
Light-Dependent Regulatory Interactions between the Redox System and miRNAs and Their Biochemical and Physiological Effects in Plants



Abstract
1. Introduction
2. Modulation of the Redox System by Light
2.1. Effect of Light Intensity on the Redox System
2.2. Regulation of the Redox System by Light Spectrum
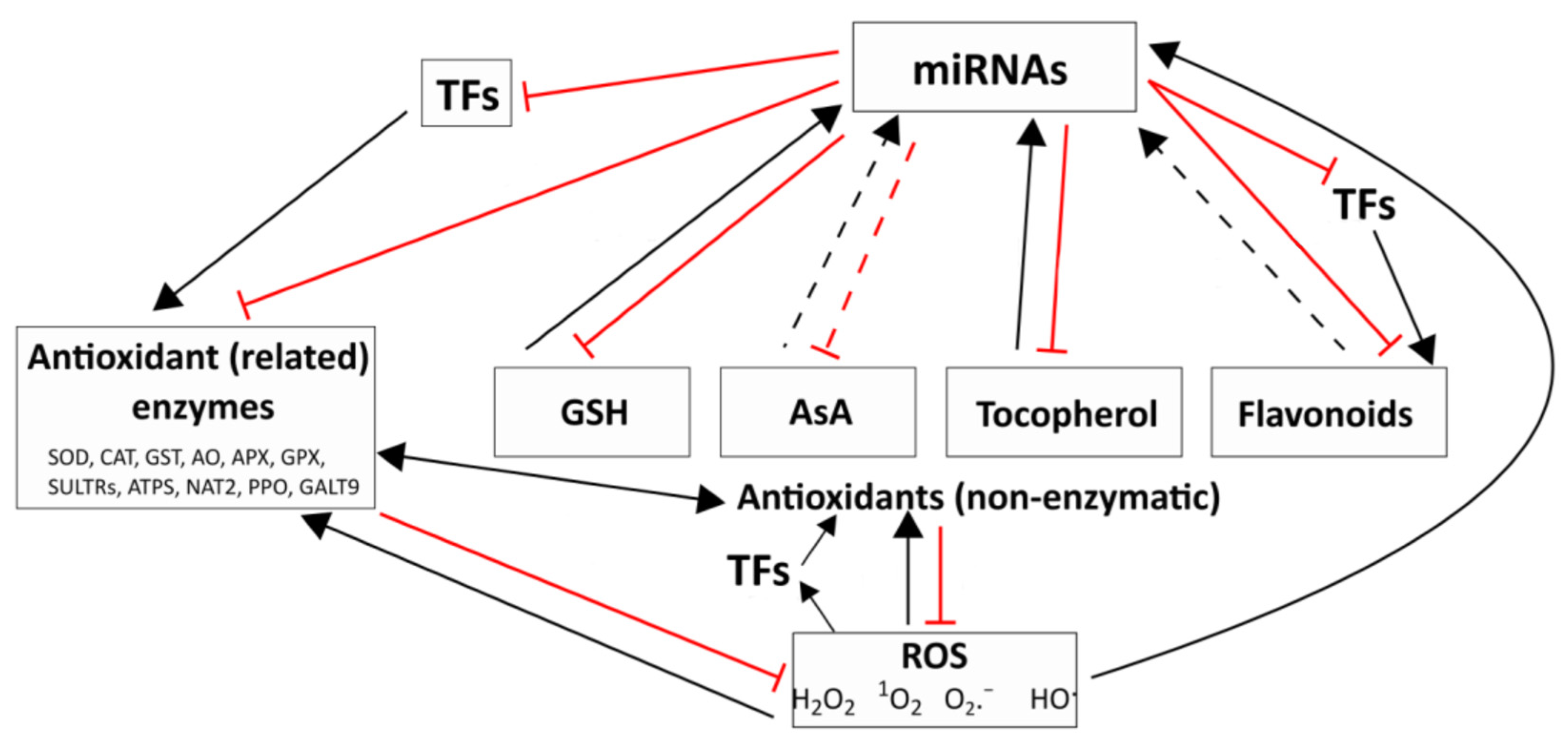
3. Regulatory Relationships between the Redox System and the miRNAs
3.1. Interactions between ROS, ROS-Processing Enzymes and miRNAs
3.2. Interplay between the Main Non-Enzymatic and Enzymatic Antioxidants and miRNAs
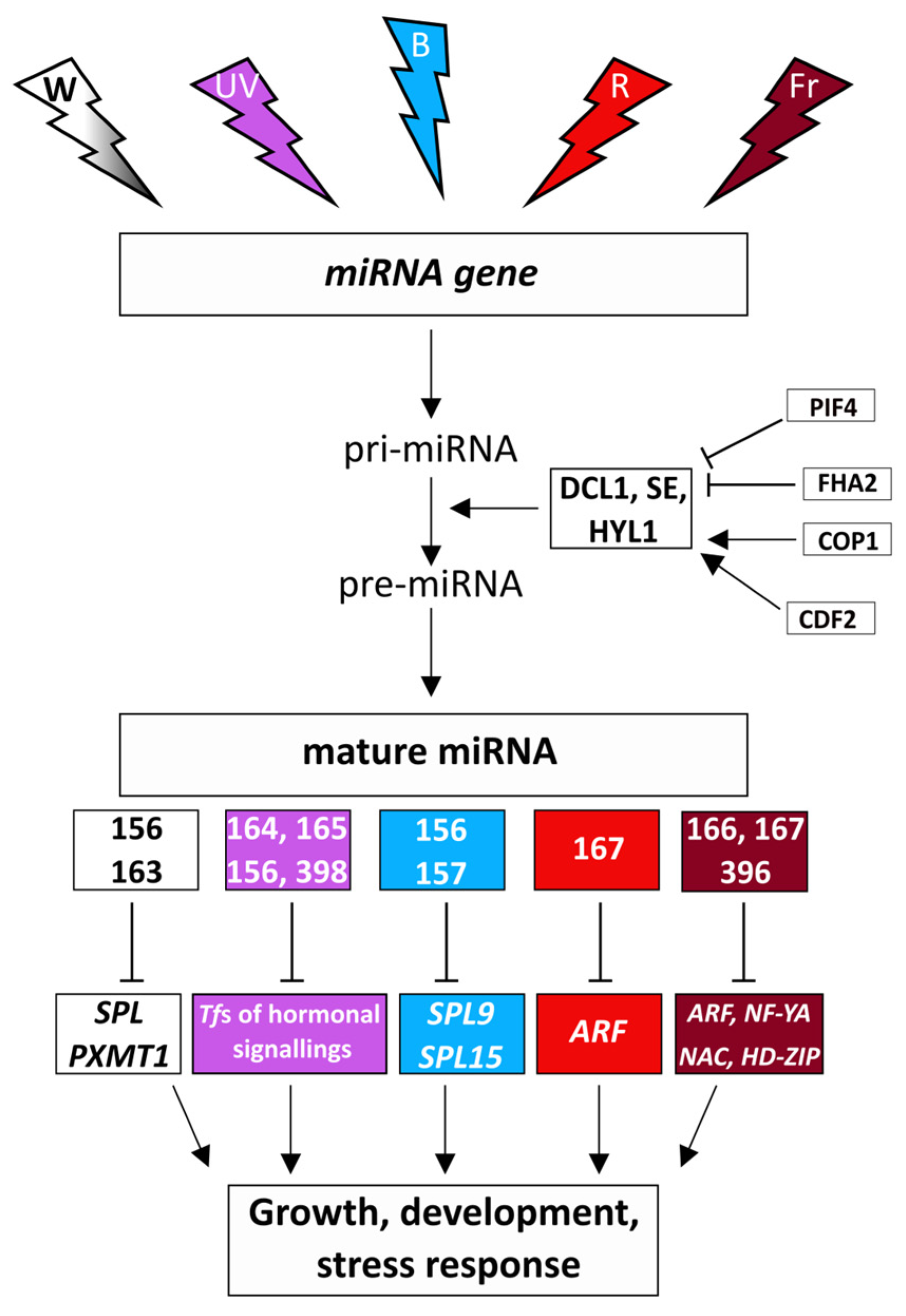
4. Control of miRNAs by Light
4.1. Regulatory Relationships between the Light Intensity and the miRNAs
4.2. Spectral Control of miRNAs
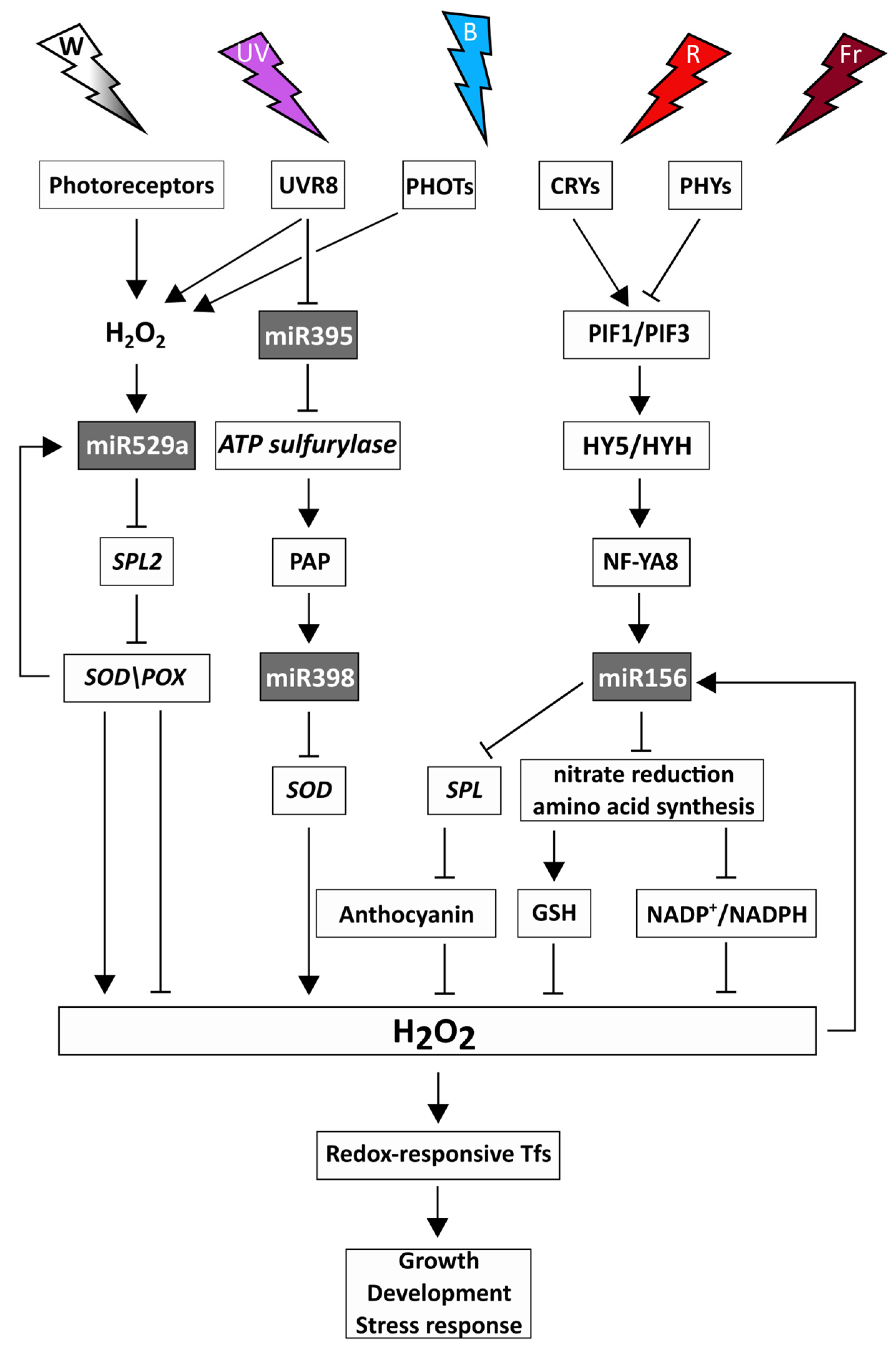
5. Light-Dependent Interaction between the Redox System and miRNAs
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Usuda, H. Evaluation of the Effect of Photosynthesis on Biomass Production with Simultaneous Analysis of Growth and Continuous Monitoring of CO2 Exchange in the Whole Plants of Radish, Cv Kosena under Ambient and Elevated CO2. Plant Prod. Sci. 2004, 7, 386–396. [Google Scholar] [CrossRef]
- Griffin, J.H.C.; Toledo-Ortiz, G.; Teige, M. Plant Photoreceptors and Their Signalling Components in Chloroplastic Anterograde and Retrograde Communication. J. Exp. Bot. 2022, 73, 7126–7138. [Google Scholar] [CrossRef]
- Kong, S.-G.; Okajima, K. Diverse Photoreceptors and Light Responses in Plants. J. Plant Res. 2016, 129, 111–114. [Google Scholar] [CrossRef]
- Su, J.; Liu, B.; Liao, J.; Yang, Z.; Lin, C.; Oka, Y. Coordination of Cryptochrome and Phytochrome Signals in the Regulation of Plant Light Responses. Agronomy 2017, 7, 25. [Google Scholar] [CrossRef]
- Roeber, V.M.; Bajaj, I.; Rohde, M.; Schmülling, T.; Cortleven, A. Light Acts as a Stressor and Influences Abiotic and Biotic Stress Responses in Plants. Plant. Cell Environ. 2021, 44, 645–664. [Google Scholar] [CrossRef] [PubMed]
- Borbély, P.; Gasperl, A.; Pálmai, T.; Ahres, M.; Asghar, M.A.; Galiba, G.; Müller, M.; Kocsy, G. Light Intensity- and Spectrum-Dependent Redox Regulation of Plant Metabolism. Antioxidants 2022, 11, 1311. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Managing the Cellular Redox Hub in Photosynthetic Organisms. Plant. Cell Environ. 2012, 35, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Kocsy, G.; Tari, I.; Vanková, R.; Zechmann, B.; Gulyás, Z.; Poór, P.; Galiba, G. Redox Control of Plant Growth and Development. Plant Sci. 2013, 211, 77–91. [Google Scholar] [CrossRef]
- Considine, M.J.; Foyer, C.H. Oxygen and Reactive Oxygen Species-Dependent Regulation of Plant Growth and Development. Plant Physiol. 2021, 186, 79–92. [Google Scholar] [CrossRef]
- Corpas, F.J.; Gupta, D.K.; Palma, J.M. Production Sites of Reactive Oxygen Species (ROS) in Organelles from Plant Cells. In Reactive Oxygen Species and Oxidative Damage in Plants Under Stress; Gupta, D.K., Corpas, F.J., Palma, J.M., Eds.; Springer International Publishing: New York, NY, USA, 2015; pp. 1–22. [Google Scholar] [CrossRef]
- Viola, I.L.; Camoirano, A.; Gonzalez, D.H. Redox-Dependent Modulation of Anthocyanin Biosynthesis by the TCP Transcription Factor TCP15 during Exposure to High Light Intensity Conditions in Arabidopsis. Plant Physiol. 2015, 170, 74–85. [Google Scholar] [CrossRef]
- Qiu, Z.; Zhu, L.; He, L.; Chen, D.; Zeng, D.; Chen, G.; Hu, J.; Zhang, G.; Ren, D.; Dong, G.; et al. DNA Damage and Reactive Oxygen Species Cause Cell Death in the Rice Local Lesions 1 Mutant under High Light and High Temperature. New Phytol. 2019, 222, 349–365. [Google Scholar] [CrossRef] [PubMed]
- Saed-Moucheshi, A.; Pakniyat, H.; Pirasteh-Anosheh, H.; Azooz, M.M. Role of ROS as Signaling Molecules in Plants. In Oxidative Damage to Plants; Ahmad, P., Ed.; Academic Press: San Diego, CA, USA, 2014; pp. 585–620. [Google Scholar] [CrossRef]
- Foyer, C.H. Reactive Oxygen Species, Oxidative Signaling and the Regulation of Photosynthesis. Environ. Exp. Bot. 2018, 154, 134–142. [Google Scholar] [CrossRef]
- Gallé, Á.; Czékus, Z.; Bela, K.; Horváth, E.; Ördög, A.; Csiszár, J.; Poór, P. Plant Glutathione Transferases and Light. Front. Plant Sci. 2019, 9, 1944. [Google Scholar] [CrossRef]
- Cimini, S.; Gualtieri, C.; Macovei, A.; Balestrazzi, A.; De Gara, L.; Locato, V. Redox Balance-DDR-MiRNA Triangle: Relevance in Genome Stability and Stress Responses in Plants. Front. Plant Sci. 2019, 10, 989. [Google Scholar] [CrossRef]
- Sanei, M.; Chen, X. Mechanisms of MicroRNA Turnover. Curr. Opin. Plant Biol. 2015, 27, 199–206. [Google Scholar] [CrossRef]
- Wang, S.; Wu, W.; Claret, F.X. Mutual Regulation of MicroRNAs and DNA Methylation in Human Cancers. Epigenetics 2017, 12, 187–197. [Google Scholar] [CrossRef]
- Borges, F.; Martienssen, R.A. The Expanding World of Small RNAs in Plants. Nat. Rev. Mol. Cell Biol. 2015, 16, 727–741. [Google Scholar] [CrossRef]
- Bajczyk, M.; Jarmolowski, A.; Jozwiak, M.; Pacak, A.; Pietrykowska, H.; Sierocka, I.; Swida-Barteczka, A.; Szewc, L.; Szweykowska-Kulinska, Z. Recent Insights into Plant MiRNA Biogenesis: Multiple Layers of MiRNA Level Regulation. Plants 2023, 12, 342. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, M.B.; Tentori, A.M.; Zhang, W.C.; Slack, F.J.; Doyle, P.S. Spatially Resolved and Multiplexed MicroRNA Quantification from Tissue Using Nanoliter Well Arrays. Microsystems Nanoeng. 2020, 6, 51. [Google Scholar] [CrossRef]
