

The Light-Regulated SsMYB106 Transcription Factor Promotes Flavonoids in Spatholobus suberectus

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
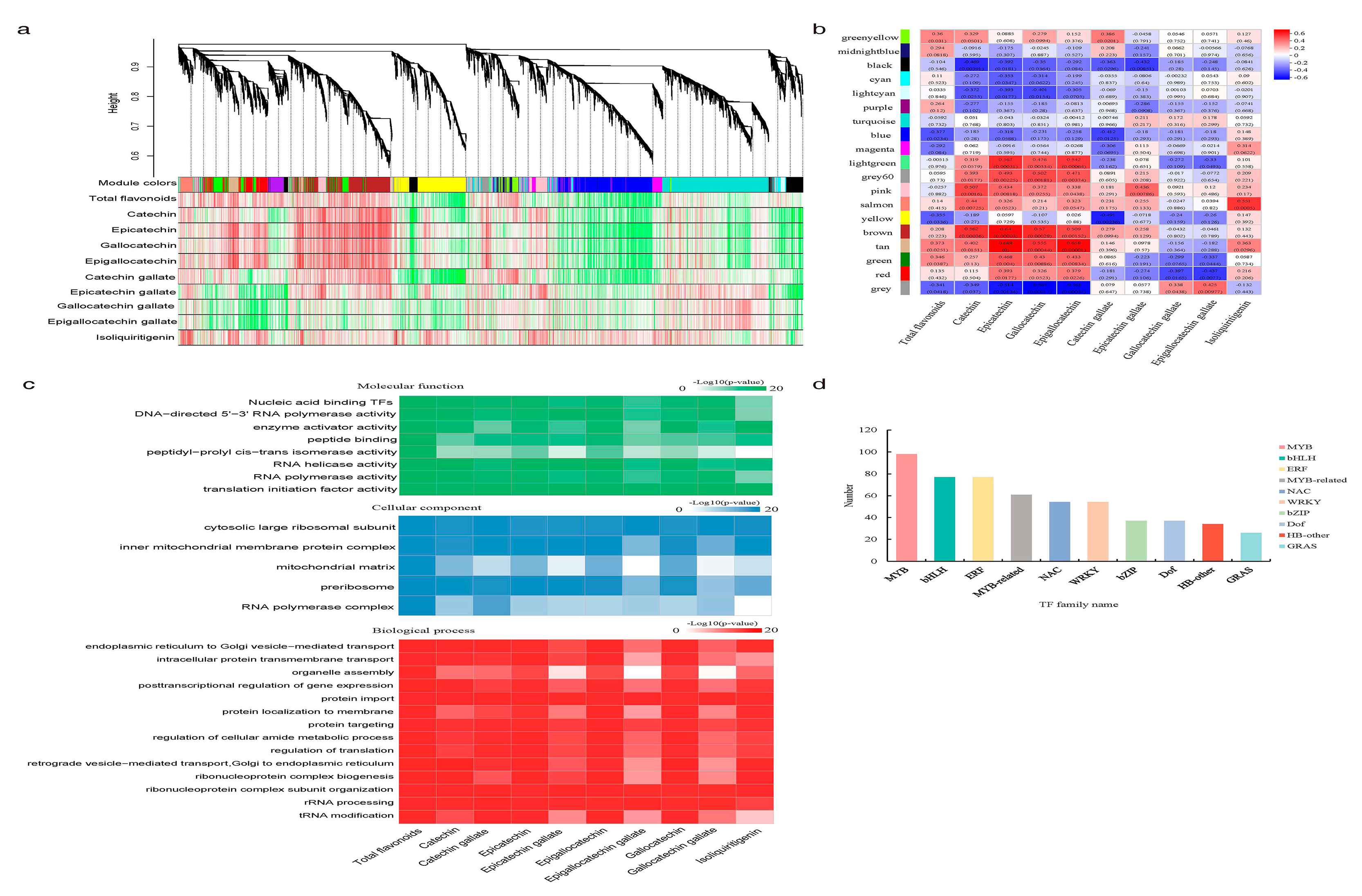
2.1. WGCNA Revealed MYB TFs Were Highly Associated with Light-Induced Flavonoid Content Changes in S. suberectus
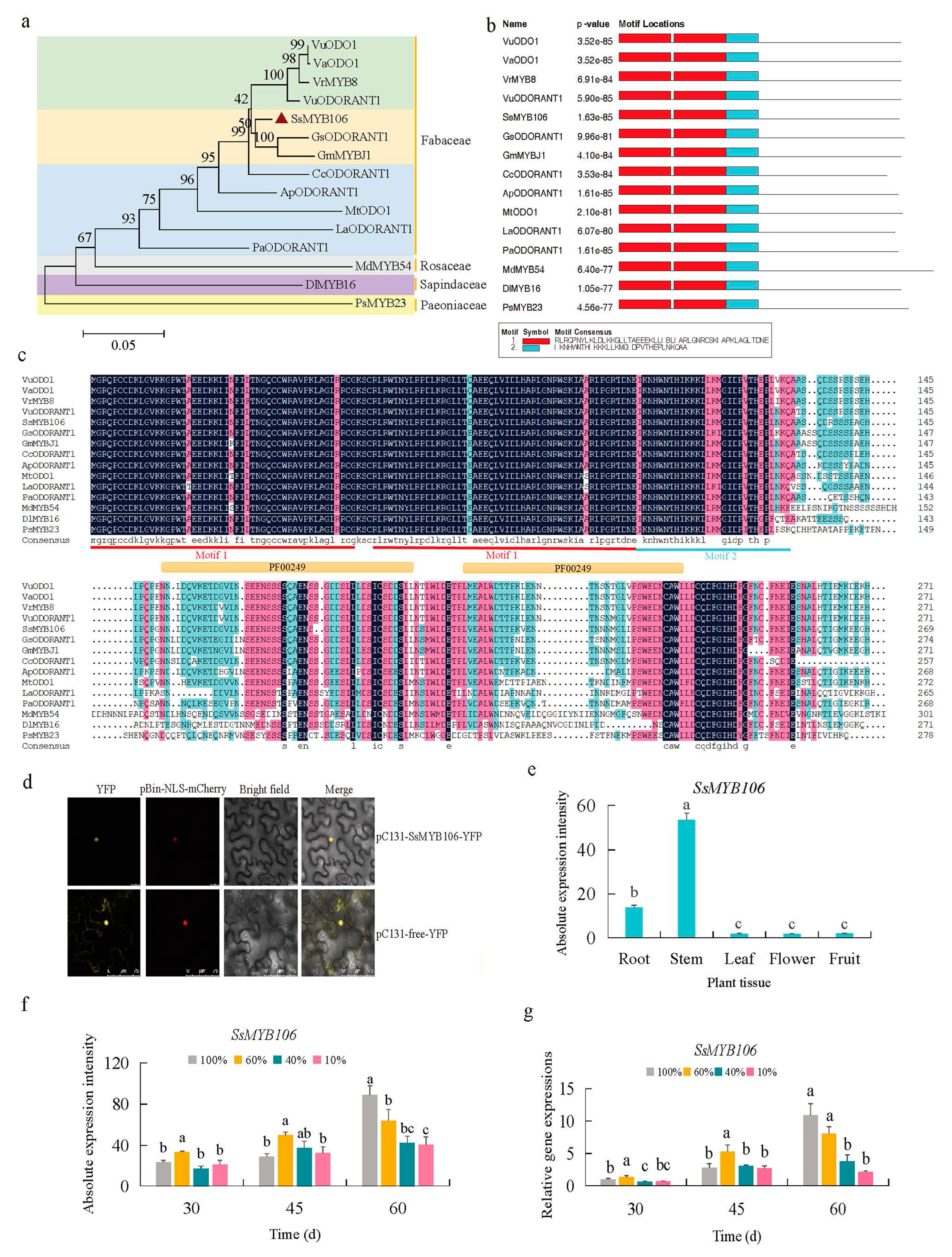
2.2. SsMYB106 Was Characterized as a Light-Regulated R2R3-MYB TF in S. suberectus
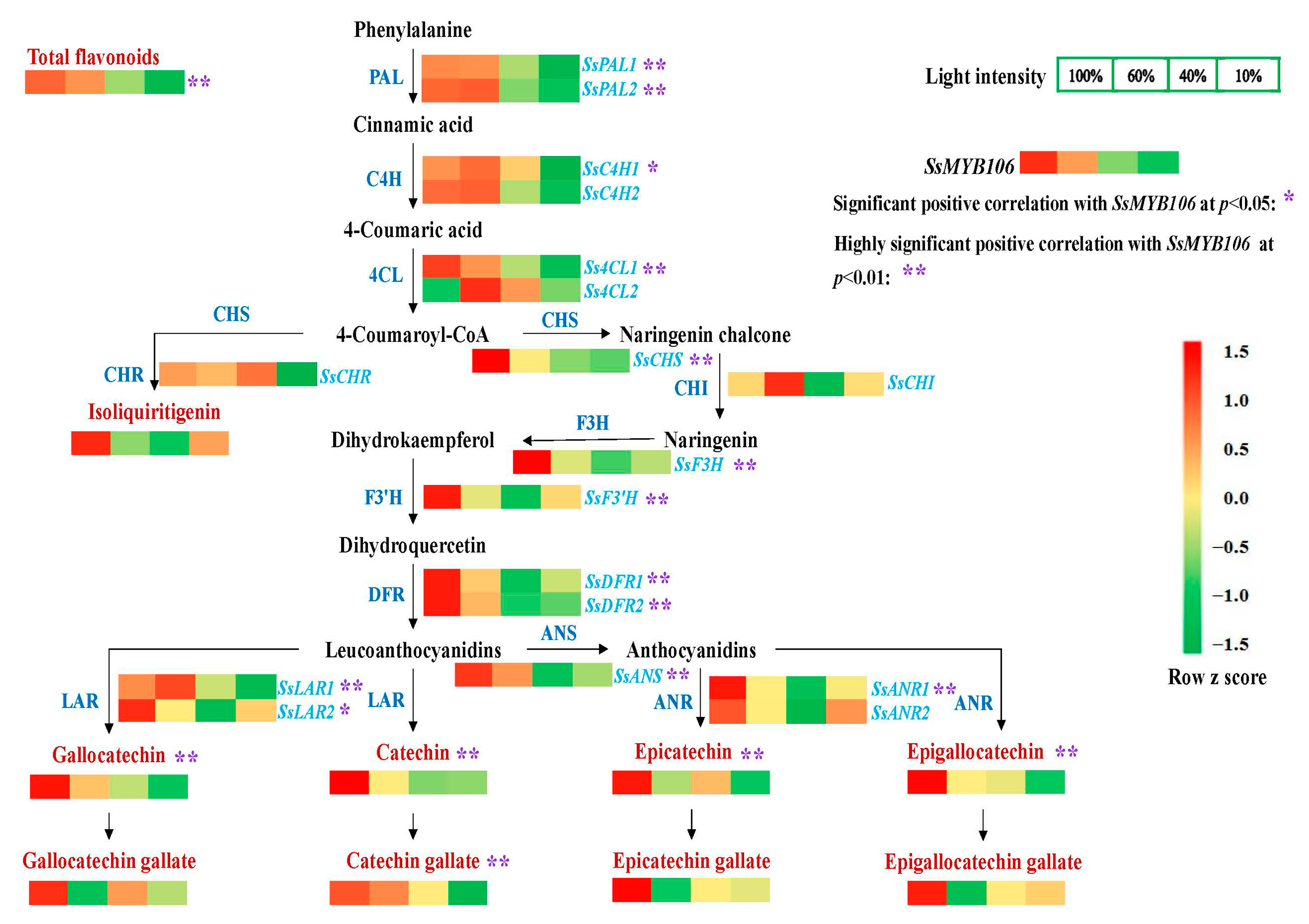
2.3. SsMYB106 Was Closely Related to the Light-Intensity-Induced Changes of Flavonoids Contents and Pathway Genes Expressions in S. suberectus
2.4. SsMYB106 Transient Overexpression Enhanced Flavonoids Accumulation, Increased All but One Pathway Genes Expressions in S. suberectus
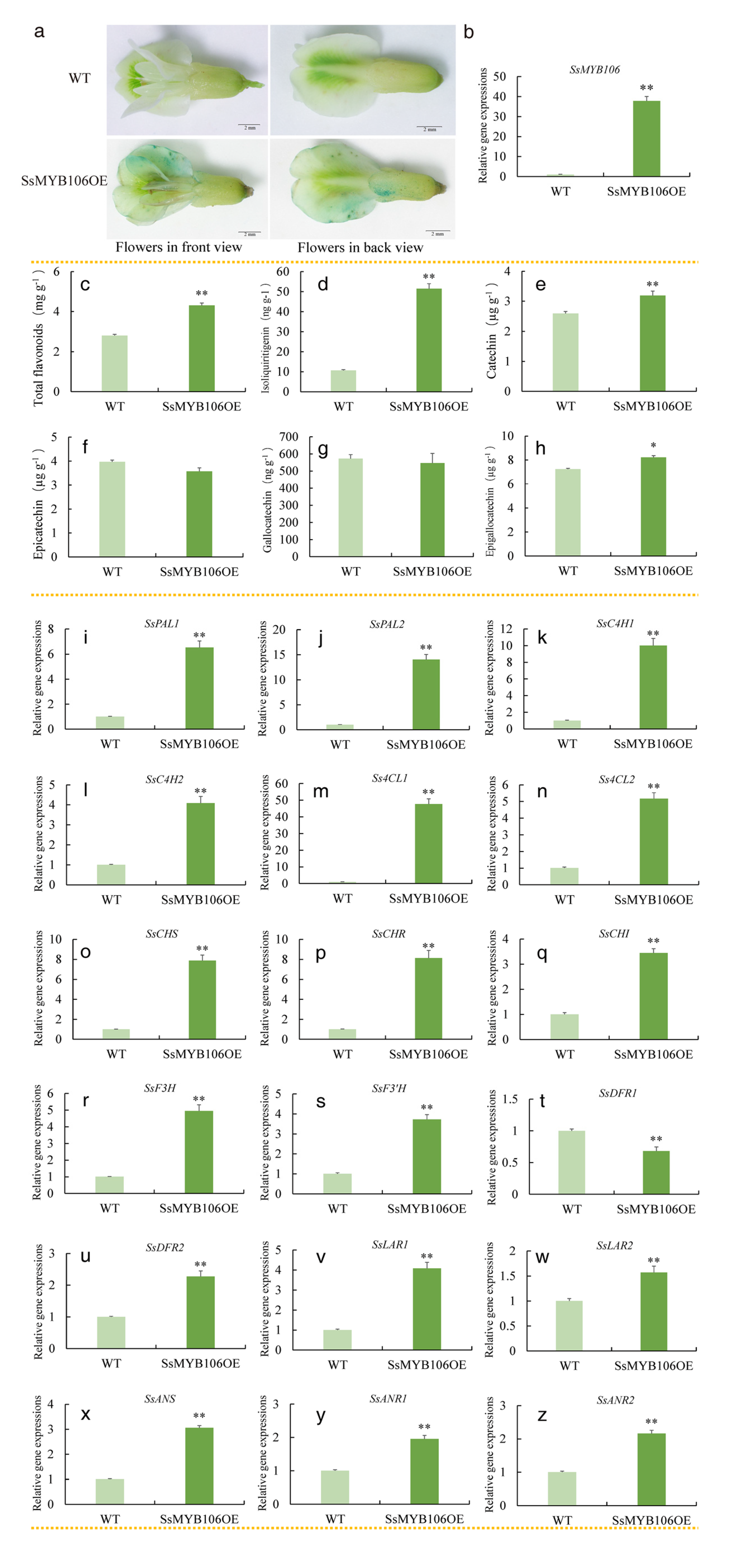
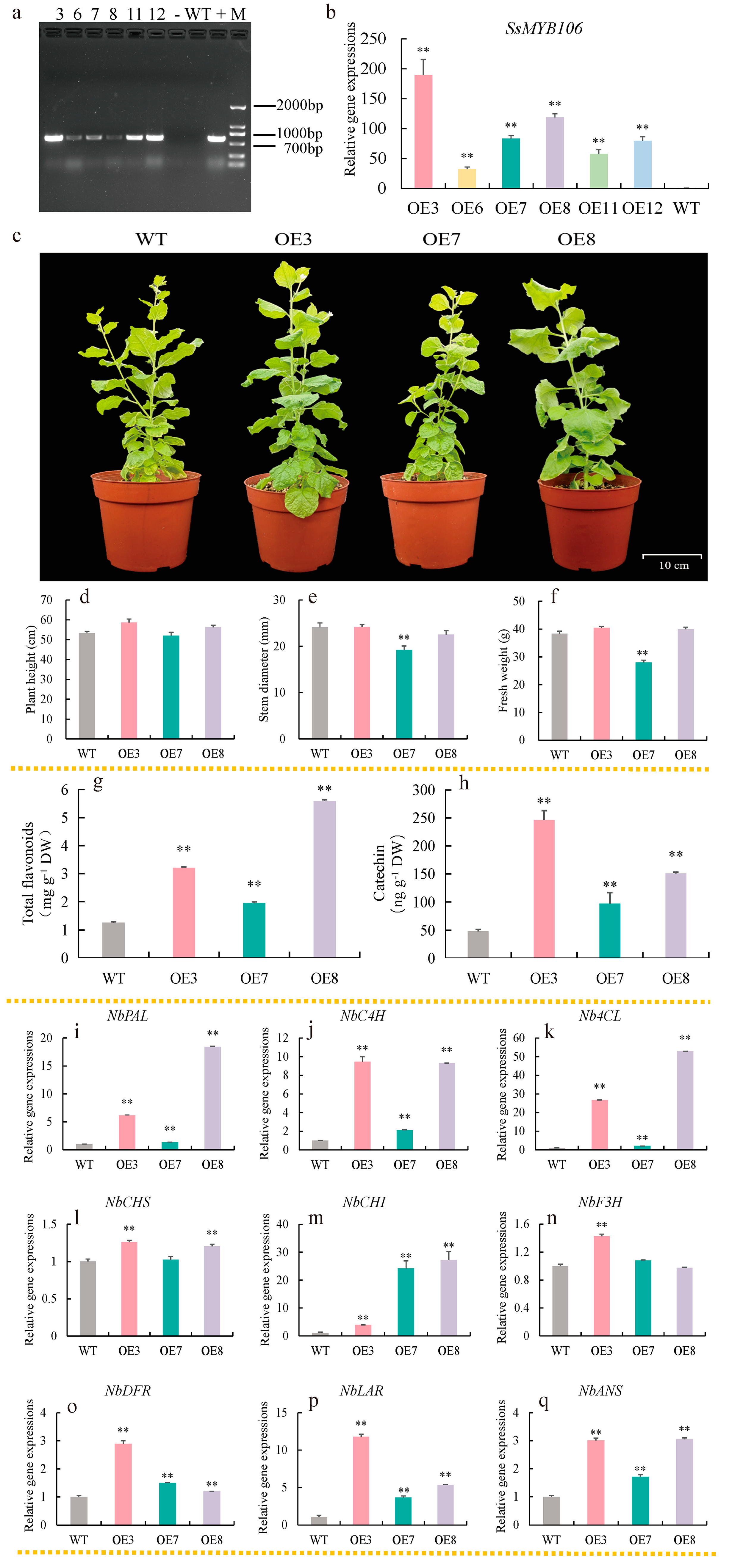
2.5. SsMYB106 Stable Overexpression Promoted Flavonoids Accumulation, Activated the Majority of Pathway Genes Expressions in N. benthamiana
3. Discussion
3.1. SsMYB106 Is Light-Regulated and Involved in Controlling the Light-Intensity-Induced Responses of S. suberectus
3.2. SsMYB106 Has Multifaceted Roles, Primarily Promotes Flavonoids Accumulation
3.2.1. First, SsMYB106 Overexpression Enhanced Flavonoids Accumulation
3.2.2. Second, SsMYB106 Overexpression Preferably Directed Metabolic Flux into Catechin Biosynthesis
In S. suberectus
In N. benthamiana
3.2.3. Third, SsMYB106 Overexpression Might Inhibit Lignin Biosynthesis
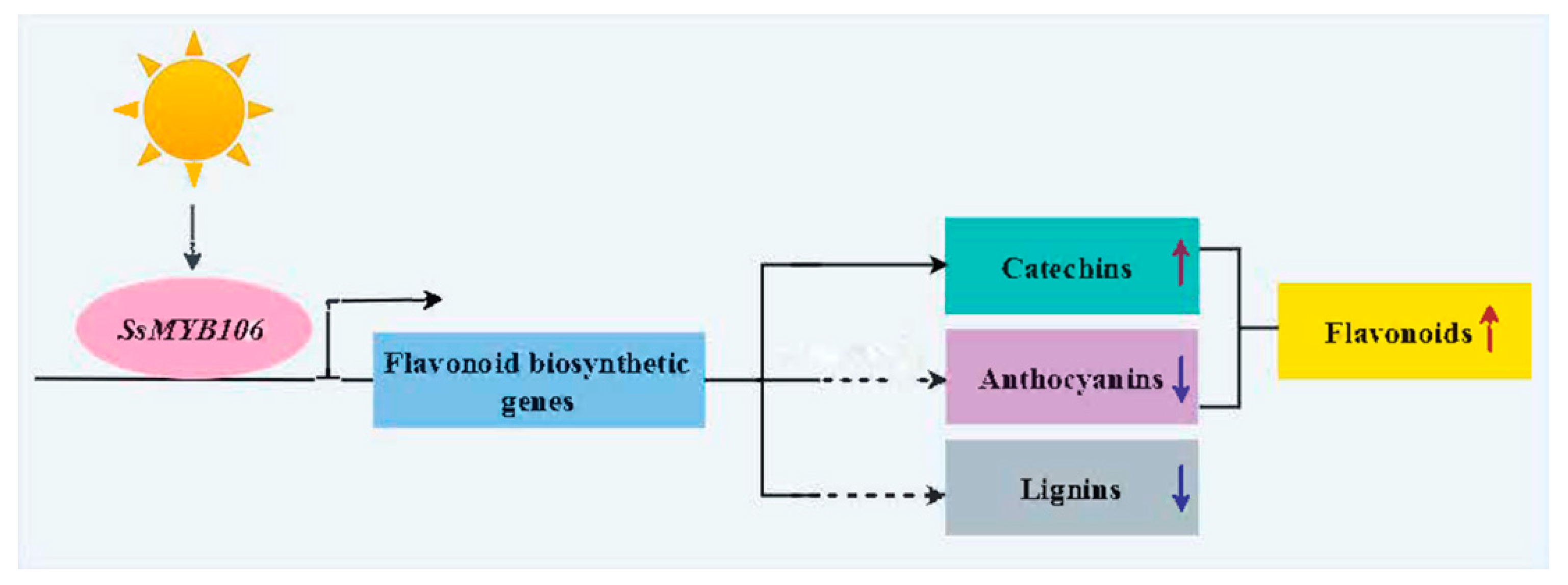
3.3. Proposed Model and Future Work
3.3.1. Proposed Model
3.3.2. Future Work
4. Materials and Methods
4.1. Plant Material
4.2. Light Treatments
4.3. Flavonoids Content Determination
4.4. Transcriptomic Gene Expression
4.5. RT-qPCR
4.6. WGCNA
4.7. Phylogeny Tree
4.8. Motif Discovery
4.9. Subcellular Localization
4.10. Gene Promoter CREs Analysis
4.11. SsMYB106 Transient Overexpression in S. suberectus Flowers
4.12. SsMYB106 Stable Overexpression in N. benthamiana
