
MicroRNAs Present in Malignant Pleural Fluid Increase the Migration of Normal Mesothelial Cells In Vitro and May Help Discriminate between Benign and Malignant Effusions

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Study Population
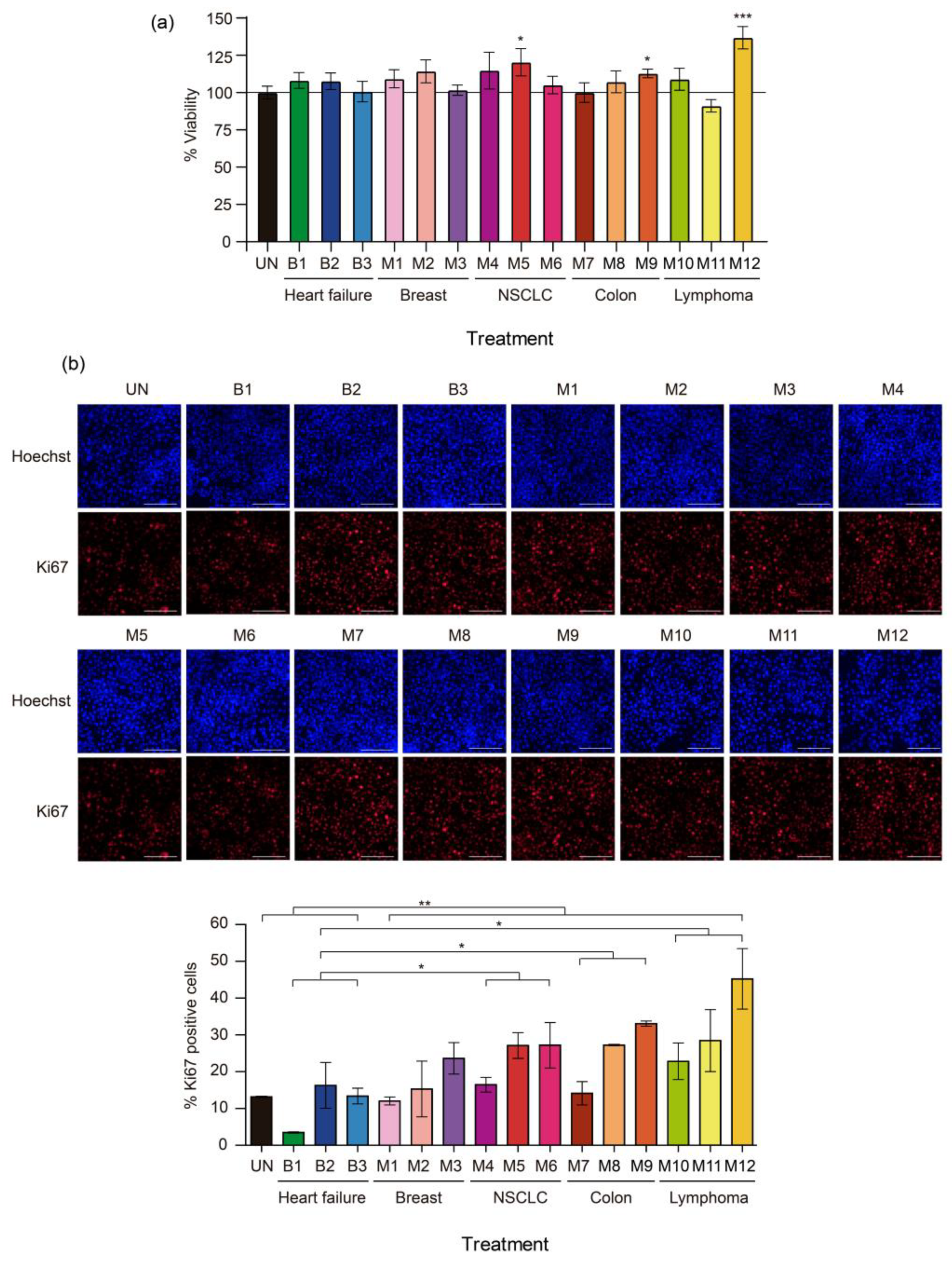
2.2. Effect of Pleural Fluid on the Viability of MeT-5A Cells
2.3. Effect of Pleural Fluid on the Proliferation of MeT-5A Cells
