


The Combination of Immune Checkpoint Blockade with Tumor Vessel Normalization as a Promising Therapeutic Strategy for Breast Cancer: An Overview of Preclinical and Clinical Studies
 ,
,  ,
,  ,
,  ,
,  and
and
Abstract
1. Introduction
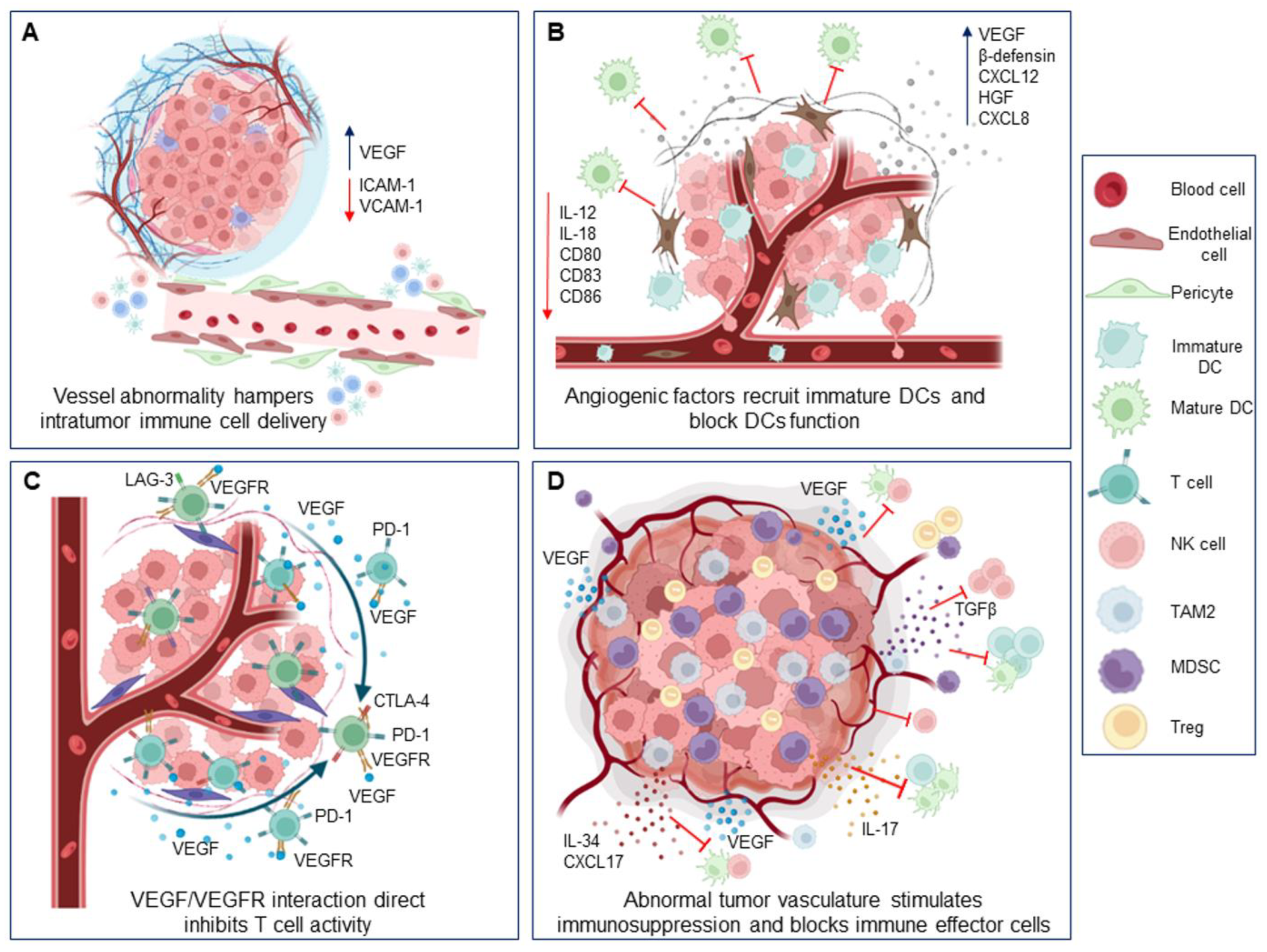
2. The Abnormal Tumor Vasculature of Breast Cancer
The Molecular Players of Aberrant Vasculature in Breast Cancer
3. Impact of Abnormal Breast Tumor Vascular on Immune Cells
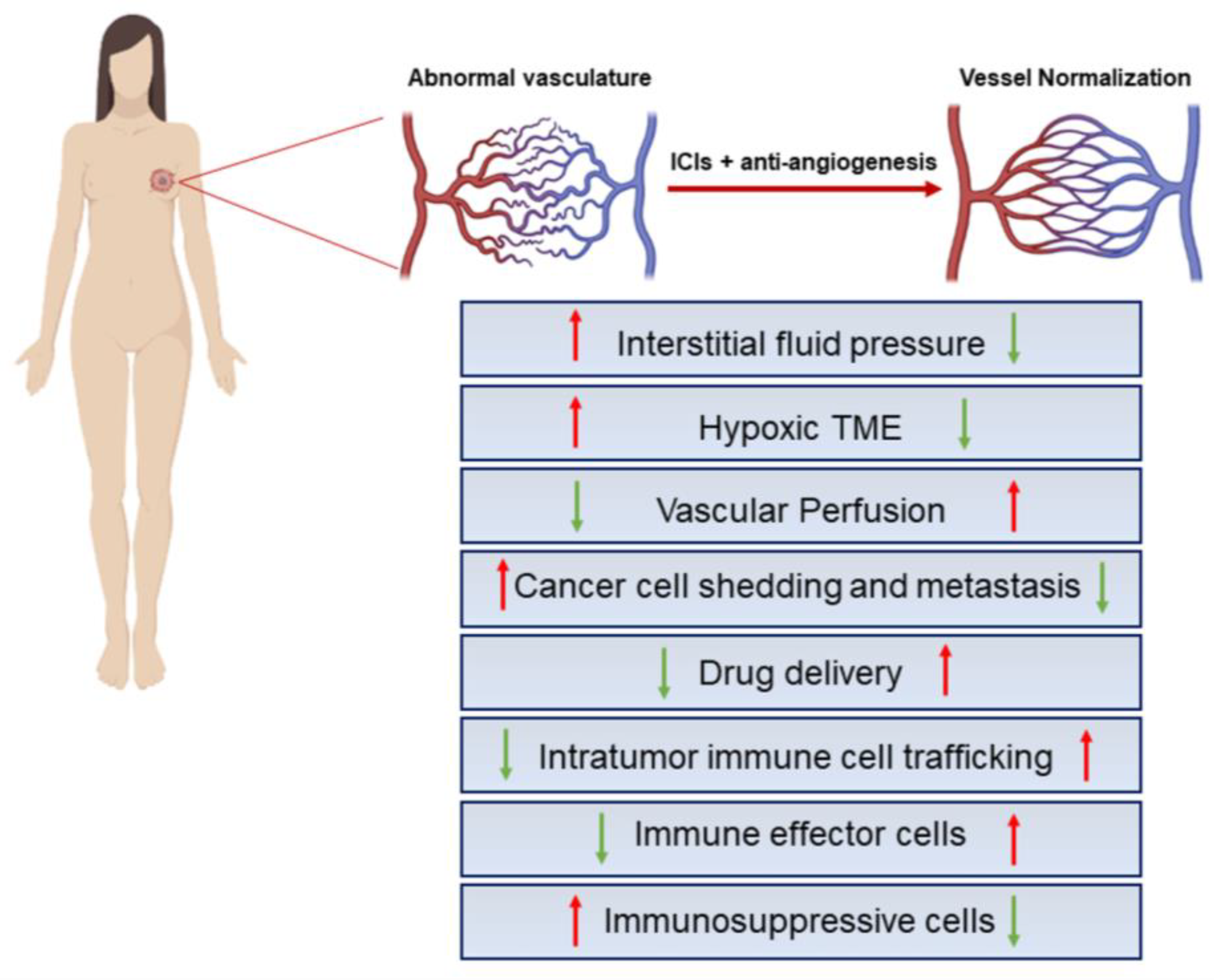
4. Vessel Normalization Strategies in Breast Cancer
Vessel Normalization Improves Immunotherapy and Vice Versa: Preclinical Evidence in Breast Cancer
5. Effect of Antiangiogenic Agents Combined with Immune Checkpoint Inhibitors in Breast Cancer: Clinical Studies
6. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| BC | breast cancer |
| CTL | cytotoxic T lymphocytes |
| CTLA-4 | cytotoxic T-lymphocyte-associated protein |
| DC | dendritic cell |
| FGF | fibroblast growth factor |
| HER2 | human epidermal growth factor receptor 2 |
| HIF | hypoxia-inducible transcription factor |
| IC | immune checkpoint |
| ICIs | immune checkpoint inhibitors |
| IL-8 | interleukin 8 |
| Lag3 | lymphocyte activation gene 3 |
| LDHA | lactate dehydrogenase |
| MDSCs | myeloid-derived suppressor cells |
| MMPs | matrix metalloproteinases |
| pCR | pathologic complete response |
| PDGF | platelet-derived growth factor |
| PD-L1 | programmed death-ligand 1 |
| TAM | M2-like protumor macrophages |
| TGFβ-1 | transforming growth factor beta-1 |
| TILs | tumor-infiltrating lymphocytes |
| Tim-3 | T-cell immunoglobulin mucin-3 |
| TME | tumor microenvironment |
| TNBCs | triple-negative BCs |
| Tregs | regulatory T cells |
| VEGF | vascular endothelial growth factor |
References
- Mellman, I.; Coukos, G.; Dranoff, G. Cancer immunotherapy comes of age. Nature 2011, 480, 480–489. [Google Scholar] [CrossRef]
- Topalian, S.L.; Taube, J.M.; Anders, R.A.; Pardoll, D.M. Mechanism-driven biomarkers to guide immune checkpoint blockade in cancer therapy. Nat. Rev. Cancer 2016, 16, 275–287. [Google Scholar] [CrossRef]
- Melaiu, O.; Lucarini, V.; Giovannoni, R.; Fruci, D.; Gemignani, F. News on immune checkpoint inhibitors as immunotherapy strategies in adult and pediatric solid tumors. Semin. Cancer Biol. 2020, 79, 18–43. [Google Scholar] [CrossRef] [PubMed]
- Mezni, E.; Behi, K.; Gonçalves, A. Immunotherapy and breast cancer: An overview. Curr. Opin. Oncol. 2022, 34, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Zhang, H.; Zheng, J.; Zhang, L. Turning Cold into Hot: Firing up the Tumor Microenvironment. Trends Cancer 2020, 6, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Howard, F.M.; Villamar, D.; He, G.; Pearson, A.T.; Nanda, R. The emerging role of immune checkpoint inhibitors for the treatment of breast cancer. Expert Opin. Investig. Drugs 2021, 31, 531–548. [Google Scholar] [CrossRef]
- Mph, K.D.M.; Fidler-Benaoudia, M.; Keegan, T.H.; Hipp, H.S.; Jemal, A.; Siegel, R.L.; Keegan, M.T.H.; Dvm, A.J.; Mph, R.L.S. Cancer statistics for adolescents and young adults, 2020. CA Cancer J. Clin. 2020, 70, 443–459. [Google Scholar] [CrossRef]
- Alexandrov, L.B.; Nik-Zainal, S.; Wedge, D.C.; Aparicio, S.A.J.R.; Behjati, S.; Biankin, A.V.; Bignell, G.R.; Bolli, N.; Borg, A.; Børresen-Dale, A.-L.; et al. Signatures of mutational processes in human cancer. Nature 2013, 500, 415–421. [Google Scholar] [CrossRef] [PubMed]
- The Cancer Genome Atlas (TCGA) Research Network. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef]
- Polyak, K. Breast cancer: Origins and evolution. J. Clin. Investig. 2007, 117, 3155–3163. [Google Scholar] [CrossRef]
- Dai, X.-L.; Han, Z.-B.; Yang, Y.-T.; Qiu, J.; Liu, Y.-F.; Feng, Y.-Z. Efficacy and prognosis of neoadjuvant chemotherapy is correlated with breast cancer molecular classification. Int. J. Clin. Pharmacol. Ther. 2015, 53, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Loi, S.; Sirtaine, N.; Piette, F.; Salgado, R.; Viale, G.; Van Eenoo, F.; Rouas, G.; Francis, P.; Crown, J.P.; Hitre, E.; et al. Prognostic and Predictive Value of Tumor-Infiltrating Lymphocytes in a Phase III Randomized Adjuvant Breast Cancer Trial in Node-Positive Breast Cancer Comparing the Addition of Docetaxel to Doxorubicin With Doxorubicin-Based Chemotherapy: BIG 02-98. J. Clin. Oncol. 2013, 31, 860–867. [Google Scholar] [CrossRef]
- Mittendorf, E.A.; Philips, A.V.; Meric-Bernstam, F.; Qiao, N.; Wu, Y.; Harrington, S.; Su, X.; Wang, Y.; Gonzalez-Angulo, A.M.; Akcakanat, A.; et al. PD-L1 Expression in Triple-Negative Breast Cancer. Cancer Immunol. Res. 2014, 2, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Luen, S.; Virassamy, B.; Savas, P.; Salgado, R.; Loi, S. The genomic landscape of breast cancer and its interaction with host immunity. Breast 2016, 29, 241–250. [Google Scholar] [CrossRef]
- Atezolizumab Combo Approved for PD-L1–Positive TNBC. Cancer Discov. 2019, 9, OF2. [CrossRef] [PubMed]
- Cortes, J.; Rugo, H.S.; Cescon, D.W.; Im, S.-A.; Yusof, M.M.; Gallardo, C.; Lipatov, O.; Barrios, C.H.; Perez-Garcia, J.; Iwata, H.; et al. Pembrolizumab plus Chemotherapy in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2022, 387, 217–226. [Google Scholar] [CrossRef]
- Ge, Y.; Domschke, C.; Stoiber, N.; Schott, S.; Heil, J.; Rom, J.; Blumenstein, M.; Thum, J.; Sohn, C.; Schneeweiss, A.; et al. Metronomic cyclophosphamide treatment in metastasized breast cancer patients: Immunological effects and clinical outcome. Cancer Immunol. Immunother. 2011, 61, 353–362. [Google Scholar] [CrossRef]
- Garufi, G.; Palazzo, A.; Paris, I.; Orlandi, A.; Cassano, A.; Tortora, G.; Scambia, G.; Bria, E.; Carbognin, L. Neoadjuvant therapy for triple-negative breast cancer: Potential predictive biomarkers of activity and efficacy of platinum chemotherapy, PARP- and immune-checkpoint-inhibitors. Expert Opin. Pharmacother. 2020, 21, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Nishino, M.; Ramaiya, N.H.; Hatabu, H.; Hodi, F.S. Monitoring immune-checkpoint blockade: Response evaluation and biomarker development. Nat. Rev. Clin. Oncol. 2017, 14, 655–668. [Google Scholar] [CrossRef]
