



Protective Role of Ethanol Extract of Cibotium barometz (Cibotium Rhizome) against Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes

Abstract
1. Introduction
2. Results
2.1. HPLC Analysis
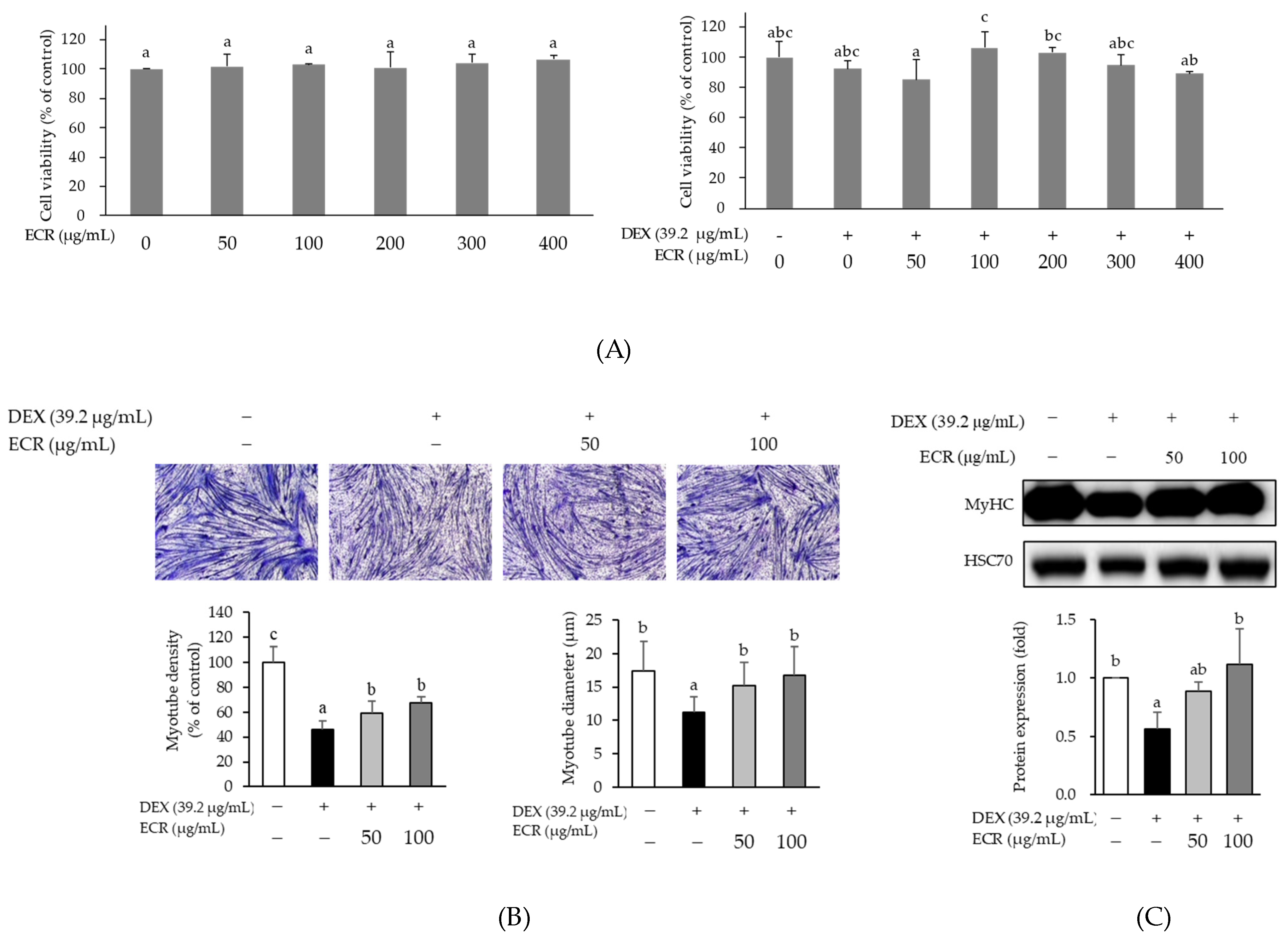
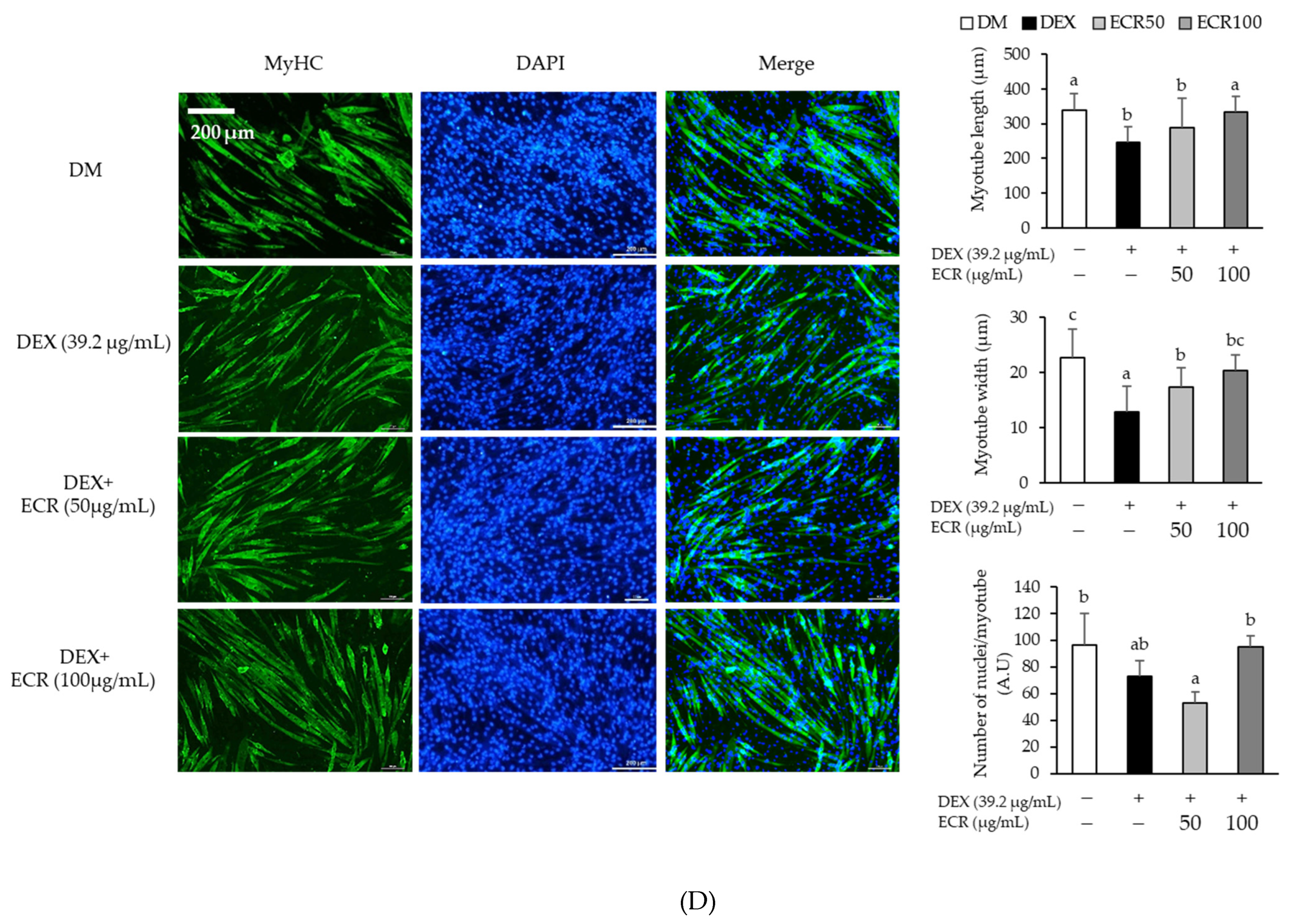
2.2. Alleviating Effects of ECR in DEX-Induced Muscle Atrophic Cells
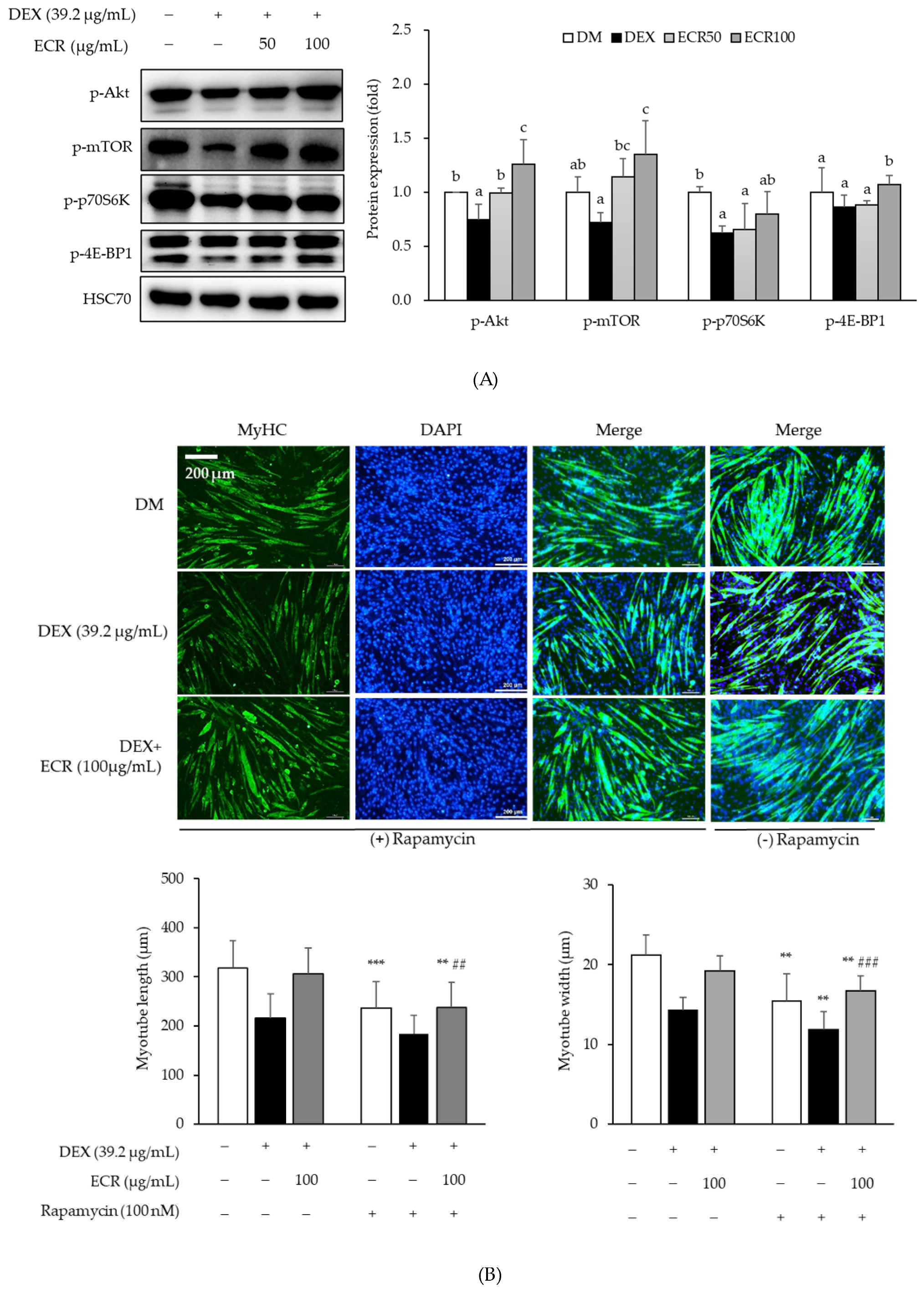
2.3. Effects of ECR on Protein Synthesis via the Akt/mTOR/S6K1 Pathway
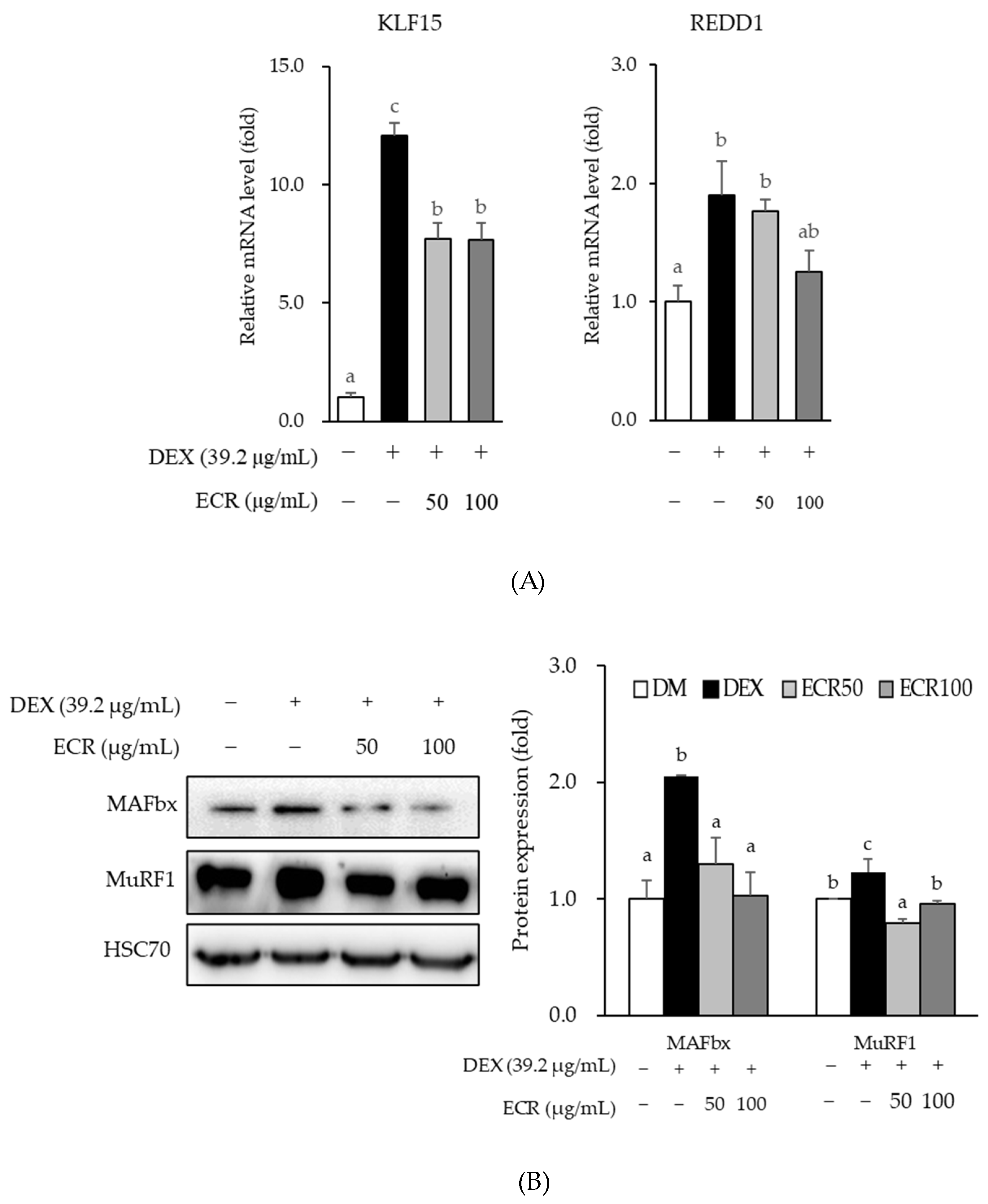
2.4. Effects of ECR on Protein Degradation in Myotubes
2.5. Component-Target-Sarcopenia/Muscle Atrophy Pathway Interaction
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Sample Preparation
4.3. Compound Analysis of ECR by HPLC
4.4. Cell Culture and Differentiation
4.5. Jenner–Giemsa Staining
4.6. Western Blot Analysis
4.7. Quantitative Real-Time-PCR
4.8. Immunofluorescence Staining
4.9. Computational Database Analysis
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, N.-H.; Kim, C.Y. Association of micronutrients and handgrip strength in Korean older population: A cross-sectional study. Healthcare 2022, 10, 1980. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Baeyens, J.P.; Bauer, J.M.; Boirie, Y.; Cederholm, T.; Landi, F.; Martin, F.C.; Michel, J.P.; Rolland, Y.; Schneider, S.M.; et al. Sarcopenia: European consensus on definition and diagnosis: Report of the European Working Group on Sarcopenia in Older People. Age Ageing 2010, 39, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, H.; Zeng, Z.; Lv, J.; Huang, J.; Wu, X.; Wang, M.; Xu, J.; Fan, J.; Chen, N. MicroRNA profiling of different exercise interventions for alleviating skeletal muscle atrophy in naturally aging rats. J. Cachexia Sarcopenia Muscle 2023, 14, 356–368. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Sayer, A.A. Sarcopenia. Lancet 2019, 393, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Bonaldo, P.; Sandri, M. Cellular and molecular mechanisms of muscle atrophy. Dis. Models Mech. 2013, 6, 25–39. [Google Scholar] [CrossRef]
- Sandri, M.; Sandri, C.; Gilbert, A.; Skurk, C.; Calabria, E.; Picard, A.; Walsh, K.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 2004, 117, 399–412. [Google Scholar] [CrossRef] [PubMed]