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pan, Y.; Luo, X.; Gong, P. Spatholobi caulis: A systematic review of its traditional uses, chemical constituents, biological activities and clinical applications. J. Ethnopharmacol. 2023, 317, 116854. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Ganesan, K.; Liu, Q.; Chen, J. A review of the pharmacological potential of Spatholobus suberectus Dunn on cancer. Cells 2022, 11, 2885. [Google Scholar] [CrossRef]
- Araguirang, G.E.; Richter, A.S. Activation of anthocyanin biosynthesis in high light—What is the initial signal? New Phytol. 2022, 236, 2037–2043. [Google Scholar] [CrossRef]
- Liu, W.; Feng, Y.; Yu, S.; Fan, Z.; Li, X.; Li, J.; Yin, H. The flavonoid biosynthesis network in plants. Int. J. Mol. Sci. 2021, 22, 12824. [Google Scholar] [CrossRef]
- Jiang, L.; Gao, Y.; Han, L.; Zhang, W.; Fan, P. Designing plant flavonoids: Harnessing transcriptional regulation and enzyme variation to enhance yield and diversity. Front. Plant Sci. 2023, 14, 1220062. [Google Scholar] [CrossRef] [PubMed]
- Khusnutdinov, E.; Sukhareva, A.; Panfilova, M.; Mikhaylova, E. Anthocyanin biosynthesis genes as model genes for genome editing in plants. Int. J. Mol. Sci. 2021, 22, 8752. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Wei, K.; Cui, Z.; Liang, Y.; Li, M.; Gu, L.; Yang, C.; Zhou, X.; Li, L.; Xu, W.; et al. Comparative genomics of Spatholobus suberectus and insight into flavonoid biosynthesis. Front. Plant Sci. 2020, 11, 528108. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Liang, Y.; Wei, G.; Wei, F.; Wei, K.; Chen, X. Shade responses and resistant mechanisms in Spatholobus suberectus. Heliyon 2024, 10, e28077. [Google Scholar] [CrossRef]
- Yan, H.; Pei, X.; Zhang, H.; Li, X.; Zhang, X.; Zhao, M.; Chiang, V.L.; Sederoff, R.R.; Zhao, X. MYB-mediated regulation of anthocyanin biosynthesis. Int. J. Mol. Sci. 2021, 22, 3103. [Google Scholar] [CrossRef]
- Jiang, L.; Yue, M.; Liu, Y.; Zhang, N.; Lin, Y.; Zhang, Y.; Wang, Y.; Li, M.; Luo, Y.; Zhang, Y.; et al. A novel R2R3-MYB transcription factor FaMYB5 positively regulates anthocyanin and proanthocyanidin biosynthesis in cultivated strawberries (Fragaria × ananassa). Plant Biotechnol. J. 2023, 21, 1140–1158. [Google Scholar] [CrossRef]
