
Genome-Wide Identification of GmARF9b/GmARF2a Negatively Regulate Root Growth in Soybean



Abstract
1. Introductions
2. Results
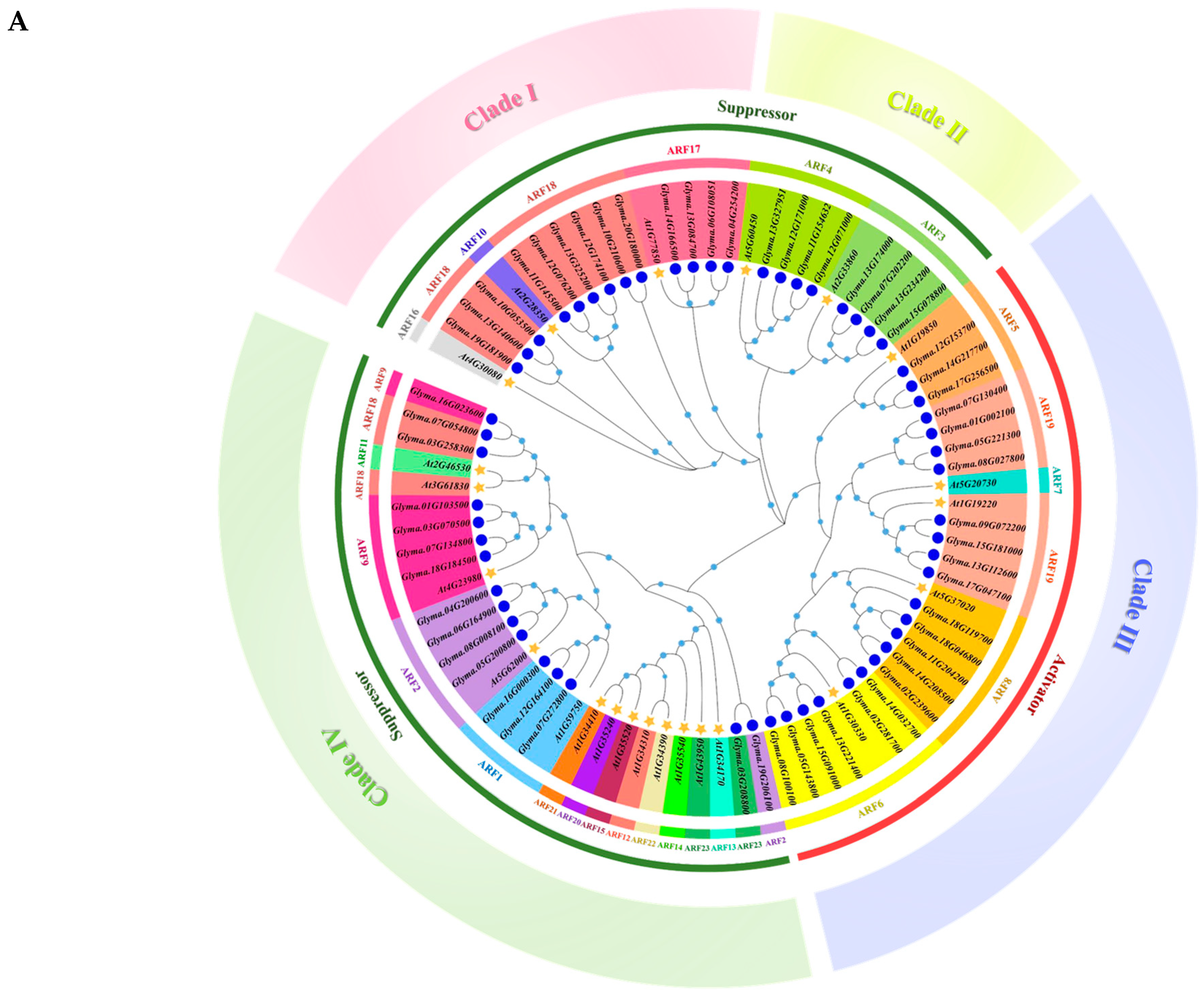
2.1. Phylogenetic, Classification, and Gene Haplotype Analysis of GmARFs
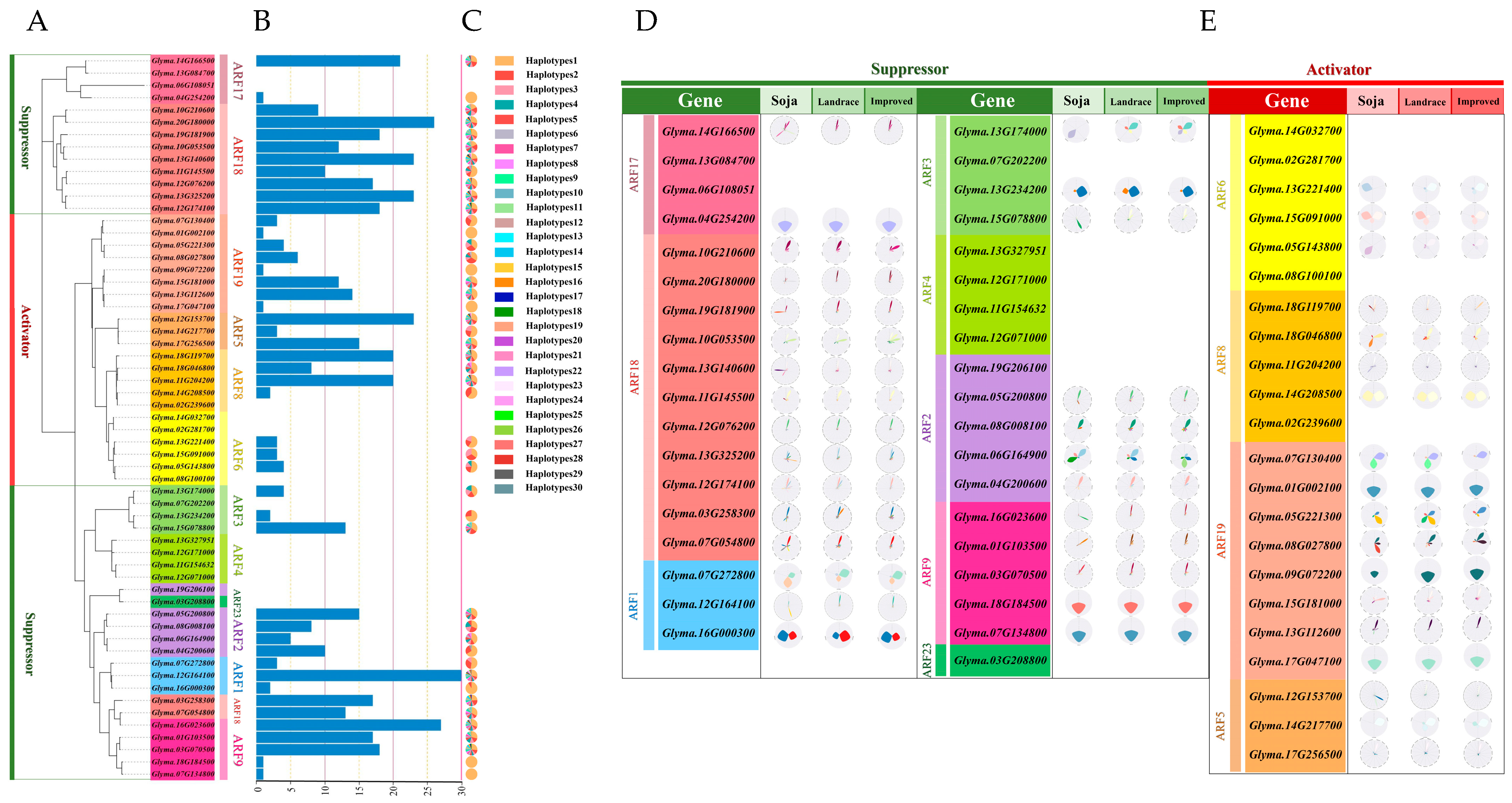
2.2. Analysis of the Association Between GmARF Gene Haplotypes and Natural Selection
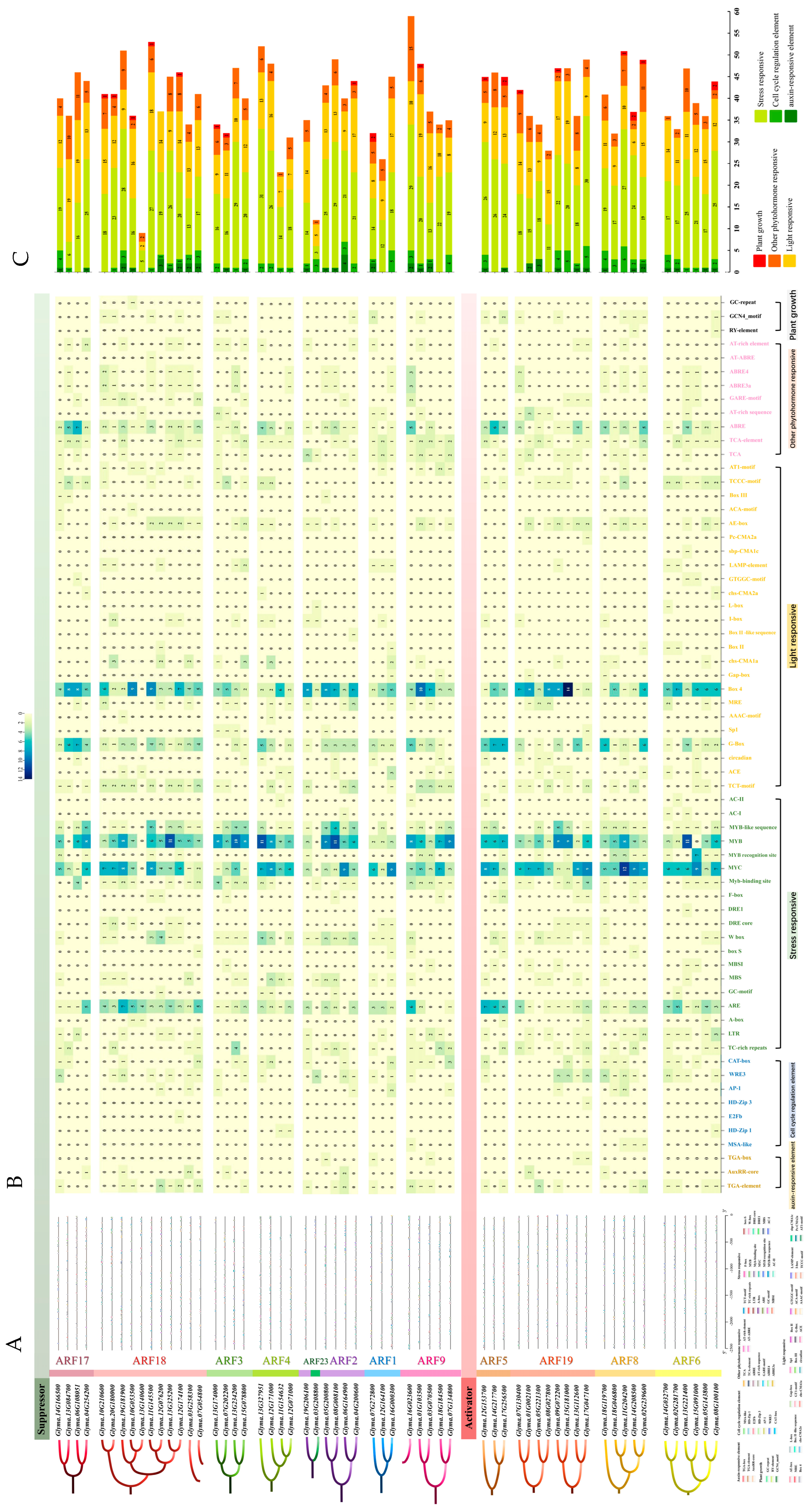
2.3. Gene Structure and Cis-Regulatory Element Analysis of GmARF Members
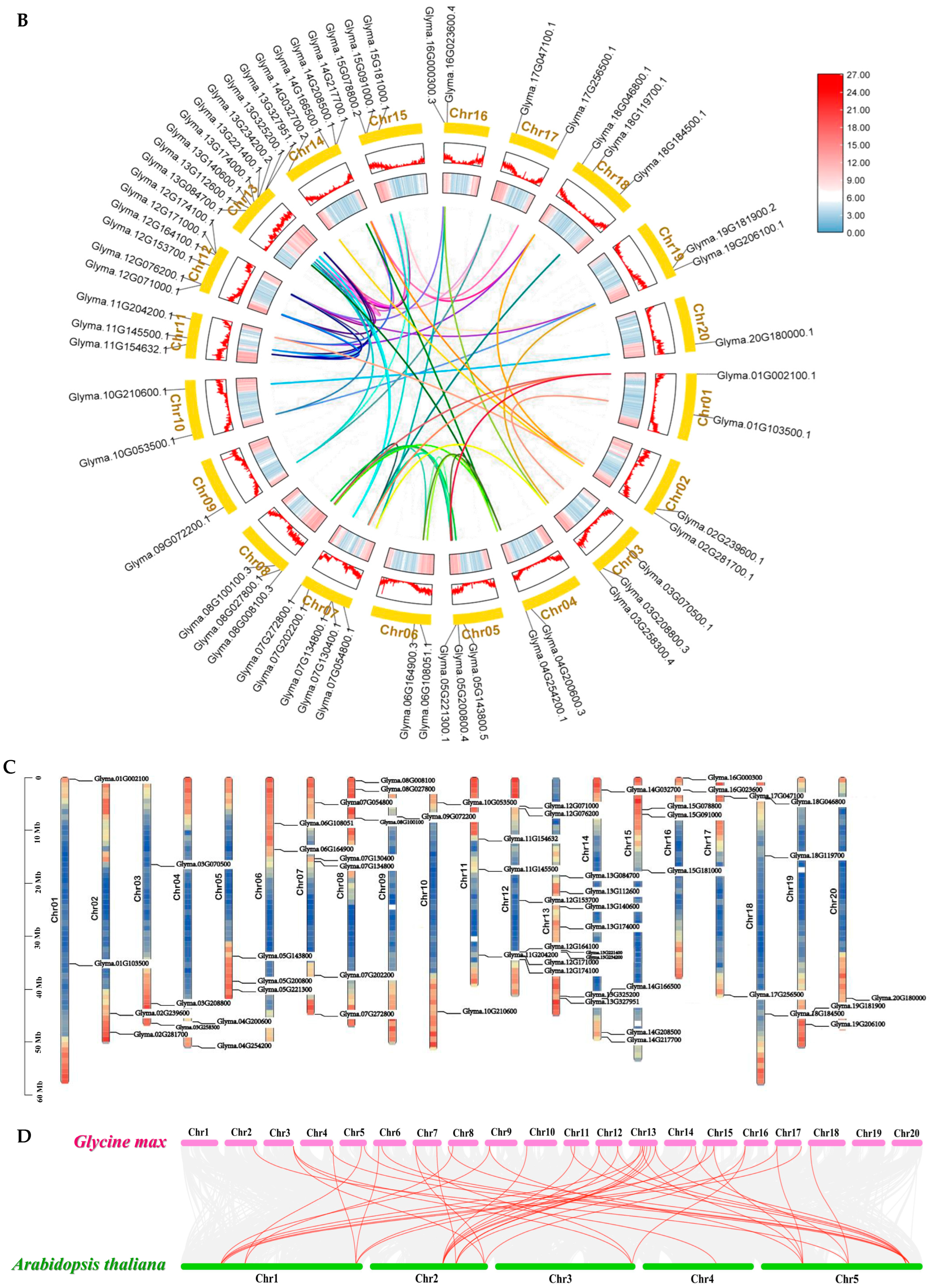
2.4. Whole-Genome Characterization of GmARF Genes in Soybean
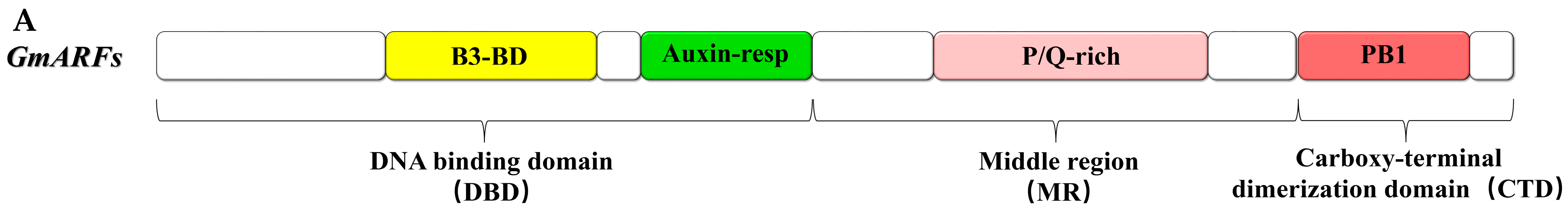
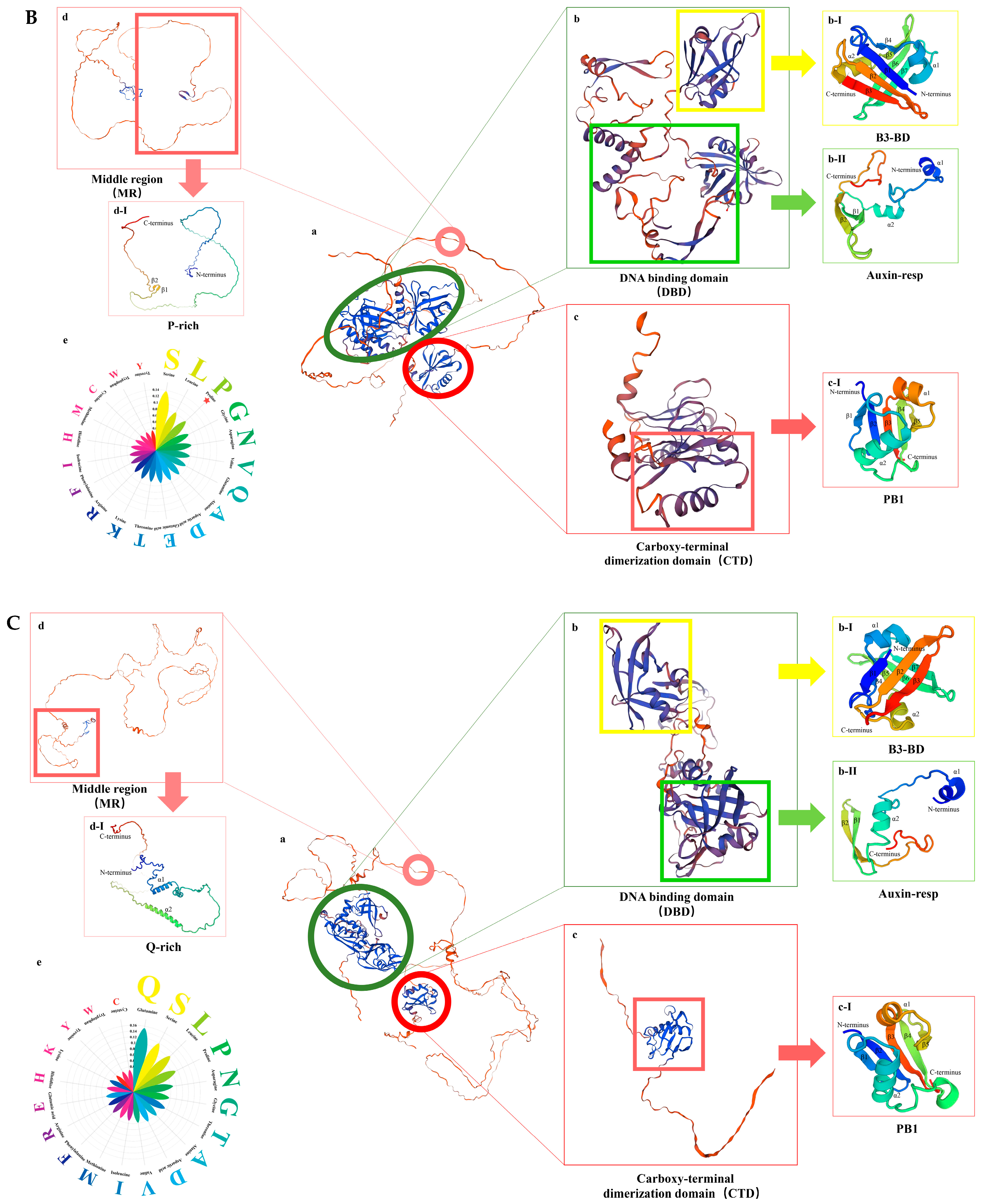
2.5. Conserved Motifs and Protein Structural Analysis of GmARF Genes
2.6. Cis-Regulatory Element Analysis of GmARF Genes
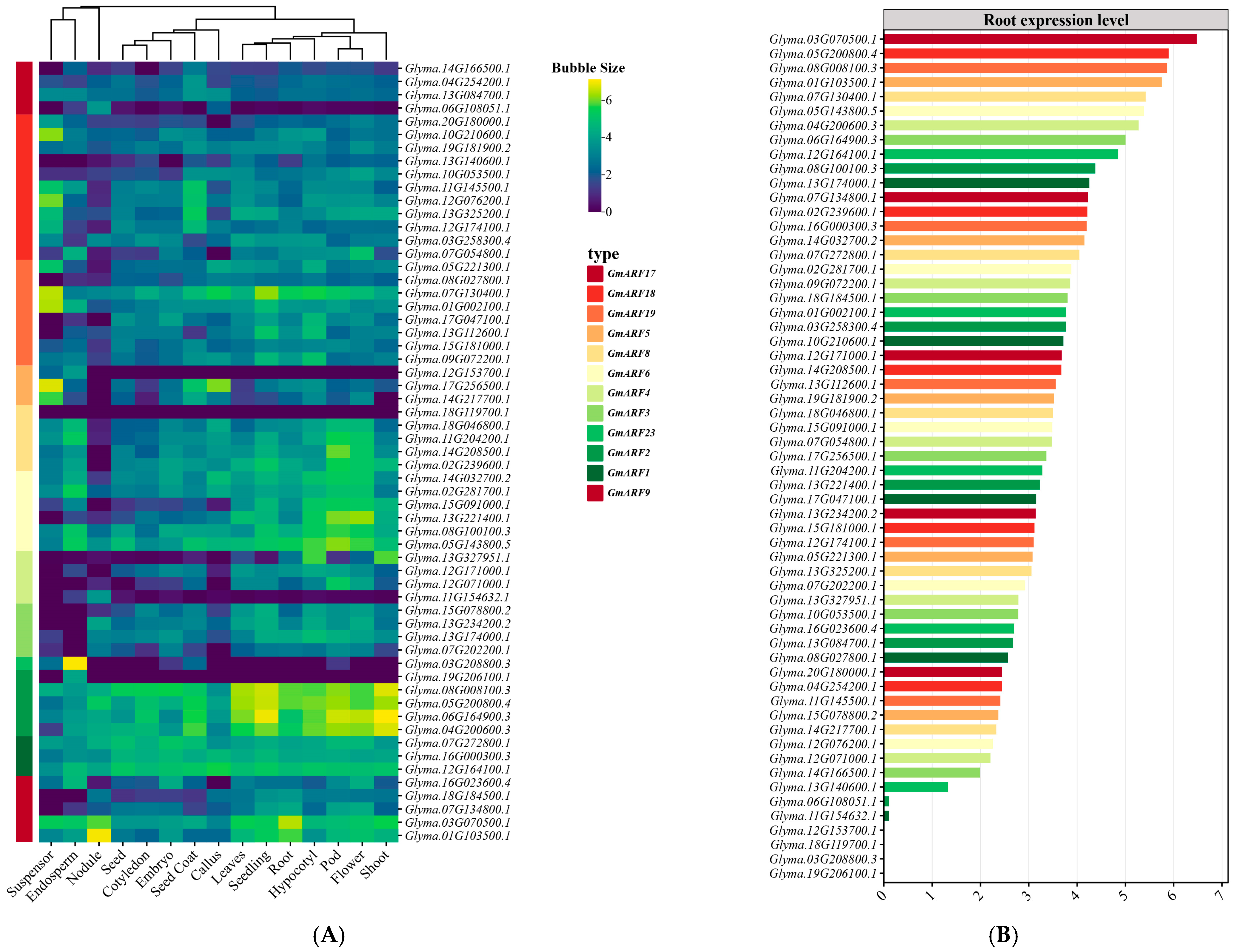
2.7. Expression Analysis of GmARF Genes
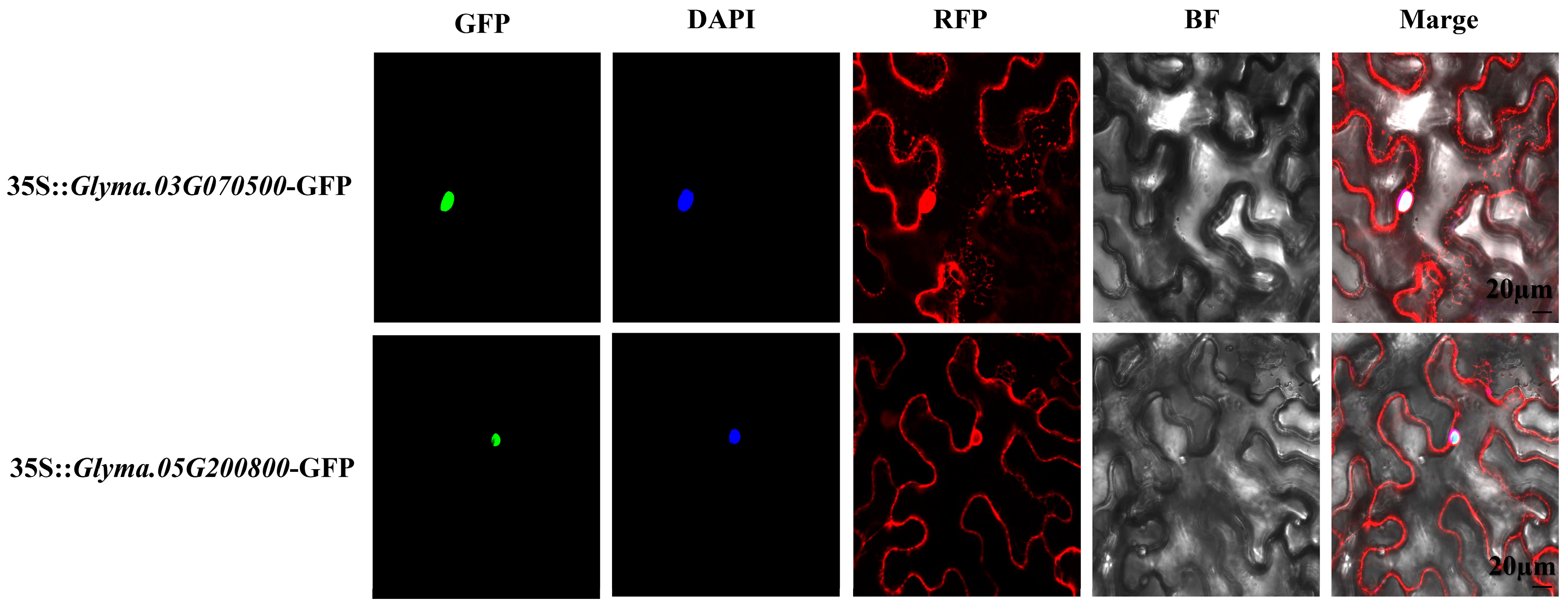
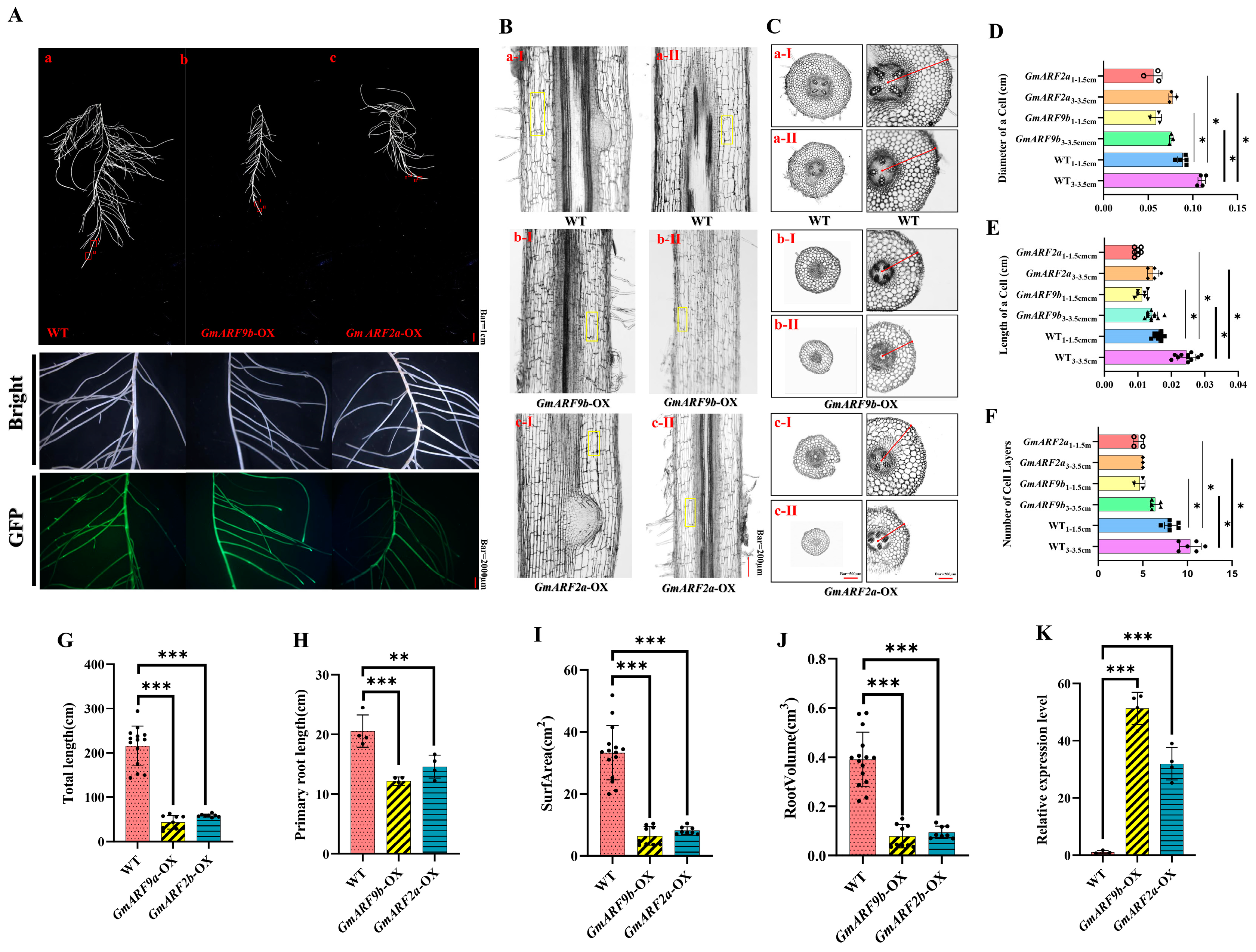
2.8. Functional Study of GmARF9b (Glyma.03G070500) and GmARF2a (Glyma.05G200800) in Soybean
3. Discussion
3.1. Phylogenetic Analysis, Classification, and Gene Haplotype of GmARF Genes
3.2. Dynamic Evolution of GmARF Gene Haplotypes Under Natural Selection in Soybean
3.3. Divergence of Conserved Motifs in GmARF Protein Domains
3.4. Biological Function Analysis of GmARF9b (Glyma.03G070500) and GmARF2a (Glyma.05G200800) in Glycine Max
4. Materials and Methods
4.1. Phylogenetic Tree Analysis of the ARF Gene Family
4.2. Chromosome Distribution and Gene Duplication Analysis of ARFs
4.3. Haplotype Analysis of GmARF Gene Family
4.4. Gene Structure and Protein Conserved Domain Prediction of GmARFs
4.5. Whole-Genome Characterization Analysis of the GmARF Gene Family in Soybean
4.6. Protein Structure Prediction and Motif Analysis of the GmARFs Gene Family in Soybean
4.7. Cis-Regulatory Element Analysis of the GmARF Gene Family in Soybean
4.8. Analysis of Gene Expression Patterns of the GmARF Family in Soybean
4.9. Acquisition of Transgenic Hairy Roots Overexpressing GmARF9b (Glyma.03G070500) and GmARF2a (Glyma.05G200800) in Soybean
4.10. Tissue Localization Analysis of GmARF9b (Glyma.03G070500) and GmARF2a (Glyma.05G200800) in Soybean
4.11. Subcellular Localization of GmARF9b (Glyma.03G070500) and GmARF2a (Glyma.05G200800) in Soybean