- Ben-Baruch, A. Host microenvironment in breast cancer development: Inflammatory cells, cytokines and chemokines in breast cancer progression: Reciprocal tumor–microenvironment interactions. Breast Cancer Res. 2002, 5, 31–36. [Google Scholar] [CrossRef]
- Jain, R.K. Normalizing Tumor Microenvironment to Treat Cancer: Bench to Bedside to Biomarkers. J. Clin. Oncol. 2013, 31, 2205–2218. [Google Scholar] [CrossRef]
- De Heer, E.C.; Jalving, M.; Harris, A.L. HIFs, angiogenesis, and metabolism: Elusive enemies in breast cancer. J. Clin. Investig. 2020, 130, 5074–5087. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhang, J.; Huang, Z.; Zuo, T.; Lu, Q.; Wu, G.; Shen, Q. Reducing Interstitial Fluid Pressure and Inhibiting Pulmonary Metastasis of Breast Cancer by Gelatin Modified Cationic Lipid Nanoparticles. ACS Appl. Mater. Interfaces 2017, 9, 29457–29468. [Google Scholar] [CrossRef]
- Palucka, A.K.; Coussens, L.M. The basis of oncoimmunology. Cell 2016, 164, 1233–1247. [Google Scholar] [CrossRef]
- Priya, S.K.; Nagare, R.; Sneha, V.; Sidhanth, C.; Bindhya, S.; Manasa, P.; Ganesan, T. Tumour angiogenesis-Origin of blood vessels. Int. J. Cancer 2016, 139, 729–735. [Google Scholar] [CrossRef]
- Hanahan, D.; Folkman, J. Patterns and Emerging Mechanisms of the Angiogenic Switch during Tumorigenesis. Cell 1996, 86, 353–364. [Google Scholar] [CrossRef]
- Majidpoor, J.; Mortezaee, K. Angiogenesis as a hallmark of solid tumors-clinical perspectives. Cell. Oncol. 2021, 44, 715–737. [Google Scholar] [CrossRef] [PubMed]
- Moschetta, M.; Mishima, Y.; Sahin, I.; Manier, S.; Glavey, S.; Vacca, A.; Roccaro, A.M.; Ghobrial, I.M. Role of endothelial progenitor cells in cancer progression. Biochim. et Biophys. Acta (BBA)-Rev. Cancer 2014, 1846, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Pezzella, F. Vascular Co-Option and Other Alternative Modalities of Growth of Tumor Vasculature in Glioblastoma. Front. Oncol. 2022, 12, 874554. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef]
- Badodekar, N.; Sharma, A.; Patil, V.; Telang, G.; Sharma, R.; Patil, S.; Vyas, N.; Somasundaram, I. Angiogenesis induction in breast cancer: A paracrine paradigm. Cell Biochem. Funct. 2021, 39, 860–873. [Google Scholar] [CrossRef]
- Linderholm, B.; Tavelin, B.; Grankvist, K.; Henriksson, R. Does vascular endothelial growth factor (VEGF) predict local relapse and survival in radiotherapy-treated node-negative breast cancer? Br. J. Cancer 1999, 81, 727–732. [Google Scholar] [CrossRef]
- Weidner, N.; Semple, J.P.; Welch, W.R.; Folkman, J. Tumor angiogenesis and metastasis—Correlation in invasive breast carcinoma. N. Engl. J. Med. 1991, 324, 1–8. [Google Scholar] [CrossRef]
- Vakoc, B.J.; Lanning, R.M.; Tyrrell, J.A.; Padera, T.; Bartlett, L.A.; Stylianopoulos, T.; Munn, L.L.; Tearney, G.J.; Fukumura, D.; Jain, R.K.; et al. Three-dimensional microscopy of the tumor microenvironment in vivo using optical frequency domain imaging. Nat. Med. 2009, 15, 1219–1223. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.K. Antiangiogenesis Strategies Revisited: From Starving Tumors to Alleviating Hypoxia. Cancer Cell 2014, 26, 605–622. [Google Scholar] [CrossRef]
- Cho, H.-H.; Kim, H.; Nam, S.Y.; Lee, J.E.; Han, B.-K.; Ko, E.Y.; Choi, J.S.; Park, H.; Ko, E.S. Measurement of Perfusion Heterogeneity within Tumor Habitats on Magnetic Resonance Imaging and Its Association with Prognosis in Breast Cancer Patients. Cancers 2022, 14, 1858. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, K.; Holm, C.; Landberg, G. Common Molecular Mechanisms of Mammary Gland Development and Breast Cancer. Cell. Mol. Life Sci. 2007, 64, 3233–3247. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Wang, M.; Schmid, T.; Xin, Z.; Kozhuharova, L.; Yu, W.-K.; Huang, Y.; Cai, F.; Biskup, E. Hypoxia in Breast Cancer—Scientific Translation to Therapeutic and Diagnostic Clinical Applications. Front. Oncol. 2021, 11, 652266. [Google Scholar] [CrossRef] [PubMed]
- Forster, J.C.; Harriss-Phillips, W.M.; Douglass, M.; Bezak, E. A review of the development of tumor vasculature and its effects on the tumor microenvironment. Hypoxia 2017, 5, 21–32. [Google Scholar] [CrossRef]
- Potente, M.; Mäkinen, T. Vascular heterogeneity and specialization in development and disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 477–494. [Google Scholar] [CrossRef]
- Herbert, S.P.; Stainier, D.Y.R. Molecular control of endothelial cell behaviour during blood vessel morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Luk, C.K.; Veinot-Drebot, L.; Tjan, E.; Tannock, I.F. Effect of Transient Hypoxia on Sensitivity to Doxorubicin in Human and Murine Cell Lines. Gynecol. Oncol. 1990, 82, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, D.; Kloepper, J.; Amoozgar, Z.; Duda, D.G.; Jain, R.K. Enhancing cancer immunotherapy using antiangiogenics: Opportunities and challenges. Nat. Rev. Clin. Oncol. 2018, 15, 325–340. [Google Scholar] [CrossRef]
- Chen, W.; Hoffmann, A.D.; Liu, H.; Liu, X. Organotropism: New insights into molecular mechanisms of breast cancer metastasis. npj Precis. Oncol. 2018, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kozłowski, J.; Kozłowska, A.; Kocki, J. Breast cancer metastasis–insight into selected molecular mechanisms of the phenomenon. Postep. Hig. I Med. Dosw. 2015, 69, 447–451. [Google Scholar] [CrossRef]
- Ribatti, D. Immunosuppressive effects of vascular endothelial growth factor (Review). Oncol. Lett. 2022, 24, 1–6. [Google Scholar] [CrossRef]
- Oshi, M.; Newman, S.; Tokumaru, Y.; Yan, L.; Matsuyama, R.; Endo, I.; Nagahashi, M.; Takabe, K. Intra-Tumoral Angiogenesis Is Associated with Inflammation, Immune Reaction and Metastatic Recurrence in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 6708. [Google Scholar] [CrossRef]
- Martins, S.F.; Garcia, E.A.; Luz, M.A.M.; Pardal, F.; Rodrigues, M.; Filho, A.L. Clinicopathological correlation and prognostic significance of VEGF-A, VEGF-C, VEGFR-2 and VEGFR-3 expression in colorectal cancer. Cancer Genom. Proteom. 2013, 10, 55–67. [Google Scholar]