- Hossain, M.S.; Hoang, N.T.; Yan, Z.; Tóth, K.; Meyers, B.C.; Stacey, G. Characterization of the Spatial and Temporal Expression of Two Soybean Mirnas Identifies SCL6 as a Novel Regulator of Soybean Nodulation. Front. Plant Sci. 2019, 10, 475. [Google Scholar] [CrossRef] [PubMed]
- Loreti, E.; Perata, P. Mobile Plant MicroRNAs Allow Communication within and between Organisms. New Phytol. 2022, 235, 2176–2182. [Google Scholar] [CrossRef]
- Pyott, D.E.; Molnar, A. Going Mobile: Non-Cell-Autonomous Small RNAs Shape the Genetic Landscape of Plants. Plant Biotechnol. J. 2015, 13, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Liu, X.; Li, D.; Wang, P.; Li, N.; Lu, L.; Cao, X. MicroRNA-466l Upregulates IL-10 Expression in TLR-Triggered Macrophages by Antagonizing RNA-Binding Protein Tristetraprolin-Mediated IL-10 MRNA Degradation. J. Immunol. 2010, 184, 6053–6059. [Google Scholar] [CrossRef]
- Lopez-Gomollon, S.; Mohorianu, I.; Szittya, G.; Moulton, V.; Dalmay, T. Diverse Correlation Patterns between MicroRNAs and Their Targets during Tomato Fruit Development Indicates Different Modes of MicroRNA Actions. Planta 2012, 236, 1875–1887. [Google Scholar] [CrossRef]
- Juarez, M.T.; Kui, J.S.; Thomas, J.; Heller, B.A.; Timmermans, M.C.P. MicroRNA-Mediated Repression of Rolled Leaf1 Specifies Maize Leaf Polarity. Nature 2004, 428, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.Y.; Zhang, M.; Tao, Y.; Xu, H.; Liu, Q.; Yang, K.; Wei, R.; Zhou, H.; Jin, T.; Liu, X.D.; et al. MiRNA-22 Upregulates Mtf1 in Dorsal Horn Neurons and Is Essential for Inflammatory Pain. Oxid. Med. Cell. Longev. 2022, 2022, 1–23. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T.L. A Meta-Analysis of Plant Responses to Light Intensity for 70 Traits Ranging from Molecules to Whole Plant Performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef]
- Kumar, A.; Guleria, S.; Ghosh, D.; Dogra, V.; Kumar, S. Managing Reactive Oxygen Species—Some Learnings from High Altitude Extremophytes. Environ. Exp. Bot. 2021, 189, 104525. [Google Scholar] [CrossRef]
- Ruban, A.V.; Belgio, E. The Relationship between Maximum Tolerated Light Intensity and Photoprotective Energy Dissipation in the Photosynthetic Antenna: Chloroplast Gains and Losses. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 1640. [Google Scholar] [CrossRef] [PubMed]
- Sakuraba, Y. Light-Mediated Regulation of Leaf Senescence. Int. J. Mol. Sci. 2021, 22, 3291. [Google Scholar] [CrossRef] [PubMed]
- D’Alessandro, S.; Beaugelin, I.; Havaux, M. Tanned or Sunburned: How Excessive Light Triggers Plant Cell Death. Mol. Plant 2020, 13, 1545–1555. [Google Scholar] [CrossRef]
- Shumbe, L.; Chevalier, A.; Legeret, B.; Taconnat, L.; Monnet, F.; Havaux, M. Singlet Oxygen-Induced Cell Death in Arabidopsis under High-Light Stress Is Controlled by OXI1 Kinase. Plant Physiol. 2016, 170, 1757–1771. [Google Scholar] [CrossRef]
- Xiong, H.; Hua, L.; Reyna-Llorens, I.; Shi, Y.; Chen, K.M.; Smirnoff, N.; Kromdijk, J.; Hibberd, J.M. Photosynthesis-Independent Production of Reactive Oxygen Species in the Rice Bundle Sheath during High Light Is Mediated by NADPH Oxidase. Proc. Natl. Acad. Sci. USA 2021, 118, e2022702118. [Google Scholar] [CrossRef] [PubMed]
- Fichman, Y.; Xiong, H.; Sengupta, S.; Morrow, J.; Loog, H.; Azad, R.K.; Hibberd, J.M.; Liscum, E.; Mittler, R. Phytochrome B Regulates Reactive Oxygen Signaling during Abiotic and Biotic Stress in Plants. New Phytol. 2023, 237, 1711–1727. [Google Scholar] [CrossRef]
- Aswani, V.; Rajsheel, P.; Bapatla, R.B.; Sunil, B.; Raghavendra, A.S. Oxidative Stress Induced in Chloroplasts or Mitochondria Promotes Proline Accumulation in Leaves of Pea (Pisum sativum): Another Example of Chloroplast-Mitochondria Interactions. Protoplasma 2019, 256, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.H.; Kim, J.H.; Kim, S.G.; Sim, H.J.; Lee, G.; Halitschke, R.; Baldwin, I.T.; Kim, J.L.; Park, C.M. Shoot Phytochrome B Modulates Reactive Oxygen Species Homeostasis in Roots via Abscisic Acid Signaling in Arabidopsis. Plant J. 2018, 94, 790–798. [Google Scholar] [CrossRef]
- Karpinski, S.; Reynolds, H.; Karpinska, B.; Wingsle, G.; Creissen, G.; Mullineaux, P. Systemic Signaling and Acclimation in Response to Excess Excitation Energy in Arabidopsis. Science 1999, 284, 654–657. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.N.; Xu, Q.Z.; Li, W.C.; Yang, X.H.; Zheng, Q.; Li, B.; Li, Z.S.; Li, H.W. Long-Term High Light Stress Induces Leaf Senescence in Wheat (Triticum aestivum L.). Photosynthetica 2019, 57, 830–840. [Google Scholar] [CrossRef]
- Luschin-Ebengreuth, N.; Zechmann, B. Compartment-Specific Investigations of Antioxidants and Hydrogen Peroxide in Leaves of Arabidopsis Thaliana during Dark-Induced Senescence. Acta Physiol. Plant. 2016, 38, 133. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Salazar, C.; Mondal, H.A.; Shulaev, E.; Cortes, D.F.; Shuman, J.L.; Luo, X.; Shah, J.; Schlauch, K.; et al. Temporal-Spatial Interaction between Reactive Oxygen Species and Abscisic Acid Regulates Rapid Systemic Acclimation in Plants. Plant Cell 2013, 25, 3553–3569. [Google Scholar] [CrossRef] [PubMed]
- Zechmann, B.; Müller, M.; Zellnig, G. Modified Levels of Cysteine Affect Glutathione Metabolism in Plant Cells. In Sulfur Assimilation and Abiotic Stress in Plants; Khan, N.A., Singh, S., Umar, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 193–206. [Google Scholar]
