1. Introduction
In vivo reactive oxygen species (ROS) are a secondary product in many cellular processes, such as the electron transport chain, lipid peroxidation, and the xanthine oxidase system. The traditional views of ROS only address their roles related to aging and disease. However, in recent years, they have gained attention as signaling molecules in many physiological processes, such as cell differentiation, vascular pressure regulation, and sperm physiological maturation, i.e., capacitation and hyperactivation [
1].
ROS include dioxygen (O
2) molecules reduced by one (radical) or two (nonradical) electrons, and these are classified by their reduction potential. Superoxide anion (O
2•-) and hydrogen peroxide (H
2O
2) are ROS that have the potential to serve as second messengers because cells have specialized systems for ROS production, such as the NADPH oxidase (NOX) system [
1,
2]. NOXs catalyze ROS production by transferring electrons from NADPH to O
2 in a highly regulated manner. NOXs’ function and regulation were first described in phagocytic cells. The phagocytic ROS production system Nox2/gp91
phox is composed of several subunits: the catalytic and transmembrane subunit flavocytochrome b588 integrated by Nox2/gp91
phox and p22
phox, and the cytosolic and regulatory proteins p47
phox, p67
phox, as well as the small GTPase, Rac1 [
2,
3].
In mammalian sperm processes such as capacitation, acrosome reaction and motility are sensitive to ROS such as O
2•-, H
2O
2, and nitric oxide (NO) [
4,
5]. Human spermatozoa were the first cells in which physiological ROS production was reported [
6]. The detection of NOX5, whose activity is calcium-dependent, is responsible for the oxidative stress produced during capacitation and regulates capacitation and motility in human spermatozoa [
7]. However, in other mammals, such as rodents (i.e., mice, rats, and guinea pigs), the
nox5 gene is absent from their genomes [
2]. Even so, ROS production has been reported in mouse and rat spermatozoa. Members of the NOX family, NOX2 and NOX3, respectively, have been observed in sperm cells, which suggests that ROS production by NOX family members plays a significant role in physiological processes, such as capacitation, acrosome reaction, and mobility development [
7,
8,
9]. Interestingly, the NOX1 and NOX2 activator p67phox is a Rac1 target, so Rac is essential to regulate NOX1 and NOX2 activity [
2,
3].
ROS production through NADPH oxidases has been correlated with the activation of calcium-dependent proteases and calpains [
10,
11]. Calpain activity is essential for fertilization [
12], since it allows mammalian spermatozoa to acquire their fertilizing ability by participating in physiological processes such as capacitation, acrosomal reaction, and sperm mobility [
13,
14,
15,
16,
17], as well as in fowl sperm mobility [
18]. Calpains-1 and -2 are the two isoforms of the calpain family members found in human, mouse, and guinea pig spermatozoa [
12,
14,
17]. Although the mechanism by which these proteases regulate capacitation, acrosomal reaction, and mobility is not well understood, calpain-1 is known to regulate remodeling of the spectrin cytoskeleton [
14], lipid raft rearrangement, and activation of the Src kinase family [
15].
Given the existence of multiple members of the NOX family in mammal cells, and the presence of calpains [
13,
14,
15,
16,
17] and Rac1 [
19,
20] in mammalian spermatozoa, as well as the importance of ROS in several sperm processes that prepare them for fertilization, such as capacitation, acrosomal reaction, and mobility [
21], the objective of this work was to determine which members of the NOX family are responsible for the production of ROS in guinea pig spermatozoa. Furthermore, we examined the participation of ROS produced by NOX in capacitation, acrosomal reaction, and mobility, and whether the activity of the NOX present in guinea pig spermatozoa is dependent on calpain. The results showed that, unlike human and stallion spermatozoa [
7,
22], two different NOX members are present in guinea pig spermatozoa. We also found that the production of ROS by these NOXs is related to capacitation, acrosomal integrity, and mobility. The results also suggested that NOX2 and NOX4 interact in the non-capacitated state but not in the capacitated state. Such interactions and NOX activity are dependent on calpain activity.
3. Discussion
ROS production in mammalian spermatozoa is a consequence of the different signaling pathways in these cells. ROS activate other signaling pathways related to processes such as capacitation, acrosomal reaction, and motility [
1,
4,
21]. Despite the physiological and pathological evidence of the effects of ROS on spermatozoa, the identity of the NADPH oxidases responsible for ROS production remains uncertain, although two different NOXs, NOX2 and NOX5, are present in mammalian spermatozoa [
7,
9,
23,
24,
25]. The present work demonstrates clear evidence of the presence of two different NOXs in mouse and guinea pig spermatozoa, NOX2 and NOX4, which increase the levels of O
2•- and H
2O
2 during capacitation. The evidence presented here directly relates NOX2 and NOX4 to sperm processes, such as capacitation, spontaneous acrosomal reaction (sAR), and motility. Notably, our results reveal an unprecedented mechanism related to NOX family activation, specifically NOX2 and NOX4, which occurs during capacitation and involves the Ca
2+-dependent protease calpain.
