
Lipid Peroxidation of the Docosahexaenoic Acid/Arachidonic Acid Ratio Relating to the Social Behaviors of Individuals with Autism Spectrum Disorder: The Relationship with Ferroptosis




Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Individuals with ASD
2.2. Dietary Nutrients
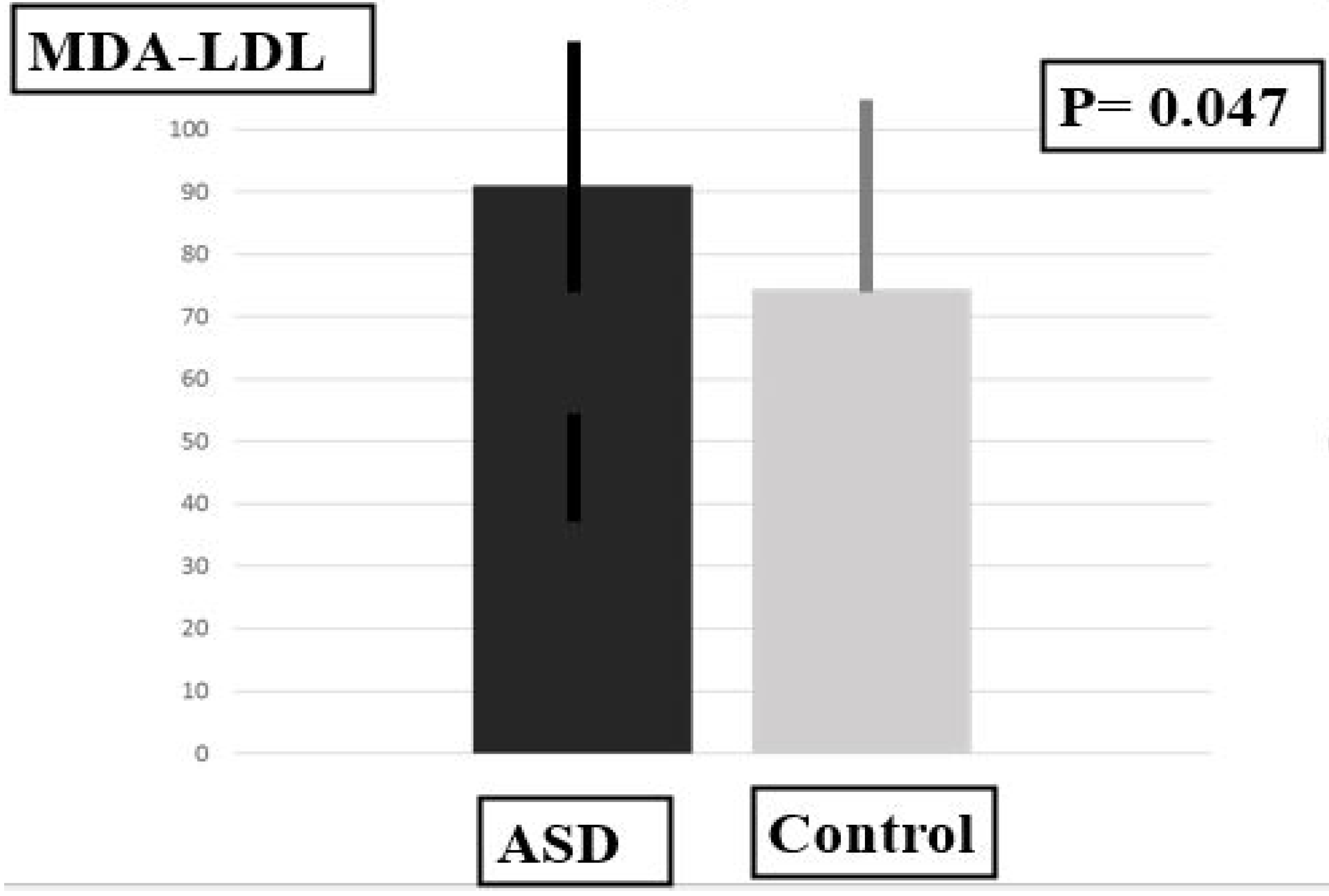
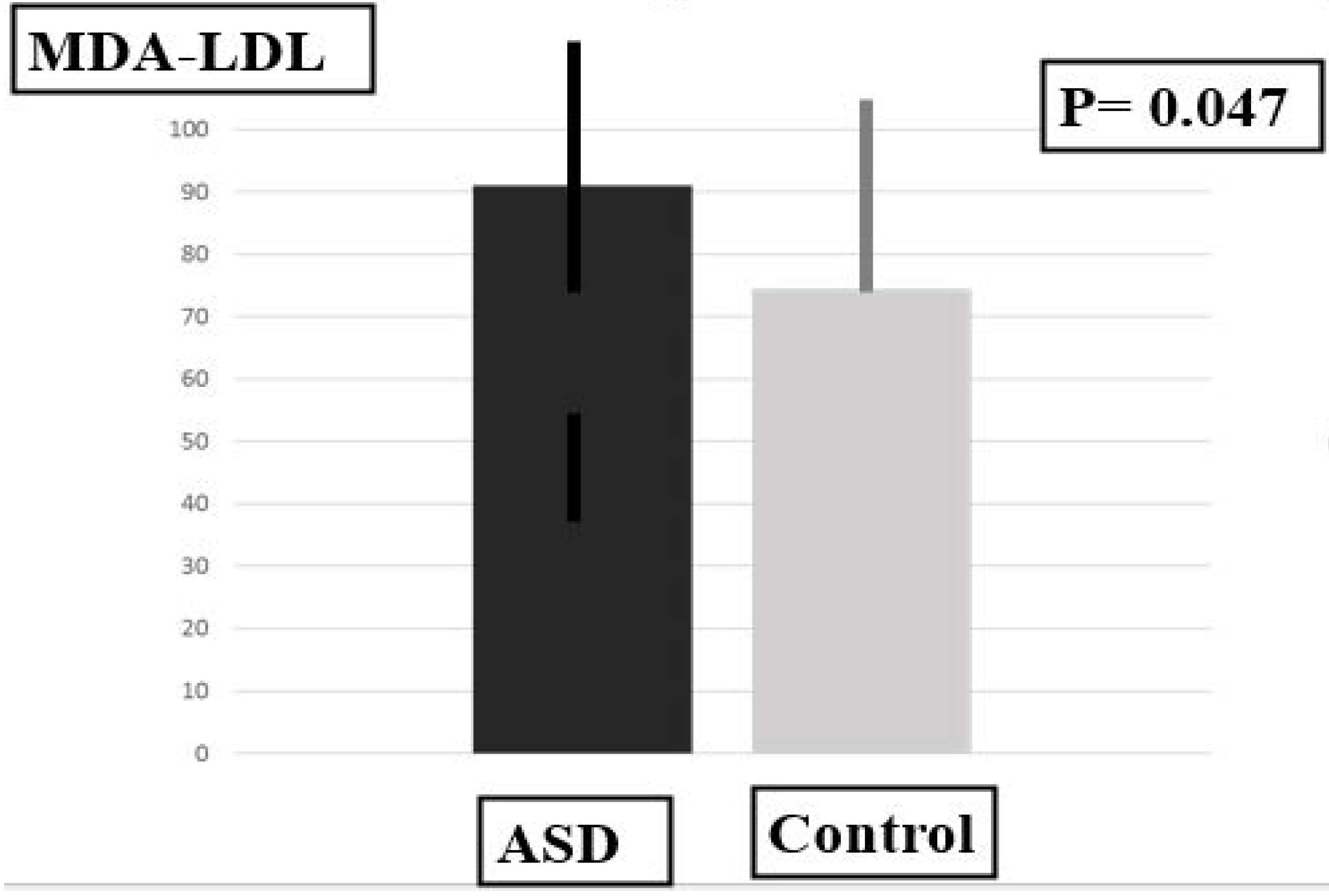
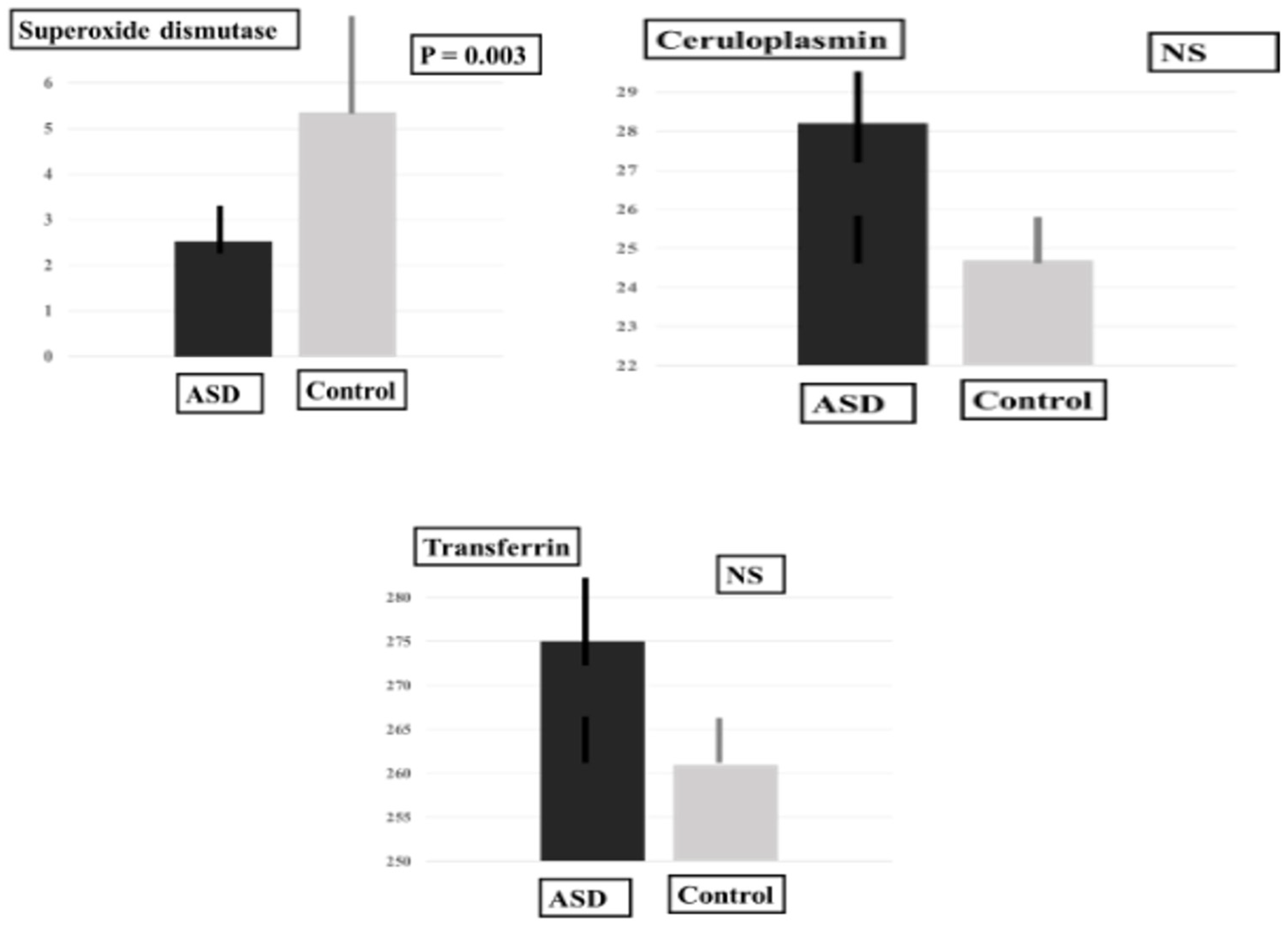
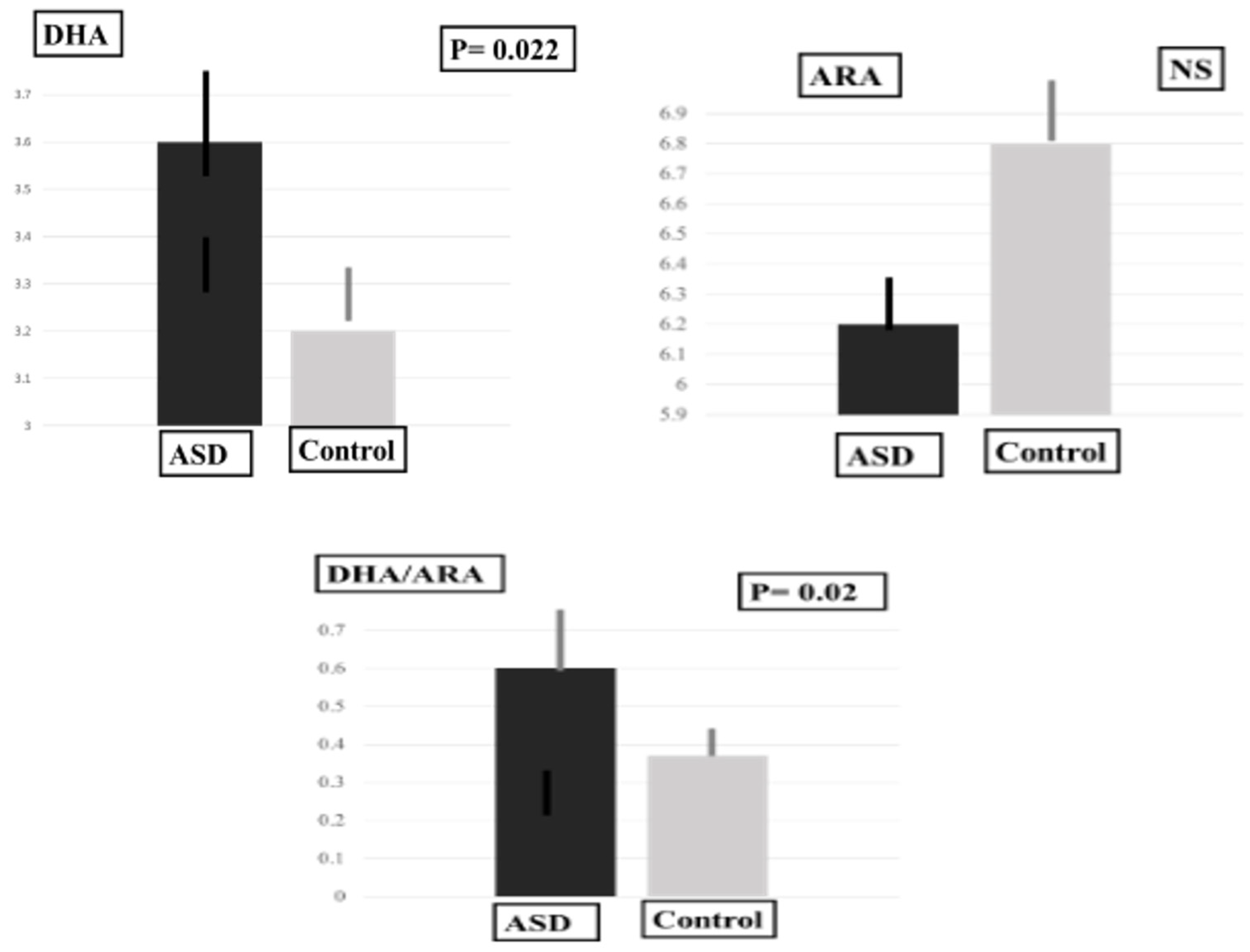
2.3. Plasma Levels of MDA-LDL, SOD and PUFAS
2.4. Gender Difference
2.5. Results of the Multiple Linear Regression Analysis
2.6. Results of the Adaptive Lasso Analysis
2.7. Coefficients of Variation
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Precautions for Mitigating the Effects of Small Sample Size
4.3. Assessment of Social Behaviors
4.4. Controlling for Dietary Intake and Assessment of Nutrient Intake
4.5. Measurement of Plasma PUFA, Cp, SOD and Tf Levels
4.5.1. Blood-Sampling Procedures
4.5.2. Plasma Levels of PUFAs
4.5.3. Plasma Levels of SOD
4.5.4. Plasma Levels of CP
4.5.5. Plasma Levels of TF
4.6. Plasma Levels of MDA-LDL
4.7. Sex Differences in Plasma Variables and the Total SRS Scores
4.8. Statistical Analyses
5. Conclusions
6. Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cao, L.M.; Zhang, X.J.; Chu, B. Targeting ferroptosis as a vulnerability in pulmonary diseases. Cell Death Dis. 2022, 13, 649. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, G.D. Perspective: The Saturated fat-unsaturated oil dilemma: Relations of dietary fatty acids and serum cholesterol, atherosclerosis, inflammation, iancer, and all-cause mortality. Adv. Nutr. 2021, 12, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Alic, L.; Binder, C.J.; Papac-Milicevic, N. The OSE complotype and its clinical potential. Front. Immunol. 2022, 13, 1010893. [Google Scholar] [CrossRef]
- Liu, X.; Lin, J.; Zhang, H.; Khan, N.U.; Zhang, J.; Tang, X.; Cao, X.; Shen, L. Oxidative stress in autism spectrum disorder-current progress of mechanisms and biomarkers. Front. Psychiatry 2022, 13, 813304. [Google Scholar] [CrossRef] [PubMed]
- de Souza, I.; Ramalho, M.C.C.; Guedes, C.B.; Osawa, I.Y.A.; Monteiro, L.K.S.; Gomes, L.R.; Rocha, C.R.R. Ferroptosis Modulation: Potential Therapeutic Target for Glioblastoma Treatment. Int. J. Mol. Sci. 2022, 23, 6879. [Google Scholar] [CrossRef]
- Mortensen, M.S.; Ruiz, J.; Watts, J.L. Polyunsaturated Fatty Acids Drive Lipid Peroxidation during Ferroptosis. Cells 2023, 12, 804. [Google Scholar] [CrossRef] [PubMed]
- Asperti, M.; Bellini, S.; Grillo, E.; Gryzik, M.; Cantamessa, L.; Ronca, R.; Maccarinelli, F.; Salvi, A.; De Petro, G.; Arosio, P.; et al. H-ferritin suppression and pronounced mitochondrial respiration make Hepatocellular Carcinoma cells sensitive to RSL3-induced ferroptosis. Free Radic. Biol. Med. 2021, 169, 294–303. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, Z.; Yan, M.; Zhang, Q.; Jiang, T.; Xue, J. Calycosin decreases cerebral ischemia/reperfusion injury by suppressing ACSL4-dependent ferroptosis. Arch. Biochem. Biophys. 2023, 734, 109488. [Google Scholar] [CrossRef]
- An, J.R.; Su, J.N.; Sun, G.Y.; Wang, Q.F.; Fan, Y.D.; Jiang, N.; Yang, Y.F.; Shi, Y. Liraglutide Alleviates Cognitive Deficit in db/db Mice: Involvement in Oxidative Stress, Iron Overload, and Ferroptosis. Neurochem. Res. 2022, 47, 279–294. [Google Scholar] [CrossRef]