- Ibrahim, I.A.; Jabbour, A.A.; Abdulmajeed, A.M.; Elhady, M.E.; Almaroai, Y.A.; Hashim, A.M. Adaptive Responses of Four Medicinal Plants to High Altitude Oxidative Stresses through the Regulation of Antioxidants and Secondary Metabolites. Agronomy 2022, 12, 3032. [Google Scholar] [CrossRef]
- Hashim, A.M.; Alharbi, B.M.; Abdulmajeed, A.M.; Elkelish, A.; Hassan, H.M.; Hozzein, W.N. Oxidative Stress Responses of Some Endemic Plants to High Altitudes by Intensifying Antioxidants and Secondary Metabolites Content. Plants 2020, 9, 869. [Google Scholar] [CrossRef] [PubMed]
- Gyugos, M.; Ahres, M.; Gulyás, Z.; Szalai, G.; Darkó, É.; Végh, B.; Boldizsár, Á.; Mednyánszky, Z.; Kar, R.K.; Dey, N.; et al. Role of Light-Intensity-Dependent Changes in Thiol and Amino Acid Metabolism in the Adaptation of Wheat to Drought. J. Agron. Crop Sci. 2019, 205, 562–570. [Google Scholar] [CrossRef]
- Choudhury, F.K.; Devireddy, A.R.; Azad, R.K.; Shulaev, V.; Mittler, R. Rapid Accumulation of Glutathione During Light Stress in Arabidopsis. Plant Cell Physiol. 2018, 59, 1817–1826. [Google Scholar] [CrossRef]
- Müller-Moulé, P.; Golan, T.; Niyogi, K.K. Ascorbate-Deficient Mutants of Arabidopsis Grow in High Light Despite Chronic Photooxidative Stress. Plant Physiol. 2004, 134, 1163. [Google Scholar] [CrossRef]
- Ntagkas, N.; Woltering, E.J.; Marcelis, L.F.M. Light Regulates Ascorbate in Plants: An Integrated View on Physiology and Biochemistry. Environ. Exp. Bot. 2018, 147, 271–280. [Google Scholar] [CrossRef]
- Ferreyra, M.L.F.; Serra, P.; Casati, P. Recent Advances on the Roles of Flavonoids as Plant Protective Molecules after UV and High Light Exposure. Physiol. Plant. 2021, 173, 736–749. [Google Scholar] [CrossRef]
- Araguirang, G.E.; Richter, A.S. Activation of Anthocyanin Biosynthesis in High Light—What Is the Initial Signal? New Phytol. 2022, 236, 2037–2043. [Google Scholar] [CrossRef]
- Zheng, X.T.; Yu, Z.C.; Tang, J.W.; Cai, M.L.; Chen, Y.L.; Yang, C.W.; Chow, W.S.; Peng, C.L. The Major Photoprotective Role of Anthocyanins in Leaves of Arabidopsis Thaliana under Long-Term High Light Treatment: Antioxidant or Light Attenuator? Photosynth. Res. 2021, 149, 25–40. [Google Scholar] [CrossRef]
- Cignoni, E.; Lapillo, M.; Cupellini, L.; Acosta-Gutiérrez, S.; Gervasio, F.L.; Mennucci, B. A Different Perspective for Nonphotochemical Quenching in Plant Antenna Complexes. Nat. Commun. 2021, 12, 7152. [Google Scholar] [CrossRef] [PubMed]
- El-Esawi, M.; Arthaut, L.D.; Jourdan, N.; D’Harlingue, A.; Link, J.; Martino, C.F.; Ahmad, M. Blue-Light Induced Biosynthesis of ROS Contributes to the Signaling Mechanism of Arabidopsis Cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef]
- Sun, T.; Rao, S.; Zhou, X.; Li, L. Plant Carotenoids: Recent Advances and Future Perspectives. Mol. Hortic. 2022, 2, 1–21. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, X.; Zhou, Y.; Tan, J.; Zhou, Y.; Gao, F. Small RNA Sequencing Revealed That MiR4415, a Legume-Specific MiRNA, Was Involved in the Cold Acclimation of Ammopiptanthus nanus by Targeting an L-Ascorbate Oxidase Gene and Regulating the Redox State of Apoplast. Front. Genet. 2022, 13, 870446. [Google Scholar] [CrossRef]
- Cui, G.; Li, B.; He, W.; Yin, X.; Liu, S.; Lian, L.; Zhang, Y.; Liang, W.; Zhang, P. Physiological Analysis of the Effect of Altitudinal Gradients on Leymus Secalinus on the Qinghai-Tibetan Plateau. PLoS ONE 2018, 13, e0202881. [Google Scholar] [CrossRef] [PubMed]
- Lima, C.S.; Ferreira-Silva, S.L.; Carvalho, F.E.L.; Lima Neto, M.C.; Aragão, R.M.; Silva, E.N.; Sousa, R.M.J.; Silveira, J.A.G. Antioxidant Protection and PSII Regulation Mitigate Photo-Oxidative Stress Induced by Drought Followed by High Light in Cashew Plants. Environ. Exp. Bot. 2018, 149, 59–69. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium Deficiency and High Light Intensity Enhance Activities of Superoxide Dismutase, Ascorbate Peroxidase, and Glutathione Reductase in Bean Leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Ghorbanpour, M.; Hatami, M. Spray Treatment with Silver Nanoparticles plus Thidiazuron Increases Anti-Oxidant Enzyme Activities and Reduces Petal and Leaf Abscission in Four Cultivars of Geranium (Pelargonium zonale) during Storage in the Dark. J. Hortic. Sci. Biotechnol. 2015, 89, 712–718. [Google Scholar] [CrossRef]
- Zhu, H.; Li, X.; Zhai, W.; Liu, Y.; Gao, Q.; Liu, J.; Ren, L.; Chen, H.; Zhu, Y. Effects of Low Light on Photosynthetic Properties, Antioxidant Enzyme Activity, and Anthocyanin Accumulation in Purple Pak-Choi (Brassica campestris ssp. Chinensis Makino). PLoS ONE 2017, 12, e0179305. [Google Scholar] [CrossRef]
- Cao, K.; Yu, J.; Xu, D.; Ai, K.; Bao, E.; Zou, Z. Exposure to Lower Red to Far-Red Light Ratios Improve Tomato Tolerance to Salt Stress. BMC Plant Biol. 2018, 18, 10–15. [Google Scholar] [CrossRef]
- Yu, W.; Liu, Y.; Song, L.; Jacobs, D.F.; Du, X.; Ying, Y.; Shao, Q.; Wu, J. Effect of Differential Light Quality on Morphology, Photosynthesis, and Antioxidant Enzyme Activity in Camptotheca Acuminata Seedlings. J. Plant Growth Regul. 2016, 1, 148–160. [Google Scholar] [CrossRef]
- Kang, C.H.; Yoon, E.K.; Muthusamy, M.; Kim, J.A.; Jeong, M.J.; Lee, S.I. Blue LED Light Irradiation Enhances L-Ascorbic Acid Content While Reducing Reactive Oxygen Species Accumulation in Chinese Cabbage Seedlings. Sci. Hortic. 2020, 261, 108924. [Google Scholar] [CrossRef]
