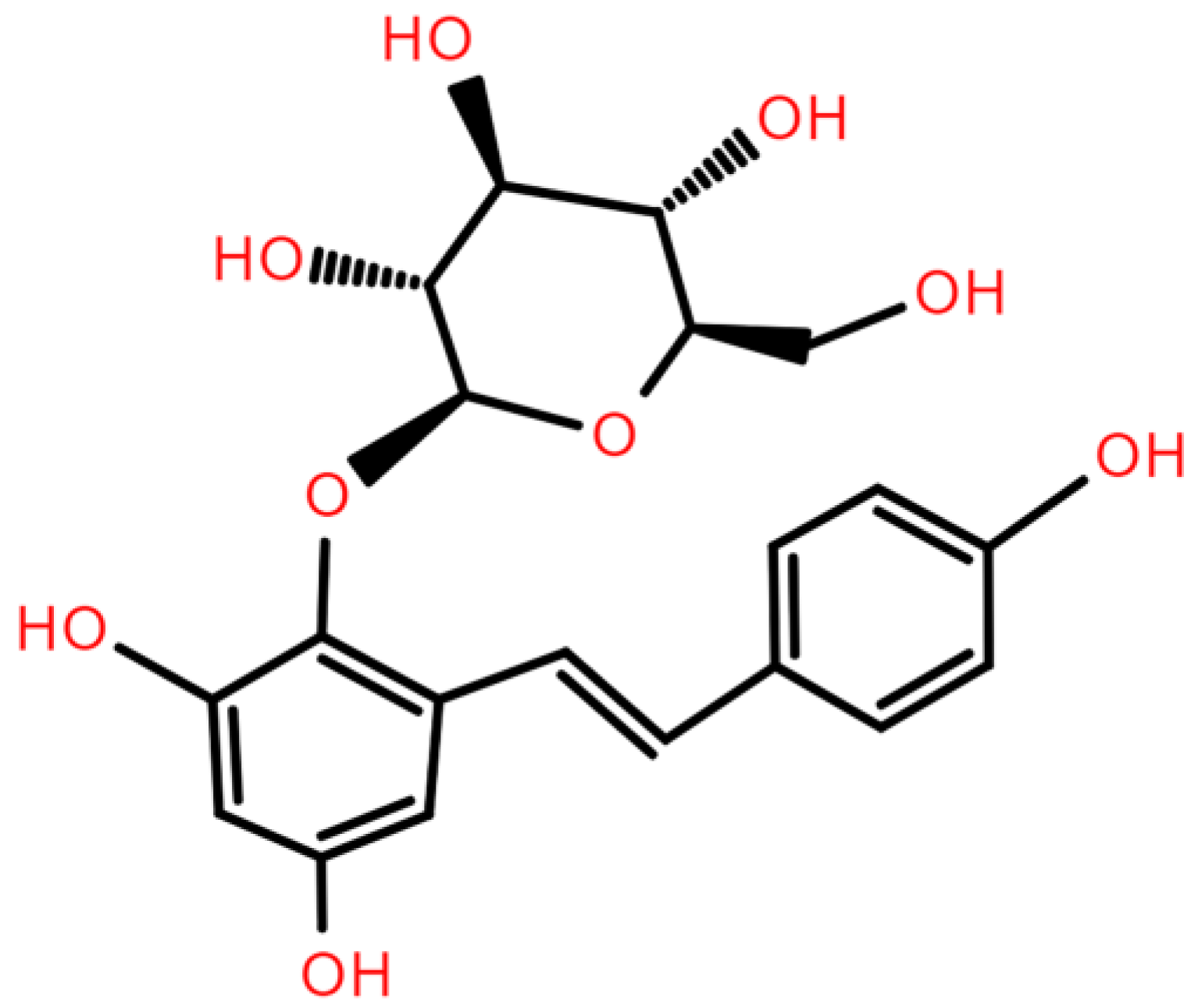
2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside (TSG) from Polygonum multiflorum Thunb.: A Systematic Review on Anti-Aging

 ,
,  ,
,
Abstract
1. Introduction
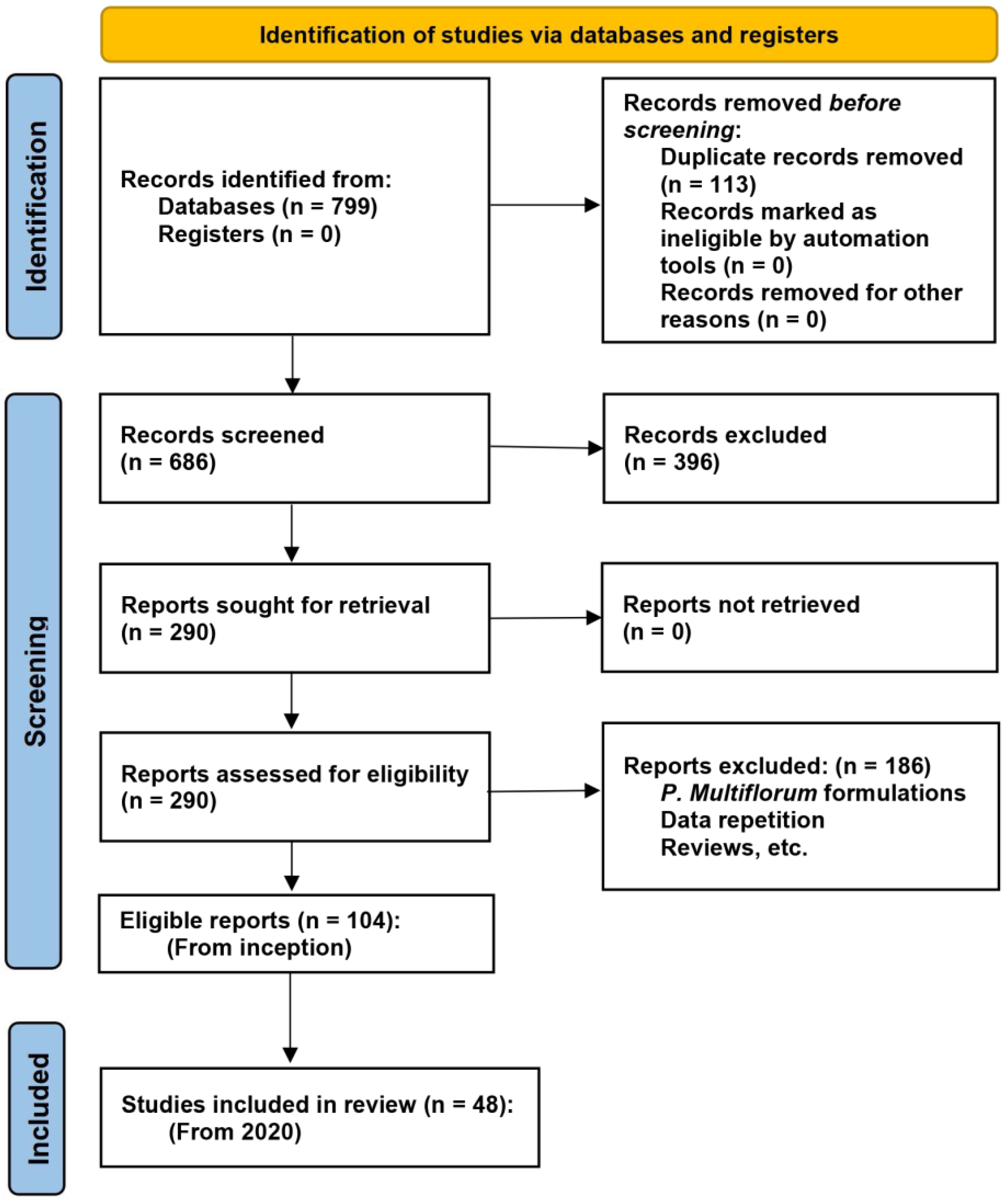
2. Methods
2.1. Search Strategy
2.2. Eligibility and Study Selection
3. Results and Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | Aging Model (Inducer; Object) | TSG Treatment (Dose; Duration) | Chemical Purity | Potential Mechanisms | Author (Year) | References |
|---|---|---|---|---|---|---|
| Lifespan extension | H2O2; larval zebrafish | 25, 50, and 100 μg/mL; 24 h | >98% | Oxidative stress↓, inflammation↓ (SA-β-gal↓, ROS↓, SOD↑, catalase↑, il-1β↓, il-6↓, cxcl-c1c↓, il-8↓) | Xia et al. (2023) | [9] |
| C. elegans | 100, 200, and 400 μM; until death | Unclear (standard) | Mean lifespan↑, mitochondrial function↑ (DAF-16/SKN-1/SIR-2.1 pathways; DAF-16↑, SKN-1↑, SIR-2.1↑, SIRT1↑, Aβ↓, Tau↓, ROS↓, MMP↑) | Sun et al. (2024) | [10] | |
| Neuroprotection | Radiation; C57BL/6J mice, or Tet2−/− mice | 40, 80, and 120 mg/kg/d; 5 months | 98% | Inflammation↓, neurogenesis↑ (AMPK/Tet2 pathway; AMPK↑, Tet2↑, NLRP3↓) | Miao et al. (2022) | [11] |
| LPS/ATP + Aβ; BV2, N2a, and SH-SY5Y cells, and primary microglia | 10, 100 nM, and 1, 10 μM; 24 h | >98% | Inflammation↓, mitophagy↑, mitosis↑ (AMPK/PINK1/Parkin pathway; AMPK↑, PINK1↑, Parkin↑, NLRP3↓, LC3-II/LC3-I↑, p62↓; iNOS↓, COX-2↓, Drp1↑, MIRO↓, Mfn2↑, MFF↓) | Gao et al. (2020) | [12] | |
| Okadaic acid; SH⁃SY5Y cells | 100 μM; 24 h | Unclear (standard) | Apoptosis↓ (PI3K/AKT pathway; PI3K↑, AKT↑, Bcl⁃2↑, Bax↓) | Kang et al. (2024) | [13] | |
| High-glucose; HT-22 cells | 200 µM; 48 h | Unclear (standard) | Apoptosis↓ (HAT↓, HDAC↑, Bcl-2↑, Bax↓, caspase-3↓) | Chen et al. (2022) | [14] | |
| Alleviating AD | ① AD model: APP/PS1 double transgenic mice ② LPS/IFN-γ; BV2 cells | ① 40 and 80 mg/kg/d; 5 months ② 25, 50, and 100 μM; 20 h | Unclear (standard) | Inflammation↓ (cGAS-STING pathway; cGAS↓, STING↓, NF-κB↓, NLRP3↓; IL-1β↓, IL-6↓, TNF-α↓, IFN-α↓, IFN-β↓, IFIT1↓, IRF7↓) | Gao et al. (2023) | [15] |
| AD model: APP/PS1/Tau triple transgenic mice | 0.033, 0.1, and 0.3 g/kg/d; 60 days | ≥70% | CDK5↓, MAPK1↓, PP1↑, Tau↓, p39↓ | Wu et al. (2022) | [16] | |
| AD model: Aβ25–35; SD rats | 0.033, 0.1, and 0.3 g/kg; 4 or 8 weeks | 98% | Apoptosis↓, improving neuronal morphology (PI3K/AKT/GSK-3β pathway; MKK7/JNK pathway; PI3K↑, AKT↑, GSK-3β↓, Tau↓; MKK7↓, JNK↓) | Xia et al. (2023); Li, YB et al. (2023); Li, Y et al. (2023) | [17,18,19] | |
| AD model: N2a/APP695swe cells | 100 μM; 48h | 98% | Apoptosis↓, improving mitochondrial function (MMP↑; PACS-2↓) | Wang et al. (2024) | [20] | |
| AD model: APP/PS1 mouse | 120 mg/kg; 8 weeks | Unclear (standard) | MAPK pathway, chemokine pathway and autophagy—animal | Gao et al. (2024) | [21] | |
| Ameliorating PD | ① PD model: MPTP; C57BL/6J mice ② MPP+; mesencephalic DA neurons or SH-SY5Y cells | ① 20 mg/kg; 7 days ② unclear | ≥98% | Apoptosis↓, neurotoxicity↓ (FGF2-Akt, BDNF-TrkB axis; FGF2↑, Akt↑, DA↑, TH↑, BDNF↑, TrkB↑, Bcl⁃2↑, caspase-3↓) | Yu et al. (2019) | [22] |
| Mouse neural stem cells | 10 μM; 2 weeks | ≥98% | DA neuron differentiation (Wnt/β-catenin pathway; Wnt1↑, Wnt3a↑, Wnt5a↑, β-catenin↑, Nurr1↑) | Zhang et al. (2021) | [23] | |
| Inhibiting AS | High-fat diet; ApoE-deficient (ApoE−/−) mice | 0.035 and 0.07 mg/g/d; 8 weeks | ≥98% | Inflammation↓, lipid accumulation↓, AS plaque↓, and regulating intestinal microbiota (TG↓, ox-LDL↓, IL- 6↓, TNF-α↓, VCAM-1↓, MCP-1↓) | Li et al. (2020) | [24] |
| ① ox-LDL; BMDCs ② ApoE−/− mice | ① 40 and 80 µM, 2 h ② 40 mg/kg/d, 5 weeks | ≥98% | Autophagy↓, DCs maturation↓, Treg differentiation↑, inflammation↓, lipid accumulation↓, (PI3K/AKT/mTOR pathway; PI3K↓, AKT↓, mTOR↓, P62↓; TC↓, TG↓, LDL-C↓; IL-6↓, IL-17A↓, IL-10↑) | Yang et al. (2024) | [25] | |