- Cañadas, I.; Taus, Á.; Villanueva, X.; Arpí, O.; Pijuan, L.; Rodríguez, Y.; Menéndez, S.; Mojal, S.; Rojo, F.; Albanell, J.; et al. Angiopoietin-2 is a negative prognostic marker in small cell lung cancer. Lung Cancer 2015, 90, 302–306. [Google Scholar] [CrossRef]
- Melaiu, O.; Catalano, C.; De Santi, C.; Cipollini, M.; Figlioli, G.; Pellè, L.; Barone, E.; Evangelista, M.; Guazzelli, A.; Boldrini, L.; et al. Inhibition of the platelet-derived growth factor receptor beta (PDGFRB) using gene silencing, crenolanib besylate, or imatinib mesylate hampers the malignant phenotype of mesothelioma cell lines. Genes Cancer 2017, 8, 438–452. [Google Scholar] [CrossRef]
- Kern, F.G.; Lippman, M.E. The role of angiogenic growth factors in breast cancer progression. Cancer Metastasis Rev. 1996, 15, 213–219. [Google Scholar] [CrossRef]
- Niu, G. Vascular Endothelial Growth Factor as an Anti-Angiogenic Target for Cancer Therapy. Curr. Drug Targets 2010, 11, 1000–1017. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Nico, B.; Ruggieri, S.; Tamma, R.; Simone, G.; Mangia, A. Angiogenesis and Antiangiogenesis in Triple-Negative Breast cancer. Transl. Oncol. 2016, 9, 453–457. [Google Scholar] [CrossRef]
- Longatto-Filho, A.; Lopes, J.M.; Schmitt, F.C. Angiogenesis and Breast Cancer. J. Oncol. 2010, 2010, 1–7. [Google Scholar] [CrossRef]
- Eroğlu, A.; Ersöz, C.; Karasoy, D.; Sak, S.D. Vascular endothelial growth factor (VEGF)-C, VEGF-D, VEGFR-3 and D2-40 expressions in primary breast cancer: Association with lymph node metastasis. Adv. Clin. Exp. Med. 2017, 26, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Srabovic, N.; Mujagic, Z.; Mujanovic-Mustedanagic, J.; Softic, A.; Muminovic, Z.; Rifatbegovic, A.; Begic, L. Vascular Endothelial Growth Factor Receptor-1 Expression in Breast Cancer and Its Correlation to Vascular Endothelial Growth Factor A. Int. J. Breast Cancer 2013, 2013, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Colbert, L.S.; Fuller, M.; Zhang, Y.; Gonzalez-Perez, R.R. Vascular endothelial growth factor receptor-2 in breast cancer. Biochim. Biophys. Acta Rev. Cancer 2010, 1806, 108–121. [Google Scholar] [CrossRef]
- Schneider, B.P.; Miller, K.D. Angiogenesis of Breast Cancer. J. Clin. Oncol. 2005, 23, 1782–1790. [Google Scholar] [CrossRef] [PubMed]
- Sahana, K.R.; Akila, P.; Prashant, V.; Chandra, B.S.; Suma, M.N. Quantitation of Vascular Endothelial Growth Factor and Interleukin-6 in Different Stages of Breast Cancer. Rep. Biochem. Mol. Biol. 2017, 6, 33–39. [Google Scholar]
- Chelouche-Lev, D.; Miller, C.P.; Tellez, C.; Ruiz, M.; Bar-Eli, M.; Price, J.E. Different signalling pathways regulate VEGF and IL-8 expression in breast cancer: Implications for therapy. Eur. J. Cancer 2004, 40, 2509–2518. [Google Scholar] [CrossRef]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 Directly Enhanced Endothelial Cell Survival, Proliferation, and Matrix Metalloproteinases Production and Regulated Angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Razmkhah, M.; Jaberipour, M.; Hosseini, A.; Safaei, A.; Khalatbari, B.; Ghaderi, A. Expression profile of IL-8 and growth factors in breast cancer cells and adipose-derived stem cells (ASCs) isolated from breast carcinoma. Cell. Immunol. 2010, 265, 80–85. [Google Scholar] [CrossRef]
- Marjon, P.L.; Bobrovnikova-Marjon, E.V.; Abcouwer, S.F. Expression of the pro-angiogenic factors vascular endothelial growth factor and interleukin-8/CXCL8 by human breast carcinomas is responsive to nutrient deprivation and endoplasmic reticulum stress. Mol. Cancer 2004, 3, 4. [Google Scholar] [CrossRef]
- Lin, Y.; Huang, R.; Chen, L.; Li, S.; Shi, Q.; Jordan, C.; Huang, R.-P. Identification of interleukin-8 as estrogen receptor-regulated factor involved in breast cancer invasion and angiogenesis by protein arrays. Int. J. Cancer 2004, 109, 507–515. [Google Scholar] [CrossRef]
- Martin, D.; Galisteo, R.; Gutkind, J. CXCL8/IL8 Stimulates Vascular Endothelial Growth Factor (VEGF) Expression and the Autocrine Activation of VEGFR2 in Endothelial Cells by Activating NFκB through the CBM (Carma3/Bcl10/Malt1) Complex. J. Biol. Chem. 2009, 284, 6038–6042. [Google Scholar] [CrossRef]
- Gyanchandani, R.; Alves, M.V.O.; Myers, J.N.; Kim, S. A Proangiogenic Signature Is Revealed in FGF-Mediated Bevacizumab-Resistant Head and Neck Squamous Cell Carcinoma. Mol. Cancer Res. 2013, 11, 1585–1596. [Google Scholar] [CrossRef] [PubMed]
- Nasarre, P.; Thomas, M.; Kruse, K.; Helfrich, I.; Wolter, V.; Deppermann, C.; Schadendorf, D.; Thurston, G.; Fiedler, U.; Augustin, H.G. Host-Derived Angiopoietin-2 Affects Early Stages of Tumor Development and Vessel Maturation but Is Dispensable for Later Stages of Tumor Growth. Cancer Res. 2009, 69, 1324–1333. [Google Scholar] [CrossRef] [PubMed]
- Gerald, D.; Chintharlapalli, S.; Augustin, H.G.; Benjamin, L.E. Angiopoietin-2: An Attractive Target for Improved Antiangiogenic Tumor Therapy. Cancer Res. 2013, 73, 1649–1657. [Google Scholar] [CrossRef]
- Augustin, H.G.; Koh, G.Y.; Thurston, G.; Alitalo, K. Control of vascular morphogenesis and homeostasis through the angiopoietin–Tie system. Nat. Rev. Mol. Cell Biol. 2009, 10, 165–177. [Google Scholar] [CrossRef]
- Dhami, S.P.S.; Patmore, S.; Comerford, C.; Byrne, C.M.; Cavanagh, B.; Castle, J.; Kirwan, C.C.; Kenny, M.; Schoen, I.; O’Donnell, J.S.; et al. Breast cancer cells mediate endothelial cell activation, promoting von Willebrand factor release, tumor adhesion, and transendothelial migration. J. Thromb. Haemost. 2022, 20, 2350–2365. [Google Scholar] [CrossRef]
- Relf, M.; Lejeune, S.; Scott, P.A.; Fox, S.; Smith, K.; Leek, R.; Moghaddam, A.; Whitehouse, R.; Bicknell, R.; Harris, A.L. Expression of the angiogenic factors vascular endothelial cell growth factor, acidic and basic fibroblast growth factor, tumor growth factor beta-1, platelet-derived endothelial cell growth factor, placenta growth factor, and pleiotrophin in human primary breast cancer and its relation to angiogenesis. . Cancer Res. 1997, 57, 963–969. [Google Scholar]
- Luo, Q.; Wang, J.; Zhao, W.; Peng, Z.; Liu, X.; Li, B.; Zhang, H.; Shan, B.; Zhang, C.; Duan, C. Vasculogenic mimicry in carcinogenesis and clinical applications. J. Hematol. Oncol. 2020, 13, 1–15. [Google Scholar] [CrossRef]