- Danon, A.; Lin, R.; Xing, D.; Chai, T.; Zhou, J.; Liu, J. LSD1 and HY5 Antagonistically Regulate Red Light Induced-Programmed Cell Death in Arabidopsis. Front. Plant Sci. 2015, 1, 292. [Google Scholar]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d’Harlingue, A.; et al. Blue-Light Dependent Reactive Oxygen Species Formation by Arabidopsis Cryptochrome May Define a Novel Evolutionarily Conserved Signaling Mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef] [PubMed]
- Toldi, D.; Gyugos, M.; Darkó, É.; Szalai, G.; Gulyás, Z.; Gierczik, K.; Székely, A.; Boldizsár, Á.; Galiba, G.; Müller, M.; et al. Light Intensity and Spectrum Affect Metabolism of Glutathione and Amino Acids at Transcriptional Level. PLoS ONE 2019, 14, e0227271. [Google Scholar] [CrossRef]
- Li, Y.; Wu, L.; Jiang, H.; He, R.; Song, S.; Su, W.; Liu, H. Supplementary Far-Red and Blue Lights Influence the Biomass and Phytochemical Profiles of Two Lettuce Cultivars in Plant Factory. Molecules 2021, 26, 7405. [Google Scholar] [CrossRef]
- Grodzinski, B.; Lee, T.-M.; Pratap Singh, V.; Liu, W.; Zha, L.; Yang, Q.; Zhang, Y.; Zhou, C.; Shao, M. Regulation of Ascorbate Accumulation and Metabolism in Lettuce by the Red:Blue Ratio of Continuous Light Using LEDs. Front. Plant Sci. 2020, 11, 704. [Google Scholar]
- Badmus, U.O.; Crestani, G.; O’Connell, R.D.; Cunningham, N.; Jansen, M.A.K. UV-B Induced Accumulation of Tocopherol in Arabidopsis thaliana Is Not Dependent on Individual UV Photoreceptors. Plant Stress 2022, 5, 100105. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, S.; Yang, W.; Shang, X.; Fu, X. Light Quality Affects Flavonoid Production and Related Gene Expression in Cyclocarya Paliurus. J. Photochem. Photobiol. B 2018, 179, 66–73. [Google Scholar] [CrossRef]
- Laoué, J.; Fernandez, C.; Ormeño, E. Plant Flavonoids in Mediterranean Species: A Focus on Flavonols as Protective Metabolites under Climate Stress. Plants 2022, 11, 172. [Google Scholar] [CrossRef]
- Pizarro, L.; Stange, C. Light-Dependent Regulation of Carotenoid Biosynthesis in Plants. Cienc. e Investig. Agrar. 2009, 36, 143–162. [Google Scholar] [CrossRef]
- Kong, Y.; Nemali, K. Blue and Far-Red Light Affect Area and Number of Individual Leaves to Influence Vegetative Growth and Pigment Synthesis in Lettuce. Front. Plant Sci. 2021, 12, 1375. [Google Scholar] [CrossRef] [PubMed]
- Nam, T.G.; Kim, D.O.; Eom, S.H. Effects of Light Sources on Major Flavonoids and Antioxidant Activity in Common Buckwheat Sprouts. Food Sci. Biotechnol. 2018, 27, 169. [Google Scholar] [CrossRef] [PubMed]
- Rehman, M.; Fahad, S.; Saleem, M.H.; Hafeez, M.; Ur Rahman, M.H.; Liu, F.; Deng, G. Red Light Optimized Physiological Traits and Enhanced the Growth of Ramie (Boehmeria nivea L.). Photosynthetica 2020, 58, 922–931. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Park, Y.G.; Jeong, B.R. Physiological and Proteomic Insights Into Red and Blue Light-Mediated Enhancement of in Vitro Growth in Scrophularia Kakudensis—A Potential Medicinal Plant. Front. Plant Sci. 2021, 11, 607007. [Google Scholar] [CrossRef] [PubMed]
- Balogh, E.; Kalapos, B.; Ahres, M.; Boldizsár, Á.; Gierczik, K.; Gulyás, Z.; Gyugos, M.; Szalai, G.; Novák, A.; Kocsy, G. Far-Red Light Coordinates the Diurnal Changes in the Transcripts Related to Nitrate Reduction, Glutathione Metabolism and Antioxidant Enzymes in Barley. Int. J. Mol. Sci. 2022, 23, 7479. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Li, Y.F.; Jagadeeswaran, G. Functions of MicroRNAs in Plant Stress Responses. Trends Plant Sci. 2012, 17, 196–203. [Google Scholar] [CrossRef]
- Carbonell, T.; Gomes, A.V. MicroRNAs in the Regulation of Cellular Redox Status and Its Implications in Myocardial Ischemia-Reperfusion Injury. Redox Biol. 2020, 36, 101607. [Google Scholar] [CrossRef]
- He, J.; Jiang, B.-H. Interplay between Reactive Oxygen Species and MicroRNAs in Cancer. Curr. Pharmacol. Rep. 2016, 2, 82–90. [Google Scholar] [CrossRef]
- Barczak-Brzyżek, A.; Brzyżek, G.; Koter, M.; Siedlecka, E.; Gawroński, P.; Filipecki, M. Plastid Retrograde Regulation of MiRNA Expression in Response to Light Stress. BMC Plant Biol. 2022, 22, 150. [Google Scholar] [CrossRef]
- Cheng, X.; He, Q.; Tang, S.; Wang, H.; Zhang, X.; Lv, M.; Liu, H.; Gao, Q.; Zhou, Y.; Wang, Q.; et al. The MiR172/IDS1 Signaling Module Confers Salt Tolerance through Maintaining ROS Homeostasis in Cereal Crops. New Phytol. 2021, 230, 1017–1033. [Google Scholar] [CrossRef]
- Wang, L.; Bai, X.; Qiao, Y.; Si, L.; Yu, Z.; Ni, C.; Li, T.; Guo, C.; Xiao, K. Tae-MiR9674a, a MicroRNA Member of Wheat, Confers Plant Drought and Salt Tolerance through Modulating the Stomata Movement and ROS Homeostasis. Plant Biotechnol. Rep. 2022, 1, 3. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, M.; Shi, G.; Wang, L.; Ni, C.; Wang, Z.; Zhang, Y.; Xue, H.; Xiao, K. TaMIR5086, a MicroRNA Member in Triticum Aestivum, Confers Plants Drought Tolerance via Modulating Stomata Movement and Antioxidant Enzyme Activities. Int. J. Agric. Biol. 2021, 26, 384–392. [Google Scholar]
- Chen, C.; Liu, Y.; Xia, R. Jack of Many Trades: The Multifaceted Role Of MiR528 in Monocots. Mol. Plant 2019, 12, 1044–1046. [Google Scholar] [PubMed]