- Wang, Y.; Lin, H.; Huang, W.; Liu, Z.; Chen, Z.; Zhao, X.; Ding, T.; Qin, W.; Shen, Y. Curcumin Attenuates Periodontal Injury via Inhibiting Ferroptosis of Ligature-Induced Periodontitis in Mice. Int. J. Mol. Sci. 2023, 24, 9835. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, S.; Zhou, H.; Li, R.; Xia, X.; Xiong, H. Identification of MTHFD2 as a prognostic biomarker and ferroptosis regulator in triple-negative breast cancer. Front. Oncol. 2023, 13, 1098357. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Nam, M.; Son, H.Y.; Hyun, K.; Jang, S.Y.; Kim, J.W.; Kim, M.W.; Jung, Y.; Jang, E.; Yoon, S.J.; et al. Polyunsaturated fatty acid biosynthesis pathway determines ferroptosis sensitivity in gastric cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 32433–32442. [Google Scholar] [CrossRef]
- Liao, Y.; Cao, P.; Luo, L. Identification of novel arachidonic acid 15-lipoxygenase inhibitors based on the bayesian classifier model and computer-aided high-throughput virtual screening. Pharmaceuticals 2022, 15, 1440. [Google Scholar] [CrossRef] [PubMed]
- Meguid, N.; Dardir, A.A.; Abdel-Raouf, E.R.; Hashish, A. Evaluation of oxidative stress in autism: Defective antioxidant enzymes and increased lipid peroxidation. Biol. Trace Elem. Res. 2011, 143, 58–65. [Google Scholar] [CrossRef]
- González-Fraguela, M.E.; Hung, M.D.; Vera, H.; Maragoto, C.; Noris, E.; Blanco, L.; Galvizu, R.; Robinson, M. Oxidative stress markers in children with autism spectrum disorders. Br. J. Med. Med. Res. 2013, 3, 307–317. [Google Scholar] [CrossRef]
- Mu, H.; Wan, W.; Song, J.; Kuang, R.; Deng, T. Mitochondrial Lipid Peroxidation and Microsomal Drug-metabolizing Enzyme Activity of Rat Hepatotoxicity under Heavy Metals from Slag Waste Exposure. Cell Biochem. Biophys. 2023, 81, 285–298. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2017, 217, 1915–1928. [Google Scholar] [CrossRef]
- Zoroglu, S.S.; Armutcu, F.; Ozen, S.; Gurel, A.; Sivasli, E.; Yetkin, O.; Meram, I. Increased oxidative stress and altered activities of erythrocyte free radical scavenging enzymes in autism. Eur. Arch. Psychiatry Clin. Neurosci. 2004, 254, 143–147. [Google Scholar] [CrossRef]
- Al-Gadani, Y.; El-Ansary, A.; Attas, O.; Al-Ayadhi, L. Metabolic biomarkers related to oxidative stress and antioxidant status in Saudi autistic children. Clin. Biochem. 2009, 42, 1032–1040. [Google Scholar] [CrossRef]
- Altun, H.; Şahin, N.; Kurutaş, E.B.; Karaaslan, U.; Hatice Sevgen, F.; Fındıklı, E. Assessment of malondialdehyde levels, superoxide dismutase, and catalase activity in children with autism spectrum disorders. Psychiatry Clin. Psychopharmacol. 2018, 28, 408–415. [Google Scholar] [CrossRef]
- Liu, L.; Kang, X.X. ACSL4 is overexpressed in psoriasis and enhances inflammatory responses by activating ferroptosis. Biochem. Biophys. Res. Commun. 2022, 623, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.D.; Zhu, A.; Li, L.D.; Zhang, T.; Wang, S.; Shan, D.P.; Li, Y.Z.; Wang, Q. Cytotoxicity and underlying mechanism of evodiamine in HepG2 cells. Beijing Da Xue Xue Bao Yi Xue Ban 2021, 53, 1107–1114. [Google Scholar] [CrossRef]
- Mu, Y.; Sun, J.; Li, Z.; Zhang, W.; Liu, Z.; Li, C.; Peng, C.; Cui, G.; Shao, H.; Du, Z. Activation of pyroptosis and ferroptosis is involved in the hepatotoxicity induced by polystyrene microplastics in mice. Chemosphere 2022, 291 Pt 2, 132944. [Google Scholar] [CrossRef]
- Ornoy, A.; Weinstein-Fudim, L.; Tfilin, M.; Ergaz, Z.; Yanai, J.; Szyf, M.; Turgeman, G. S-adenosyl methionine prevents ASD like behaviors triggered by early postnatal valproic acid exposure in very young mice. Neurotoxicol. Teratol. 2019, 71, 64–74. [Google Scholar] [CrossRef]