- Andonegui-Elguera, M.A.; Alfaro-Mora, Y.; Cáceres-Gutiérrez, R.; Caro-Sánchez, C.H.S.; Herrera, L.A.; Díaz-Chávez, J. An Overview of Vasculogenic Mimicry in Breast Cancer. Front. Oncol. 2020, 10, 220. [Google Scholar] [CrossRef]
- Horton, B.L.; Fessenden, T.B.; Spranger, S. Tissue Site and the Cancer Immunity Cycle. Trends Cancer 2019, 5, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Huinen, Z.R.; Huijbers, E.J.M.; van Beijnum, J.R.; Nowak-Sliwinska, P.; Griffioen, A.W. Anti-angiogenic agents—overcoming tumour endothelial cell anergy and improving immunotherapy outcomes. Nat. Rev. Clin. Oncol. 2021, 18, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Mellman, I. Oncology Meets Immunology: The Cancer-Immunity Cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef]
- Hammerl, D.; Martens, J.W.M.; Timmermans, M.; Smid, M.; Trapman-Jansen, A.M.; Foekens, R.; Isaeva, O.I.; Voorwerk, L.; Balcioglu, H.E.; Wijers, R.; et al. Spatial immunophenotypes predict response to anti-PD1 treatment and capture distinct paths of T cell evasion in triple negative breast cancer. Nat. Commun. 2021, 12, 5668. [Google Scholar] [CrossRef]
- Dirkx, A.E.M.; Egbrink, M.G.A.O.; Kuijpers, M.J.E.; Van Der Niet, S.T.; Heijnen, V.V.T.; Steege, J.C.A.B.-T.; Wagstaff, J.; Griffioen, A.W. Tumor angiogenesis modulates leukocyte-vessel wall interactions in vivo by reducing endothelial adhesion molecule expression. Cancer Res. 2003, 63, 2322–2329. [Google Scholar] [PubMed]
- Steege, J.C.A.B.-T.; Baeten, C.I.M.; Thijssen, V.L.J.L.; Satijn, S.A.; Verhoeven, I.C.L.; Hillen, H.F.P.; Wagstaff, J.; Griffioen, A.W. Angiogenic Profile of Breast Carcinoma Determines Leukocyte Infiltration. Clin. Cancer Res. 2004, 10, 7171–7178. [Google Scholar] [CrossRef]
- Gabrilovich, D.; Ishida, T.; Oyama, T.; Ran, S.; Kravtsov, V.; Nadaf, S.; Carbone, D.P. Vascular Endothelial Growth Factor Inhibits the Development of Dendritic Cells and Dramatically Affects the Differentiation of Multiple Hematopoietic Lineages In Vivo. Blood 1998, 92, 4150–4166. [Google Scholar] [CrossRef]
- Voron, T.; Colussi, O.; Marcheteau, E.; Pernot, S.; Nizard, M.; Pointet, A.-L.; Latreche, S.; Bergaya, S.; Benhamouda, N.; Tanchot, C.; et al. VEGF-A modulates expression of inhibitory checkpoints on CD8+ T cells in tumors. J. Exp. Med. 2015, 212, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Gabrilovich, D.I.; Chen, H.L.; Girgis, K.R.; Cunningham, H.T.; Meny, G.M.; Nadaf, S.; Kavanaugh, D.; Carbone, D.P. Production of vascular endothelial growth factor by human tumors inhibits the functional maturation of dendritic cells. Nat. Med. 1996, 2, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Terme, M.; Pernot, S.; Marcheteau, E.; Sandoval, F.; Benhamouda, N.; Colussi, O.; Dubreuil, O.; Carpentier, A.F.; Tartour, E.; Taieb, J. VEGFA-VEGFR Pathway Blockade Inhibits Tumor-Induced Regulatory T-cell Proliferation in Colorectal Cancer. Cancer Res. 2013, 73, 539–549. [Google Scholar] [CrossRef] [PubMed]
- Ohm, J.E.; Gabrilovich, D.I.; Sempowski, G.D.; Kisseleva, E.; Parman, K.S.; Nadaf, S.; Carbone, D.P. VEGF inhibits T-cell development and may contribute to tumor-induced immune suppression. Blood 2003, 101, 4878–4886. [Google Scholar] [CrossRef]
- Lucarini, V.; Melaiu, O.; Tempora, P.; D’Amico, S.; Locatelli, F.; Fruci, D. Dendritic Cells: Behind the Scenes of T-Cell Infiltration into the Tumor Microenvironment. Cancers 2021, 13, 433. [Google Scholar] [CrossRef]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef]
- Asselin-Paturel, C.; Trinchieri, G. Production of type I interferons. J. Exp. Med. 2005, 202, 461–465. [Google Scholar] [CrossRef]
- Lee, W.S.; Yang, H.; Chon, H.J.; Kim, C. Combination of anti-angiogenic therapy and immune checkpoint blockade normalizes vascular-immune crosstalk to potentiate cancer immunity. Exp. Mol. Med. 2020, 52, 1475–1485. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, L.; Zhang, S.; Li, Y. Inhibition of vascular endothelial growth factor by small interfering RNA upregulates differentiation, maturation and function of dendritic cells. Exp. Ther. Med. 2014, 9, 120–124. [Google Scholar] [CrossRef]
- Roland, C.L.; Dineen, S.P.; Lynn, K.D.; Sullivan, L.A.; Dellinger, M.T.; Sadegh, L.; Sullivan, J.P.; Shames, D.S.; Brekken, R.A. Inhibition of vascular endothelial growth factor reduces angiogenesis and modulates immune cell infiltration of orthotopic breast cancer xenografts. Mol. Cancer Ther. 2009, 8, 1761–1771. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, X.; Dikov, M.M.; Novitskiy, S.V.; Mosse, C.A.; Yang, L.; Carbone, D.P. Distinct roles of VEGFR-1 and VEGFR-2 in the aberrant hematopoiesis associated with elevated levels of VEGF. Blood 2007, 110, 624–631. [Google Scholar] [CrossRef]
- Meder, L.; Schuldt, P.; Thelen, M.; Schmitt, A.; Dietlein, F.; Klein, S.; Borchmann, S.; Wennhold, K.; Vlasic, I.; Oberbeck, S.; et al. Combined VEGF and PD-L1 Blockade Displays Synergistic Treatment Effects in an Autochthonous Mouse Model of Small Cell Lung Cancer. Cancer Res. 2018, 78, 4270–4281. [Google Scholar] [CrossRef] [PubMed]
- Wherry, E.J. T cell exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Hirakata, T.; Kurozumi, S.; Tokuda, S.; Nakazawa, Y.; Obayashi, S.; Yajima, R.; Oyama, T.; Shirabe, K. VEGF-A Is Associated with the Degree of TILs and PD-L1 Expression in Primary Breast Cancer. Vivo 2020, 34, 2641–2646. [Google Scholar] [CrossRef]
- Palazon, A.; Tyrakis, P.A.; Macias, D.; Veliça, P.; Rundqvist, H.; Fitzpatrick, S.; Vojnovic, N.; Phan, A.T.; Loman, N.; Hedenfalk, I.; et al. An HIF-1α/VEGF-A Axis in Cytotoxic T Cells Regulates Tumor Progression. Cancer Cell 2017, 32, 669–683.e5. [Google Scholar] [CrossRef] [PubMed]
- DeNardo, D.G.; Brennan, D.J.; Rexhepaj, E.; Ruffell, B.; Shiao, S.L.; Madden, S.F.; Gallagher, W.M.; Wadhwani, N.; Keil, S.D.; Junaid, S.A.; et al. Leukocyte Complexity Predicts Breast Cancer Survival and Functionally Regulates Response to Chemotherapy. Cancer Discov. 2011, 1, 54–67. [Google Scholar] [CrossRef]