- Cao, Y.; Li, K.; Li, Y.; Zhao, X.; Wang, L. MYB transcription factors as regulators of secondary metabolism in plants. Biology 2020, 9, 61. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, D.; Wang, D.; Sun, M.; Zhang, G.; Wu, Y.; Zhang, Y.; Cheng, B. Study on the causes of changes in colour during Hibiscus syriacus flowering based on transcriptome and metabolome analyses. BMC Plant Biol. 2024, 24, 431. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Huang, Y.; Dong, Q.; Wan, M.; Wang, A.; Chen, Y.; Li, C.; Wu, Q.; Chen, H.; Zhao, H. FtMYB6, a light-induced SG7 R2R3-MYB transcription factor, promotes flavonol biosynthesis in tartary buckwheat (Fagopyrum tataricum). J. Agric. Food. Chem. 2020, 68, 13685–13696. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, G.; Tang, Q.; Song, W.; Gao, Q.; Xiang, G.; Li, X.; Liu, G.; Fan, W.; Li, X.; et al. EbMYBP1, a R2R3-MYB transcription factor, promotes flavonoid biosynthesis in Erigeron breviscapus. Front. Plant Sci. 2022, 13, 946827. [Google Scholar] [CrossRef]
- Naik, J.; Misra, P.; Trivedi, P.K.; Pandey, A. Molecular components associated with the regulation of flavonoid biosynthesis. Plant Sci. 2022, 317, 111196. [Google Scholar] [CrossRef]
- Wang, P.; Ma, G.; Zhang, L.; Li, Y.; Fu, Z.; Kan, X.; Han, Y.; Wang, H.; Jiang, X.; Liu, Y.; et al. A sucrose-induced MYB (SIMYB) transcription factor promoting proanthocyanidin accumulation in the tea plant ( Camellia sinensis). J. Agric. Food. Chem. 2019, 67, 1418–1428. [Google Scholar] [CrossRef]
- Wang, X.C.; Wu, J.; Guan, M.L.; Zhao, C.H.; Geng, P.; Zhao, Q. Arabidopsis MYB4 plays dual roles in flavonoid biosynthesis. Plant J. 2020, 101, 637–652. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, K.; Qi, Y.; Lv, G.; Ren, X.; Liu, Z.; Ma, F. Transcriptional regulation of anthocyanin synthesis by MYB-bHLH-WDR complexes in Kiwifruit (Actinidia chinensis). J. Agric. Food. Chem. 2021, 69, 3677–3691. [Google Scholar] [CrossRef]
- Qin, S.; Wei, F.; Liang, Y.; Tang, D.; Lin, Q.; Miao, J.; Wei, K. Genome-wide analysis of the R2R3-MYB gene family in Spatholobus suberectus and identification of its function in flavonoid biosynthesis. Front. Plant Sci. 2023, 14, 1219019. [Google Scholar] [CrossRef]
- Alexander, R.D.; Wendelboe-Nelson, C.; Morris, P.C. The barley transcription factor HvMYB1 is a positive regulator of drought tolerance. Plant Physiol. Biochem. 2019, 142, 246–253. [Google Scholar] [CrossRef]