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Tian, H.Y.; De, S.I.; Ding, Z.J. Shaping a root system: Regulating lateral versus primary root growth. Trends Plant Sci. 2014, 19, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Eva, B.; Agnieszka, B. Lateral root organogenesis—From cell to organ. Curr. Opin. Plant Biol. 2010, 13, 677–683. [Google Scholar]
- Mohammad, S.; Rammyani, B.; Mark, E. SCFTIR1/AFB-based auxin perception: Mechanism and role in plant growth and development. Plant Cell 2015, 27, 9–19. [Google Scholar]
- Wang, J.W.; Wang, L.J.; Mao, Y.B.; Cai, W.J.; Xue, H.W.; Chen, X.Y. Control of root cap formation by microRNA-targeted auxin response factors in Arabidopsis. Plant Cell 2005, 17, 2204–2216. [Google Scholar] [CrossRef]
- Wang, S.; Bai, Y.; Shen, C.; Wu, Y.; Zhang, S.; Jiang, D.; Guilfoyle, T.J.; Chen, M.; Qi, Y. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genom. 2010, 10, 533–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Nie, L.; Ma, J.; Zhou, B.; Han, X.; Cheng, J.; Lu, X.; Fan, Z.; Li, Y.; Cao, Y. Transcriptomic variations and network hubs controlling seed size and weight during maize seed development. Front. Plant Sci. 2022, 13, 828923. [Google Scholar] [CrossRef]
- Qiao, L.; Zhang, W.; Li, X.; Zhang, L.; Zhang, X.; Li, X.; Guo, H.; Ren, Y.; Zheng, J.; Chang, Z. Characterization and expression patterns of auxin response factors in wheat. Front. Plant Sci. 2018, 9, 1395. [Google Scholar] [CrossRef]
- Tiwari, S.B.; Hagen, G.; Guilfoyle, T.J. The roles of auxin response factor domains in auxin-responsive transcription. Plant Cell 2003, 15, 533–543. [Google Scholar] [CrossRef]
- Guilfoyle, T.J.; Hagen, G. Auxin response factors: Recent advances in auxin biology. J. Plant Growth Regul. 2001, 10, 281–291. [Google Scholar] [CrossRef]
- Krogan, N.T.; Marcos, D.; Weiner, A.I.; Berleth, T. The auxin response factor MONOPTEROS controls meristem function and organogenesis in both the shoot and root through the direct regulation of PIN genes. New Phytol. 2016, 212, 42–50. [Google Scholar] [CrossRef]
- Hardtke, C.S. Root development—Branching into novel spheres. Curr. Opin. Plant Biol. 2006, 9, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Schlereth, A.; Möller, B.; Liu, W.; Kientz, M.; Flipse, J.; Rademacher, E.H.; Schmid, M.; Jürgens, G.; Weijers, D. MONOPTEROS controls embryonic root initiation by regulating a mobile transcription factor. Nature 2010, 464, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Tian, C.E.; Muto, H.; Higuchi, K.; Matamura, T.; Tatematsu, K.; Koshiba, T.; Yamamoto, K.T. Disruption and overexpression of auxin response