| ① ApoE−/− mice ② Macrophages in the aorta cells of mice (in ①) | 100 mg/kg/d, 8 weeks | 99% | Atherosclerotic lesions↓, dyslipidemia symptoms↓, and regulating lipid metabolism (Srepb-1c↓, Fasn↓, Scd1↓, Gpat1↓, Dgat1↓, Pparα↑ and Cpt1α↑; Srebp2↓, Hmgcr↓, Ldlr↑, Acat1↓, Acat2↓, and Cyp7a1↑) | Li et al. (2024) | [26] | |
| ① High-fat diet; LDLr−/− mice ② ox-LDL; HAECs | ① 50 and 100 mg/kg/d; 12 weeks ② 1, 10, and 100 μM; 24 h | >98% | Oxidative stress↓, endothelial senescence↓, telomerase activity↑, mitochondrial damage↓, and improving lipid profiles (PGC-1α pathway; PGC-1α↑, TC↓, TG↓, LDL-c↓, ox-LDL↓; γ-H2AX↓, p53↓, p21↓, p16↓; TERT↑; mitoROS↓, NRF1↑, TFAM↑; ROS↓, MDA↓; β-gal↓, MMP↑, mtDNA↓, SOD↑, CAT↑) | Wang et al. (2022) | [27] | |
| Cardiovascular protection | ① Natural aging C57BL/6J mice, Tet2 Mut mice ② IMR-90 fibroblasts | ① 120 mg/kg/d; 60 days ② 10 and 100 μM; 48 h | Unclear (standard) | HSC aging↓, repopulation potential↑, epigenetic reprograming↑, stemness↑ (AMPK-Tet2 axis; CLPs↑, B lymphocytes↑) | Gao et al. (2024) | [28] |
| Ang Ⅱ; HUVECs | 50 and 100 μM; 24 h | Unclear (standard) | Endothelial senescence↓ (SA-β-gal↓, p53↓, PAI-1↓, SIRT1↑) | Fan et al. (2021) | [29] | |
| Anti-hypertension | U46619; superior mesenteric artery of SD rats | Concentration accumulation: 10−5 ∼10−2 M; | ≥98% | Vasodilation (SIRT1/TP pathway; SIRT1↑, TP↓) | Chen et al. (2022) | [30] |
| HHcy; C57BL/6 mice | 40, 80, and 160 mg/kg; 4 weeks | 98% | Inhibiting vasoconstriction (ERK1/2/NF-κB pathway; p-ERK1/2↓, p-p65↓, endothelin-1↓; BP↓, Hcy↓) | Jia et al. (2022) | [31] | |
| ① ZDF rats, OMT−/− mice ② HUVEC and mature adipocyte co-culture | ① 50, 100, and 200 mg/kg/d; 2 weeks ② 100 μM; 24 h | ≥98% | Oxidative/nitrative stress↓, improving endothelial function (Akt/eNOS/NO pathway; SBP↓, omentin-1↑, Akt↓, eNOS↓, NO↓; NOX2↓, p22phox↓, SOD↓, peroxynitrite anion↓, PPAR-γ↑, Itln-1↑) | Dong et al. (2021) | [32] | |
| Reproductive protection | H2O2 + FeSO4; rat testicular Leydig cells | 150 μM; 48 h | Unclear (standard) | Oxidative stress↓, cell senescence↓ (Insulin/IGF-1pathway; SA-β-gal↓, IRS1↑, IGF-1↑, IRS2↑, INSR↑, IGFBP3↓) | Li et al. (2021) | [33] |
| Normal C57BL/6J mice | 10 mg/kg/d; 32, or 16 weeks | 95% | Oocyte quantity and quality↑, mitochondrial biogenesis↑, steroidogenesis ↑ (AMH↑, PR-B↑, atp6↑, pgc1α↑, cyp11a↑, cyp19↑, er-β↑) | Lin et al. (2022) | [34] | |
| Estrogenic activity | ER (+) MCF-7 cells | 100 nM; 24 h | Unclear (standard) | Cell proliferation↑, acting as phytoestrogen (ERα↑, ERβ↑, pS2↑) | Akter et al. (2023) | [35] |
| Reducing OP | ① OP model: OVX; SD rats ② H2O2; MC3T3-E1 cells | ① 80 mg/kg/d; 3 months ② 10 μM; 24 h | ≥98% | Oxidative stress↑, apoptosis↓, bone resorption↓ (miR-34a↑, SIRT1↓; Conn.D↑, Tb.N↑, BMD↑, MDA↓, GSH-Px↑; ALP↑, OPN↑, COL-1↑, OCN↑) | Wang et al. (2022) | [36] |
| Diabetic OP model: Streptozotocin; C57BL/6J mice | 10 and 40 mg/kg; 8 weeks | Unclear (standard) | Regulating osteogenesis and osteoclast genesis (Ca↑, RUNX-2↑, COL-I↑, OCN↑, β-catenin↑, RAS↓, OPG↑, RANKL↓, sclerostin↓) | Zhang et al. (2019) | [37] | |
| Bone protection | BMSCs | 10−6, 10−5, and 10−4 M; 3 or 7 days | Unclear (standard) | Cell proliferation↑, osteogenic differentiation↑ (ALP↑, OCN↑, Col1a1↑, Runx2↑, β-catenin↑) | Liang et al. (2022) | [38] |
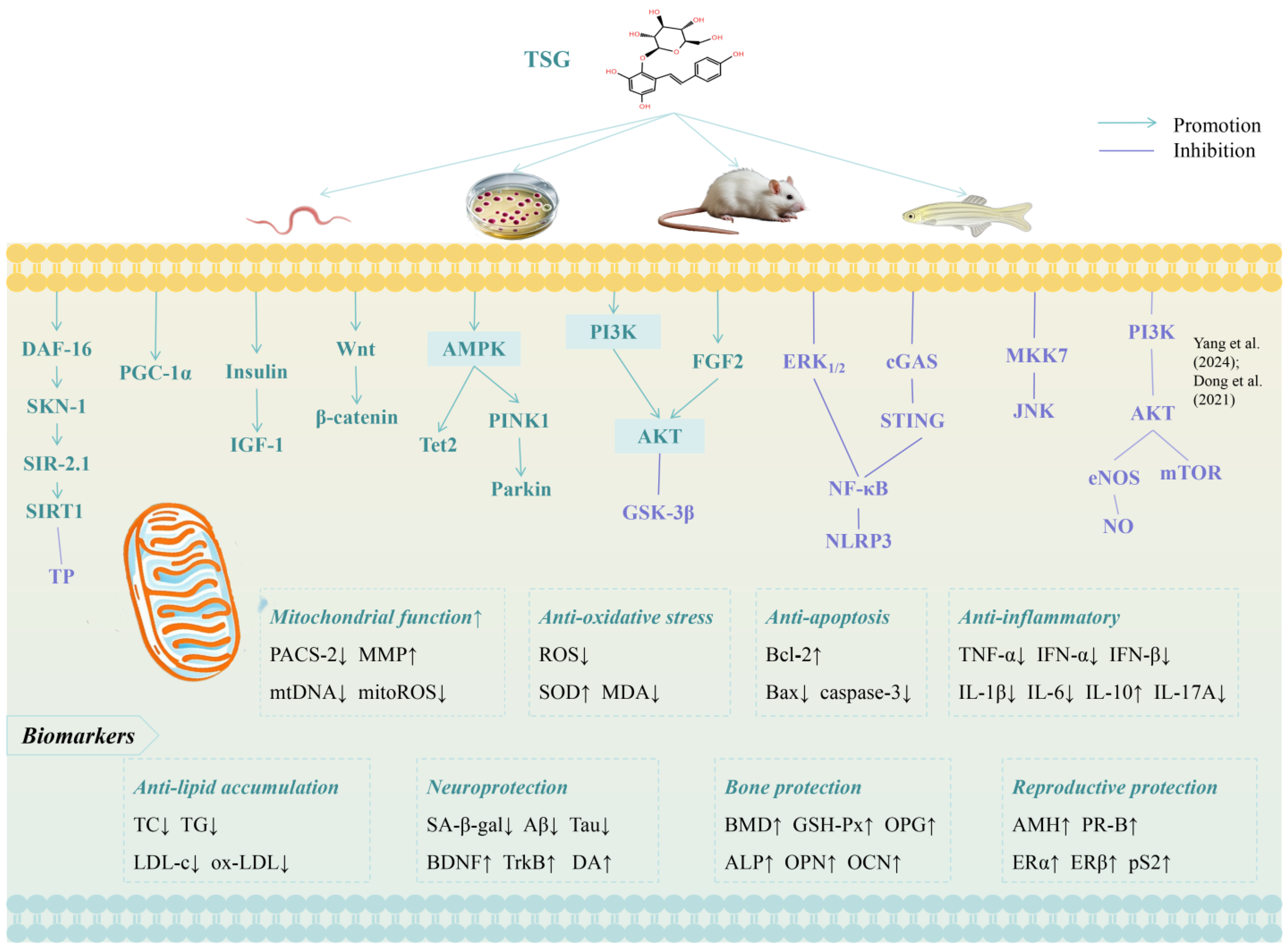
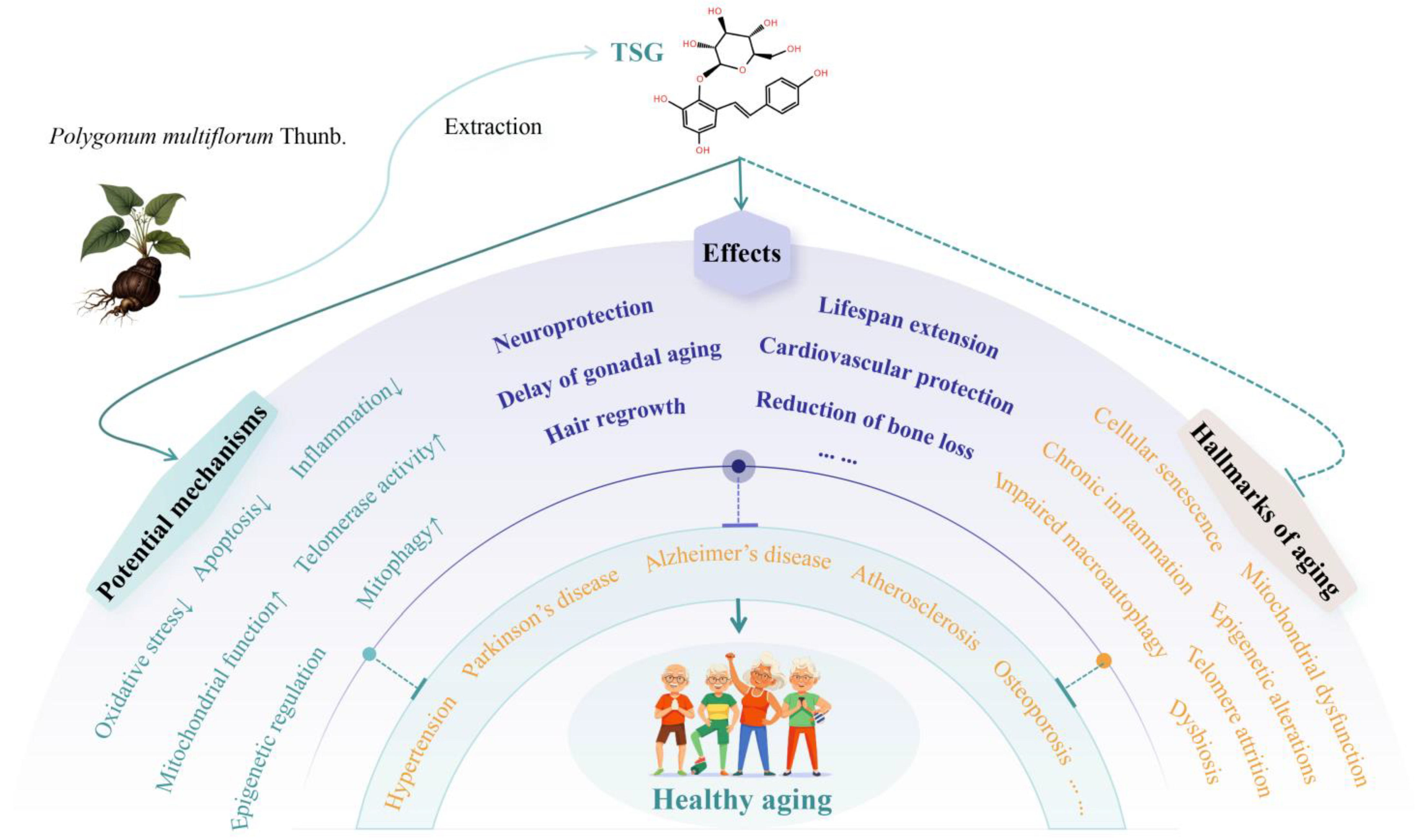
3.1. Lifespan-Extending Effects of TSG
3.2. Neuroprotective Effects of TSG
3.2.1. Attenuation of Alzheimer’s Disease (AD)
3.2.2. Amelioration of Parkinson’s Disease (PD)
3.3. Cardiovascular Protective Effects of TSG
3.3.1. Inhibition of Vascular Senescence and Atherosclerosis (AS)
3.3.2. Anti-Hypertensive Effects
3.4. Reproductive Protective Effects of TSG
3.5. Bone Protective Effects of TSG
3.6. Other Protective Effects of TSG
3.7. Effects of Other P. multiflorum Extracts Against Aging and Age-Related Diseases
3.7.1. Identified Compounds
3.7.2. Ethanol Extract
3.7.3. Aqueous Extract
4. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Aβ | β-Amyloid |