Different reports suggest that ROS production in mammalian spermatozoa could be the consequence of a single member of the NOX family. NOX5 occurs in human, stallion, and canine spermatozoa [
7,
22,
23,
26], and NOX2 in mouse, rat and goat spermatozoa [
8,
9,
25]. This suggestion is supported by the study by Musset et al., which demonstrated that human spermatozoa do not express NOX1, NOX2, or NOX4 [
7]. Our results show that, as in other rodents [
24,
25], NOX2 is also expressed in guinea pig spermatozoa (
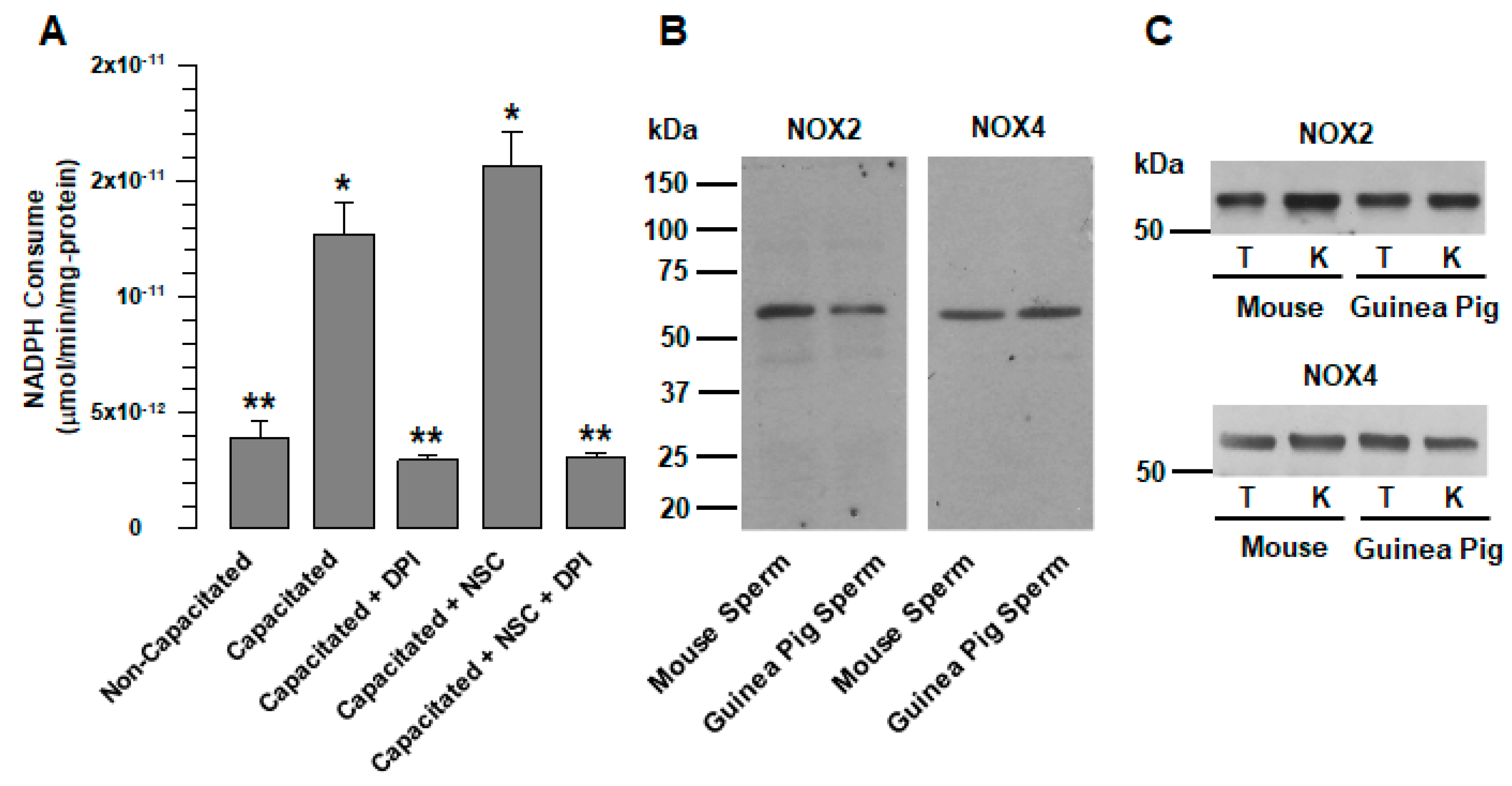
Figure 1B). At the same time, analysis of the consumption of NADPH revealed the presence of another member of the NOX family in guinea pig spermatozoa, NOX4. ROS production by NOX2 requires active Rac1 [
3]; therefore, the increase in NADPH consumption that occurs during capacitation (
Figure 1A) was not suppressed by a specific Rac1 inhibitor (NSC23766). In contrast, NADPH consumption increased (
Figure 1A); this is indicative of the presence of other NOXs not regulated by Rac1, and resulted in NOX4, which was corroborated by WB (
Figure 1B) and immunolocalization (
Figure 6A). It is possible that spermatozoa express not only NOX2 and NOX4 but also NOX1 and NOX3. Nevertheless, this remains to be clarified in a later study.