- Linderholm, B.K.; Lindh, B.; Beckman, L.; Erlanson, M.; Edin, K.; Tavelin, B.; Bergh, J.; Grankvist, K.; Henriksson, R. Prognostic Correlation of Basic Fibroblast Growth Factor and Vascular Endothelial Growth Factor in 1307 Primary Breast Cancers. Clin. Breast Cancer 2003, 4, 340–347. [Google Scholar] [CrossRef]
- Missiaen, R.; Mazzone, M.; Bergers, G. The reciprocal function and regulation of tumor vessels and immune cells offers new therapeutic opportunities in cancer. Semin. Cancer Biol. 2018, 52, 107–116. [Google Scholar] [CrossRef]
- Paluskievicz, C.M.; Cao, X.; Abdi, R.; Zheng, P.; Liu, Y.; Bromberg, J.S. T Regulatory cells and priming the suppressive tumor microenvironment. Front. Immunol. 2019, 10, 2453. [Google Scholar] [CrossRef]
- Munn, L.L.; Jain, R.K. Vascular regulation of antitumor immunity. Science 2019, 365, 544–545. [Google Scholar] [CrossRef]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.K.; Kadel, S.; Townsend, M.G.; Oliwa, M.; Guerriero, J.L. Macrophage Biology and Mechanisms of Immune Suppression in Breast Cancer. Front. Immunol. 2021, 12, 643771. [Google Scholar] [CrossRef] [PubMed]
- Ceci, C.; Atzori, M.G.; Lacal, P.M.; Graziani, G. Role of VEGFs/VEGFR-1 Signaling and Its Inhibition in Modulating Tumor Invasion: Experimental Evidence in Different Metastatic Cancer Models. Int. J. Mol. Sci. 2020, 21, 1388. [Google Scholar] [CrossRef]
- Dong, F.; Ruan, S.; Wang, J.; Xia, Y.; Le, K.; Xiao, X.; Hu, T.; Wang, Q. M2 macrophage-induced lncRNA PCAT6 facilitates tumorigenesis and angiogenesis of triple-negative breast cancer through modulation of VEGFR2. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Sun, X.; Bernhardt, S.M.; Glynn, D.J.; Hodson, L.J.; Woolford, L.; Evdokiou, A.; Yan, C.; Du, H.; Robertson, S.A.; Ingman, W.V. Attenuated TGFB signalling in macrophages decreases susceptibility to DMBA-induced mammary cancer in mice. Breast Cancer Res. 2021, 23, 1–16. [Google Scholar] [CrossRef]
- Jia, X.-H.; Feng, G.-W.; Wang, Z.-L.; Du, Y.; Shen, C.; Hui, H.; Peng, D.; Li, Z.-J.; Kong, D.-L.; Tian, J. Activation of mesenchymal stem cells by macrophages promotes tumor progression through immune suppressive effects. Oncotarget 2016, 7, 20934–20944. [Google Scholar] [CrossRef] [PubMed]
- Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. The Yin-Yang of tumor-associated macrophages in neoplastic progression and immune surveillance. Immunol. Rev. 2008, 222, 155–161. [Google Scholar] [CrossRef]
- Vroling, L.; Yuana, Y.; Schuurhuis, G.J.; van Hinsbergh, V.W.M.; Gundy, C.; de Haas, R.; van Cruijsen, H.; Boven, E.; Hoekman, K.; Broxterman, H.J. VEGFR2 expressing circulating (progenitor) cell populations in volunteers and cancer patients. Thromb. Haemost. 2007, 98, 440–450. [Google Scholar] [CrossRef] [PubMed]
- Kajal, K.; Bose, S.; Panda, A.K.; Chakraborty, D.; Chakraborty, S.; Pati, S.; Sarkar, T.; Dhar, S.; Roy, D.; Saha, S.; et al. Transcriptional regulation of VEGFA expression in T-regulatory cells from breast cancer patients. Cancer Immunol. Immunother. 2021, 70, 1877–1891. [Google Scholar] [CrossRef]
- Gupta, S.; Joshi, K.; Wig, J.; Arora, S.K. Intratumoral FOXP3 expression in infiltrating breast carcinoma: Its association with clinicopathologic parameters and angiogenesis. Acta Oncol. 2007, 46, 792–797. [Google Scholar] [CrossRef]
- Kajihara, N.; Kobayashi, T.; Otsuka, R.; Nio-Kobayashi, J.; Oshino, T.; Takahashi, M.; Imanishi, S.; Hashimoto, A.; Wada, H.; Seino, K.-I. Tumor-derived interleukin-34 creates an immunosuppressive and chemoresistant tumor microenvironment by modulating myeloid-derived suppressor cells in triple-negative breast cancer. Cancer Immunol. Immunother. 2022, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.-L.; Yen, M.-C.; Chang, W.-A.; Tsai, P.-H.; Pan, Y.-C.; Liao, S.-H.; Kuo, P.-L. CXCL17-derived CD11b+Gr-1+ myeloid-derived suppressor cells contribute to lung metastasis of breast cancer through platelet-derived growth factor-BB. Breast Cancer Res. 2019, 21, 1–13. [Google Scholar] [CrossRef]
- He, Q.; Jamalpour, M.; Bergquist, E.; Anderson, R.L.; Gustafsson, K.; Welsh, M. Mouse Breast Carcinoma Monocytic/Macrophagic Myeloid-Derived Suppressor Cell Infiltration as a Consequence of Endothelial Dysfunction in Shb-Deficient Endothelial Cells Increases Tumor Lung Metastasis. Int. J. Mol. Sci. 2021, 22, 11478. [Google Scholar] [CrossRef] [PubMed]
- Bergenfelz, C.; Roxå, A.; Mehmeti, M.; Leandersson, K.; Larsson, A.-M. Clinical relevance of systemic monocytic-MDSCs in patients with metastatic breast cancer. Cancer Immunol. Immunother. 2020, 69, 435–448. [Google Scholar] [CrossRef] [PubMed]
- Sceneay, J.; Parker, B.S.; Smyth, M.J.; Möller, A. Hypoxia-driven immunosuppression contributes to the pre-metastatic niche. Oncoimmunology 2013, 2, e22355. [Google Scholar] [CrossRef]
- Wicks, E.E.; Semenza, G.L. Hypoxia-inducible factors: Cancer progression and clinical translation. J. Clin. Investig. 2022, 132, e159839. [Google Scholar] [CrossRef] [PubMed]
- Giatromanolaki, A.; Gkegka, A.G.; Pouliliou, S.; Biziota, E.; Kakolyris, S.; Koukourakis, M. Hypoxia and anaerobic metabolism relate with immunologically cold breast cancer and poor prognosis. Breast Cancer Res. Treat. 2022, 194, 13–23. [Google Scholar] [CrossRef]
- Folkman, J. Tumor angiogenesis: Therapeutic implications. N. Engl. J. Med. 1971, 285, 1182–1186. [Google Scholar] [CrossRef]
- Comunanza, V.; Bussolino, F. Therapy for Cancer: Strategy of Combining Anti-Angiogenic and Target Therapies. Front. Cell Dev. Biol. 2017, 5, 101. [Google Scholar] [CrossRef]
- Jain, R.K. Normalization of Tumor Vasculature: An Emerging Concept in Antiangiogenic Therapy. Science 2005, 307, 58–62. [Google Scholar] [CrossRef]
- Duda, D.G.; Jain, R.K.; Willett, C.G. Antiangiogenics: The Potential Role of Integrating This Novel Treatment Modality with Chemoradiation for Solid Cancers. J. Clin. Oncol. 2007, 25, 4033–4042. [Google Scholar] [CrossRef] [PubMed]