| AD | Alzheimer’s disease |
| AKT | Protein kinase B |
| Ang II | Angiotensin II |
| APP | Amyloid precursor protein |
| AS | Atherosclerosis |
| BALP | Bone alkaline phosphatase |
| Bcl-2 | B-cell lymphoma-2 |
| Bax | Bcl-2-associated X protein |
| BDNF | Brain-derived neurotrophic factor |
| BMD | Bone mineral density |
| BMSCs | Bone mesenchymal stem cells |
| BP | Blood pressure |
| CDKN1A | Cyclin-dependent kinase inhibitor 1A |
| C. elegans | Caenorhabditis elegans |
| cGAS | Cyclic GMP-AMP synthase |
| CLPs | Common lymphoid progenitors |
| Conn.D | Connectivity density |
| DA | Dopaminergic |
| DAF-16 | Abnormal dauer formation-16 |
| DCs | Dendritic cells |
| DE | Diabetic encephalopathy |
| D-gal | D-galactose |
| DOR | Diminished ovarian reserve |
| ERK | Extracellular signal-regulated kinase |
| ET-1 | Endothelin-1 |
| FGF2 | Fibroblast growth factor 2 |
| FOXO | Forkhead box O |
| GSH-Px | Glutathione peroxidase |
| GSK-3β | Glycogen synthase kinase-3β |
| HAECs | Human aortic endothelial cells |
| HDAC | Histone deacetylase |
| HHcy | Hyperhomocysteinemia |
| HSCs | Hematopoietic stem cells |
| HUVECs | Human umbilical vein endothelial cells |
| IGF | Insulin-like growth factor |
| IRS1 | Insulin receptor substrate 1 |
| JNK | Jun N-terminal kinase |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| MCP-1 | Monocyte chemotactic protein-1 |
| MMP | Mitochondrial membrane potential |
| MPTP | 1-Methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine |
| MPP+ | 1-Methyl-4-phenylpyridinium |
| mTOR | Mechanistic target of rapamycin |
| NLRP3 | Nod-like receptor protein 3 |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| NSCs | Neural stem cells |
| OCN | Osteocalcin |
| OPG | Osteoprotegerin |
| OPN | Osteopontin |
| OVX | Ovariectomized |
| PACS-2 | Phosphofurin acidic cluster sorting protein-2 |
| PAI-1 | Plasminogen activator inhibitor-1 |
| PD | Parkinson’s disease |
| PGC-1α | Proliferator-activated receptor γ coactivator 1α |
| PI3K | Phosphatidylinositol-3-kinase |
| PMOP | Postmenopausal osteoporosis |
| POF | Premature ovarian failure |
| POI | Premature ovarian insufficiency |
| P.multiflorum | Polygonum multiflorum Thunb. |
| PPAR-γ | Peroxisome proliferator-activated receptor-γ |
| RANKL | Receptor activator for nuclear factor kappa B ligand |
| ROS | Reactive oxygen species |
| RUNX-2 | Runt-related transcription factor 2 |
| SA-β-gal | Senescence-associated β-galactosidase |
| SIRT1 | Silent information regulator 1 |
| SKN-1 | Skinhead-1 |
| SOD | Superoxide dismutase |
| STING | Stimulator of interferon genes |
| Tb.N | Trabecular number |
| TC | Total cholesterol |
| TG | Triglyceride |
| TNF-α | Tumor necrosis factor-α |
| TRAP | Tartrate-resistant acid phosphatase |
| TrkB | Tropomyosin receptor kinase-B |
| TSG | 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside |
| VaD | Vascular dementia |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| ZDF | Zucker diabetic fatty |
| 6-OHDA | 6-Hydroxydopamine |
References
- Stakos, D.A.; Stamatelopoulos, K.; Bampatsias, D.; Sachse, M.; Zormpas, E.; Vlachogiannis, N.I.; Tual-Chalot, S.; Stellos, K. The Alzheimer’s Disease Amyloid-Beta Hypothesis in Cardiovascular Aging and Disease: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 952–967. [Google Scholar] [CrossRef]
- López-Otín, C.; Pietrocola, F.; Roiz-Valle, D.; Galluzzi, L.; Kroemer, G. Meta-hallmarks of aging and cancer. Cell Metab. 2023, 35, 12–35. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Simon, M.; Seluanov, A.; Gorbunova, V. DNA damage and repair in age-related inflammation. Nat. Rev. Immunol. 2023, 23, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Qu, J.; Zhang, W.; Liu, G.H. Stress, epigenetics, and aging: Unraveling the intricate crosstalk. Mol. Cell 2024, 84, 34–54. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, C.; Zhang, W.; Wang, Y.; Qian, P.; Huang, H. Inflammation and aging: Signaling pathways and intervention therapies. Signal Transduct. Target. Ther. 2023, 8, 239. [Google Scholar] [CrossRef]