- Dziobekm, I.; Gold, S.M.; Wolf, O.T.; Convit, A. Hypercholesterolemia in Asperger syndrome: Independence from lifestyle, obsessive-compulsive behavior, and social anxiety. Psychiatry Res. 2007, 149, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, Y.; Shi, J.; Gao, M.; Liu, Y.; Cong, Y.; Li, Y.; Wang, Y.; Yu, M.; Lu, Y.; et al. Entire peroxidation reaction system of myeloperoxidase correlates with progressive low-density lipoprotein modifications via reactive aldehydes in atherosclerotic patients with hypertension. Cell Physiol. Biochem. 2018, 50, 1245–1254. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Tanaka, T.; Nagoshi, T.; Sekiyama, H.; Arase, S.; Minai, K.; Ogawa, T.; Yoshimura, M. Increase in the oxidised low-density lipoprotein level by smoking and the possible inhibitory effect of statin therapy in patients with cardiovascular disease: A retrospective study. BMJ Open 2015, 5, e005455. [Google Scholar] [CrossRef]
- Shortreed, S.M.; Ertefaie, A. Outcome-adaptive lasso: Variable selection for causal inference. Biometrics 2017, 73, 1111–1122. [Google Scholar] [CrossRef] [PubMed]
- Wahid, A.; Khan, D.M.; Hussain, L. Robust Adaptive Lasso method for parameter’s estimation and variable selection in high-dimensional sparse models. PLoS ONE 2017, 12, e0183518. [Google Scholar] [CrossRef]
- Abdul-Razik Ismail, E. Behavior of lasso quantile regression with small sample sizes. J. Multidiscip. Eng. Sci. Technol. 2015, 2, 388–394. [Google Scholar]
- Singh, K.; Connors, S.L.; Macklin, E.A.; Smith, K.D.; Fahey, J.W.; Talalay, P.; Zimmerman, A.W. Sulforaphane treatment of autism spectrum disorder (ASD). Proc. Nat. Acad. Sci. USA 2014, 111, 15550–15555. [Google Scholar] [CrossRef]
- Ji, Y.; Ji, Y.; Zhu, H.L.; Cheng, S.M.; Zou, X.B.; Zhu, F.L. Examine sex differences in autism spectrum disorder in school-aged children and adolescents with fluent language. Front. Psychiatry 2023, 14, 1151596. [Google Scholar] [CrossRef] [PubMed]
- Baliga, K.; Coggins, A.; Warburton, S.; Mathias, D.; Yamada, N.K.; Fuerch, J.H.; Halamek, L.P. Pilot study of the DART tool—An objective healthcare simulation debriefing assessment instrument. BMC Med. Educ. 2022, 22, 636. [Google Scholar] [CrossRef] [PubMed]
- Fourchet, F.; Materne, O.; Horobeanu, C.; Hudacek, T.; Buchheit, M. Reliability of a novel procedure to monitor the flexibility of lower limb muscle groups in highly-trained adolescent athletes. Phys. Ther. Sport 2013, 14, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Childress, A.C.; Stark, J.G. Diagnosis and treatment of attention-deficit/Hyperactivity disorder in preschool-aged children. J. Child. Adolesc. Psychopharmacol. 2018, 28, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Kim, S.; Chung, H.J.; Lee, W.; Chun, S.; Min, W.K. Evaluation of the MassTrak Immunosuppressant XE kit for the determination of everolimus and cyclosporin A in human whole blood employing isotopically labeled internal standards. Clin. Chem. Lab. Med. 2011, 49, 2021–2027. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.J.; Hecht, P.M.; Jasarevic, E.; Beversdorf, D.Q.; Will, M.J.; Fritsche, K.; Gillespie, C.H. Sex-specific effects of docosahexaenoic acid (DHA) on the microbiome and behavior of socially-isolated mice. Brain Behav. Immun. 2017, 59, 38–48. [Google Scholar] [CrossRef]