- Stitt, T.N.; Drujan, D.; Clarke, B.A.; Panaro, F.; Timofeyva, Y.; Kline, W.O.; Gonzalez, M.; Yancopoulos, G.D.; Glass, D.J. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol. Cell 2004, 114, 395–403. [Google Scholar] [CrossRef]
- Shah, O.J.; Kimball, S.R.; Jefferson, L.S. Acute attenuation of translation initiation and protein synthesis by glucocorticoids in skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E76–E82. [Google Scholar] [CrossRef]
- Shah, O.J.; Kimball, S.R.; Jefferson, L.S. Glucocorticoids abate p70 (S6k) and eIF4E function in L6 skeletal myoblasts. Am. J. Physiol. Endocrinol. Metab. 2000, 279, E74–E82. [Google Scholar] [CrossRef]
- Chung, J.; Kuo, C.J.; Crabtree, G.R.; Blenis, J. Rapamycin-FKBP specifically blocks growth-dependent activation of and signaling by the 70 kd S6 protein kinases. Cell 1992, 69, 1227–1236. [Google Scholar] [CrossRef]
- Kuo, C.J.; Chung, J.; Fiorentino, D.F.; Flanagan, W.M.; Blenis, J.; Crabtree, G.R. Rapamycin selectively inhibits interleukin-2activation of p70 S6 kinase. Nature 1992, 358, 70–73. [Google Scholar] [CrossRef]
- Goldberg, A.L. Protein turnover in skeletal muscle. II. Effects of denervation and cortisone on protein catabolism in skeletal muscle. J. Biol. Chem. 1969, 244, 3223–3229. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Mallidis, C.; Bhasin, S.; Mahabadi, V.; Artaza, J.; Gonzalez-Cadavid, N.; Arias, J.; Salehian, B. Glucocorticoid-induced skeletal muscle atrophy is associated with upregulation of myostatin gene expression. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E363–E371. [Google Scholar] [CrossRef] [PubMed]
- Watson, M.L.; Baehr, L.M.; Reichardt, H.M.; Tuckermann, J.P.; Bodine, S.C.; Furlow, J.D. A cell autonomous role for the glucocorticoid receptor in skeletal muscle atrophy induced by systemic glucocorticoid exposure. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1210–E1220. [Google Scholar] [CrossRef] [PubMed]
- Jhuo, C.F.; Hsieh, S.K.; Chen, W.Y.; Tzen, J.T.C. Attenuation of Skeletal Muscle Atrophy Induced by Dexamethasone in Rats by Teaghrelin Supplementation. Molecules 2023, 28, 688. [Google Scholar] [CrossRef]
- Shimizu, N.; Yoshikawa, N.; Ito, N.; Maruyama, T.; Suzuki, Y.; Takeda, S.; Nakae, J.; Tagata, Y.; Nishitani, S.; Takehana, K.; et al. Crosstalk between glucocorticoid receptor and nutritional sensor mTOR in skeletal muscle. Cell Metab. 2011, 13, 170–182. [Google Scholar] [CrossRef]