- Viallard, C.; Audiger, C.; Popovic, N.; Akla, N.; Lanthier, K.; Legault-Navarrete, I.; Melichar, H.; Costantino, S.; Lesage, S.; Larrivée, B. BMP9 signaling promotes the normalization of tumor blood vessels. Oncogene 2020, 39, 2996–3014. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Fang, Z.; Liu, X.; Deng, S.; Zhou, P.; Wang, X.; Zhang, C.; Yin, R.; Hu, H.; Chen, X.; et al. Increased vessel perfusion predicts the efficacy of immune checkpoint blockade. J. Clin. Investig. 2018, 128, 2104–2115. [Google Scholar] [CrossRef] [PubMed]
- Carretero, R.; Sektioglu, I.M.; Garbi, N.; Salgado, O.C.; Beckhove, P.; Hämmerling, G.J. Eosinophils orchestrate cancer rejection by normalizing tumor vessels and enhancing infiltration of CD8+ T cells. Nat. Immunol. 2015, 16, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Yin, R.; Zhou, P.; Liu, X.; Fan, P.; Qian, L.; Dong, L.; Zhang, C.; Zheng, X.; Deng, S.; et al. DLL1 orchestrates CD8+T cells to induce long-term vascular normalization and tumor regression. Proc. Natl. Acad. Sci. USA 2021, 118, e2020057118. [Google Scholar] [CrossRef]
- Huang, Y.; Yuan, J.; Righi, E.; Kamoun, W.S.; Ancukiewicz, M.; Nezivar, J.; Santosuosso, M.; Martin, J.D.; Martin, M.R.; Vianello, F.; et al. Vascular normalizing doses of antiangiogenic treatment reprogram the immunosuppressive tumor microenvironment and enhance immunotherapy. Proc. Natl. Acad. Sci. USA 2012, 109, 17561–17566. [Google Scholar] [CrossRef]
- Secondini, C.; Coquoz, O.; Spagnuolo, L.; Spinetti, T.; Peyvandi, S.; Ciarloni, L.; Botta, F.; Bourquin, C.; Rüegg, C. Arginase inhibition suppresses lung metastasis in the 4T1 breast cancer model independently of the immunomodulatory and anti-metastatic effects of VEGFR-2 blockade. Oncoimmunology 2017, 6, e1316437. [Google Scholar] [CrossRef]
- Zhang, H.; Xia, W.; Liang, C.; Wang, X.; Zhi, L.; Guo, C.; Niu, Z.; Zhu, W. VEGF165b and its mutant demonstrate immunomodulatory, not merely anti-angiogenic functions, in tumor-bearing mice. Mol. Immunol. 2020, 122, 132–140. [Google Scholar] [CrossRef]
- Manning, E.A.; Ullman, J.G.; Leatherman, J.M.; Asquith, J.M.; Hansen, T.R.; Armstrong, T.D.; Hicklin, D.J.; Jaffee, E.M.; Emens, L.A. A Vascular Endothelial Growth Factor Receptor-2 Inhibitor Enhances Antitumor Immunity through an Immune-Based Mechanism. Clin. Cancer Res. 2007, 13, 3951–3959. [Google Scholar] [CrossRef]
- Huang, X.; Wong, M.K.; Yi, H.; Watkins, S.; Laird, A.D.; Wolf, S.F.; Gorelik, E. Combined therapy of local and metastatic 4T1 breast tumor in mice using SU6668, an inhibitor of angiogenic receptor tyrosine kinases, and the immunostimulator B7.2-IgG fusion protein. Cancer Res. 2002, 62, 5727–5735. [Google Scholar]
- Allen, E.; Jabouille, A.; Rivera, L.B.; Lodewijckx, I.; Missiaen, R.; Steri, V.; Feyen, K.; Tawney, J.; Hanahan, D.; Michael, I.P.; et al. Combined antiangiogenic and anti–PD-L1 therapy stimulates tumor immunity through HEV formation. Sci. Transl. Med. 2017, 9, eaak9679. [Google Scholar] [CrossRef] [PubMed]
- Schmittnaegel, M.; Rigamonti, N.; Kadioglu, E.; Cassará, A.; Rmili, C.W.; Kiialainen, A.; Kienast, Y.; Mueller, H.-J.; Ooi, C.-H.; Laoui, D.; et al. Dual angiopoietin-2 and VEGFA inhibition elicits antitumor immunity that is enhanced by PD-1 checkpoint blockade. Sci. Transl. Med. 2017, 9, eaak9670. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.T.; Man, S.; Xu, P.; Chow, A.; Paez-Ribes, M.; Lee, C.R.; Pirie-Shepherd, S.R.; Emmenegger, U.; Kerbel, R.S. Efficacy of Cotargeting Angiopoietin-2 and the VEGF Pathway in the Adjuvant Postsurgical Setting for Early Breast, Colorectal, and Renal Cancers. Cancer Res. 2016, 76, 6988–7000. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Jia, W.; Deng, H.; Li, G.; Deng, W.; Chen, J.; Kim, B.Y.; Jiang, W.; Liu, Q.; et al. Low-Dose Anti-Angiogenic Therapy Sensitizes Breast Cancer to PD-1 Blockade. Clin. Cancer Res. 2020, 26, 1712–1724. [Google Scholar] [CrossRef]
- Wu, F.; Xu, P.; Chow, A.; Man, S.; Krüger, J.; Khan, K.; Paez-Ribes, M.; Pham, E.; Kerbel, R.S. Pre- and post-operative anti-PD-L1 plus anti-angiogenic therapies in mouse breast or renal cancer models of micro- or macro-metastatic disease. Br. J. Cancer 2018, 120, 196–206. [Google Scholar] [CrossRef]
- Tian, L.; Goldstein, A.; Wang, H.; Lo, H.C.; Kim, I.S.; Welte, T.; Sheng, K.; Dobrolecki, L.E.; Zhang, X.; Putluri, N.; et al. Mutual regulation of tumour vessel normalization and immunostimulatory reprogramming. Nature 2017, 544, 250–254. [Google Scholar] [CrossRef]
- Kabir, A.U.; Subramanian, M.; Lee, D.H.; Wang, X.; Krchma, K.; Wu, J.; Naismith, T.; Halabi, C.M.; Kim, J.Y.; Pulous, F.E.; et al. Dual role of endothelial Myct1 in tumor angiogenesis and tumor immunity. Sci. Transl. Med. 2021, 13, eabb6731. [Google Scholar] [CrossRef]
- Panagi, M.; Voutouri, C.; Mpekris, F.; Papageorgis, P.; Martin, M.R.; Martin, J.D.; Demetriou, P.; Pierides, C.; Polydorou, C.; Stylianou, A.; et al. TGF-β inhibition combined with cytotoxic nanomedicine normalizes triple negative breast cancer microenvironment towards anti-tumor immunity. Theranostics 2020, 10, 1910–1922. [Google Scholar] [CrossRef]
- Zeng, Y.; Yu, T.; Zhang, S.; Song, G.; Meng, T.; Yuan, H.; Hu, F. Combination of tumor vessel normalization and immune checkpoint blockade for breast cancer treatment via multifunctional nanocomplexes. Biomater. Sci. 2022, 10, 4140–4155. [Google Scholar] [CrossRef]
- Thomssen, C.; Pierga, J.-Y.; Pritchard, K.I.; Biganzoli, L.; Cortes-Funes, H.; Petráková, K.; Kaufman, B.; Duenne, A.; Smith, I. First-Line Bevacizumab-Containing Therapy for Triple-Negative Breast Cancer: Analysis of 585 Patients Treated in the ATHENA Study. Oncology 2012, 82, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Robert, N.J.; Diéras, V.; Glaspy, J.; Brufsky, A.M.; Bondarenko, I.; Lipatov, O.N.; Perez, E.A.; Yardley, D.A.; Chan, S.Y.; Zhou, X.; et al. RIBBON-1: Randomized, Double-Blind, Placebo-Controlled, Phase III Trial of Chemotherapy With or Without Bevacizumab for First-Line Treatment of Human Epidermal Growth Factor Receptor 2–Negative, Locally Recurrent or Metastatic Breast Cancer. J. Clin. Oncol. 2011, 29, 1252–1260. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, E.; Kimmick, G.; Hopkins, J.; Marcom, P.K.; Rocha, G.; Welch, R.; Broadwater, G.; Blackwell, K. Nab-Paclitaxel/Bevacizumab/Carboplatin Chemotherapy in First-Line Triple Negative Metastatic Breast Cancer. Clin. Breast Cancer 2013, 13, 416–420. [Google Scholar] [CrossRef]