Our results show the presence of two different NOXs in guinea pigs and mouse spermatozoa, but further evidence also suggests that both NOX2 and NOX4 are active in guinea pig spermatozoa. In the case of NOX2, we know that the main product of this enzyme is the O
2•-, which is transmuted to H
2O
2 [
2]. NSC23766 and VAS2870 inhibit superoxide ion production at levels found in non-capacitated spermatozoa (
Figure 2B). Thus, the production of the O
2•- is blocked by inhibiting Rac1, indicating that this anion is produced by a NOX dependent on Rac1, such as NOX2. However, Rac1 inhibition did not reduce H
2O
2 levels; these were reduced by ML171 (
Figure 2C), a more specific inhibitor for NOX4 that does not affect NOX2 activity [
27]. Therefore, we suggest that the production of H
2O
2 under this condition must be due to a NOX not dependent on Rac1, such as NOX4. The fact that NOX2 inhibition does not reduce H
2O
2 levels during capacitation does not rule out the transmutation from O
2•- to H
2O
2 under normal conditions. It is essential to consider that the reduction in H
2O
2 levels compared to those observed for non-capacitated spermatozoa is only achieved when NSC23766 and ML171 were tested together (
Figure 2C). In conclusion, our data clearly show that NOX2 and NOX4 are responsible for ROS production during capacitation.
The use of ML171 (2 µM) in this work implies the possibility that NOX1, NOX4, and NOX5 activity may be affected [
27]. First, it is known that NOX5 is an NADPH oxidase exclusive to human beings [
7]. Since the gene does not exist in organisms of the rodent order [
2], there is no interference in the production of ROS in the spermatozoa of rodents such as guinea pigs and mice. In the case of NOX1, ML171 concentrations less than 0.5 µM specifically inhibit NOX1 [
28]. However, at these concentrations, the production of H
2O
2 did not show changes concerning capacitated spermatozoa in the absence of ML171. NOX1, if present in rodent spermatozoa, would not be involved in the different physiological processes experienced by the sperm before fertilizing the egg. This is supported by the fact that NOX1-deficient mice are fertile [
28]. Finally, the NOX2 activity could not have been altered by ML171, since it is only affected at concentrations greater than 10 µM [
27,
28].
During capacitation, O
2•- and H
2O
2 levels rose due to the increased activity of the enzymes responsible for producing these ROS [
1,
4,
29]. Several enzyme pathways responsible for ROS production and their participation in processes such as capacitation, acrosomal reaction, and motility have been characterized [
29]. However, except for NOX5 [
7], little is known about the participation of NOX2 and NOX4 in ROS production and its impact on sperm physiology. Our results clearly show that NOX2 and NOX4 inhibition by an agonist of both NOXs and VAS2879 [
27] almost entirely prevents the production of O
2•- and reduced the levels of H
2O
2 (
Figure 2B,C). However, this low level of H
2O
2 production does not support capacitation (
Figure 4 and
Figure 5) and directly affects sperm processes such as acrosomal reaction and the activation of progressive and hyperactivated motility. Therefore, the inhibition of O
2•- and H
2O
2 production by VAS2870 during capacitation directly relates NOX2 and NOX4 to physiological processes such as capacitation and motility, which require an adequate level of O
2•- and H
2O
2 produced by NOX2 and NOX4 for these sperm processes to occur correctly. In this regard, it is well known that O
2•- and H
2O
2 are related to capacitation, acrosomal reaction, and motility by activating different kinases, such as PKA, PKC, and MEK, which are fully involved in sperm physiology [
29,
30].