- Commission, C.P. Pharmacopoeia of the People’s Republic of China 2020; China Medical Science and Technology Press: Beijing, China, 2020. [Google Scholar]
- Xia, H.; Cheng, X.; Cao, M.; Sun, X.; He, F.; Yao, X.; Liu, H. Tetrahydroxystilbene Glucoside Attenuates Oxidative Stress-Induced Aging by Regulating Oxidation Resistance and Inflammation in Larval Zebrafish. Zebrafish 2023, 20, 55–66. [Google Scholar] [CrossRef]
- Sun, M.; Wei, C.; Gao, Y.; Chen, X.; Zhong, K.; Li, Y.; Yang, Z.; Gao, Y.; Wang, H. TSG Extends the Longevity of Caenorhabditis elegans by Targeting the DAF-16/SKN-1/SIR-2.1-Mediated Mitochondrial Quality Control Process. Antioxidants 2024, 13, 1086. [Google Scholar] [CrossRef]
- Miao, B.B.; Gao, D.; Hao, J.P.; Li, Y.L.; Li, L.; Wang, J.B.; Xiao, X.H.; Yang, C.C.; Zhang, L. Tetrahydroxy stilbene glucoside alters neurogenesis and neuroinflammation to ameliorate radiation-associated cognitive disability via AMPK/Tet2. Int. Immunopharmacol. 2022, 110, 108928. [Google Scholar] [CrossRef]
- Gao, Y.; Li, J.; Li, J.; Hu, C.; Zhang, L.; Yan, J.; Li, L.; Zhang, L. Tetrahydroxy stilbene glycoside alleviated inflammatory damage by mitophagy via AMPK related PINK1/Parkin signaling pathway. Biochem. Pharmacol. 2020, 177, 113997. [Google Scholar] [CrossRef]
- Kang, B.Q.; Li, Y.; He, X.X.; Xiao, Z.; Hu, R.; Luo, C.L.; Qiao, M.Y.; Wu, G.Y.; Li, Z.Z.; Zhu, X.Y.; et al. Mechanism of stilbene glycosides on apoptosis of SH⁃SY5Y cells via regulating PI3K/AKT signaling pathway. J. Hainan Med. Univ. 2024, 30, 8–14. [Google Scholar] [CrossRef]
- Chen, G.; Xu, Y.J.; Huang, C.X.D.; Lin, H.R.; Yang, T.T.; Zhu, L.Y.; Li, X.; Pan, W. Tetrahydroxystilbene glucoside and emodin improves the hippocampal neuronal apoptosis in mice induced by high glucose. Tianjin Med. J. 2022, 50, 561–565. [Google Scholar] [CrossRef]
- Gao, D.; Hao, J.P.; Li, B.Y.; Zheng, C.C.; Miao, B.B.; Zhang, L.; Li, Y.L.; Li, L.; Li, X.J.; Zhang, L. Tetrahydroxy stilbene glycoside ameliorates neuroinflammation for Alzheimer’s disease via cGAS-STING. Eur. J. Pharmacol. 2023, 953, 175809. [Google Scholar] [CrossRef]
- Wu, W.X.; Su, Y.Z.; Li, Z.Z.; Liu, C.Y.; Meng, W.Y.; Huang, J.; Zhu, X.Y.; Liu, J.B.; Liu, D.M.; Huang, Z.S. Effects of stilbene glycoside on CDK5, MAPK1 and PP1 in Alzheimer’s disease mice. Chin. J. Gerontol. 2022, 42, 2206–2210. [Google Scholar] [CrossRef]
- Xia, X.Y.; Li, Y.B.; He, X.X.; Li, Y.; Kang, B.Q.; Li, Z.Z.; Huang, Z.S. Effect of stilbene glycoside on Ser199 posphorylation in Alzheimer’s disease rats. Chin. J. Clin. Pharmacol. 2023, 39, 2368–2372. [Google Scholar] [CrossRef]
- Li, Y.B. Study on the Intervention of Stilbene Glycoside on Tau Protein Phosphorylation in Rats with Alzheimer’s Disease via PI3K/AKT/GSK-3β Signaling Pathway. Master’s Thesis, Youjiang Medical University for Nationalities, Baise, China, 2023. [Google Scholar]
- Li, Y.; Kang, B.Q.; He, X.X.; Hu, R.; Xiao, Z.; Luo, C.L.; Wu, G.Y.; Huang, Z.S. The effect and mechanism of stilbene glycosides on improving neuronal injury in Alzheimer’s disease rats by regulating ASK/MKK7/JNK pathway. J. Hainan Med. Univ. 2023, 29, 1601–1606+1613. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Ye, Q.Y.; Qian, J.; Liu, Z.P.; Luo, H.B.; Li, Y. Effect of PACS-2 on the development of Alzheimer’s disease. China Mod. Dr. 2024, 62, 11–15. [Google Scholar]
- Gao, Y.; Li, J.; Hu, K.; Wang, S.; Yang, S.; Ai, Q.; Yan, J. Phosphoproteomic analysis of APP/PS1 mice of Alzheimer’s disease by DIA based mass spectrometry analysis with PRM verification. J. Proteom. 2024, 299, 105157. [Google Scholar] [CrossRef]
- Yu, Y.; Lang, X.Y.; Li, X.X.; Gu, R.Z.; Liu, Q.S.; Lan, R.; Qin, X.Y. 2,3,5,4’-Tetrahydroxystilbene-2-O-β-d-glucoside attenuates MPP+/MPTP-induced neurotoxicity in vitro and in vivo by restoring the BDNF-TrkB and FGF2-Akt signaling axis and inhibition of apoptosis. Food Funct. 2019, 10, 6009–6019. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, H. Promotive effects of tetrahydroxystilbene glucoside on the differentiation of neural stem cells from the mesencephalon into dopaminergic neurons. Neurosci. Lett. 2021, 742, 135520. [Google Scholar] [CrossRef]