- Nabi, B.G.; Mukhtar, K.; Ahmed, W.; Manzoor, M.F.; Ranjha, M.M.A.N.; Kieliszek, M.; Bhat, Z.F.; Aadil, R.M.J.F.B. Natural pigments: Anthocyanins, carotenoids, chlorophylls, and betalains as food colorants in food products. Food Biosci. 2023, 52, 102403. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Yan, Q.; Jia, Y.; Dong, F.; Shen, Y.; Li, F.; Zhang, M. Metabolomics uncovers the mechanisms of nitrogen response to anthocyanins synthesis and grain quality of colored grain wheat (Triticum aestivum L.). J. Agric. Food. Chem. 2024, 72, 19003–19015. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Xu, Z.; Fang, Z.; Zheng, X.; Cao, L.; Kang, T.; Xu, Y.; Zhang, X.; Zhan, Q.; Wang, H.; et al. NAC transcription factor PpNAP4 promotes chlorophyll degradation and anthocyanin synthesis in the skin of peach fruit. J. Agric. Food. Chem. 2024, 72, 19826–19837. [Google Scholar] [CrossRef]
- Marin-Recinos, M.F.; Pucker, B. Genetic factors explaining anthocyanin pigmentation differences. BMC Plant Biol. 2024, 24, 627. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Li, M.; Ma, C.; Li, X.; Bai, P.; Lin, A.; Wang, C.; Zhang, L.; Kuang, H.; Lian, H. Loss-of-function mutation in anthocyanidin reductase activates the anthocyanin synthesis pathway in strawberry. Mol. Hortic. 2024, 4, 33. [Google Scholar] [CrossRef]
- Zhang, J.; Han, N.; Zhao, A.; Wang, Z.; Wang, D. ZbMYB111 expression positively regulates ZbUFGT-mediated anthocyanin biosynthesis in Zanthoxylum bungeanum with the involvement of ZbbHLH2. J. Agric. Food. Chem. 2024, 72, 16941–16954. [Google Scholar] [CrossRef]
- Zhao, D.; Tao, J. Recent advances on the development and regulation of flower color in ornamental plants. Front. Plant Sci. 2015, 6, 261. [Google Scholar] [CrossRef]
- Li, C.; Yu, W.; Xu, J.; Lu, X.; Liu, Y. Anthocyanin biosynthesis induced by MYB transcription factors in plants. Int. J. Mol. Sci. 2022, 23, 11701. [Google Scholar] [CrossRef]
- Moss, S.M.A.; Zhou, Y.; Butelli, E.; Waite, C.N.; Yeh, S.M.; Cordiner, S.B.; Harris, N.N.; Copsey, L.; Schwinn, K.E.; Davies, K.M.; et al. Painted flowers: Eluta generates pigment patterning in Antirrhinum. New Phytol. 2024, 243, 738–752. [Google Scholar] [CrossRef]