factor 8 gene of Arabidopsis affect hypocotyl elongation and root growth habit, indicating its possible involvement in auxin homeostasis in light condition. Plant J. 2004, 40, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Wilmoth, J.C.; Wang, S.; Tiwari, S.B.; Joshi, A.D.; Hagen, G.; Guilfoyle, T.J.; Alonso, J.M.; Ecker, J.R.; Reed, J.W. NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. Plant J. 2005, 43, 118–130. [Google Scholar] [CrossRef]
- Lee, H.W.; Kim, N.Y.; Lee, D.J.; Kim, J. LBD18/ASL20 regulates lateral root formation in combination with LBD16/ASL18 downstream of ARF7 and ARF19 in Arabidopsis. Plant Physiol. 2009, 151, 1377–1389. [Google Scholar] [CrossRef]
- Yong, H.L.; Sha, Q.H.; Yan, H.Q. Advances in structure and function of auxin response factor in plants. J. Integr. Plant Biol. 2023, 16, 617–632. [Google Scholar]
- Li, W.C.; Kai, Y.; Chun, H.C.; Bo, Z.; Ya, N.H.; Wan, Q.W.; Pu, Z.; Kai, L.W.; Bin, H.W.; Ying, X.; et al. Chromosome-level reference genome and resequencing of 322 accessions reveal evolution, genomic imprint and key agronomic traits in adzuki bean. Plant Biotechnol. J. 2024, 22, 2173–2185. [Google Scholar]
- Shahid, A.; Wan, P.W.; Zhao, H.Z.; Li, N.X.; Roeland, B.; Naeem, K. Identification and confirmation of novel genetic loci and domestication gene GmGA20ox1 regulating primary root length in soybean seedling stage. Ind. Crops Prod. 2024, 30, 118814. [Google Scholar]
- Shobha, S.; Stacy, C.; Elena, S.; Dakshesh, D.; Larry, C.; Ilene, K.M.; Conrad, L.S. The NCBI BioCollections Database. Database 2018, 1, bay006. [Google Scholar]
- Samantha, K.P.; Alex, S.H.; David, A.K.; Katherine, H.S.; Natalie, M.C.; Hong, W.J.; Ryan, E.; Soeun, H.; Eric, T.; Ildoo, H.; et al. Nucleo-cytoplasmic Partitioning of ARF Proteins Controls Auxin Responses in Arabidopsis thaliana. Mol. Cell 2019, 76, 177–190. [Google Scholar]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Grey, M.; Thanvi, S.; Pablo, C.B.; Claude, B.; Mariele, L.; Moises, E.A.; Marie, K.; Julia, H.; Manuela, N.; Daniel, K.; et al. Mutation bias reflects natural selection in Arabidopsis thaliana. Nature 2022, 602, 101–105. [Google Scholar]
- Yoko, O.; Hidehiro, F.; Makoto, O.; Athanasios, T.; Masao, T. ARF7 and ARF19 regulate lateral root formation via direct activation of LBD/ASL genes in Arabidopsis. Plant Cell 2007, 19, 118–130. [Google Scholar]