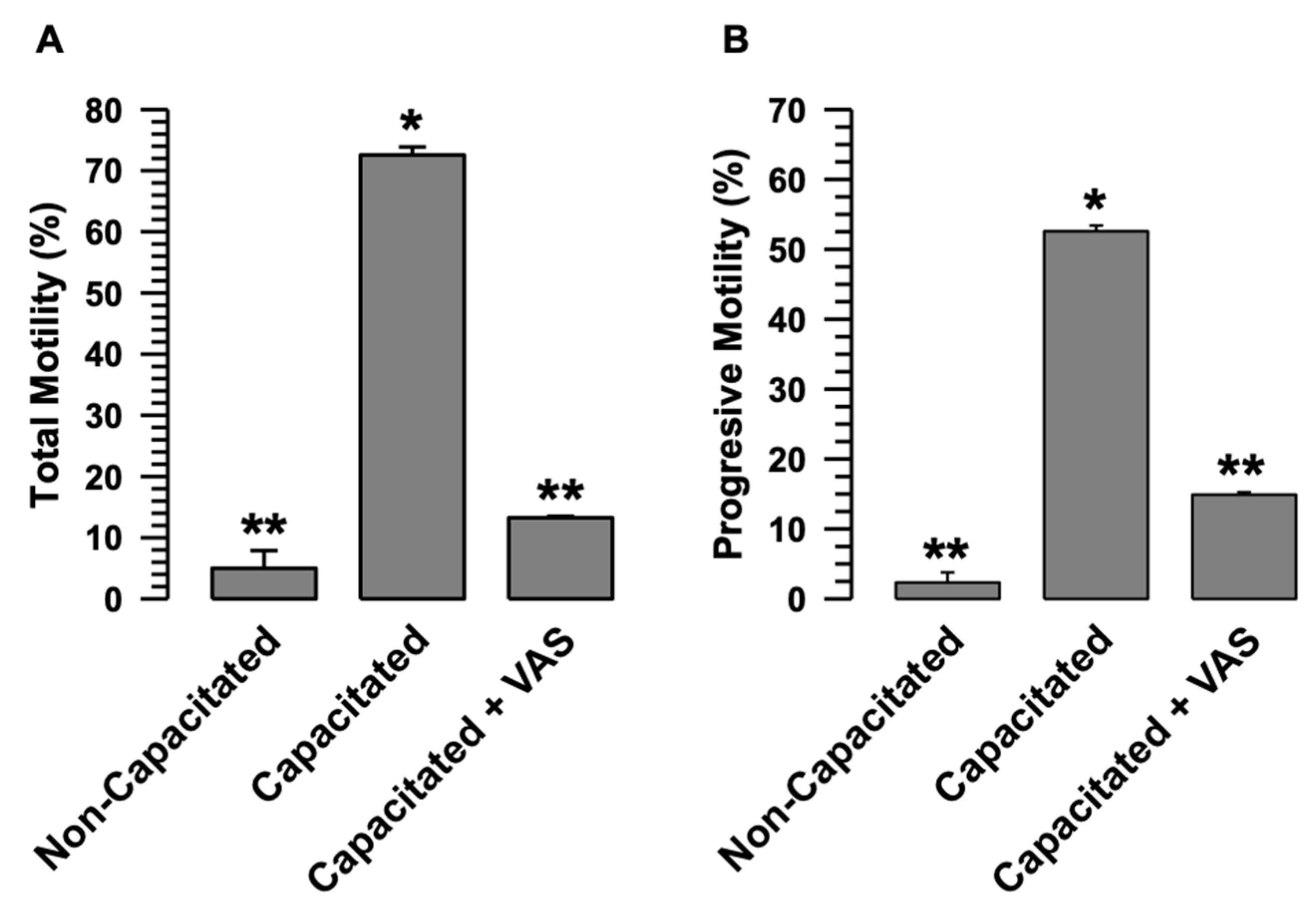
The early increase in the B and AR patterns experienced by sperm when VAS2870 inhibits H
2O
2 production suggests that spermatozoa undergo early capacitation; therefore, sAR is also increased early (
Figure 3A,B). These data appear to be contradictory regarding the non-increase in PYP during capacitation in the presence of VAS2870, and suggest that the spermatozoa would not undergo capacitation (
Figure 3C,D). The development of the B and AR patterns through the CTC assay is dependent on Ca
2+, since CTC interacts with membrane proteins in the presence of this cation [
31]. While the adequate production of H
2O
2 during capacitation is related to the activation of different kinases, such as PKC and PKA, these kinases show low activity when capacitation is performed in an environment with low levels of H
2O
2 [
29]. We suggest that both processes, the association of CTC with membrane proteins and PYP, occur independently during capacitation, since, despite the low production of H
2O
2 when NOX2 and NOX4 are inhibited, a continuous sperm influx Ca
2+ (
Figure 5) and, therefore, the association between CTCs and membrane proteins, should occur during capacitation. In this sense, a recent study reported that in human sperm, during capacitation, progesterone increases Ca
2+ influx and ROS production by NOX5. Still, the inhibition of NOX5 by DPI, in both the presence and absence of progesterone, reduces the production of ROS but not the influx of Ca
2+ [
32]. We also suggest that normal H
2O
2 production is required for capacitation and AR events to occur normally. Low H
2O
2 production during capacitation induces a higher Ca
2+ uptake, leading to the early occurrence of these events.