- Zhu, H.; Chen, C.; Zeng, J.; Yun, Z.; Liu, Y.; Qu, H.; Jiang, Y.; Duan, X.; Xia, R. MicroRNA528, a Hub Regulator Modulating ROS Homeostasis via Targeting of a Diverse Set of Genes Encoding Copper-Containing Proteins in Monocots. New Phytol. 2020, 225, 385–399. [Google Scholar] [CrossRef]
- Wang, M.; Guo, W.; Li, J.; Pan, X.; Pan, L.; Zhao, J.; Zhang, Y.; Cai, S.; Huang, X.; Wang, A.; et al. The MiR528- AO Module Confers Enhanced Salt Tolerance in Rice by Modulating the Ascorbic Acid and Abscisic Acid Metabolism and ROS Scavenging. J. Agric. Food Chem. 2021, 69, 8634–8648. [Google Scholar] [CrossRef]
- Mishra, V.; Singh, A.; Gandhi, N.; Das, S.S.; Yadav, S.; Kumar, A.; Sarkar, A.K. A Unique MiR775-GALT9 Module Regulates Leaf Senescence in Arabidopsis during Post-Submergence Recovery by Modulating Ethylene and the Abscisic Acid Pathway. Development 2022, 149, dev199974. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Feng, C.; Wang, Y.-T.; Zhang, X.; Wang, Y.-Y.; Sun, Y.-T.; Xiao, Y.-Q.; Zhai, Z.-F.; Zhou, X.; Du, B.-Y.; et al. MiR164g-MsNAC022 Acts as a Novel Module Mediating Drought Response by Transcriptional Regulation of Reactive Oxygen Species Scavenging Systems in Apple. Hortic. Res. 2022, 9, uhac192. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Xue, H.; Zhang, F.; Jiang, Q.; Yang, S.; Yue, P.; Wang, F.; Zhang, Y.; Li, L.; He, P.; et al. The MiR156/SPL Module Regulates Apple Salt Stress Tolerance by Activating MdWRKY100 Expression. Plant Biotechnol. J. 2021, 19, 311–323. [Google Scholar] [CrossRef]
- Dey, S.; Sarkar, A.; Chowdhury, S.; Singh, R.; Mukherjee, A.; Ghosh, Z.; Kundu, P. Heightened MiR6024-NLR Interactions Facilitate Necrotrophic Pathogenesis in Tomato. Plant Mol. Biol. 2022, 109, 717–739. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Y.-P.; Zhou, X.-H.; Yang, X.-M.; He, X.-R.; Feng, Q.; Zhu, Y.; Li, G.-B.; Wang, H.; Zhao, J.-H.; et al. Rice MiR1432 Fine-Tunes the Balance of Yield and Blast Disease Resistance via Different Modules. Rice 2021, 14, 87. [Google Scholar] [CrossRef]
- Feng, Y.; Qi, N.; Lei, P.; Wang, Y.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y.; Zhu, X. Gma-MiR408 Enhances Soybean Cyst Nematode Susceptibility by Suppressing Reactive Oxygen Species Accumulation. Int. J. Mol. Sci. 2022, 23, 14022. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Mhamdi, A.; Chaouch, S.; Han, Y.; Neukermans, J.; Marquez-Garcia, B.; Queval, G.; Foyer, C.H. Glutathione in Plants: An Integrated Overview. Plant. Cell Environ. 2012, 35, 454–484. [Google Scholar] [CrossRef] [PubMed]
- Sunkar, R.; Kapoor, A.; Zhu, J.K. Posttranscriptional Induction of Two Cu/Zn Superoxide Dismutase Genes in Arabidopsis Is Mediated by Downregulation of MiR398 and Important for Oxidative Stress Tolerance. Plant Cell 2006, 18, 2051–2065. [Google Scholar] [CrossRef]
- Dong, Q.; Hu, B.; Zhang, C. MicroRNAs and Their Roles in Plant Development. Front. Plant Sci. 2022, 13, 315. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C.; Mato, J.M.; Espinosa-Diez, C.; Lamas, S. MicroRNA-Mediated Regulation of Glutathione and Methionine Metabolism and Its Relevance for Liver Disease. Free Radic. Biol. Med. 2016, 100, 66. [Google Scholar] [CrossRef]
- Datta, R.; Mandal, K.; Boro, P.; Sultana, A.; Chattopadhyay, S. Glutathione Imparts Stress Tolerance against Alternaria Brassicicola Infection via MiRNA Mediated Gene Regulation. Plant Signal. Behav. 2022, 17, 2047352. [Google Scholar] [CrossRef]
- Cao, J.; Gulyás, Z.; Kalapos, B.; Boldizsár, Á.; Liu, X.; Pál, M.; Yao, Y.; Galiba, G.; Kocsy, G. Identification of a Redox-Dependent Regulatory Network of MiRNAs and Their Targets in Wheat. J. Exp. Bot. 2019, 70, 85–99. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Glutathione in Plants: Biosynthesis and Physiological Role in Environmental Stress Tolerance. Physiol. Mol. Biol. Plants 2017, 23, 249–268. [Google Scholar] [CrossRef]
- Jagadeeswaran, G.; Li, Y.F.; Sunkar, R. Redox Signaling Mediates the Expression of a Sulfate-Deprivation-Inducible MicroRNA395 in Arabidopsis. Plant J. 2014, 77, 85–96. [Google Scholar] [CrossRef]
- Liang, G.; Yang, F.; Yu, D. MicroRNA395 Mediates Regulation of Sulfate Accumulation and Allocation in Arabidopsis thaliana. Plant J. 2010, 62, 1046–1057. [Google Scholar] [CrossRef]
- Matthewman, C.A.; Kawashima, C.G.; Húska, D.; Csorba, T.; Dalmay, T.; Kopriva, S. MiR395 Is a General Component of the Sulfate Assimilation Regulatory Network in Arabidopsis. FEBS Lett. 2012, 586, 3242–3248. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; Ángeles Bermúdez, M.; Romero, L.C.; Gotor, C.; García, I. Cysteine Homeostasis Plays an Essential Role in Plant Immunity. New Phytol. 2012, 193, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Barozai, M.Y.K.; Baloch, I.A.; Din, M. Identification of MicroRNAs and Their Targets in Helianthus. Mol. Biol. Rep. 2012, 39, 2523–2532. [Google Scholar] [CrossRef]
- Hossain, R.; Quispe, C.; Saim Mohammad Saikat, A.; Jain, D.; Habib, A.; Janmeda, P.; Torequl Islam, M.; Durna Daştan, S.; Kumar, M.; Butnariu, M.; et al. Biosynthesis of Secondary Metabolites Based on the Regulation of MicroRNAs. Hindawi BioMed Res. Int. 2022, 2022, 20. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhao, G.; Zhang, S.; Li, Y.; Gu, H.; Li, Y.; Zhao, Q.; Qi, Y. Chloroplast-to-Nucleus Signaling Regulates MicroRNA Biogenesis in Arabidopsis. Dev. Cell 2019, 48, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Spencer, J.P.E.; Rice-Evans, C. Flavonoids: Antioxidants or Signalling Molecules? Free Radic. Biol. Med. 2004, 36, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Biosynthesis, F.; Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar]