- Metherel, A.H.; Irfan, M.; Klingel, S.L.; Mutch, D.M.; Bazinet, R.P. Higher increase in plasma DHA in females compared to males following EPA supplementation may be influenced by a polymorphism in ELOVL2: An exploratory study. Lipids 2021, 56, 211–228. [Google Scholar] [CrossRef]
- Walker, C.G.; Browning, L.M.; Mander, A.P.; Madden, J.; West, A.L.; Calder, P.C.; Jebb, S.A. Age and sex differences in the incorporation of EPA and DHA into plasma fractions, cells and adipose tissue in humans. Br. J. Nutr. 2014, 111, 679–689. [Google Scholar] [CrossRef]
- Cho, S.; Cola, M.; Knox, A.; Pelella, M.R.; Russell, A.; Hauptmann, A.; Covello, M.; Cieri, C.; Liberman, M.; Schultz, R.T.; et al. Sex differences in the temporal dynamics of autistic children’s natural conversations. Mol. Autism 2023, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Usui, N.; Shimada, S. Prenatal sex hormone exposure is associated with the development of autism spectrum disorder. Int. J. Mol. Sci. 2023, 24, 2203. [Google Scholar] [CrossRef] [PubMed]
- Beggiato, A.; Peyre, H.; Maruani, A.; Scheid, I.; Rastam, M.; Amsellem, F.; Gillberg, C.I.; Leboyer, M.; Bourgeron, T.; Gillberg, C.; et al. Gender differences in autism spectrum disorders: Divergence among specific core symptoms. Autism Res. 2017, 10, 680–689. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, O.H.; Oktar, S.; Aydin, M.; Kucukatay, V. Effect of sulfite on antioxidant enzymes and lipid peroxidation in normal and sulfite oxidase-deficient rat erythrocytes. J. Physiol. Biochem. 2010, 66, 205–212. [Google Scholar] [CrossRef]
- Frackowiak, J.l.; Mazur-Kolecka, B.; Schanen, N.C.; Brown, W.T.; Wegiel, J. The link between intraneuronal N-truncated amyloid-β peptide and oxidatively modified lipids in idiopathic autism and dup (15.q11.2-q13)/autism. Acta Neuropathol. Commun. 2013, 1, 61. [Google Scholar] [CrossRef]
- White-Springer, S.H.; Vineyard, K.R.; Kivipelto, J.; Warren, I.K. Dietary omega-3 fatty acid supplementation does not impair vitamin E status or promote lipid peroxidation in growing horses. J. Anim. Sci. 2021, 99, skab177. [Google Scholar] [CrossRef]
- Huun, M.U.; Garberg, H.T.; Escobar, J.; Chafer, C.; Vento, M.; Holme, I.M.; Saugstad, O.D.; Solberg, R. A reduces oxidative stress following hypoxia-ischemia in newborn piglets: A study of lipid peroxidation products in urine and plasma. J. Perinat. Med. 2018, 46, 209–217. [Google Scholar] [CrossRef]
- Yang, W.S.; Kim, K.J.; Gaschler, M.M.; Patel, M.; Shchepinov, M.S.; Stockwell, B.S. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc. Natl. Acad. Sci. USA 2016, 113, E4966–E4975. [Google Scholar] [CrossRef]
- Park, M.W.; Cha, H.W.; Kim, J.; Kim, J.H.; Yang, H.; Yoon, S.; Boonpraman, N.; Yi, S.S.; Yoo, I.D.; Moon, J.S. NOX4 promotes ferroptosis of astrocytes by oxidative stress-induced lipid peroxidation via the impairment of mitochondrial metabolism in Alzheimer’s diseases. Redox Biol. 2021, 41, 101947. [Google Scholar] [CrossRef]
- Afif, I.Y.; Farkhan, M.; Kurdi, O.; Maula, M.I.; Ammarullah, M.I.; Setiyana, B.; Jamari, J.; Winarni, T.I. Effect of Short-Term Deep-Pressure Portable Seat on behavioral and biological stress in children with autismspectrum disorders: A Pilot Study. Boengineering 2022, 9, 48. [Google Scholar] [CrossRef]
- Krombach, T.; Miltenberger, R.J. The effects of stability ball seating on the behavior of children with autism during instructional activities. Autism Dev. Disord. 2020, 50, 551–559. [Google Scholar] [CrossRef]