- Pratyusha, D.S.; Sarada, D.V.L. MYB transcription factors-master regulators of phenylpropanoid biosynthesis and diverse developmental and stress responses. Plant Cell Rep. 2022, 41, 2245–2260. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Zhang, S.; Zhao, Q.; Wu, Y.; Li, H. The CsMYB123 and CsbHLH111 are involved in drought stress-induced anthocyanin biosynthesis in Chaenomeles speciosa. Mol. Hortic. 2023, 3, 25. [Google Scholar] [CrossRef]
- Yang, G.; Xue, Z.; Lin-Wang, K.; Chen, G.; Zhao, Y.; Chang, Y.; Xu, S.; Sun, M.; Xue, C.; Li, J.; et al. An ‘activator-repressor’ loop controls the anthocyanin biosynthesis in red-skinned pear. Mol. Hortic. 2024, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Bulanov, A.N.; Andreeva, E.A.; Tsvetkova, N.V.; Zykin, P.A. Regulation of flavonoid biosynthesis by the MYB-bHLH-WDR (MBW) complex in plants and its specific features in cereals. Int. J. Mol. Sci. 2025, 26, 734. [Google Scholar] [CrossRef]
- Shan, X.; Li, Y.; Yang, S.; Yang, Z.; Qiu, M.; Gao, R.; Han, T.; Meng, X.; Xu, Z.; Wang, L.; et al. The spatio-temporal biosynthesis of floral flavonols is controlled by differential phylogenetic MYB regulators in Freesia hybrida. New Phytol. 2020, 228, 1864–1879. [Google Scholar] [CrossRef]
- Liang, B.; Ye, X.; Li, H.; Li, F.; Wang, S.; Jiang, C.; Wang, J.; Wang, P. Genome-wide identification and analysis of anthocyanidin reductase gene gamily in Lychee (Litchi chinensis Sonn.). Genes 2024, 15, 757. [Google Scholar] [CrossRef]
- Eichenberger, M.; Schwander, T.; Hüppi, S.; Kreuzer, J.; Mittl, P.R.E.; Peccati, F.; Jiménez-Osés, G.; Naesby, M.; Buller, R.M. The catalytic role of glutathione transferases in heterologous anthocyanin biosynthesis. Nat. Catal. 2023, 6, 927–938. [Google Scholar] [CrossRef]
- Tohge, T.; Nishiyama, Y.; Hirai, M.Y.; Yano, M.; Nakajima, J.; Awazuhara, M.; Inoue, E.; Takahashi, H.; Goodenowe, D.B.; Kitayama, M.; et al. Functional genomics by integrated analysis of metabolome and transcriptome of Arabidopsis plants over-expressing an MYB transcription factor. Plant J. 2005, 42, 218–235. [Google Scholar] [CrossRef] [PubMed]