- Gupta, O.P.; Karkute, S.G.; Banerjee, S.; Meena, N.L.; Dahuja, A. Contemporary Understanding of MiRNA-Based Regulation of Secondary Metabolites Biosynthesis in Plants. Front. Plant Sci 2017, 8, 374. [Google Scholar] [CrossRef]
- Yang, K.; Han, H.; Li, Y.; Ye, J.; Xu, F. Significance of MiRNA in Enhancement of Flavonoid Biosynthesis. Plant Biol. 2022, 24, 217–226. [Google Scholar] [CrossRef]
- Prakash Gupta, O.; Dahuja, A.; Sachdev, A.; Kumari, S.; Pradeep, J.K.; Vinutha, T.; Praveen, S. Conserved MiRNAs Modulate the Expression of Potential Transcription Factors of Isoflavonoid Biosynthetic Pathway in Soybean Seeds. Mol. Biol. Rep. 2019, 46, 3713–3730. [Google Scholar] [CrossRef]
- Dai, Z.; Tan, J.; Zhou, C.; Yang, X.; Yang, F.; Zhang, S.; Sun, S.; Miao, X.; Shi, Z. The OsmiR396-OsGRF8-OsF3H-Flavonoid Pathway Mediates Resistance to the Brown Planthopper in Rice (Oryza sativa). Plant Biotechnol. J. 2019, 17, 1657–1669. [Google Scholar] [CrossRef] [PubMed]
- Verhoeyen, M.E.; Bovy, A.; Collins, G.; Muir, S.; Robinson, S.; De Vos, C.H.R.; Colliver, S. Increasing Antioxidant Levels in Tomatoes through Modification of the Flavonoid Biosynthetic Pathway. J. Exp. Bot. 2002, 53, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Cheng, C.; Lin, Y.; Zhu, Q.; Lin, J.; Lai, Z. Combined Small RNA and Degradome Sequencing Reveals Complex MicroRNA Regulation of Catechin Biosynthesis in Tea (Camellia sinensis). PLoS ONE 2017, 12, e0171173. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Z.; Liu, D.; Zhang, K.; Li, A.; Mao, L. SQUAMOSA Promoter-Binding Protein-like Transcription Factors: Star Players for Plant Growth and Development. J. Integr. Plant Biol. 2010, 52, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative Regulation of Anthocyanin Biosynthesis in Arabidopsis by a MiR156-Targeted SPL Transcription Factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [CrossRef]
- Liu, R.; Lai, B.; Hu, B.; Qin, Y.; Hu, G.; Zhao, J. Identification of MicroRNAs and Their Target Genes Related to the Accumulation of Anthocyanins in Litchi Chinensis by High-Throughput Sequencing and Degradome Analysis. Front. Plant Sci. 2017, 7, 2059. [Google Scholar] [CrossRef]
- Qian, M.; Ni, J.; Niu, Q.; Bai, S.; Bao, L.; Li, J.; Sun, Y.; Zhang, D.; Teng, Y. Response of MiR156-SPL Module during the Red Peel Coloration of Bagging-Treated Chinese Sand Pear (Pyrus pyrifolia Nakai). Front. Physiol. 2017, 8, 550. [Google Scholar] [CrossRef]
- Saminathan, T.; Bodunrin, A.; Singh, N.V.; Devarajan, R.; Nimmakayala, P.; Jeff, M.; Aradhya, M.; Reddy, U.K. Genome-Wide Identification of MicroRNAs in Pomegranate (Punica granatum L.) by High-Throughput Sequencing. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef]
- Qu, D.; Yan, F.; Meng, R.; Jiang, X.; Yang, H.; Gao, Z.; Dong, Y.; Yang, Y.; Zhao, Z. Identification of MicroRNAs and Their Targets Associated with Fruit-Bagging and Subsequent Sunlight Re-Exposure in the “Granny Smith” Apple Exocarp Using High-Throughput Sequencing. Front. Plant Sci. 2016, 7, 550. [Google Scholar] [CrossRef]
- Zhao, C.; Li, T.; Zhao, Y.; Zhang, B.; Li, A.; Zhao, S.; Hou, L.; Xia, H.; Fan, S.; Qiu, J.; et al. Integrated Small RNA and MRNA Expression Profiles Reveal MiRNAs and Their Target Genes in Response to Aspergillus Flavus Growth in Peanut Seeds. BMC Plant Biol. 2020, 20, 1–16. [Google Scholar] [CrossRef]
- Wang, L.; Liu, N.; Wang, T.; Li, J.; Wen, T.; Yang, X.; Lindsey, K.; Zhang, X. The GhmiR157a- GhSPL10 Regulatory Module Controls Initial Cellular Dedifferentiation and Callus Proliferation in Cotton by Modulating Ethylene-Mediated Flavonoid Biosynthesis. J. Exp. Bot. 2018, 69, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Tong, Z.; He, F.; Li, X.; Sun, J. Integrated MRNA and MicroRNA Expression Analysis of Root Response to Phosphate Deficiency in Medicago Sativa. Front. Plant Sci. 2022, 13, 3444. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Burd, S.; Lers, A. MiR408 Is Involved in Abiotic Stress Responses in Arabidopsis. Plant J. 2015, 84, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Dixon, D.P.; Lapthorn, A.; Edwards, R. Plant Glutathione Transferases. Genome Biol. 2002, 3, 3004. [Google Scholar] [CrossRef]
- Wang, N.H.; Zhou, X.Y.; Shi, S.H.; Zhang, S.; Chen, Z.H.; Ali, M.A.; Ahmed, I.M.; Wang, Y.; Wu, F. An MiR156-Regulated Nucleobase-Ascorbate Transporter 2 Confers Cadmium Tolerance via Enhanced Anti-Oxidative Capacity in Barley. J. Adv. Res. 2023, 44, 23–37. [Google Scholar] [CrossRef]
- Wang, Y.; Feng, C.; Zhai, Z.; Peng, X.; Wang, Y.; Sun, Y.; Li, J.; Shen, X.; Xiao, Y.; Zhu, S.; et al. The Apple MicroR171i-SCARECROW-LIKE PROTEINS26.1 Module Enhances Drought Stress Tolerance by Integrating Ascorbic Acid Metabolism. Plant Physiol. 2020, 184, 194–211. [Google Scholar] [CrossRef]
- Hernando, C.E.; Garcia, C.; Mateos, J.L. Casting Away the Shadows: Elucidating the Role of Light-Mediated Posttranscriptional Control in Plants. Photochem. Photobiol. 2017, 93, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Szakonyi, D.; Mora-Garcia, S.; Umesh, A.; Reddy, K.; Henriques, R.; Sánchez-Retuerta, C.; Suaréz-López, P. Under a New Light: Regulation of Light-Dependent Pathways by Non-Coding RNAs. Front. Plant Sci. 2018, 9, 962. [Google Scholar]