The presented results reveal a novel mechanism for the activation of NOXs, in which calpain-1, the calpain expressed in guinea pig spermatozoa [
14], plays a predominant role. In spermatozoa in a non-capacitated state, NOX2 and NOX4 are physically associated. In this physiological state, the production of O
2•- and H
2O
2 is deficient (
Figure 3 and
Figure 7). Therefore, we suggest that the interaction between NOX2 and NOX4 keeps these two NADPH oxidases inactive or at very low activity levels (
Figure 7A). The interaction between NOX2 and NOX4 changes when spermatozoa enter the capacitated state, separating and increasing their activity (
Figure 3 and
Figure 7). NOX2-NOX4 interaction occurs independently of their activity since, even in the presence of VAS2870, the NOXs are separated (
Figure 6B). Additionally, as H
2O
2 production increases only in the sperm-capacitated state (
Figure 3 and
Figure 7), we propose that the separation of NOX2 and NOX4 is a crucial point in the activation of these two NOXs, and also suggest that the increase in ROS causes capacitation and hypermotility to occur without alterations.
Furthermore, calpain-1 is known for its importance in sperm processes, including capacitation, acrosomal reaction, and motility, in different mammals’ spermatozoa [
12,
13,
14,
16,
18]. Although different relationships between NOXs and calpain have been reported, wherein NOX2 and NOX4 regulate calpain activity in neuropathological processes [
10,
11,
33], a mechanism where calpain-1 is related to the activation of NOX2 and NOX4 has not been previously reported. Our results suggest that calpain-1 activation is an essential and critical requirement for capacitation and motility to occur through the separation of NOX2 and NOX4, leading to an increase in O
2•- and H
2O
2, which are directly related to capacitation and sperm motility [
1,
4,
29].
We do not know whether the NOX2/NOX4 interaction is direct or whether other proteins are involved. The relationship between NOX2 and NOX5 and caveolin-1 has been characterized, and it has been suggested that caveolin-1 is a negative regulator of these two NOXs [
34,
35]. Additionally, the caveolin-1/NOX2 interaction was observed in goat spermatozoa, and NOX2 activity is closely related to the reorganization of lipid rafts that occurs during capacitation [
9]. The relocation experienced by NOX2 and NOX4 during capacitation (
Figure 7A) is similar to that experienced by lipid rafts in guinea pig spermatozoa [
15]. Although neither caveolins nor flotillins are targeted by calpains, we suggest that these proteins, which are related to lipid rafts, could be associated with the reorganizing of these two NOXs that occurs after their separation. Recently, in cardiomyocytes, it was reported that the NOX2 and NOX4 activity depends on their interaction with LRRC8A through its LRRD domain [
34]. However, like lipid raft proteins, LRRC8A is not targeted by calpain. It is important to note that, in the work of Huo et al. (2021), as in this work, the involved NADPH oxidases are NOX2 and NOX4, suggesting that these two NOXs could have an important relationship in multiple cellular and physiological processes. Therefore, it remains to be elucidated whether the NOX2/NOX4 interaction is direct or mediated by other proteins.
In conclusion, our results indicate that the production of O2•- and H2O2 during capacitation is related to at least two NOXs: NOX2 and NOX4. The inhibition of these two NOXs implies a low production of O2•- and H2O2 during capacitation and, as a result, capacitation, acrosome reaction and motility are affected, directly relating these two NOXs with the physiological processes that enable spermatozoa to fertilize. Therefore, the normal production of ROS by NOX2 and NOX4 during capacitation is essential for events such as capacitation, acrosomal reaction, and motility to occur normally. Finally, we present a new regulatory mechanism for NOXs, directed by the calcium-dependent protease calpain.
4. Materials and Methods
4.1. Reagents
All reagents used in this work were obtained from Sigma-Aldrich (St. Louis, MO, USA), except where otherwise indicated.
4.2. Experimental Animals
Male Dunkin-Hartley guinea pigs (Cavia porcellus) with a mean weight from 800 to 900 g were used to isolate testicles, epididymis, and vas deferens. Experimental animals were handled in accordance with the Mexican Official Norm for Laboratory Animals (NOM-062-ZOO-1999) and protocols from the Internal Committee for the Care and Use of Laboratory Animals, Cinvestav-IPN (CICUAL No. 321-02), following American Veterinary Medical Association guidelines.