- Pucker, B.; Selmar, D. Biochemistry and molecular basis of intracellular flavonoid transport in plants. Plants 2022, 11, 963. [Google Scholar] [CrossRef]
- Enaru, B.; Drețcanu, G.; Pop, T.D.; Stǎnilǎ, A.; Diaconeasa, Z. Anthocyanins: Factors affecting their stability and degradation. Antioxidants 2021, 10, 1967. [Google Scholar] [CrossRef]
- Zhao, Y.W.; Wang, C.K.; Huang, X.Y.; Hu, D.G. Anthocyanin stability and degradation in plants. Plant Signaling Behav. 2021, 16, 1987767. [Google Scholar] [CrossRef] [PubMed]
- LaFountain, A.M.; Yuan, Y.W. Repressors of anthocyanin biosynthesis. New Phytol. 2021, 231, 933–949. [Google Scholar] [CrossRef]
- Li, Z.; Geng, G.; Xie, H.; Zhou, L.; Wang, L.; Qiao, F. Metabolomic and transcriptomic reveal flavonoid biosynthesis and regulation mechanism in Phlomoides rotata from different habitats. Genomics 2024, 116, 110850. [Google Scholar] [CrossRef]
- Luo, P.; Ning, G.; Wang, Z.; Shen, Y.; Jin, H.; Li, P.; Huang, S.; Zhao, J.; Bao, M. Disequilibrium of flavonol synthase and dihydroflavonol-4-reductase expression associated tightly to white vs. red color flower formation in plants. Front. Plant Sci. 2015, 6, 1257. [Google Scholar] [CrossRef]
- Miosic, S.; Thill, J.; Milosevic, M.; Gosch, C.; Pober, S.; Molitor, C.; Ejaz, S.; Rompel, A.; Stich, K.; Halbwirth, H. Dihydroflavonol 4-reductase genes encode enzymes with contrasting substrate specificity and show divergent gene expression profiles in Fragaria species. PLoS ONE 2014, 9, e112707. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Fan, W.; Li, H.; Yang, J.; Huang, J.; Zhang, P. Functional characterization of Dihydroflavonol-4-reductase in anthocyanin biosynthesis of purple sweet potato underlies the direct evidence of anthocyanins function against abiotic stresses. PLoS ONE 2013, 8, e78484. [Google Scholar] [CrossRef]
- Fatihah, H.N.N.; Moñino López, D.; van Arkel, G.; Schaart, J.G.; Visser, R.G.F.; Krens, F.A. The ROSEA1 and DELILA transcription factors control anthocyanin biosynthesis in Nicotiana benthamiana and Lilium flowers. Sci. Hortic. 2019, 243, 327–337. [Google Scholar] [CrossRef]