- Waddell, D.S.; Baehr, L.M.; van den Brandt, J.; Johnsen, S.A.; Reichardt, H.M.; Furlow, J.D.; Bodine, S.C. The glucocorticoid receptor and FOXO1 synergistically activate the skeletal muscle atrophy-associated MuRF1 gene. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E785–E797. [Google Scholar] [CrossRef]
- Black, A.J.; Gordon, B.S.; Dennis, M.D.; Jefferson, L.S.; Kimball, S.R. Regulation of protein and mRNA expression of the mTORC1 repressor REDD1 in response to leucine and serum. Biochem. Biophys. Rep. 2016, 8, 296–301. [Google Scholar] [CrossRef][Green Version]
- Kimball, S.R.; Do, A.N.D.; Kutzler, L.; Cavener, D.R.; Jefferson, L.S. Rapid turnover of the mTOR complex 1 (mTORC1) repressor REDD1 and activation of mTORC1 signaling following inhibition of protein synthesis. J. Biol. Chem. 2008, 283, 3465–3475. [Google Scholar] [CrossRef]
- Kim, S.-T.; Han, Y.N.; Shon, Y.K.; Jang, H.S.; Kim, S.J.; Shin, J.S. Isolation of the efficacy constituent for neuronal regeneration from Cibotium barometz. Yakhak Hoeji 2002, 46, 398–404. Available online: https://www.yakhak.org/journal/view.html?spage=398&volume=46&number=6 (accessed on 5 January 2023).
- Liu, H.; Gao, J.-H.; Xie, Y.M.; Li, Y.Y.; Li, S.H.; Zhang, L.; Xu, Z.J.; Huang, F.; Jin, J.; Li, G. Interpretation of Expert consensus on Shujin Jianyao Pills in clinical practice. Zhongguo Zhong Yao Za Zhi 2020, 45, 3336–3339. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J.Y.; Kim, C.Y. Allium macrostemon whole extract ameliorates obesity-induced inflammation and endoplasmic reticulum stress in adipose tissue of high-fat diet-fed C57BL/6N mice. Food Nutr. Res. 2023, 18, 67. [Google Scholar] [CrossRef]
- Silva, E.L.; Almeida-Lafetá, R.C.; Borges, R.M.; Staerk, D. Athenolide A, a New Steroidal Lactone from the Leaves of Athenaea martiana (Solanaceae) Determined by Means of HPLC-HR-MS-SPE-NMR Analysis. Chem. Biodivers. 2008, 15, e1700455. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rajaonson, S.; Diallo, B.; Mol, A.; El Jaziri, M.; Baucher, M. Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Appl. Environ. Microbiol. 2010, 76, 243–253. [Google Scholar] [CrossRef]