- Li, F.; Zhang, T.; He, Y.; Gu, W.; Yang, X.; Zhao, R.; Yu, J. Inflammation inhibition and gut microbiota regulation by TSG to combat atherosclerosis in ApoE(-/-) mice. J. Ethnopharmacol. 2020, 247, 112232. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bai, D.; Jiang, L.; Chen, Y.; Wang, M.; Wang, W.; Wang, H.; He, Q.; Bu, G.; Long, J.; et al. Stilbene glycosides alleviate atherosclerosis partly by promoting lipophagy of dendritic cells. Int. Immunopharmacol. 2024, 143 Pt 1, 113223. [Google Scholar] [CrossRef]
- Li, M.; Meng, Y.; Hong, X.; Chai, H.; Huang, J.; Wang, F.; Zhang, W.; Wang, J.; Liu, Q.; Xu, Y. Anti-atherosclerotic effect of tetrahydroxy stilbene glucoside via dual-targeting of hepatic lipid metabolisms and aortic M2 macrophage polarization in ApoE(-/-) mice. J. Pharm. Biomed. Anal. 2024, 248, 116338. [Google Scholar] [CrossRef]
- Wang, C.Y.; Wang, J.; Cao, J.; Xu, J.; Wu, R.M.; Xu, X.L. Activating PGC-1α-mediated signaling cascades in the aorta contributes to the amelioration of vascular senescence and atherosclerosis by 2,3,4’,5-tetrahydroxystilbene-2-O-β-d-glycoside. Phytomedicine 2022, 99, 154017. [Google Scholar] [CrossRef]
- Gao, D.; Yi, W.W.; Liu, B.; Zhang, C.E.; Yang, C.C.; Zeng, L.; Li, L.; Luo, G.; Zhang, L.; Ju, Z.Y.; et al. Tetrahydroxy stilbene glucoside rejuvenates aging hematopoietic stem cells with predilection for lymphoid differentiation via AMPK and Tet2. J. Adv. Res. 2024, 70, 515–529. [Google Scholar] [CrossRef]
- Fan, W.; Guo, Y.; Cao, S.; Cao, S.; Xie, Y.; Liu, X.; Jin, B. Tetrahydroxystilbene glucoside alleviates angiotensin II induced HUVEC senescence via SIRT1. Can. J. Physiol. Pharmacol. 2021, 99, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.L.; Qin, Q.H.; Hou, Y.; Zhang, H.; Jia, M. Study on the mechanism of stilbene glycosides vasodilation via SIRT1-TP pathway based on molecular docking technology. Chin. J. Hosp. Pharm. 2022, 42, 1771–1775. [Google Scholar] [CrossRef]
- Jia, M.; Su, X.; Qin, Q.; Li, Y.; Wang, S.; Chen, Y. Tetrahydroxystilbene glucoside attenuated homocysteine-upregulated endothelin receptors in vascular smooth muscle cells via the ERK(1) (/2) /NF-κB signaling pathway. Phytother. Res. 2022, 36, 3352–3361. [Google Scholar] [CrossRef]
- Dong, Q.; Xing, W.; Li, K.; Zhou, X.; Wang, S.; Zhang, H. Tetrahydroxystilbene glycoside improves endothelial dysfunction and hypertension in obese rats: The role of omentin-1. Biochem. Pharmacol. 2021, 186, 114489. [Google Scholar] [CrossRef]
- Li, S.H.; Jiang, L.P.; Niu, J.Y.; Wang, L.; Niu, S.Y.; Qi, F. Regulation mechanism of tetrahydroxy stilbene glycoside on insulin / IGF-1 signaling pathway to delaying the senescence of rat leydig cells. J. Hebei Univ. (Nat. Sci. Ed.) 2021, 41, 297–303. [Google Scholar]
- Lin, H.Y.; Yang, Y.N.; Chen, Y.F.; Huang, T.Y.; Crawford, D.R.; Chuang, H.Y.; Chin, Y.T.; Chu, H.R.; Li, Z.L.; Shih, Y.J.; et al. 2,3,5,4’-Tetrahydroxystilbene-2-O-β-D-Glucoside improves female ovarian aging. Front. Cell Dev. Biol. 2022, 10, 862045. [Google Scholar] [CrossRef] [PubMed]
- Akter, R.; Yang, D.U.; Ahn, J.C.; Awais, M.; Nahar, J.; Ramadhania, Z.M.; Kim, J.Y.; Lee, G.J.; Kwak, G.Y.; Lee, D.W.; et al. Comparison of In Vitro Estrogenic Activity of Polygoni multiflori Radix and Cynanchi wilfordii Radix via the Enhancement of ERα/β Expression in MCF7 Cells. Molecules 2023, 28, 2199. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, P.; Sun, J.J.; Yao, S.H. Effect and mechanism of stilbene glucoside regulating miR-34a/SIRT1 on osteoporosis in rats. Chin. J. Osteoporos. 2022, 28, 185–191. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, X.; Chen, B.; Tong, L.; Zhang, Y. Tetrahydroxy stilbene glucoside protected against diabetes-induced osteoporosis in mice with streptozotocin-induced hyperglycemia. Phytother. Res. 2019, 33, 442–451. [Google Scholar] [CrossRef]