- Choi, S.W.; Ryu, M.Y.; Viczián, A.; Jung, H.J.; Kim, G.M.; Arce, A.L.; Achkar, N.P.; Manavella, P.; Dolde, U.; Wenkel, S.; et al. Light Triggers the MiRNA-Biogenetic Inconsistency for De-Etiolated Seedling Survivability in Arabidopsis thaliana. Mol. Plant 2020, 13, 431–445. [Google Scholar] [CrossRef]
- Park, S.J.; Choi, S.W.; Kim, G.M.; Møller, C.; Pai, H.S.; Yang, S.W. Light-Stabilized FHA2 Suppresses MiRNA Biogenesis through Interactions with DCL1 and HYL1. Mol. Plant 2021, 14, 647–663. [Google Scholar] [CrossRef]
- Cho, S.K.; Chaabane, S.B.; Shah, P.; Poulsen, C.P.; Yang, S.W. COP1 E3 Ligase Protects HYL1 to Retain MicroRNA Biogenesis. Nat. Commun. 2014, 5, 5867. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.; Habermann, K.; Arif, M.A.; Top, O.; Frank, W. Identification of Small RNAs During High Light Acclimation in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 1179. [Google Scholar] [CrossRef] [PubMed]
- Chung, P.J.; Park, B.S.; Wang, H.; Liu, J.; Jang, I.C.; Chua, N.H. Light-Inducible MiR163 Targets PXMT1 Transcripts to Promote Seed Germination and Primary Root Elongation in Arabidopsis. Plant Physiol. 2016, 170, 1772–1782. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Xu, J.; Lin, R.; Song, J.; Shao, S.; Yu, J.; Zhou, Y. Light-Induced HY5 Functions as a Systemic Signal to Coordinate the Photoprotective Response to Light Fluctuation. Plant Physiol. 2020, 184, 1181–1193. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Lian, H.; Li, H.; Xu, Y.; Zhang, H. HY5 Regulates Light-Responsive Transcription of MicroRNA163 to Promote Primary Root Elongation in Arabidopsis Seedlings. J. Integr. Plant Biol. 2021, 63, 1437–1450. [Google Scholar] [CrossRef]
- Zhang, H.; He, H.; Wang, X.; Wang, X.; Yang, X.; Li, L.; Deng, X.W. Genome-Wide Mapping of the HY5-Mediated Gene Networks in Arabidopsis That Involve Both Transcriptional and Post-Transcriptional Regulation. Plant J. 2011, 65, 346–358. [Google Scholar] [CrossRef]
- Panigrahy, M.; Panigrahi, K.C.S.; Poli, Y.; Ranga, A.; Majeed, N. Integrated Expression Analysis of Small RNA, Degradome and Microarray Reveals Complex Regulatory Action of MiRNA during Prolonged Shade in Swarnaprabha Rice. Biology 2022, 11, 798. [Google Scholar] [CrossRef]
- Sekhar, S.; Das, S.; Panda, D.; Mohanty, S.; Mishra, B.; Kumar, A.; Navadagi, D.B.; Sah, R.P.; Pradhan, S.K.; Samantaray, S.; et al. Identification of MicroRNAs That Provide a Low Light Stress Tolerance-Mediated Signaling Pathway during Vegetative Growth in Rice. Plants 2022, 11, 2558. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, Y.; Wang, H.; Ma, X.; Wang, B.; Wu, G.; Wang, H. Phytochrome-Interacting Factors Directly Suppress MIR156 Expression to Enhance Shade-Avoidance Syndrome in Arabidopsis. Nat. Commun. 2017, 8, 348. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, D.; Peng, Z.; Wang, L.; Gao, Z. Identification and Characterization of MicroRNAs in the Leaf of Ma Bamboo (Dendrocalamus latiflorus) by Deep Sequencing. PLoS ONE 2013, 8, e78755. [Google Scholar] [CrossRef]
- Li, Y.; Varala, K.; Hudson, M.E. A Survey of the Small RNA Population during Far-Red Light-Induced Apical Hook Opening. Front. Plant Sci. 2014, 5, 156. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Li, M.; Zhou, Y.; Guo, T.; Liu, Y.; Zhang, H.; Fang, Y. Coordinated Regulation of Arabidopsis MicroRNA Biogenesis and Red Light Signaling through Dicer-like 1 and Phytochrome-Interacting Factor 4. PLoS Genet. 2018, 14, e1007247. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, A.M.; Singh, A.; Singh, R.; Verma, A.K.; Roy, S. Modulation of MiRNA Expression in Natural Populations of A. thaliana along a Wide Altitudinal Gradient of Indian Himalayas. Sci. Reports 2019, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Facella, P.; Carbone, F.; Placido, A.; Perrotta, G. Cryptochrome 2 Extensively Regulates Transcription of the Chloroplast Genome in Tomato. FEBS Open Bio 2017, 7, 456–471. [Google Scholar] [CrossRef]
- Zhou, B.; Fan, P.; Li, Y.; Yan, H.; Xu, Q. Exploring MiRNAs Involved in Blue/UV-A Light Response in Brassica Rapa Reveals Special Regulatory Mode during Seedling Development. BMC Plant Biol. 2016, 16, 111. [Google Scholar] [CrossRef] [PubMed]
- Pashkovskiy, P.; Kreslavski, V.; Khudyakova, A.; Kosobryukhov, A.; Kuznetsov, V.V.; Allakhverdiev, S.I. Influence of Phytochromes on MicroRNA Expression, Phenotype, and Photosynthetic Activity in A. thaliana Phy Mutants under Light with Different Spectral Composition. Photosynthetica 2022, 61, 135–144. [Google Scholar] [CrossRef]
- Dong, F.; Wang, C.; Dong, Y.; Hao, S.; Wang, L.; Sun, X.; Liu, S. Differential Expression of MicroRNAs in Tomato Leaves Treated with Different Light Qualities. BMC Genom. 2020, 21, 37. [Google Scholar] [CrossRef]
- Tsai, H.L.; Li, Y.H.; Hsieh, W.P.; Lin, M.C.; Ahn, J.H.; Wu, S.H. HUA ENHANCER1 Is Involved in Posttranscriptional Regulation of Positive and Negative Regulators in Arabidopsis Photomorphogenesis. Plant Cell 2014, 26, 2858–2872. [Google Scholar] [CrossRef]
- Chen, D.; Xu, G.; Tang, W.; Jing, Y.; Ji, Q.; Fei, Z.; Lin, R. Antagonistic Basic Helix-Loop-Helix/BZIP Transcription Factors Form Transcriptional Modules That Integrate Light and Reactive Oxygen Species Signaling in Arabidopsis. Plant Cell 2013, 25, 1657–1673. [Google Scholar] [CrossRef]