- Meng, Q.C.; Chen, X.; Tao, W.; Li, H.Z.; Yu, X.W.; Gui, X.L.; Jing, Y.Y.; Ji, M.X.; Rong, L.; Zhi, Y.W.; et al. An LRH-RSL4 feedback regulatory loop controls the determinate growth of root hairs in Arabidopsis. Curr. Biol. 2024, 22, 313–326. [Google Scholar]
- Philippe, L.; Tanya, Z.B.; Donghui, L.; David, S.; Christopher, W.; Rajkumar, S.; Robert, M.; Kate, D.; Debbie, L.A.; Margarita, G.H.; et al. The Arabidopsis Information Resource (TAIR): Improved gene annotation and new tools. Nucleic Acids Res. 2012, 40, 1202–1210. [Google Scholar]
- David, M.G.; Sheng, Q.S.; Russell, H.; Rochak, N.; Richard, D.H.; Joni, F.; Therese, M.; William, D.; Uffe, H.; Nicholas, P.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, 1178–1186. [Google Scholar]
- Koichiro, T.; Daniel, P.; Nicholas, P.; Glen, S.; Masatoshi, N.; Sudhir, K. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum Parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar]
- Ivica, L.; Peer, B. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, 293–296. [Google Scholar]
- Martin, K.; Jacqueline, S.; Inanç, B.; Joseph, C.; Randy, G.; Doug, H.; Steven, J.J.; Marco, A.M. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar]
- Yu, Q.W.; Hai, B.T.; Jeremy, D.D.B.; Xu, T.; Jing, Q.L.; Xi, Y.W.; Tae-ho, L.; Hui, Z.J.; Barry, M.; Hui, G.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, 49. [Google Scholar]
- Cheng, J.C.; Hao, C.; Yi, Z.; Hannah, R.; Thomas, H.R.; Margaret, H.F.; Ye, H.H.; Rui, X. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 8, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.Z.; Ying, H.L.; Yan, F.L.; Sheng, R.Z.; Tian, L.G.; Chun, C.W.; Fan, Z.; Muhiuddin, F.; Lina, Z.; Xiang, Y.W.; et al. A general model for “germplasm-omics” data sharing and mining: A case study of SoyFGB v2.0. Sci. Bull. 2022, 15, 1716–1719. [Google Scholar]
- Robert, D.F.; Teresa, K.A.; Patricia, C.B.; Alex, B.; Peer, B.; Alan, J.B.; Hsin, Y.C.; Zsuzsanna, D.; Sara, E.G.; Matthew, F.; et al. InterPro in 2017-beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, 190–199. [Google Scholar]
- Scott, F. The NCBI Taxonomy database. Nucleic Acids Res. 2012, 8, 136–143. [Google Scholar]
- Paul, H.; Keun-Joon, P.; Takeshi, O.; Naoya, F.; Hajime, H.; Adams-Collier, C.J.; Kenta, N. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, 585–587. [Google Scholar]
- Andrew, W.; Martino, B.; Stefan, B.; Gabriel, S.; Gerardo, T.; Rafal, G.; Florian, T.H.; Tjaart, A.P.B.; Christine, R.; Lorenza, B.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, 296–303. [Google Scholar]