- Grützner, R.; König, K.; Horn, C.; Engler, C.; Laub, A.; Vogt, T.; Marillonnet, S. A transient expression tool box for anthocyanin biosynthesis in Nicotiana benthamiana. Plant Biotechnol. J. 2024, 22, 1238–1250. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shan, X.; Gao, R.; Han, T.; Zhang, J.; Wang, Y.; Kimani, S.; Wang, L.; Gao, X. MYB repressors and MBW activation complex collaborate to fine-tune flower coloration in Freesia hybrida. Commun. Biol. 2020, 3, 396. [Google Scholar] [CrossRef]
- Gui, J.; Lam, P.Y.; Tobimatsu, Y.; Sun, J.; Huang, C.; Cao, S.; Zhong, Y.; Umezawa, T.; Li, L. Fibre-specific regulation of lignin biosynthesis improves biomass quality in Populus. New Phytol. 2020, 226, 1074–1087. [Google Scholar] [CrossRef]
- Yoon, J.; Choi, H.; An, G. Roles of lignin biosynthesis and regulatory genes in plant development. J. Integr. Plant Biol. 2015, 57, 902–912. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Long, C.; Hao, X.; Zhang, R.; Li, C.; Song, Y. Identification of key genes involved in lignin and flavonoid accumulation during Tilia tuan seed maturation. Plant Cell Rep. 2024, 43, 205. [Google Scholar] [CrossRef]
- Miao, Y.; Duan, W.; Li, A.; Yuan, M.; Meng, J.; Wang, H.; Pan, L.; Sun, S.; Cui, G.; Shi, C.; et al. The MYB transcription factor PpMYB5 regulates Pp4CL1/Pp4CL2 expression to promote lignin biosynthesis of fruit russeting in the flat nectarine. Plant Cell Rep. 2024, 43, 231. [Google Scholar] [CrossRef]
- Shi, J.; Yan, X.; Sun, T.; Shen, Y.; Shi, Q.; Wang, W.; Bao, M.; Luo, H.; Nian, F.; Ning, G. Homeostatic regulation of flavonoid and lignin biosynthesis in phenylpropanoid pathway of transgenic tobacco. Gene 2022, 809, 146017. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Liu, X.Y.; Ni, R.; Fu, J.; Tan, H.; Cheng, A.X.; Lou, H.X. Molecular cloning and functional analysis of 4-coumarate: CoA ligases from Marchantia paleacea and their roles in lignin and flavanone biosynthesis. PLoS ONE 2024, 19, e0296079. [Google Scholar] [CrossRef]
- Zhang, C.; Zang, Y.; Liu, P.; Zheng, Z.; Ouyang, J. Characterization, functional analysis and application of 4-Coumarate: CoA ligase genes from Populus trichocarpa. J. Biotechnol. 2019, 302, 92–100. [Google Scholar] [CrossRef]