- Brufsky, A.; Valero, V.; Tiangco, B.; Dakhil, S.; Brize, A.; Rugo, H.S.; Rivera, R.; Duenne, A.; Bousfoul, N.; Yardley, D.A. Second-line bevacizumab-containing therapy in patients with triple-negative breast cancer: Subgroup analysis of the RIBBON-2 trial. Breast Cancer Res. Treat. 2012, 133, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Sasich, L.D.; Sukkari, S.R. The US FDAs withdrawal of the breast cancer indication for Avastin (bevacizumab). Saudi Pharm. J. 2011, 20, 381–385. [Google Scholar] [CrossRef]
- Cameron, D.; Brown, J.; Dent, R.; Jackisch, C.; Mackey, J.; Pivot, X.; Steger, G.G.; Suter, T.M.; Toi, M.; Parmar, M.; et al. Adjuvant bevacizumab-containing therapy in triple-negative breast cancer (BEATRICE): Primary results of a randomised, phase 3 trial. Lancet Oncol. 2013, 14, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Rini, B.I.; Plimack, E.R.; Stus, V.; Gafanov, R.; Hawkins, R.; Nosov, D.; Pouliot, F.; Alekseev, B.; Soulières, D.; Melichar, B.; et al. Pembrolizumab plus Axitinib versus Sunitinib for Advanced Renal-Cell Carcinoma. N. Engl. J. Med. 2019, 380, 1116–1127. [Google Scholar] [CrossRef]
- Socinski, M.A.; Jotte, R.M.; Cappuzzo, F.; Orlandi, F.; Stroyakovskiy, D.; Nogami, N.; Rodríguez-Abreu, D.; Moro-Sibilot, D.; Thomas, C.A.; Barlesi, F.; et al. Atezolizumab for First-Line Treatment of Metastatic Nonsquamous NSCLC. N. Engl. J. Med. 2018, 378, 2288–2301. [Google Scholar] [CrossRef]
- Finn, R.S.; Qin, S.; Ikeda, M.; Galle, P.R.; Ducreux, M.; Kim, T.-Y.; Kudo, M.; Breder, V.; Merle, P.; Kaseb, A.O.; et al. Atezolizumab plus Bevacizumab in Unresectable Hepatocellular Carcinoma. N. Engl. J. Med. 2020, 382, 1894–1905. [Google Scholar] [CrossRef]
- Makker, V.; Rasco, D.; Vogelzang, N.J.; Brose, M.S.; Cohn, A.L.; Mier, J.; Di Simone, C.; Hyman, D.M.; Stepan, D.E.; Dutcus, C.E.; et al. Lenvatinib plus pembrolizumab in patients with advanced endometrial cancer: An interim analysis of a multicentre, open-label, single-arm, phase 2 trial. Lancet Oncol. 2019, 20, 711–718. [Google Scholar] [CrossRef]
- Wu, S.-Y.; Xu, Y.; Chen, L.; Fan, L.; Ma, X.-Y.; Zhao, S.; Song, X.-Q.; Hu, X.; Yang, W.-T.; Chai, W.-J.; et al. Combined angiogenesis and PD-1 inhibition for immunomodulatory TNBC: Concept exploration and biomarker analysis in the FUTURE-C-Plus trial. Mol. Cancer 2022, 21, 1–15. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Y.; Tian, Z.; Lin, Y.; Li, H.; Zhu, Z.; Liu, Q.; Su, S.; Zeng, Y.; Jia, W.; et al. Multicenter phase II trial of Camrelizumab combined with Apatinib and Eribulin in heavily pretreated patients with advanced triple-negative breast cancer. Nat. Commun. 2022, 13, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Quintela-Fandino, M.; Holgado, E.; Manso, L.; Morales, S.; Bermejo, B.; Colomer, R.; Apala, J.V.; Blanco, R.; Muñoz, M.; Caleiras, E.; et al. Immuno-priming durvalumab with bevacizumab in HER2-negative advanced breast cancer: A pilot clinical trial. Breast Cancer Res. 2020, 22, 1–14. [Google Scholar] [CrossRef]
- Quintela-Fandino, M.; Sànchez, L.M.; Martín, E.H.; Moreno, M.; Murillo, S.M.; Heras, B.B.D.L.; Gimenez, D.M.; Bosch, R.C.; Cortijo, L.G.; Hornedo, J.; et al. Addition of durvalumab (Dur) upon progression to bevacizumab (Bev) maintenance in advanced HER2-negative (HERNEG) breast cancer (BC): Safety, efficacy and biomarkers. Ann. Oncol. 2018, 29, ix24. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, J.; Jiang, Y. Partial Response After Toripalimab Plus Anlotinib for Advanced Metaplastic Breast Carcinoma: A Case Report. Front. Endocrinol. 2022, 13, 810747. [Google Scholar] [CrossRef]
- Xuelian, C.; Lanying, M.; Xi, W.; Hongnan, M.; Dawei, W.; Bo, L.; Dong, Q.; Hongtu, Z.; Jing, H.; Binghe, X. Reactive capillary hemangiomas: A novel dermatologic toxicity following anti-PD-1 treatment with SHR-1210. Cancer Biol. Med. 2019, 16, 173–181. [Google Scholar] [CrossRef]
- Fadlallah, A.; Chelala, E.; Dirani, A. Intravitreal anti-VEGF injection for the treatment of progressive juxtapapillary retinal capillary hemangioma: A case report and mini review of the literature. Clin. Ophthalmol. 2013, 7, 2143–2146. [Google Scholar] [CrossRef]
- Qin, L.; Li, X.; Stroiney, A.; Qu, J.; Helgager, J.; Reardon, D.A.; Young, G.S. Advanced MRI assessment to predict benefit of anti-programmed cell death 1 protein immunotherapy response in patients with recurrent glioblastoma. Neuroradiology 2017, 59, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, W.S.; Ley, C.D.; Farrar, C.; Duyverman, A.M.; Lahdenranta, J.; Lacorre, D.A.; Batchelor, T.T.; Di Tomaso, E.; Duda, D.G.; Munn, L.L.; et al. Edema Control by Cediranib, a Vascular Endothelial Growth Factor Receptor–Targeted Kinase Inhibitor, Prolongs Survival Despite Persistent Brain Tumor Growth in Mice. J. Clin. Oncol. 2009, 27, 2542–2552. [Google Scholar] [CrossRef]
- Chen, W.; Shen, L.; Jiang, J.; Zhang, L.; Zhang, Z.; Pan, J.; Ni, C.; Chen, Z. Antiangiogenic therapy reverses the immunosuppressive breast cancer microenvironment. Biomark. Res. 2021, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-L.; Sainson, R.C.; Oon, C.E.; Turley, H.; Leek, R.; Sheldon, H.; Bridges, E.; Shi, W.; Snell, C.; Bowden, E.T.; et al. DLL4-Notch Signaling Mediates Tumor Resistance to Anti-VEGF Therapy In Vivo. Cancer Res. 2011, 71, 6073–6083. [Google Scholar] [CrossRef]
- Galluzzi, L.; Humeau, J.; Buqué, A.; Zitvogel, L.; Kroemer, G. Immunostimulation with chemotherapy in the era of immune checkpoint inhibitors. Nat. Rev. Clin. Oncol. 2020, 17, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Wang, Y.; He, P.; Shao, B.; Liu, F.; Xiang, Z.; Yang, T.; Zeng, Y.; He, T.; Ma, J.; et al. Effective Combinations of Immunotherapy and Radiotherapy for Cancer Treatment. Front. Oncol. 2022, 12, 809304. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Rank | NCT Number | Status | Conditions | Interventions |
|---|---|---|---|---|
| 1 | NCT03961698 | Active, not recruiting | Breast Cancer, Renal Cell Carcinoma | IPI-549 (eganelisib), Atezolizumab, nab-paclitaxel, Bevacizumab |