- Timothy, L.B.; James, J.; Charles, E.G.; William, S.N. The MEME Suite. Nucleic Acids Res. 2015, 43, 39–49. [Google Scholar]
- Magali, L.; Patrice, D.; Gert, T.; Kathleen, M.; Yves, M.; Yves, V.P.; Pierre, R.; Stephane, R. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar]
- Fabricio, A.; Francisnei, P.S.; Thiago, M.V. The Soybean Expression Atlas v2: A comprehensive database of over 5000 RNA-seq samples. Plant J. 2023, 116, 1041–1051. [Google Scholar]
- Xiao, Z.; Jia, X.C.; Wen, T.L.; Hui, W.Z.; Ying, H.; Xin, X.L.; Hong, L. Molecular module GmPTF1a/b-GmNPLa regulates rhizobia infection and nodule formation in soybean. New Phytol. 2024, 241, 1813–1828. [Google Scholar]
- Han, Y.X.; Yan, J.L.; Ke, F.Z.; Ming, J.L.; Si, Y.F.; Ying, Z.T.; Tong, F.Q.; Xin, X.L.; Yong, J.Z.; Hong, L. miR169c-NFYA-C-ENOD40 modulates nitrogen inhibitory effects in soybean nodulation. New Phytol. 2021, 229, 3377–3392. [Google Scholar]
- Varaud, E.; Brioudes, F.; Szécsi, J.; Leroux, J.; Brown, S.; Perrot Rechenmann, C.; Bendahmane, M. AUXIN RESPONSE FACTOR 8 regulates Arabidopsis petal growth by interacting with the bHLH transcription factor BIGPETALp. Plant Cell 2011, 23, 973–983. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | Gene Name | Gene Loci | Chromosome | Gene Position | Size (aa) | MW (Da) | PI | Instability Index | A.I. | GRAVY | Predicted Location | Arabidopsis Orthologues | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Start | End | Gene Name | Gene Loci | |||||||||||
| Suppressor | GmARF1 | Glyma.07G272800 | chr07 | 44,814,499 | 44,821,841 | 701 | 78,043.97 | 5.83 | 58.93 | 75.48 | −0.475 | Nucleus | AtARF1 | AtlG59750 |
| Glyma.12G164100 | chr12 | 33,328,006 | 33,334,622 | 665 | 74,273.52 | 5.75 | 60.82 | 67.98 | −0.523 | Nucleus | ||||
| Glyma.16G000300 | chr16 | 25,166 | 31,978 | 666 | 74,250.40 | 5.62 | 58.72 | 70.36 | −0.494 | Nucleus | ||||
| GmARF2 | Glyma.05G200800 | chr05 | 38,470,758 | 38,476,414 | 858 | 94,845.13 | 6.35 | 52.13 | 67.02 | −0.638 | Nucleus | AtARF2 | At5G62000 | |
| Glyma.08G008100 | chr08 | 643,657 | 649,224 | 853 | 95,522.61 | 6.50 | 53.98 | 71.98 | −0.577 | Nucleus | ||||
| Glyma.06G164900 | chr06 | 13,626,690 | 13,632,154 | 843 | 93,667.20 | 6.19 | 59.40 | 63.49 | −0.654 | Nucleus | ||||
| Glyma.04G200600 | chr04 | 46,137,859 | 46,143,355 | 850 | 94,721.43 | 6.35 | 56.01 | 64.93 | −0.661 | Nucleus | ||||
| Glyma.19G206100 | chr19 | 46,694,888 | 46,706,151 | 677 | 75,645.75 | 7.11 | 57.20 | 73.87 | −0.489 | Nucleus | ||||