- Taher, R.F.; Raslan, M.A.; Masoud, M.A.; Nassar, M.I.; Aboutabl, M.E. HPLC-ESI/MS profiling, phytoconstituent isolation and evaluation of renal function, oxidative stress and inflammation in gentamicin-induced nephrotoxicity in rats of Ficus spragueana Mildbr. & Burret. Biomed. Chromatogr. 2021, 35, e5135. [Google Scholar] [CrossRef]
- Khan, M.R.U.Z.; Yanase, E.; Trivedi, V. Extraction, phytochemical characterization and anti-cancer mechanism of Haritaki churna: An ayurvedic formulation. PLoS ONE 2023, 18, e0286274. [Google Scholar] [CrossRef] [PubMed]
- Taban, A.K.; Gürbüz, P.; Doğan, Ş.D.; Emerce, E.; Gören, A.C.; Polat, R.; Süntar, İ. Two new eudesmane-type sesquiterpene derivatives from Lecokia cretica (Lam.) DC. Nat. Prod. Res. 2022, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kirk, B.; Feehan, J.; Lombardi, G.; Duque, G. Muscle, Bone, and Fat Crosstalk: The Biological Role of Myokines, Osteokines, and Adipokines. Curr. Osteoporos. Rep. 2020, 18, 388–400. [Google Scholar] [CrossRef]
- Bosco, F.; Musolino, V.; Gliozzi, M.; Nucera, S.; Carresi, C.; Zito, M.C.; Scarano, F.; Scicchitano, M.; Reale, F.; Ruga, S.; et al. The muscle to bone axis (and viceversa): An encrypted language affecting tissues and organs and yet to be codified? Pharmacol. Res. 2021, 165, 105427. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.K. Cibotium barometz. Edible Medicinal and Non-Medicinal. Plants 2015, 22, 82–91. [Google Scholar] [CrossRef]
- Li, L.; Xie, M.P.; Sun, H.; Lu, A.Q.; Zhang, B.; Zhang, D.; Wang, S.J. Bioactive phenolic acid-substituted glycoses and glycosides from rhizomes of Cibotium barometz. J. Asian Nat. Prod. Res. 2019, 10, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Mai, W.; Chen, D.; Li, X. Antioxidant Activity of Rhizoma Cibotii in vitro. Adv. Pharm. Bull. 2012, 2, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.Z.; Zang, J.P. Comparative study on essential oil in Rhizoma Cibotii and precessed product. China J. Chin. Mater. Med. 1996, 21, 216–217. Available online: https://typeset.io/journals/china-journal-of-chinese-materia-medica-3s54tb30/1996 (accessed on 23 January 2023).
- Wu, Q.; Yang, X.W. The constituents of Cibotium barometz and their permeability in the human Caco-2 monolayer cell model. J. Ethnopharmacol. 2009, 125, 417–422. [Google Scholar] [CrossRef]