- Casati, P. Analysis of UV-B Regulated MiRNAs and Their Targets in Maize Leaves. Plant Signal. Behav. 2013, 8, e26758. [Google Scholar] [CrossRef]
- Jia, X.; Ren, L.; Chen, Q.J.; Li, R.; Tang, G. UV-B-Responsive MicroRNAs in Populus tremula. J. Plant Physiol. 2009, 166, 2046–2057. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, G.; Zhang, W. UV-B Responsive MicroRNA Genes in Arabidopsis thaliana. Mol. Syst. Biol. 2007, 3, 103. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Ortiz, G.; Johansson, H.; Lee, K.P.; Bou-Torrent, J.; Stewart, K.; Steel, G.; Rodríguez-Concepción, M.; Halliday, K.J. The HY5-PIF Regulatory Module Coordinates Light and Temperature Control of Photosynthetic Gene Transcription. PLoS Genet. 2014, 10, e1004416. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Yang, Y.; Yang, Y.; Zhang, X.; Su, Y.; Guo, J.; Que, Y.; Xu, L. Identification of Low-Nitrogen-Related MiRNAs and Their Target Genes in Sugarcane and the Role of MiR156 in Nitrogen Assimilation. Int. J. Mol. Sci. 2022, 23, 13187. [Google Scholar] [CrossRef]
- Zhao, H.; Lin, K.; Ma, L.; Chen, Q.; Gan, S.; Li, G. Arabidopsis NUCLEAR FACTOR Y A8 Inhibits the Juvenile-to-Adult Transition by Activating Transcription of MIR156s. J. Exp. Bot. 2020, 71, 4890–4902. [Google Scholar] [CrossRef]
- Xing, L.; Zhu, M.; Luan, M.; Zhang, M.; Jin, L.; Liu, Y.; Zou, J.; Wang, L.; Xu, M. MiR169q and NUCLEAR FACTOR YA8 Enhance Salt Tolerance by Activating PEROXIDASE1 Expression in Response to ROS. Plant Physiol. 2022, 188, 608–623. [Google Scholar] [CrossRef]
- Myers, Z.A.; Kumimoto, R.W.; Siriwardana, C.L.; Gayler, K.K.; Risinger, J.R.; Pezzetta, D.; Holt, B.F. Nuclear factor y, Subunit C (NF-YC) Transcription Factors Are Positive Regulators of Photomorphogenesis in Arabidopsis thaliana. PLoS Genet. 2016, 12, e1006333. [Google Scholar] [CrossRef]
- Rusaczonek, A.; Czarnocka, W.; Willems, P.; Sujkowska-Rybkowska, M.; Van Breusegem, F.; Karpiński, S. Phototropin 1 and 2 Influence Photosynthesis, Uv-c Induced Photooxidative Stress Responses, and Cell Death. Cells 2021, 10, 200. [Google Scholar] [CrossRef]
- Czégény, G.; Wu, M.; Dér, A.; Eriksson, L.A.; Strid, Å.; Hideg, É. Hydrogen Peroxide Contributes to the Ultraviolet-B (280-315 Nm) Induced Oxidative Stress of Plant Leaves through Multiple Pathways. FEBS Lett. 2014, 588, 2255–2261. [Google Scholar] [CrossRef]
- Yue, E.; Liu, Z.; Li, C.; Li, Y.; Liu, Q.; Xu, J.H. Overexpression of MiR529a Confers Enhanced Resistance to Oxidative Stress in Rice (Oryza sativa L.). Plant Cell Rep. 2017, 36, 1171–1182. [Google Scholar] [CrossRef]
- Ali, E.; Hussain, S.; Hussain, N.; Ullah Kakar, K.; Munawar Shah, J.; Hassan Raza Zaidi, S.; Jan, M.; Zhang, K.; Ali Khan, M.; Imtiaz, M. Tocopherol as Plant Protector: An Overview of Tocopherol Biosynthesis Enzymes and Their Role as Antioxidant and Signaling Molecules. Acta Physiol. Plant. 2022, 44, 20. [Google Scholar] [CrossRef]
- Carroll, B.; Xin, M.; Yan, Y.-M.; Lv, D.-W.; Zhen, S.; Zhu, G.-R.; Bian, Y.-W.; Chen, G.-X.; Han, C.-X.; Yu, Z.-T. High-Throughput Sequencing Reveals H2O2 Stress-Associated MicroRNAs and a Potential Regulatory Network in Brachypodium distachyon Seedlings. Front. Plant Sci. 2016, 7, 1567. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| miRNA | Species | Targeted Genes | Related Stress | References |
|---|---|---|---|---|
| miR156 | Malus domestica | SPL | Salinity | [91] |
| miR6024 | Solanum lycopersicum | NLR | Biotic stress | [92] |
| miR775 | Arabidopsis thaliana | GALT9 | Hypoxia | [89] |
| miR164g | Malus domestica | MsNAC022 | Drought | [90] |
| miR9674a | Triticum aesivum | Mta/sah nucleosidase | Drought, Salt, Osmotic | [84] |
| Serine/threonie protein | ||||
| TRAP dicarboxylate transporter | ||||
| AUX/IAA1 | ||||
| peptidase S16 | ||||
| RUBP activase A | ||||
| TRAP dicarboxylate transporter | ||||
| miR5086 | Triticum aesivum | TaTIF, TaTP, TaRPS | Drought | [85] |
| TaRPT, TaSF, TaAP | ||||
| miR172 | Triticum aesivum | IDS1 | Salinity | [83] |
| miR528 | Oryza sativa | AO | Salinity | [88] |
| miR1432 | Oryza sativa | OsEFH1 | Biotic stress | [93] |
| miR408 | Glycine max | PR1, OXI1 | Biotic stress | [94] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gulyás, Z.; Székely, A.; Kulman, K.; Kocsy, G. Light-Dependent Regulatory Interactions between the Redox System and miRNAs and Their Biochemical and Physiological Effects in Plants. Int. J. Mol. Sci. 2023, 24, 8323. https://doi.org/10.3390/ijms24098323
Gulyás Z, Székely A, Kulman K, Kocsy G. Light-Dependent Regulatory Interactions between the Redox System and miRNAs and Their Biochemical and Physiological Effects in Plants. International Journal of Molecular Sciences. 2023; 24(9):8323. https://doi.org/10.3390/ijms24098323
Chicago/Turabian StyleGulyás, Zsolt, András Székely, Kitti Kulman, and Gábor Kocsy. 2023. "Light-Dependent Regulatory Interactions between the Redox System and miRNAs and Their Biochemical and Physiological Effects in Plants" International Journal of Molecular Sciences 24, no. 9: 8323. https://doi.org/10.3390/ijms24098323
APA StyleGulyás, Z., Székely, A., Kulman, K., & Kocsy, G. (2023). Light-Dependent Regulatory Interactions between the Redox System and miRNAs and Their Biochemical and Physiological Effects in Plants. International Journal of Molecular Sciences, 24(9), 8323. https://doi.org/10.3390/ijms24098323