4.3. Capacitation Assay
Spermatozoa were obtained from the vas deferens of guinea pigs in 2 mL of phosphate-buffered saline (PBS, pH 7.4) perfusion. The cells were adjusted to 3.5 × 107 cells/mL and incubated at 37 °C during and until capacitation. We used minimal capacitate medium supplemented with sodium pyruvate and lactic acid (MCM-PL (NaCl 105.8 mM, CaCl2 1.8 mM, NaHCO3 25.1 mM, lactic acid 20 mM, sodium pyruvate 0.25 mM and HEPES 2.5 mM, pH 7.8)) to induce capacitation. For non-capacitive conditions, the cell suspension (3.5 × 107 cell/mL) was incubated in MCM-PL lacking CaCl2 and NaHCO3 at 37 °C.
4.4. Pharmacologic Inhibitors
ROS production in guinea pig spermatozoa was assessed under non-capacitive and capacitive conditions, with or without specific NOX inhibitor, VAS2879 (40 µM), a pan-NOX inhibitor [
27]; ML171 (2 µM) or NSC23766, a small GTPase Rac1 inhibitor (100 µM). We also tested the effect of calpain inhibition on ROS production using calpeptin (10 µM), a calpain-specific inhibitor.
4.5. Assessment of NADPH Consumption
NADPH consumption was assessed following the method described by Andrade-Pavon et al. [
36]. Whole native protein extracts obtained from spermatozoa under non-capacitation and capacitation conditions were tested with or without specific inhibitors of NOXs in the presence of NADPH (18 nM). The decrease in absorbance at 340 nm was measured for 10 min, and the difference between the initial and final absorbance was used to calculate the enzymatic activity (µmol/min/mg–protein)
. 4.6. Assessment of Oxidative Stress
Intracellular peroxide assay. Intracellular ROS levels were determined using the oxidation-sensitive fluorescent probe 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA) to analyze the intracellular content of peroxides (H
2O
2) following a procedure modified from Guthrie and Welch [
37]. Spermatozoa (3.5 × 10
7 cell/mL) were incubated under conditions of capacitation in the presence of H2DCF-DA (50 µM final concentration), at 37 °C for 60 min in the dark. ROS production was evaluated (Ex/Em, 490/525) using an LS55 Luminescence Spectrometer (Perkin Elmer, Waltham Mass. USA). Detection of extracellular H
2O
2 by Amplex UltraRed (AUR) assay in the presence of HRP; spermatozoa (3.5 × 10
7 cell/mL) were incubated under conditions of capacitation with or without specific inhibitor in the presence of AUR (50 µM final concentration) and HRP (0.1 mM final concentration), at 37 °C for 60 min in the dark. Samples were centrifuged (5000×
g) for 3 min and supernatants were placed in a 96-well plate. Absorbance was read on a Microplate Reader at 540 nm (Bio rad, microplate reader, model 550).
Superoxide anion assay: Nitrotetrazolium Blue (NBT, 0.6 mM) was used to determine O
2•- [
38]. ROS production assay was performed at 3.5 × 10
7 cell/mL during capacitation (60 min). The NBT deposited inside the cells was then dissolved by adding KOH (2 M) to solubilize cell membranes and then adding DMSO to dissolve Formazan complex by gentle shaking for 10 min at room temperature. Absorbance was measured at 620 nm. Relative production was obtained when sperm were incubated in PBS, relative solution was then transferred to a 96-well plate and absorbance was read on a Multi-Function Microplate Reader at 620 nm (Power Wave X 340, Bio-Tek Instruments, Winooski, VT, USA).
4.7. Effect of VAS2870, NSC23766, and DPI on Sperm Viability
Sperm viability was determined following the method described by Roa-Espitia et al. [
39]. Sperm suspensions were incubated in MCM-PL at pH 7.8 and 37 °C for 60 min in the absence and presence of VAS2870 (40 µM) or NSC23766 (100 µM). Once the incubation was finished, we added a PI solution (1 μg/mL) to a sample of spermatozoa to a 1:1 ratio and mixed and incubated the mixture at room temperature for 30 min. The spermatozoa were washed and the number of stained and unstained spermatozoa was counted (500 cell X sample,
n = 3) under an epifluorescence microscope (Olympus BX500, Tokyo, Japan). Images were registered and analyzed using the software Nikon Elements 3.1.