- Liu, L.; Lai, Y.; Zhan, Z.; Fu, Q.; Jiang, Y.I. Dentification of ferroptosis-related molecular clusters and immune characterization in autism spectrum disorder. Front. Genet. 2022, 13, 911119. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Li, R.; Michael Greenlief, C.; Fritsche, K.L.; Gu, Z.; Cui, J.; Lee, J.C.; Beversdorf, D.Q.; Sun, G.Y. Yin-Yang mechanisms regulating lipid peroxidation of docosahexaenoic acid and arachidonic acid in the central nervous system. Front. Neurol. 2019, 10, 642. [Google Scholar] [CrossRef] [PubMed]
- Marmunti, M.; Catalá, A. Arachidonic acid hydroperoxide stimulates lipid peroxidation in rat liver nuclei and chromatin fractions. Mol. Cell. Biochem. 2007, 298, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Sakayori, N.; Kikkawa, T.; Tokuda, H.; Kiryu, E.; Yoshizaki, K.; Kawashima, H.; Yamada, T.; Arai, H.; Kang, J.X.; Katagir, H.; et al. Maternal dietary imbalance between omega-6 and omega-3 polyunsaturated fatty acids impairs neocortical development via epoxy metabolites. Stem Cells 2016, 34, 470–482. [Google Scholar] [CrossRef]
- Hoffman, D.R.; Boettcher, J.A.; Diersen-Schade, D.A. Toward optimizing vision and cognition in term infants by dietary docosahexaenoic and arachidonic acid supplementation: A review of randomized controlled trials. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 151–158. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, Y.; Duan, R.; Wei, H.; Jiang, S.; Peng, J. Lower dietary n-6: N-3 ratio and high-dose vitamin E supplementation improve sperm morphology and oxidative stress in boars. Reprod. Fertil. Dev. 2017, 29, 940–949. [Google Scholar] [CrossRef]
- McAnulty, S.R.; Nieman, D.C.; Fox-Rabinovich, M.; Duran, V.; McAnulty, L.S.; Henson, D.A.; Jin, F.; Landram, M. Effect of n-3 fatty acids and antioxidants on oxidative stress after exercise. J. Med. Sci. Sports Exerc. 2010, 42, 1704–1711. [Google Scholar] [CrossRef]
- Schuchardt, J.P.; Huss, M.; Stauss-Grabo, M.; Hahn, A. Significance of long-chain polyunsaturated fatty acids (PUFAs) for the development and behaviour of children. Eur. J. Pediatr. 2010, 169, 149–164. [Google Scholar] [CrossRef]
- Sarajlic, P.; Vigor, C.; Avignon, A.; Zhou, B.; Oger, C.; Galano, J.M.; Durand, T.; Sultan, A.; Bäck, M. Omega-3 to omega-6 fatty acid oxidation ratio as a novel inflammation resolution marker for metabolic complications in obesity. Nutr. Metab. Cardiovasc. Dis. 2023, 33, 1206–1213. [Google Scholar] [CrossRef]
- Li, J.; Jia, B.; Cheng, Y.; Song, Y.; Li, Q.; Luo, C. Targeting molecular mediators of ferroptosis and oxidative stress for neurological disorders. Oxid. Med. Cell. Longev. 2022, 2022, 3999083. [Google Scholar] [CrossRef] [PubMed]
- Dierge, E.; Debock, E.; Guilbaud, C.; Corbet, C.; Mignolet, E.; Mignard, L.; Bastien, E.; Dessy, C.; Larondelle, Y.; Feron, O. Peroxidation of n-3 and n-6 polyunsaturated fatty acids in the acidic tumor environment leads to ferroptosis-mediated anticancer effects. Cell Metab. 2021, 33, 1701–1715. [Google Scholar] [CrossRef] [PubMed]
- Delesderrier, E.; Monteiro, J.D.C.; Freitas, S.; Pinheiro, I.C.; Batista, M.S.; Citelli, M. Can iron and polyunsaturated fatty acid supplementation induce ferroptosis? Cell Physiol. Biochem. 2023, 57, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Brouwers, H.; Jónasdóttir, H.S.; Kuiper, M.E.; Kwekkeboom, J.C.; Auger, J.L.; Gonzalez-Torres, M.; López-Vicario, C.; Clària, J.; Freysdottir, J.; Hardardottir, I.; et al. Anti-inflammatory and proresolving effects of the omega-6 polyunsaturated fatty acid adrenic acid. J. Immunol. 2020, 205, 2840–2849. [Google Scholar] [CrossRef]