- Liang, Y.Y.; Zhen, L.; Chen, X.L.; Chen, J.M.; Cui, L.; Liu, Y.Y. Effects and mechanism of tetrahydroxystilbene glycoside on the proliferation and differentiation of bone marrow mesenchymal stem cells. Chin. J. Hosp. Pharm. 2022, 42, 1978–1982. [Google Scholar] [CrossRef]
- Büchter, C.; Zhao, L.; Havermann, S.; Honnen, S.; Fritz, G.; Proksch, P.; Wätjen, W. TSG (2,3,5,4’-Tetrahydroxystilbene-2-O-β-D-glucoside) from the Chinese Herb Polygonum multiflorum Increases Life Span and Stress Resistance of Caenorhabditis elegans. Oxid. Med. Cell Longev. 2015, 2015, 124357. [Google Scholar] [CrossRef]
- Zhou, X.X. Study on the Preparation Process of High Purity Tetrahydroxystilbene Glucoside and Its Anti-Aging Mechanism. Ph.D. Thesis, The Fourth Military Medical University, Xian, China, 2013. [Google Scholar]
- Wu, D.J.; Fan, F.; Chen, J.Y.; Ge, Z.L. Effect of Gstm3 on Tetrahydroxy stilbene glycoside delaying senescence of SH-SY5Y cells. J. Zunyi Med. Univ. 2022, 45, 296–303. [Google Scholar] [CrossRef]
- Lin, F.; Zhou, Y.; Shi, W.; Wan, Y.; Zhang, Z.; Zhang, F. Tetrahydroxystilbene Glucoside Improves Neurotrophic Factors Release in Cultured Astroglia. CNS Neurol. Disord. Drug Targets 2016, 15, 514–519. [Google Scholar] [CrossRef]
- Monteiro, A.R.; Barbosa, D.J.; Remião, F.; Silva, R. Alzheimer’s disease: Insights and new prospects in disease pathophysiology, biomarkers and disease-modifying drugs. Biochem. Pharmacol. 2023, 211, 115522. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Zhang, L.; Zhang, L.; Wang, Y.L.; Li, L. Anti-amyloidgenic and neurotrophic effects of tetrahydroxystilbene glucoside on a chronic mitochondrial dysfunction rat model induced by sodium azide. J. Nat. Med. 2018, 72, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Zeng, C.H.; Cao, Z.Y.; Chi, X.Y.; Duan, Y.T.; Zhou, F.L.; Liu, S.L.; Yang, K. Action mechanism of radix polygoni multiflori in the improvement of Alzheimer’s disease based on network pharmacology and molecular docking. Chin. J. Gerontol. 2023, 43, 3705–3712. [Google Scholar] [CrossRef]
- Leite Silva, A.B.R.; Gonçalves de Oliveira, R.W.; Diógenes, G.P.; de Castro Aguiar, M.F.; Sallem, C.C.; Lima, M.P.P.; de Albuquerque Filho, L.B.; Peixoto de Medeiros, S.D.; Penido de Mendonça, L.L.; de Santiago Filho, P.C.; et al. Premotor, nonmotor and motor symptoms of Parkinson’s Disease: A new clinical state of the art. Ageing Res. Rev. 2023, 84, 101834. [Google Scholar] [CrossRef]
- Zhang, R.; Sun, F.; Zhang, L.; Sun, X.; Li, L. Tetrahydroxystilbene glucoside inhibits α-synuclein aggregation and apoptosis in A53T α-synuclein-transfected cells exposed to MPP. Can. J. Physiol. Pharmacol. 2017, 95, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Lin, F.; Wang, G.; Lu, D.; Wu, Q.; Liu, J.; Shi, J.; Zhang, F. Tetrahydroxystilbene Glucoside Produces Neuroprotection against 6-OHDA-Induced Dopamine Neurotoxicity. Oxid. Med. Cell Longev. 2018, 2018, 7927568. [Google Scholar] [CrossRef]
- Ungvari, Z.; Tarantini, S.; Sorond, F.; Merkely, B.; Csiszar, A. Mechanisms of Vascular Aging, A Geroscience Perspective: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 931–941. [Google Scholar] [CrossRef]
- Yi, B.; Nguyen, M.C.; Won, M.H.; Kim, Y.M.; Ryoo, S. Arginase Inhibitor 2,3,5,4’-Tetrahydroxystilbene-2-O-β-D-Glucoside Activates Endothelial Nitric Oxide Synthase and Improves Vascular Function. Planta Med. 2017, 83, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Gu, C.; Shao, H.; Meng, G.; Wang, H.; Jing, X.; Zhang, W. Tetrahydroxystilbene glucoside improves TNF-α-induced endothelial dysfunction: Involvement of TGFβ/Smad pathway and inhibition of vimentin expression. Am. J. Chin. Med. 2015, 43, 183–198. [Google Scholar] [CrossRef]