- Lee, J.Y.; Ko, S.H.; Lee, Y.J.; Lee, S.Y.; Park, H.J.; Shin, T.Y.; Jeon, H. Anti-inflammatory effect of MeOH extract of Cibotium barometz in IFN-γ and LPS-stimulated mouse peritoneal macrophage. Korean J. Pharmacogn. 2010, 41, 108–114. Available online: http://kpubs.org/article/articleMain.kpubs?articleANo=HKSOBF_2010_v41n2_108 (accessed on 17 November 2022).
- Manning, B.D.; Cantley, L.C. AKT/PKB Signaling: Navigating Downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef]
- Marte, B.M.; Rodriguez-Viciana, P.; Wennstrom, S.; Warne, P.H.; Downward, J. R-Ras can activate the phosphoinositide 3-kinase but not the MAP kinase arm of the Ras effector pathways. Curr. Biol. 1997, 7, 63–70. [Google Scholar] [CrossRef]
- Anderson, K.E.; Coadwell, J.; Stephens, L.R.; Hawkins, P.T. Translocation of PDK-1 to the plasma membrane is important in allowing PDK-1 to activate protein kinase B. Curr. Biol. 1998, 8, 684–691. [Google Scholar] [CrossRef]
- Kwon, D.; Kim, C.; Woo, Y.K.; Hwang, J.K. Inhibitory Effects of Chrysanthemum (Chrysanthemum morifolium Ramat.) Extract and Its Active Compound Isochlorogenic Acid A on Sarcopenia. Prev. Nutr. Food Sci. 2021, 31, 408–416. [Google Scholar] [CrossRef]
- Kim, C.; Hwang, J.K. The 5,7-Dimethoxyflavone Suppresses Sarcopenia by Regulating Protein Turnover and Mitochondria Biogenesis-Related Pathways. Nutrients 2020, 12, 1079. [Google Scholar] [CrossRef]
- Lee, J.; Kim, C.; Lee, H.; Hwang, J.K. Inhibitory Effects of Standardized Leonurus japonicus Extract and Its Bioactive Leonurine on TNF-α-Induced Muscle Atrophy in L6 Myotubes. J. Microbiol. Biotechnol. 2020, 30, 1896–1904. [Google Scholar] [CrossRef]
- Salvadori, L.; Mandrone, M.; Manenti, T.; Ercolani, C.; Cornioli, L.; Lianza, M.; Tomasi, P.; Chiappalupi, S.; Di Filippo, E.S.; Fulle, S.; et al. Identification of Withania somnifera-Silybum marianum-Trigonella foenum-graecum Formulation as a Nutritional Supplement to Contrast Muscle Atrophy and Sarcopenia. Nutrients 2020, 13, 49. [Google Scholar] [CrossRef]
- Fernando, R.; Drescher, C.; Nowotny, K.; Grune, T.; Castro, J.P. Impaired proteostasis during skeletal muscle aging. Free Radic. Biol. Med. 2019, 132, 58–66. [Google Scholar] [CrossRef]
- Jackson, M.P.; Hewitt, E.W. Cellular proteostasis: Degradation of misfolded proteins by lysosomes. Essays Biochem. 2016, 60, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Voges, D.; Zwickl, P.; Baumeister, W. The 26S proteasome: A molecular machine designed for controlled proteolysis. Annu. Rev. Biochem. 1999, 68, 1015–1068. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [PubMed]