| GmARF3 | Glyma.13G174000 | chr13 | 28,226,449 | 28,232,503 | 714 | 78,286.47 | 6.67 | 54.87 | 70.31 | −0.400 | Nucleus | AtARF3 | At2G33860 | |
| Glyma.07G202200 | chr07 | 37,447,174 | 37,452,876 | 709 | 77,934.52 | 6.67 | 52.83 | 73.27 | −0.358 | Nucleus | ||||
| Glyma.13G234200 | chr13 | 33,897,474 | 33,903,871 | 739 | 80,750.89 | 6.18 | 55.82 | 75.20 | −0.315 | Nucleus | ||||
| Glyma.15G078800 | chr15 | 6,035,900 | 6,041,756 | 736 | 80,922.02 | 6.10 | 54.51 | 75.50 | −0.309 | Plasmodesmata | ||||
| GmARF4 | Glyma.13G327951 | chr13 | 41,634,392 | 41,640,306 | 254 | 28,301.65 | 5.23 | 54.88 | 66.81 | −0.498 | Nucleus | AtARF4 | At5G60450 | |
| Glyma.12G171000 | chr12 | 34,108,206 | 34,114,864 | 799 | 88,732.31 | 6.49 | 55.71 | 70.53 | −0.438 | Nucleus | ||||
| Glyma.11G154632 | chr11 | 11,580,788 | 11,587,787 | 792 | 88,149.47 | 5.98 | 53.35 | 74.91 | −0.433 | Nucleus | ||||
| Glyma.12G071000 | chr12 | 5,182,536 | 5,190,631 | 792 | 87,731.72 | 6.00 | 53.18 | 73.83 | −0.428 | Nucleus | ||||
| GmARF9 | Glyma.18G184500 | chr18 | 44,742,372 | 44,749,324 | 664 | 74,553.01 | 5.89 | 51.79 | 76.46 | −0.472 | Nucleus | AtARF9 | At4G23980 | |
| Glyma.07G134800 | chr07 | 15,844,232 | 15,849,865 | 664 | 74,516.94 | 5.99 | 51.99 | 74.23 | −0.501 | Nucleus | ||||
| Glyma.03G070500 | chr03 | 16,448,588 | 16,454,400 | 691 | 76,860.43 | 6.02 | 46.87 | 68.67 | −0.537 | Nucleus | ||||
| Glyma.01G103500 | chr01 | 35,187,770 | 35,192,407 | 692 | 76961.62 | 6.02 | 46.67 | 70.36 | −0.504 | Nucleus | ||||
| Glyma.16G023600 | chr16 | 2,242,818 | 2,248,084 | 717 | 79,581.75 | 6.14 | 53.34 | 72.73 | −0.474 | Nucleus | ||||
| GmARF17 | Glyma.14G166500 | chr14 | 42,041,638 | 42,046,150 | 548 | 60,511.91 | 6.36 | 53.03 | 63.70 | −0.389 | Chloroplast | AtARF17 | At1G77850 | |
| Glyma.13G084700 | chr13 | 18,614,254 | 18,618,647 | 551 | 60,677.92 | 5.78 | 52.46 | 63.36 | −0.386 | Chloroplast | ||||
| Glyma.06G108051 | chr06 | 8,647,276 | 8,649,645 | 162 | 18,067.40 | 6.85 | 51.29 | 71.48 | −0.478 | Cytoplasm | ||||
| Glyma.04G254200 | chr04 | 50,908,674 | 50,913,759 | 562 | 61,209.95 | 5.53 | 48.05 | 71.46 | −0.265 | Chloroplast | ||||
| GmARF18 | Glyma.19G181900 | chr19 | 44,488,211 | 44,492,387 | 700 | 77,788.51 | 8.30 | 50.38 | 71.20 | −0.378 | Nucleus | AtARF18 | At3G61830 | |
| Glyma.13G140600 | chr13 | 24,398,045 | 24,400,077 | 514 | 57,052.25 | 8.93 | 45.45 | 74.94 | −0.344 | Nucleus | ||||