- Horas, H.; Nababa, S.; Nishiumi, S.; Kawano, Y.; Kobayashi, T.; Yoshida, M.; Azuma, T. Adrenic acid as an inflammation enhancer in non-alcoholic fatty liver disease. Arch. Biochem. Biophys. 2017, 623–624, 64–75. [Google Scholar] [CrossRef]
- Greene, R.K.; Walsh, E.; Mosner, M.G.; Dichter, G.S. A potential mechanistic role for neuroinflammation in reward processing impairments in autism spectrum disorder. Biol. Psychol. 2019, 142, 1–12. [Google Scholar] [CrossRef]
- Yui, K.; Tanuma, N.; Yamada, H.; Kawasaki, Y. Decreased total antioxidant capacity has a larger effect size than increased oxidant levels in urine in individuals with autism spectrum disorder. Environ. Sci. Pollut. Res. Int. 2017, 24, 9635–9644. [Google Scholar] [CrossRef]
- Hashimoto, M.; Hossain, S.; Tanabe, Y.; Kawashima, A.; Harada, T.; Yano, T.; Mizuguchi, K.; Shido, O. The protective effect of dietary eicosapentaenoic acid against impairment of spatial cognition learning ability in rats infused with amyloid beta(1-40). J. Nutr. Biochem. 2009, 20, 965–973. [Google Scholar] [CrossRef]
- Davis-Bruno, K.; Tassinari, M.S. Essential fatty acid supplementation of DHA and ARA and effects on neurodevelopment across animal species: A review of the literature. Birth Defects Res. B Dev. Reprod. Toxicol. 2011, 92, 240–250. [Google Scholar] [CrossRef]
- Shahmirzai, R.; Firouzifar, M. Sleep apnea and idiopathic leg edema: A case control study. Acta Med. Iran 2017, 55, 612–615. [Google Scholar]
- Nieradko-Iwanicka, B.; Przybylska, D.; Borzęcki, A. A case-control study on immunologic markers of patients with vitiligo. Biomed. Pharmacother. 2022, 156, 113785. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Gomez, A.; Aguilera, P.; Langohr, K.; Casals, G.; Pavon, C.; Marcos, J.; To-Figueras, J.; Pozo, O. Evaluation of metabolic changes in acute intermittent porphyria patients by targeted metabolomics. Int. J. Mol. Sci. 2022, 23, 3219. [Google Scholar] [CrossRef]
- Moll, I.; Voorman, J.M.; Ketelaar, M.; van Schie, P.E.; Gorter, J.W.; Lequin, M.H.; de Vries, L.S.; Vermeulen, R.J. Prognostic value of brain abnormalities for cognitive functioning in cerebral palsy: A prospective cohort study. Eur. J. Paediatr. Neurol. 2021, 32, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Packiasabapathy, S.; Prasad, V.; Rangasamy, V.; Popok, D.; Xu, X.; Novack, V.; Subramaniam, B. Cardiac surgical outcome prediction by blood pressure variability indices Poincaré plot and coefficient of variation: A retrospective study. BMC Anesthesiol. 2020, 20, 56. [Google Scholar] [CrossRef] [PubMed]
- Lichou, F.; Orazio, S.; Dulucq, S.; Etienne, G.; Longy, M.; Hubert, C.; Groppi, A.; Monnereau, A.; Mahon, F.X.; Turcq, B. Novel analytical methods to interpret large sequencing data from small sample sizes. Hum. Genomic. 2019, 13, 41. [Google Scholar] [CrossRef]
- Constantino, J.H.; Gruber, C.P. Social Responsiveness Scale-Second Edition (SRS-2). In Western Psychological Services; CAR Autism Roadmap™: Torrance, CA, USA, 2012. [Google Scholar]
- Sipsock, D.; Tokadjian, H.; Righi, G.; Morrow, E.M.; Sheinkopf, S.J.; Rhode Island Consortium for Autism Research and Treatment (RI-CART). Autism severity aggregates with family psychiatric history in a community-based autism sample. Autism Res. 2021, 14, 2524–2532. [Google Scholar] [CrossRef]