- Eo, H.; Reed, C.H.; Valentine, R.J. Imoxin prevents dexamethasone-induced promotion of muscle-specific E3 ubiquitin ligases and stimulates anabolic signaling in C2C12 myotubes. Biomed. Pharmacother. 2020, 128, 110238. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kubica, N.; Ellisen, L.W.; Jefferson, L.S.; Kimball, S.R. Dexamethasone represses signaling through the mammalian target of rapamycin in muscle cells by enhancing expression of REDD1. J. Biol. Chem. 2006, 281, 39128–39134. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Corradetti, M.N.; Inoki, K.; Guan, K.L. TSC2: Filling the GAP in the mTOR signaling pathway. Trends Biochem. Sci. 2004, 29, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; Harris, C.A.; Wang, J.C. Metabolic functions of glucocorticoid receptor in skeletal muscle. Mol. Cell. Endocrinol. 2013, 380, 79–88. [Google Scholar] [CrossRef]
- Wary, K.K.; Mariotti, A.; Zurzolo, C.; Giancotti, F.G. A requirement for caveolin-1 and associated kinase Fyn in integrin signaling and anchorage-dependent cell growth. Cell 1998, 94, 625–634. [Google Scholar] [CrossRef]
- Sha, G.; Sun, P.; Kong, X.; Han, X.; Sun, Q.; Fouillen, L.; Zhao, J.; Li, Y.; Yang, L.; Wang, Y.; et al. Genome editing of a rice CDP-DAG synthase confers multipathogen resistance. Nature 2023, 618, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.-H.; Xin, M.J.; Cha, J.Y.; Ji, S.J.; Kwon, S.U.; Jee, H.K.; Park, M.R.; Park, Y.S.; Kim, C.T.; Kim, D.K.; et al. Antitumor and Immunomodulatory Effect of Gastrodia elata on Colon Cancer In Vitro and In Vivo. Am. J. Chin. Med. 2017, 45, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Veliça, P.; Bunce, C.M. A quick, simple and unbiased method to quantify C2C12 myogenic differentiation. Muscle Nerve 2011, 44, 366–370. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lim, J.Y.; Choi, S.J. Role of l-carnitine and oleate in myogenic differentiation: Implications for myofiber regeneration. J. Exerc. Nutr. Biochem. 2018, 22, 36–42. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (CAS Number) | 2-Dimensional Chemical Structure | 3-Dimensional Chemical Structure | Retention Time (min) | Quantity (µg/g Extract) [22] | Classification | Absorption (nm) | DL OB (%) Caco-2 | Reference |