4.8. Western Blotting
Vas deferens sperm were obtained, washed in 154 mM NaCl solution, and capacitated. Samples were centrifuged (5000×
g) for 3 min and suspended in RIPA buffer (25 mM TRIS HCl pH 7.6, 150 mM NaCl, 1% NP-40, 1%, sodium deoxycholate, 0.1% SDS), supplemented with protease inhibitors (5 mg/mL soybean trypsin inhibitor, 100 mg/mL benzamidine, 30 mg/mL pepstatin A, 30 mg/mL leupeptin, 30 mg/mL aprotinin, 1 mM PMSF diluted in dimethyl sulfoxide, 20 mg/mL iodoacetamide, 1 mM sodium orthovanadate, 10 mM sodium fluoride, 10% glycerol, and 2.5% complete, Mini, EDTA-free Protease Inhibitor Cocktail [1 tablet diluted in 1 mL H2O]). The samples were then incubated for 20 min on ice and centrifuged (20,000×
g) for 20 min at 4 °C. The supernatants were collected, and their protein concentration was determined [
40]. To reduce protein disulfide bonds, supernatant aliquots were boiled for 5 min in 3X Laemmli buffer (720 mM TRIS-base, 6% SDS (
w/
v), 30% glycerol, 2-mercaptoethanol, and 0.03% bromophenol blue (
w/
v), pH 10). Then, proteins were separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on 10% polyacrylamide non-gradient gels, and subsequently transferred to nitrocellulose membranes and blocked in 5% skimmed milk in 1 × PBS/1% Triton X-100 (pH 7.4).
Phospho-tyrosine detection. First, 500 µg of total spermatic protein extract was run on SDS-PAGE gels and transferred to nitrocellulose membranes overnight at 4 °C. Membranes were washed once with PBS-Tween 20 (0.1%) and blocked with 5% nonfat dry milk in PBS-Tween 20 (0.1%) for two hours at room temperature with constant agitation. Membranes were then incubated with a 1:500 dilution of anti-phosphotyrosine (Cell Signaling, APY03, Danvers, MA, USA) antibody diluted in PBS-Tween 20 (0.1%) and incubated overnight at 4 °C with constant agitation followed by washing three times in PBS-Tween 20 (0.1% final concentration). The secondary mouse antibody HRP-conjugated IgG (Jackson Labs., 115-035-068) was diluted 1:10,000 in PBS added with nonfat milk (3% final concentration) and incubated with membranes for one hour at room temperature. Membranes were then washed three times in PBS for 10 min each.
Nox4 and Nox2 detection. Using SDS-PAGE, 300 µg of spermatic protein extract was loaded, and the proteins were separated and transferred to nitrocellulose membranes. Western blotting was performed under the same conditions used for phospho-tyrosine detection. The antibodies used were anti-NOX2/gp91phox antibody (EPR6991) (Abcam, ab129068) for NOX2 detection and anti-NADPH oxidase 4 antibody (UOTR1B492) (Abcam, ab109225, Cambridge, UK) for NOX4 detection.
In both assays, proteins were detected by chemiluminescence, which was developed using Amersham Biosciences ECL Prime Western Blotting Detection Reagent and visualized in an Odyssey
® Fc Imaging System by LI-COR Biosciences. The densitometric analysis was performed using the ImageJ software (
https://imagej.nih.gov (accessed on 3 April 2021)).
4.9. Chlortetracycline Fluorescence Assay
This procedure was previously described [
31] and modified for guinea pig spermatozoa [
39,
41] as follows: The stain solution was prepared by dissolving 250-μM chlortetracycline (CTC)-HCl in TN buffer (20 mM Tris, 130 mM NaCl, and 5 mM cysteine at pH 7.8); fresh CTC stock was prepared daily. At the time of the assay, 20 μL of non-capacitated or capacitated spermatozoa, treated or not with VAS2879, were mixed with 20 μL of pre-warmed CTC stock solution and incubated for 20 s in a water bath at 37 °C. Immediately after incubation, the CTC-sperm suspension was fixed adding 3.5 µL of 12.5% glutaraldehyde in 1.25 M Tris (pH 7.5), immediately followed by gentle mixing. Fixed samples were kept in a dark box. After 1–4 h of fixation, slides were prepared and examined under fluorescence microscope (Ex330-380/Em420 nm). All fluorescence images were obtained using an Olympus BX500 fluorescence microscope. In each sample, 100 spermatozoa were classified as expressing one of three CTC staining patterns: F pattern, a faint fluorescence in the acrosome region, which is characteristic of non-capacitated acrosome-intact cells; B pattern, a bright fluorescence in the acrosomal region with a band along the equatorial segment, which is typical of capacitated, acrosome-intact cells; AR pattern, a fluorescence in the equatorial segment and/or post-acrosomal region, which is characteristic of physiologically capacitated acrosome-reacted cells. The presence or absence of the acrosomal cap on each cell was verified using phase-contrast illumination.