| Glyma.10G053500 | chr10 | 4,804,493 | 4,809,528 | 700 | 77,266.44 | 7.26 | 46.42 | 73.39 | −0.389 | Nucleus | ||||
| Glyma.11G145500 | chr11 | 17,400,515 | 17,404,144 | 697 | 76,555.96 | 7.85 | 43.90 | 74.42 | −0.324 | Nucleus | ||||
| Glyma.12G076200 | chr12 | 5,836,558 | 5,840,461 | 701 | 77,188.67 | 8.24 | 43.03 | 73.59 | −0.348 | Nucleus | ||||
| Glyma.13G325200 | chr13 | 41,405,939 | 41,408,913 | 670 | 73,760.45 | 7.61 | 48.92 | 75.99 | −0.331 | Nucleus | ||||
| Glyma.12G174100 | chr12 | 34,605,359 | 34,608,860 | 700 | 76,824.93 | 7.60 | 48.61 | 75.07 | −0.361 | Nucleus | ||||
| Glyma.10G210600 | chr10 | 44,358,942 | 44,362,451 | 612 | 67,127.69 | 7.21 | 45.48 | 67.70 | −0.397 | Nucleus | ||||
| Glyma.20G180000 | chr20 | 41,744,451 | 41,747,715 | 593 | 65,246.96 | 6.70 | 46.04 | 70.20 | −0.336 | Nucleus | ||||
| Glyma.03G258300 | chr03 | 46,452,008 | 46,456,771 | 669 | 74,921.72 | 6.65 | 59.84 | 69.46 | −0.516 | Nucleus | ||||
| Glyma.07G054800 | chr07 | 4,782,625 | 4,787,849 | 716 | 79,530.35 | 6.07 | 52.65 | 70.80 | −0.528 | Nucleus | ||||
| GmARF23 | Glyma.03G208800 | chr03 | 42,723,037 | 42,743,487 | 786 | 88,027.81 | 8.38 | 51.78 | 86.07 | −0.235 | Plasmodesmata | AtARF23 | AtlG43950 | |
| Activator | GmARF6 | Glyma.14G032700 | chr14 | 2,368,563 | 2,377,330 | 898 | 99,065.45 | 6.13 | 66.79 | 75.04 | −0.414 | Nucleus | AtARF6 | At1G30330 |
| Glyma.02G281700 | chr02 | 48,225,437 | 48,235,389 | 896 | 98,952.51 | 6.04 | 67.86 | 75.31 | −0.410 | Nucleus | ||||
| Glyma.13G221400 | chr13 | 32,864,280 | 32,871,326 | 896 | 99,837.22 | 6.28 | 60.45 | 71.91 | −0.497 | Nucleus | ||||
| Glyma.15G091000 | chr15 | 6,970,378 | 6,977,273 | 898 | 99,689.89 | 6.22 | 57.02 | 73.82 | −0.476 | Nucleus | ||||
| Glyma.05G143800 | chr05 | 33,795,530 | 33,802,884 | 909 | 100,674.79 | 6.13 | 66.68 | 70.87 | −0.483 | Nucleus | ||||
| Glyma.08G100100 | chr08 | 7,676,663 | 7,683,563 | 907 | 100,662.05 | 6.17 | 64.72 | 72.21 | −0.483 | Nucleus | ||||
| GmARF8 | Glyma.18G119700 | chr18 | 14,818,239 | 14,819,381 | 119 | 13,215.88 | 4.99 | 59.26 | 82.69 | −0.364 | Cytoplasm | AtARF8 | At5G37020 | |
| Glyma.18G046800 | chr18 | 4,068,582 | 4,076,135 | 841 | 93,247.82 | 5.93 | 57.98 | 76.05 | −0.441 | Nucleus | ||||
| Glyma.11G204200 | chr11 | 3,3637,709 | 33,645,524 | 844 | 93,600.09 | 5.93 | 58.73 | 75.89 | −0.461 | Nucleus | ||||
| Glyma.14G208500 | chr14 | 48,263,889 | 48,272,154 | 843 | 93,795.44 | 6.01 | 60.63 | 74.12 | −0.473 | Nucleus | ||||