|---|---|---|---|---|---|---|---|---|
| Protocatechuic acid (99-50-3) |  |  | 8.724 | 562.28 ± 0.77 | Benzoates | 280 | 0.04 25.37 0.10 | [23] |
| (+)-Catechin hydrate (225937-10-0) |  |  | 11.744 | 2233.07 ± 1.56 | Catechins | 280 | 0.24 17.83 0.10 | [24] |
| p-Coumaric acid (501-98-4) |  |  | 20.995 | 166.90 ±52.64 | Monolignols | 280 | 0.04 43.29 0.46 | [25] |
| Ellagic acid (476-66-4) |  |  | 24.152 | 274.78 ± 1.48 | Tannins and Galloyl derivatives | 280 | 0.43 43.06 −0.44 | [26] |
| Chlorogenic acid (327-97-9) |  |  | 10.950 | 5068.92 ± 6.20 | Monolignols | 320 | 0.31 13.61 −1.33 | [27] |
| Caffeic acid (331-39-5) |  |  | 14.309 | 313.30 ± 65.53 | Monolignols | 320 | 0.05 54.97 0.27 | [23] |
| Ferulic acid (1135-24-6) |  |  | 24.148 | 874.05 ± 1.20 | Monolignols | 320 | 0.06 40.43 0.96 | [23] |
| Count | P. Adjust Value | |
|---|---|---|
| Biologic process | ||
| TORC2 signaling | 3 | 0.00028 |
| Mast cell differentiation | 2 | 0.0075 |
| Natural killer cell chemotaxis | 2 | 0.0075 |
| Caveola assembly | 2 | 0.0104 |
| Receptor-mediated endocytosis of virus by host cell | 2 | 0.0104 |
| Negative regulation of long-chain fatty acid import across plasma membrane | 2 | 0.0104 |
| Myeloid progenitor cell differentiation | 2 | 0.0104 |
| Postive regulation of neutrophil apoptotic process | 2 | 0.0133 |
| Establishment or maintenance of actin cytoskeleton polarity | 2 | 0.0133 |
| Mast cell chemotaxis | 2 | 0.0133 |
| Molecular function | ||
| Phosphatidylinositol-3,4-bisphosphate 5-kinase activity | 4 | 1.56 × 10−5 |
| Phosphatidylinositol-4,5-bisphosphate 3-kinase activity | 4 | 1.95 × 10−5 |
| 1-phosphatidylinositol-4-phosphate 3-kinase activity | 4 | 1.95 × 10−5 |
| 1-phosphatidylinositol-3-kinase activity | 4 | 4.60 × 10−5 |
| Phosphatidylinositol-3,4-trisphosphate binding | 4 | 0.0022 |
| Protein serine/threonine kinase activator activity | 4 | 0.0026 |
| Insulin receptor binding | 3 | 0.0120 |
| Phosphatidylinositol-3,4-bisphosphate binding | 3 | 0.0165 |
| Chitin binding | 2 | 0.0431 |
| Guanylate kinase activity | 2 | 0.0498 |
| Cellular component | ||
| TOR complex | 5 | 1.33 × 10−6 |
| Phosphatidylinositol-3-kinase complex | 5 | 2.93 × 10−5 |
| Extrinsic component of membrane | 10 | 2.93 × 10−5 |