4.10. Spermatic Motility Analysis
Sperm motility was evaluated following the methodology described by Cordero-Martínez [
41]. Guinea pig spermatozoa were capacitated for 60 min in the absence or presence of VAS2870 (40 µM). During incubation, aliquots were used for sperm motility quantitative parameter screening on a CASA instrument (TOX IVOS, software version 12.3: Hamilton Thorne Bioscience, Beverly, MA, USA) in a sample counting chamber prewarmed to 37 °C (MicroCell 20 Micron). Sperm motility was recorded at 60 frames/s for one second; velocity distribution and other kinematic parameters of motility (VAP, VSL, VCL, ALH, and BCF) were analyzed. Experiments were performed in triplicate.
4.11. Intracellular Calcium Evaluation
Spermatozoa were capacitated for 15, 30, 60 and 90 min in the absence or presence of VAS2870 (40 µM), and then spermatozoa (aliquots of 3.5 × 107 cell/mL) were loaded with the Ca2+ indicator dye Fluo-3AM (Fluo-3-pentaacetoxymethyl ester, Invitrogen, Carlsbad, CA, USA) resuspended in dimethyl sulfoxide (DMSO, 1 mM). Assays were prepared at a final concentration of 2 μM Fluo-3AM for 15 min at 37 °C in MCM-PL. The cells were washed by centrifugation at 5000× g with MCM-PL. After treatment, changes in intracellular calcium concentration ([Ca2+]i) were recorded as background substrate ratios of the corresponding excitation wavelength (Ex490/Em520 nm). Intracellular calcium evaluation was performed for 200 s on a continuous basis, with or without pharmacological inhibitors. Fluorescence was registered in an LS-55 Fluorescence Spectrometer (Perkin Elmer, Waltham MA, USA).
4.12. Immunofluorescence Assays
Non-capacitated or capacitated spermatozoa, treated or not with calpeptin, were fixed in formaldehyde (1.5% final concentration) in PBS. After one hour, the sperm were collected by centrifugation. The pelleted sperm (600 g for 3 min) were incubated in 50 mM NH4Cl for 10 min, rinsed twice with PBS, and then with bidistilled water. Microscope slides were prepared using this suspension, air-dried at room temperature overnight and stored at 4 °C. Sperm cells were permeabilized in acetone for 7 min at −20°C and washed with PBS. NOX2 or NOX4 (1:100) antibody was diluted in PBS with 1% BSA (blocking solution) added and incubated overnight at room temperature. The slides were washed with PBS and incubated for two hours at 37 °C with the appropriate TRITC-conjugated secondary antibody diluted in blocking solution. The samples were mounted on glass-covered slides using Gelvatol, sealed properly, and stored at −20 °C until observations were performed. The stained cells were imaged under a confocal laser scanning microscope (Leica TCS SP8, Wetzlar, Germany) and analyzed using LAS AF Lite’s imaging software (Version 2.6.3).
4.13. Co-Immunoprecipitation Assay
Co-immunoprecipitation experiments were performed using the Crosslink Immunoprecipitation Kit (Thermo Fisher Scientific, Waltham MA, USA, 26147). This method involves capturing 20 μg antibody on Protein A/G beaded agarose resin and covalently immobilizing it on the support by crosslinking with 2.5 mM disuccinimidyl suberate (DSS). The antibody resin was then incubated at 4 °C for 12 h with 500 μg precleared guinea pig spermatozoa protein extracts, allowing for the antibody-antigen complex to form. Proteins bound to the respective antibodies were eluted, recovered by low-speed centrifugation (3000× g), and stored at −20 °C. Only the antigen was eluted during the procedure, enabling it to be identified and analyzed with minimal interference from antibody fragments. For protein disulfide reduction, supernatant aliquots were boiled for 5 min in 3X Laemmli buffer (pH 10) containing 2-mercaptoethanol, and the proteins were then separated by SDS-PAGE. Next, proteins were transferred to nitrocellulose membranes for immunodetection. The recovered proteins were analyzed by WB using the adequate antibodies.
Negative control was performed by associating 20 µg of an IgG unrelated to NOX2 or NOX4 with the agarose-protein A/G beads. The beads were incubated with 500 µg of sperm protein extract in the same way as before. The WB carried out with the anti-NOX2 antibody did not show the presence of this protein, nor of another (
Supplemental Figure S1).
4.14. Statistical Analysis
SigmaPlot 11 was employed to perform statistical analysis. Data are expressed as mean ± S.E. Means were compared using ANOVA one way as appropriate. Statistical significance between the samples was considered when p ≤ 0.05.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