| Glyma.02G239600 | chr02 | 44,642,958 | 44,654,516 | 847 | 94,202.81 | 6.06 | 61.67 | 73.55 | −0.495 | Nucleus | ||||
| GmARF19 | Glyma.07G130400 | chr07 | 15,309,926 | 15,315,946 | 1110 | 122,978.03 | 6.30 | 63.79 | 75.15 | −0.578 | Nucleus | AtARF19 | At1G19220 | |
| Glyma.01G002100 | chr01 | 294,373 | 300,422 | 1104 | 121,957.83 | 6.51 | 63.38 | 76.10 | −0.561 | Nucleus | ||||
| Glyma.05G221300 | chr05 | 40,123,000 | 40,129,871 | 1099 | 120,828.59 | 6.01 | 61.52 | 80.35 | −0.455 | Nucleus | ||||
| Glyma.08G027800 | chr08 | 2,218,615 | 2,225,900 | 1113 | 122,551.22 | 5.86 | 63.04 | 77.85 | −0.503 | Nucleus | ||||
| Glyma.09G072200 | chr09 | 7,436,392 | 7,445,095 | 1125 | 125,840.87 | 6.02 | 67.64 | 67.01 | −0.698 | Nucleus | ||||
| Glyma.15G181000 | chr15 | 17,521,997 | 17,530,209 | 1122 | 125,284.27 | 6.19 | 67.09 | 67.9 | −0.686 | Nucleus | ||||
| Glyma.13G112600 | chr13 | 21,613,425 | 21,621,047 | 1131 | 126,747.12 | 6.08 | 65.28 | 73.47 | −0.673 | Nucleus | ||||
| Glyma.17G047100 | chr17 | 3,563,891 | 3,571,811 | 1136 | 127,058.28 | 6.05 | 67.23 | 72.90 | −0.683 | Nucleus | ||||
| GmARF5 | Glyma.12G153700 | chr12 | 23,224,445 | 23,225,261 | 109 | 120,50.98 | 7.86 | 46.01 | 83.21 | −0.184 | Cytoplasm | AtARF5 | At1G19850 | |
| Glyma.14G217700 | chr14 | 49,093,824 | 49,100,973 | 930 | 103,499.15 | 5.62 | 53.23 | 78.17 | −0.407 | Nucleus | ||||
| Glyma.17G256500 | chr17 | 41,134,622 | 41,140,694 | 933 | 103,492.16 | 5.48 | 51.50 | 77.19 | −0.384 | Nucleus | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Miao, T.; Liao, H.; Zhong, Y. Genome-Wide Identification of GmARF9b/GmARF2a Negatively Regulate Root Growth in Soybean. Int. J. Mol. Sci. 2025, 26, 4547. https://doi.org/10.3390/ijms26104547
Li D, Miao T, Liao H, Zhong Y. Genome-Wide Identification of GmARF9b/GmARF2a Negatively Regulate Root Growth in Soybean. International Journal of Molecular Sciences. 2025; 26(10):4547. https://doi.org/10.3390/ijms26104547
Chicago/Turabian StyleLi, Di, Tianle Miao, Hong Liao, and Yongjia Zhong. 2025. "Genome-Wide Identification of GmARF9b/GmARF2a Negatively Regulate Root Growth in Soybean" International Journal of Molecular Sciences 26, no. 10: 4547. https://doi.org/10.3390/ijms26104547
APA StyleLi, D., Miao, T., Liao, H., & Zhong, Y. (2025). Genome-Wide Identification of GmARF9b/GmARF2a Negatively Regulate Root Growth in Soybean. International Journal of Molecular Sciences, 26(10), 4547. https://doi.org/10.3390/ijms26104547