- (NCD-RisC), N.R.F.C. Worldwide trends in hypertension prevalence and progress in treatment and control from 1990 to 2019: A pooled analysis of 1201 population-representative studies with 104 million participants. Lancet 2021, 398, 957–980. [Google Scholar] [CrossRef]
- Hay, M.; Barnes, C.; Huentelman, M.; Brinton, R.; Ryan, L. Hypertension and Age-Related Cognitive Impairment: Common Risk Factors and a Role for Precision Aging. Curr. Hypertens. Rep. 2020, 22, 80. [Google Scholar] [CrossRef]
- Dong, Q.; Xing, W.; Fu, F.; Liu, Z.; Wang, J.; Liang, X.; Zhou, X.; Yang, Q.; Zhang, W.; Gao, F.; et al. Tetrahydroxystilbene Glucoside Inhibits Excessive Autophagy and Improves Microvascular Endothelial Dysfunction in Prehypertensive Spontaneously Hypertensive Rats. Am. J. Chin. Med. 2016, 44, 1393–1412. [Google Scholar] [CrossRef] [PubMed]
- Reid, I.R.; Billington, E.O. Drug therapy for osteoporosis in older adults. Lancet 2022, 399, 1080–1092. [Google Scholar] [CrossRef]
- Kim, S.J.; Hwang, Y.H.; Mun, S.K.; Hong, S.G.; Kim, K.J.; Kang, K.Y.; Son, Y.J.; Yee, S.T. Protective Effects of 2,3,5,4’-Tetrahydroxystilbene-2-O-β-d-glucoside on Ovariectomy Induced Osteoporosis Mouse Model. Int. J. Mol. Sci. 2018, 19, 2554. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.S.; Li, Q.; Hamdan, N.; Bian, Y.F.; Zhuang, S.; Fan, K.; Liu, Z.J. Tetrahydroxystilbene Glucoside Regulates Proliferation, Differentiation, and OPG/RANKL/M-CSF Expression in MC3T3-E1 Cells via the PI3K/Akt Pathway. Molecules 2018, 23, 2306. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, J.; Wu, J.; Yu, Y.; Yao, W.; Zhou, M.; Tian, J.; Zhang, J.; Cui, L.; Zeng, X.; et al. Tetrahydroxystilbene glucoside isolated from Polygonum multiflorum Thunb. demonstrates osteoblast differentiation promoting activity. Exp. Ther. Med. 2017, 14, 2845–2852. [Google Scholar] [CrossRef]
- Ning, Z.; Li, Y.; Liu, D.; Owoicho Orgah, J.; Zhu, J.; Wang, Y.; Zhu, Y. Tetrahydroxystilbene Glucoside Delayed Senile Symptoms in Old Mice via Regulation of the AMPK/SIRT1/PGC-1α Signaling Cascade. Gerontology 2018, 64, 457–465. [Google Scholar] [CrossRef]
- Xu, X.G. Study on the Mechanism of Kidney-Tonifying Method Antagonizing Myelosuppression After Chemotherapy by Regulating Bone Marrow Stem Cells. Master’s Thesis, Guangzhou University of Chinese Medicine, Guangzhou, China, 2018. [Google Scholar]
- Chen, L.; Duan, H.; Xie, F.; Gao, Z.; Wu, X.; Chen, F.; Wu, W. Tetrahydroxystilbene Glucoside Effectively Prevents Apoptosis Induced Hair Loss. Biomed. Res. Int. 2018, 2018, 1380146. [Google Scholar] [CrossRef]
- Hu, S.; Sun, Q.; Xu, F.; Jiang, N.; Gao, J. Age-related hearing loss and its potential drug candidates: A systematic review. Chin. Med. 2023, 18, 121. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Chen, G.; Zhu, L.; Lin, H.; Huang, C.; Wei, S.; Yang, T.; Qian, W.; Li, X.; et al. Radix polygoni multiflori protects against hippocampal neuronal apoptosis in diabetic encephalopathy by inhibiting the HDAC4/JNK pathway. Biomed. Pharmacother. 2022, 153, 113427. [Google Scholar] [CrossRef]
- Zhang, L.; Tian, W.Y.; Wang, H.S.; Lai, C.Q.; Li, M.J.; Xia, L.; Leng, C.L. Shouwu emodin against ApoE-/- mouse atherosclerosis based on PI3K/AKT/mTOR signaling pathway. Chin. J. Gerontol. 2021, 41, 2769–2773. [Google Scholar] [CrossRef]
- Kim, J.; Kim, M.M. The effect of emodin on melanogenesis through the modulation of ERK and MITF signaling pathway. Nat. Prod. Res. 2022, 36, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Lao, Z.; Fan, Y.; Huo, Y.; Liao, F.; Zhang, R.; Zhang, B.; Kong, Z.; Long, H.; Xie, J.; Sang, C.; et al. Physcion, a novel inhibitor of 5α-reductase that promotes hair growth in vitro and in vivo. Arch. Dermatol. Res. 2022, 314, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Li, Y.; Wang, Y.; Ye, Q.; Luo, H. Effects of tetrahydroxy stilbene glycoside derivatives on free radical damage and apoptosis in APP695V717I transgenic mice. Redox Rep. 2023, 28, 2259246. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Xiong, Y.; Hu, Y.; Chen, L.; Panayi, A.C.; Xue, H.; Zhou, W.; Yan, C.; Hu, L.; Xie, X.; et al. Polydatin Ameliorates Osteoporosis via Suppression of the Mitogen-Activated Protein Kinase Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 730362. [Google Scholar] [CrossRef]