| 2 | NCT03395899 | Active, not recruiting | Breast Cancer, Estrogen Receptor-positive Breast Cancer | Atezolizumab, Cobimetinib, Ipatasertib, Bevacizumab |
| 3 | NCT03280563 | Active, not recruiting | Breast Neoplasms | Atezolizumab (MPDL3280A), an engineered anti-programmed death-ligand 1 (PD-L1) antibody, Bevacizumab, Entinostat, Exemestane, Fulvestrant, Ipatasertib, Tamoxifen, Abemaciclib |
| 4 | NCT03387085 | Active, not recruiting | Triple-Negative Breast Cancer | Aldoxorubicin HCl, N-803, ETBX-011, ETBX-051, ETBX-061, GI-4000, GI-6207, GI-6301, haNK for Infusion, avelumab, bevacizumab, Capecitabine, Cisplatin, Cyclophosphamide, 5-Fluorouracil, Leucovorin, nab-Paclitaxel, SBRT |
| 5 | NCT02734004 | Active, not recruiting | Ovarian, Breast, SCLC, Gastric Cancers | Olaparib, MEDI4736, Bevacizumab |
| 6 | NCT03761914 | Active, not recruiting | Breast Cancer, Other Tumors | galinpepimut-S, Pembrolizumab |
| 7 | NCT03170960 | Active, not recruiting | Triple-Negative Breast Neoplasm, Other Solid Tumors | cabozantinib, atezolizumab |
| 8 | NCT02009449 | Active, not recruiting | Solid Tumors, Breast Cancer | Pegilodecakin, Paclitaxel or Docetaxel and Carboplatin or Cisplatin, FOLFOX (Oxaliplatin/Leucovorin/5-Fluorouracil), gemcitabine/nab-paclitaxel, Capecitabine, Pazopanib, Pembrolizumab, Paclitaxel, nivolumab, Gemcitabine/carboplatin |
| 9 | NCT05431582 | Not yet recruiting | Ovarian Cancer, Breast Cancer, Lung Cancer, Pancreatic Cancer | ZN-c3, Bevacizumab, Pembrolizumab |
| 10 | NCT04739670 | Recruiting | Metastatic Triple-Negative Breast Cancer | Atezolizumab, Bevacizumab, Gemcitabine, Carboplatin |
| 11 | NCT04732598 | Recruiting | Breast Cancer | Paclitaxel + bevacizumab therapy, Paclitaxel + bevacizumab + atezolizumab |
| 12 | NCT04408118 | Recruiting | Metastatic Breast Cancer, Advanced Breast Cancer, Triple-Negative Breast Cancer | Atezolizumab, Paclitaxel, Bevacizumab |
| 13 | NCT05180006 | Recruiting | Breast Cancer | Atezolizumab Injection, Ipatasertib, Bevacizumab, Pertuzumab, Trastuzumab |
| 14 | NCT05007106 | Recruiting | Triple-Negative Breast Neoplasm, Other Solid Tumors | Pembrolizumab/Vibostolimab Co-Formulation, Pembrolizumab, Lenvatinib, 5-Fluorouracil, Cisplatin, Paclitaxel, Gemcitabine, Carboplatin, Docetaxel, Bevacizumab, Capecitabine, Oxaliplatin |
| 15 | NCT05092373 | Recruiting | Advanced Breast Carcinoma, Advanced Other Solid Tumors | Atezolizumab, Cabozantinib S-malate, Nab-paclitaxel, Tumor Treating Fields Therapy |
| 16 | NCT04514484 | Recruiting | Advanced Triple-Negative Breast Carcinoma, Other Advanced Solid Tumors | Cabozantinib S-malate, Computed Tomography, Magnetic Resonance Imaging, Nivolumab |
| 17 | NCT04802759 | Recruiting | Inoperable, Locally Advanced or Metastatic, ER-positive Breast Cancer | Giredestrant, Abemaciclib, Ipatasertib, Inavolisib, Ribociclib, Everolimus, Samuraciclib, PH FDC SC, Palbociclib, Atezolizumab |
| 18 | NCT04591431 | Recruiting | Breast Cancer, Gastrointestinal Cancer, Non-Small-Cell Lung Cancer | Erlotinib, Trastuzumab, Trastuzumab emtansine, Pertuzumab, Lapatinib, Everolimus, Vemurafenib, Cobimetinib, Alectinib, Brigatinib, Palbociclib, Ponatinib, Vismogedib, Itacitinib, Ipatasertib, Entrectinib, Atezolizumab, Nivolumab, Ipilimumab, Pemigatinib, Oncology Drugs |
| 19 | NCT03878524 | Recruiting | Anatomic Stage IV Breast Cancer AJCC v8, Metastatic Breast Carcinoma, Other Advanced Solid Tumors | Abemaciclib, Abiraterone, Afatinib, Bevacizumab, Bicalutamide, Biospecimen Collection, Bortezomib, Cabazitaxel, Cabozantinib, Capecitabine, Carboplatin, Celecoxib, Cobimetinib, Copanlisib, Dabrafenib, Dacomitinib, Darolutamide, Dasatinib, Doxorubicin, Durvalumab, Enasidenib, Entrectinib, Enzalutamide, Erlotinib, Everolimus, Fluorouracil, Idelalisib, Imatinib, Ipilimumab, Lenvatinib, Leucovorin, Lorlatinib, Losartan, Nab-paclitaxel, Neratinib, Nivolumab, Olaparib, Oxaliplatin, Palbociclib, Panobinostat, Pembrolizumab, Pertuzumab, Ponatinib, Quality-of-Life Assessment, Regorafenib, Ruxolitinib, Sirolimus, Sorafenib, Sunitinib, Trametinib, Trastuzumab Emtansine, Tretinoin, Vemurafenib, Venetoclax, Vismodegib, Vorinostat |
| 20 | NCT02802098 | Completed | Metastatic Breast Cancer, Bevacizumab-alone Maintenance Treatment Progression | Durvalumab, Bevacizumab |
| 21 | NCT03316586 | Completed | Breast Cancer | Nivolumab, Cabozantinib |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melaiu, O.; Vanni, G.; Portarena, I.; Pistolese, C.A.; Anemona, L.; Pomella, S.; Bei, R.; Buonomo, O.C.; Roselli, M.; Mauriello, A.; et al. The Combination of Immune Checkpoint Blockade with Tumor Vessel Normalization as a Promising Therapeutic Strategy for Breast Cancer: An Overview of Preclinical and Clinical Studies. Int. J. Mol. Sci. 2023, 24, 3226. https://doi.org/10.3390/ijms24043226
Melaiu O, Vanni G, Portarena I, Pistolese CA, Anemona L, Pomella S, Bei R, Buonomo OC, Roselli M, Mauriello A, et al. The Combination of Immune Checkpoint Blockade with Tumor Vessel Normalization as a Promising Therapeutic Strategy for Breast Cancer: An Overview of Preclinical and Clinical Studies. International Journal of Molecular Sciences. 2023; 24(4):3226. https://doi.org/10.3390/ijms24043226
Chicago/Turabian StyleMelaiu, Ombretta, Gianluca Vanni, Ilaria Portarena, Chiara Adriana Pistolese, Lucia Anemona, Silvia Pomella, Roberto Bei, Oreste Claudio Buonomo, Mario Roselli, Alessandro Mauriello, and et al. 2023. "The Combination of Immune Checkpoint Blockade with Tumor Vessel Normalization as a Promising Therapeutic Strategy for Breast Cancer: An Overview of Preclinical and Clinical Studies" International Journal of Molecular Sciences 24, no. 4: 3226. https://doi.org/10.3390/ijms24043226
APA StyleMelaiu, O., Vanni, G., Portarena, I., Pistolese, C. A., Anemona, L., Pomella, S., Bei, R., Buonomo, O. C., Roselli, M., Mauriello, A., & Barillari, G. (2023). The Combination of Immune Checkpoint Blockade with Tumor Vessel Normalization as a Promising Therapeutic Strategy for Breast Cancer: An Overview of Preclinical and Clinical Studies. International Journal of Molecular Sciences, 24(4), 3226. https://doi.org/10.3390/ijms24043226