- Zheng, J.S.; Imamura, F.; Sharp, S.J.; Koulman, A.; Griffin, J.L.; Mulligan, A.A.; Luben, R.; Khaw, K.T.; Wareham, N.J.; Forouhi, N.G. Changes in plasma phospholipid fatty acid profiles over 13 years and correlates of change: European Prospective Investigation into Cancer and Nutrition-Norfolk Study. Am. J. Clin. Nutr. 2019, 109, 1527–1534. [Google Scholar] [CrossRef]
- Okuda, M.; Sasaki, S.; Bando, N.; Hashimoto, M.; Kunitsugu, I.; Sugiyama, S.; Terao, J.; Hobara, T. Carotenoid, tocopherol, and fatty acid biomarkers and dietary intake estimated by using a brief self-administered diet history questionnaire for older Japanese children and adolescents. J. Nutr. Sci. 2009, 55, 231–241. [Google Scholar] [CrossRef]
- Kobayashi, S.; Honda, S.; Murakami, K.; Sasaki, S.; Okubo, H.; Hirota, N.; Notsu, A.; Fukui, M.; Date, C. Both comprehensive and brief self-administered diet history questionnaires satisfactorily rank nutrient intakes in Japanese adults. J. Epidemiol. 2012, 22, 151–159. [Google Scholar] [CrossRef]
- Simundi, A.M.; Cornes, M.; Grankvis, K.; Lippi, G.; Nybo, M. Standardization of collection requirements for fasting samples For the Working Group on Preanalytical Phase (WG-PA) of the European Federation of Clinical Chemistry and Laboratory Medicine (EFLM). Clin. Chim. Acta 2014, 432, 33037. [Google Scholar] [CrossRef]
- Stalenhoef, A.F.; de Graaf, J. Association of fasting and nonfasting serum triglycerides with cardiovascular disease and the role of remnant-like lipoproteins and small dense LDL. Curr. Opin. Lipido. 2008, 19, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Li, S.; Bi, X.; Yuan, R.; Xiang, Y. Multicolor-Encoded DNA Framework Enables Specific and Amplified In Situ Detection of the Mitochondrial Apoptotic Signaling Pathway. Anal. Chem. 2023, 95, 12514–12520. [Google Scholar] [CrossRef] [PubMed]
- Kitano, S.; Kanno, T.; Maekawa, M.; Sakiurabayashi, I.; Kotani, K.; Hisatomi, H. Improved method for the immunological detection of malondialdehyde-modified low-density lipoproteins in human serum. Anal. Chim. Acta 2004, 509, 229–235. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Variables | ASD (n = 18) | Control (n = 8) | U | p Values | |
|---|---|---|---|---|---|
| Age (Years) | 10.9 ± 5.6 | 9.6 ± 4.0 | 63 | 0.64 | |
| Sex (Male/Female) | 12/6 | 5/3 | χ2 = 0.4 | 0.84 | |
| Subject Characteristics | Domain A | 24.4 ± 3.7 | NA | ||
| Domain B (Communication) | 12.0 ± 2.3 | NA | |||
| Domain C (Stereotyped Behaviors) | 10.8 ± 3.5 | NA | |||
| Antioxidant | Cp (mg·dL) | 28.17 ± 7.02 | 24.63 ± 6.78 | 55.00 | 0.37 |
| Tf (mg/dL) | 275.39 ± 41.89 | 261.25 ± 24.06 | 56.00 | 0.40 * | |
| SOD (U·mL) | 2.53 ± 0.46 | 5.36 ± 4.39 | 12.50 | 0.003 * | |
| Biomarkers | MDA-LDL | 91.00 ± 16.70 | 74.50 ± 18.88 | 36.50 | 0.047 * |
| DHA | 3.57 ± 0.98 | 3.25 ± 0.84 | 31.50 | 0.022 * | |
| ARA | 6.23 ± 1.08 | 6.79 ± 1.58 | 53.00 | 0.31 | |
| DHA/ARA | 0.60 ± 0.16 | 0.37 ± 0.07 | 10.50 | 0.02 * | |
| Adrenic Acid | 0.26 ± 0.25 | 0.27 ± 0.06 | 32.00 | 0.03 * |
| Model | Model | Model | Coefficients | |||
|---|---|---|---|---|---|---|
| R2 | p-Value | B | Beta | p Value | ||
| Coefficients | ||||||
| DHA | 0.991 | 0.000 ** | ||||
| α-linolenic acid | 0.535 ± 0.155 | 0.107 | 0.013 * | |||
| DPA | 0.498 ± 0.193 | 0.088 | 0.04 * | |||
| GLA | −1.166 ± 0.306 | −0.157 | 0.007 * | |||
| SOD | −0.06 ± 0.015 | −0.170 | 0.03 * | |||
| SRS total | 0.000009 ± 0.001 | 0.092 | 0.057 | |||
| Group (1 = ASD, 2 = control) | −0.057 ± 0.102 | −0.26 | 0.591 | |||
| DHA/ARA | 0.972 | 0.000 ** | ||||
| ARA | −0.057 ± 0.102 | −0.541 | 0.01 * | |||