- Fan, J.; Wang, Y.; Yang, J.; Gu, D.; Kang, S.; Liu, Y.; Jin, H.; Wei, F.; Ma, S. Anti-aging activities of neutral and acidic polysaccharides from Polygonum multiflorum Thunb in Caenorhabditis elegans. Int. J. Biol. Macromol. 2024, 257 Pt 2, 128724. [Google Scholar] [CrossRef]
- Zou, J.Y.; Yang, J.Q.; Xu, L.; Fan, F.; Ge, Z.L. Effects of Polygonum multiflorum extract on DNA methylation and gene transcription in brain tissue of aging mice. J. Third Mil. Med. Univ. 2020, 42, 9–17. [Google Scholar] [CrossRef]
- Yang, J.; He, Y.; Zou, J.; Xu, L.; Fan, F.; Ge, Z. Effect of Polygonum Multiflorum Thunb on liver fatty acid content in aging mice induced by D-galactose. Lipids Health Dis. 2019, 18, 128. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, M.; Wang, R.; Liang, Y.; Zhuang, G.; Chen, X.; Luo, S.; Cai, Y.; Song, C.; Liu, L.; et al. Alleviation of Glucocorticoid-Induced Osteoporosis in Rats by Ethanolic Reynoutria multiflora (Thunb.) Moldenke Extract. J. Med. Food 2024, 27, 287–300. [Google Scholar] [CrossRef]
- Wu, F.; Li, Y.; Liu, W.; Xiao, R.; Yao, B.; Gao, M.; Xu, D.; Wang, J. Comparative Investigation of Raw and Processed Radix Polygoni Multiflori on the Treatment of Vascular Dementia by Liquid Chromatograph-Mass Spectrometry Based Metabolomic Approach. Metabolites 2022, 12, 1297. [Google Scholar] [CrossRef]
- Sun, M.L.; Chen, X.Y.; Cao, J.J.; Cui, X.H.; Wang, H.B. Polygonum multiflorum Thunb extract extended the lifespan and healthspan of Caenorhabditis elegans via DAF-16/SIR-2.1/SKN-1. Food Funct. 2021, 12, 8774–8786. [Google Scholar] [CrossRef]
- Ren, J.; Jiang, Y.H.; Liu, S.J.; Yu, F.; Pang, H.M. Anti-aging effects of different drying methods of Polygonum multiflorum on D-galactose-induced aging mice. Pharm. Clin. Res. 2024, 32, 109–114. [Google Scholar] [CrossRef]
- Zhu, C.; Li, Y.F.; Peng, F.; Yang, C.F.; Fu, Y.Y.; Tian, M. Study on Phytoestrogen-like Effect and Mechanism of Polygoni Multiflori Radix Preparata. Chin. Arch. Tradit. Chin. Med. 2020, 38, 216–220. [Google Scholar] [CrossRef]
- Pan, F.Z.; Chen, M.X.; Yi, B.; Xue, Y.H.; Wang, Q.P.; Wu, F.Y.; Ji, E.H.; Wu, H.W.; Xu, J. Stewed Polygoni Multiflori Radix Treats androgenic Alopecia in Mice by Activating Wnt/β-catenin Signaling Pathway. Chin. J. Exp. Tradit. Med. Formulae 2024, 30, 246–253. [Google Scholar] [CrossRef]
- Liu, X.; Yang, C.; Deng, Y.; Liu, P.; Yang, H.; Du, X.; Du, Y. Polygoni Multiflori Radix Preparat Delays Skin Aging by Inducing Mitophagy. Biomed. Res. Int. 2021, 2021, 5847153. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Li, J.; Tang, W.; Li, Y.; Lin, C.; Peng, D.; Yang, C. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside (TSG) from Polygonum multiflorum Thunb.: A Systematic Review on Anti-Aging. Int. J. Mol. Sci. 2025, 26, 3381. https://doi.org/10.3390/ijms26073381
Zhu C, Li J, Tang W, Li Y, Lin C, Peng D, Yang C. 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside (TSG) from Polygonum multiflorum Thunb.: A Systematic Review on Anti-Aging. International Journal of Molecular Sciences. 2025; 26(7):3381. https://doi.org/10.3390/ijms26073381
Chicago/Turabian StyleZhu, Can, Jinhong Li, Wenchao Tang, Yaofeng Li, Chang Lin, Danhong Peng, and Changfu Yang. 2025. "2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside (TSG) from Polygonum multiflorum Thunb.: A Systematic Review on Anti-Aging" International Journal of Molecular Sciences 26, no. 7: 3381. https://doi.org/10.3390/ijms26073381
APA StyleZhu, C., Li, J., Tang, W., Li, Y., Lin, C., Peng, D., & Yang, C. (2025). 2,3,5,4′-Tetrahydroxystilbene-2-O-β-D-glucoside (TSG) from Polygonum multiflorum Thunb.: A Systematic Review on Anti-Aging. International Journal of Molecular Sciences, 26(7), 3381. https://doi.org/10.3390/ijms26073381