- Li, P.; Xia, E.; Fu, J.; Xu, Y.; Zhao, X.; Tong, W.; Tang, Q.; Tadege, M.; Fernie, A.R.; Zhao, J. Diverse roles of MYB transcription factors in regulating secondary metabolite biosynthesis, shoot development, and stress responses in tea plants (Camellia sinensis). Plant J. 2022, 110, 1144–1165. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wen, J.; Xia, Y.; Zhang, L.; Du, H. Evolution and functional diversification of R2R3-MYB transcription factors in plants. Hortic. Res. 2022, 9, uhac058. [Google Scholar] [CrossRef]
- Qin, S.; Wei, G.; Lin, Q.; Tang, D.; Li, C.; Tan, Z.; Yao, L.; Huang, L.; Wei, F.; Liang, Y. Analysis of the Spatholobus suberectus full-length transcriptome identified an R2R3-MYB transcription factor-encoding gene SsMYB158 that regulates flavonoid biosynthesis. Plant Physiol. Biochem. 2024, 214, 108929. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinf. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 1994, 2, 28–36. [Google Scholar] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L.; et al. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef] [PubMed]
- Higo, K.; Ugawa, Y.; Iwamoto, M.; Korenaga, T. Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, S.; Liang, Y.; Wei, F.; Wei, G.; Lin, Q.; Chen, X. The Light-Regulated SsMYB106 Transcription Factor Promotes Flavonoids in Spatholobus suberectus. Int. J. Mol. Sci. 2025, 26, 5292. https://doi.org/10.3390/ijms26115292
Qin S, Liang Y, Wei F, Wei G, Lin Q, Chen X. The Light-Regulated SsMYB106 Transcription Factor Promotes Flavonoids in Spatholobus suberectus. International Journal of Molecular Sciences. 2025; 26(11):5292. https://doi.org/10.3390/ijms26115292
Chicago/Turabian StyleQin, Shuangshuang, Ying Liang, Fan Wei, Guili Wei, Quan Lin, and Xiaoying Chen. 2025. "The Light-Regulated SsMYB106 Transcription Factor Promotes Flavonoids in Spatholobus suberectus" International Journal of Molecular Sciences 26, no. 11: 5292. https://doi.org/10.3390/ijms26115292
APA StyleQin, S., Liang, Y., Wei, F., Wei, G., Lin, Q., & Chen, X. (2025). The Light-Regulated SsMYB106 Transcription Factor Promotes Flavonoids in Spatholobus suberectus. International Journal of Molecular Sciences, 26(11), 5292. https://doi.org/10.3390/ijms26115292