| α-linolenic acid | −0.267 ± 0.102 | −0.316 | 0.04 * | |||
| Adrenic acid | −0.070 ± 0.147 | −0.037 | 0.650 | |||
| SOD | 0.016 ± 0.015 | 0.242 | 0.326 | |||
| SRS total score | 0.000009 ± 0.001 | −0.023 | 0.901 | |||
| Group (1 = ASD, 2 = control) | 0.020 ± 0.057 | 0.05 | 0.731 |
| Variables | Standardized Coefficients | SE | 95% CL | p Value | |
|---|---|---|---|---|---|
| Lower Bound | Upper Bound | ||||
| SRS total scores | |||||
| DHA/ARA | 61.155 | 27.353 | 7.546 | 114.776 | 0.125 |
| MDA-LDL | |||||
| DHA/ARA | 5.633 | 15.91 | −25.572 | 36.837 | 0.712 |
| SOD | −40.060 | 9.935 | −0.135 | −31.186 | <0.001 |
| ASD | Control | U | p Value | ||
|---|---|---|---|---|---|
| (n = 7) | (n = 5) | ||||
| Age (years) | 11.4 ± 4.3 | 11.4 ± 3.2 | 14.0 | 0.87 | |
| Fat (g/day) | 72.2 ± 30.1 | 87.4 ± 25.8 | 12.0 | 0.43 | |
| Unsaturated | 14.8 ± 4.4 | 18.7 ± 5.5 | 9.0 | 0.20 | |
| Fatty acid (g/day) | |||||
| PUFAs | Omega-3 PUFAs (g/day) | 2.6 ± 0.8 | 3.1 ± 0.5 | 9.0 | 0.20 |
| Omega-6 PUFAs (g/day) | 12.1 ± 3.9 | 15.9 ± 5.1 | 11.0 | 0.34 | |
| EPA (mg/day) | 181.2 ± 118.7 | 176.2 ± 73.6 | 15.5 | 0.76 | |
| DHA (mg/day) | 332.6 ± 170.4 | 345.0 ± 83.31 | 15.5 | 0.76 | |
| ARA (mg/day) | 168.1 ± 17.1 | 221.0 ± 87.7 | 11.5 | 0.34 | |
| Protein (g/day) | 78.1 ± 25.8 | 89.2 ± 25.8 | 13.0 | 0.53 | |
| Animal protein (mg/day) | 32.0 ± 9.1 | 30.4 ± 14.7 | 14.0 | 0.64 | |
| Cholesterol (mg/day) | 139.1± 186.4 | 31.9 ± 10.3 | 17.0 | 1.00 | |
| Carbohydrates (g/day) | 286.2 ± 62.1 | 304.21 ± 72.4 | 14.0 | 0.64 | |
| Copper (mg/day) | 1.0 ± 0.2 | 1.3 ± 0.5 | 7.0 | 0.56 | |
| Iron (mg·day) | 7.4 ±1.6 | 9.2 ± 3.5 | 6.0 | 0.41 | |
| MDA-LDL | DHA | ARA | DHA/ARA | Adrenic | SOD | Total SRS | |
|---|---|---|---|---|---|---|---|
| Acid | Scores | ||||||
| Male (n = 12) | 97.17 ± 17.06 | 3.85 ±1.07 | 6.16 ± 0.65 | 0.62 ± 0.16 | 0.27 ± 0.30 | 2.59 ± 0.42 | 89.03 ± 30.18 |
| Female (n = 6) | 88.67 ±17.36 | 3.00 ± 0.60 | 6.37 ± 1.72 | 0.56 ± 0.17 | 0.24 ± 0.13 | 2.40 ± 0.56 | 70.50 ± 44.36 |
| U | 34.50 | 15.50 | 33.00 | 25.50 | 33.00 | 28.50 | 24.00 |
| p values | 0.89 | 0.053 | 0.82 | 0.34 | 0.82 | 0.49 | 0.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yui, K.; Imataka, G.; Shiohama, T. Lipid Peroxidation of the Docosahexaenoic Acid/Arachidonic Acid Ratio Relating to the Social Behaviors of Individuals with Autism Spectrum Disorder: The Relationship with Ferroptosis. Int. J. Mol. Sci. 2023, 24, 14796. https://doi.org/10.3390/ijms241914796
Yui K, Imataka G, Shiohama T. Lipid Peroxidation of the Docosahexaenoic Acid/Arachidonic Acid Ratio Relating to the Social Behaviors of Individuals with Autism Spectrum Disorder: The Relationship with Ferroptosis. International Journal of Molecular Sciences. 2023; 24(19):14796. https://doi.org/10.3390/ijms241914796
Chicago/Turabian StyleYui, Kunio, George Imataka, and Tadashi Shiohama. 2023. "Lipid Peroxidation of the Docosahexaenoic Acid/Arachidonic Acid Ratio Relating to the Social Behaviors of Individuals with Autism Spectrum Disorder: The Relationship with Ferroptosis" International Journal of Molecular Sciences 24, no. 19: 14796. https://doi.org/10.3390/ijms241914796
APA StyleYui, K., Imataka, G., & Shiohama, T. (2023). Lipid Peroxidation of the Docosahexaenoic Acid/Arachidonic Acid Ratio Relating to the Social Behaviors of Individuals with Autism Spectrum Disorder: The Relationship with Ferroptosis. International Journal of Molecular Sciences, 24(19), 14796. https://doi.org/10.3390/ijms241914796