| TORC2 complex | 4 | 2.99 × 10−5 |
| Caveolar macromolecular signaling complex | 2 | 0.0087 |
| Mast cell granule | 3 | 0.0090 |
| Cytoplasmic side of plasma membrane | 5 | 0.0202 |
| Phosphatidylinositol-3-kinase complex, class1 | 2 | 0.0207 |
| TORC1 complex | 2 | 0.0207 |
| Lamellipodium | 5 | 0.0448 |
| KEGG pathways | ||
| Longevity regulating pathway-multiple | 10 | 2.99 × 10−12 |
| Acute myeloid leukemia | 10 | 4.88 × 10−12 |
| Longevity regulating pathway | 10 | 4.34 × 10−11 |
| Central carbon metabolism in cancer | 9 | 1.99 × 10−10 |
| Regulation of lipolysis in adipocytes | 8 | 1.15 × 10−9 |
| Endometrial cancer | 8 | 1.53 × 10−9 |
| Non-small cell lung cancer | 8 | 3.98 × 10−9 |
| Carbohydrate digestion and absorption | 7 | 7.12 × 10−9 |
| VEGF signaling pathway | 7 | 2.76 × 10−8 |
| GnRH secretion | 7 | 4.42 × 10−8 |
| Target Gene | Primer Sequences (5′→3′) | NCBI Accession Number |
|---|---|---|
| KLF15 | F: TGGTACCATCCTCCAACTTGAA R: CAATAGGTTTGGCGGCAATG | NM_023184.4 |
| REDD1 | F: CCTAGCCTCTGGGATCGTTTC R: ATCAGCGGCCGGAGTTC | NM_029083.2 |
| 18s rRNA | F: AACCCGTTGAACCCCATT R: CCATCCAATCGGTAGTAGCG | NR_003278.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.-H.; Lee, J.-Y.; Kim, C.Y. Protective Role of Ethanol Extract of Cibotium barometz (Cibotium Rhizome) against Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes. Int. J. Mol. Sci. 2023, 24, 14798. https://doi.org/10.3390/ijms241914798
Kim N-H, Lee J-Y, Kim CY. Protective Role of Ethanol Extract of Cibotium barometz (Cibotium Rhizome) against Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes. International Journal of Molecular Sciences. 2023; 24(19):14798. https://doi.org/10.3390/ijms241914798
Chicago/Turabian StyleKim, Na-Hyung, Joo-Yeon Lee, and Choon Young Kim. 2023. "Protective Role of Ethanol Extract of Cibotium barometz (Cibotium Rhizome) against Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes" International Journal of Molecular Sciences 24, no. 19: 14798. https://doi.org/10.3390/ijms241914798
APA StyleKim, N.-H., Lee, J.-Y., & Kim, C. Y. (2023). Protective Role of Ethanol Extract of Cibotium barometz (Cibotium Rhizome) against Dexamethasone-Induced Muscle Atrophy in C2C12 Myotubes. International Journal of Molecular Sciences, 24(19), 14798. https://doi.org/10.3390/ijms241914798