2.4. Effects of Pleural Fluid on the Migration of MeT-5A Cells
2.5. Effects of Pleural Fluid on the Phosphorylation of Proteins That Participate in Cell Survival-and Proliferation-Related Signaling Pathways
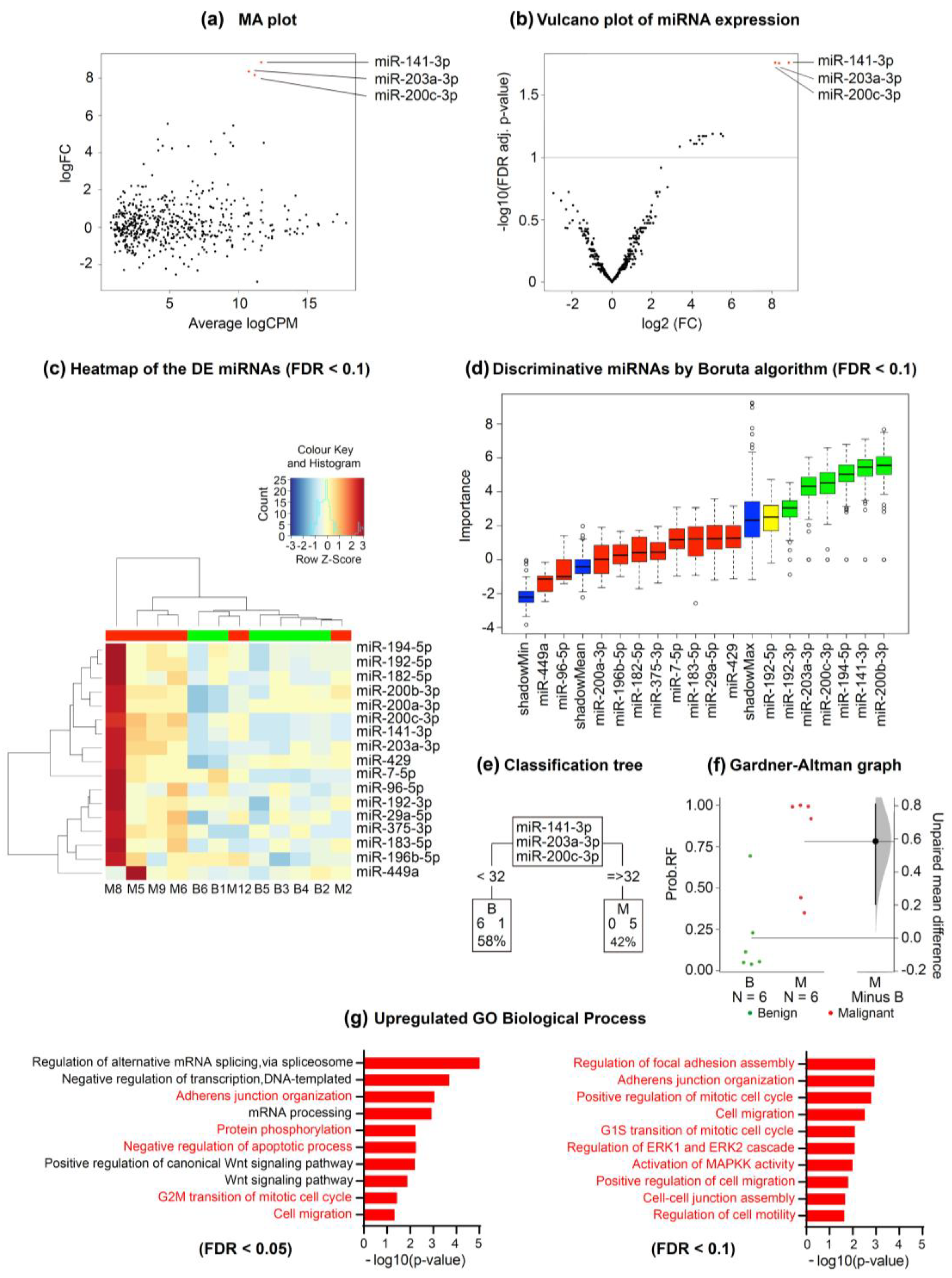
2.6. Whole Exosome-miRNA Sequencing to Discriminate Benign from Malignant Pleural Fluid Samples
3. Discussion
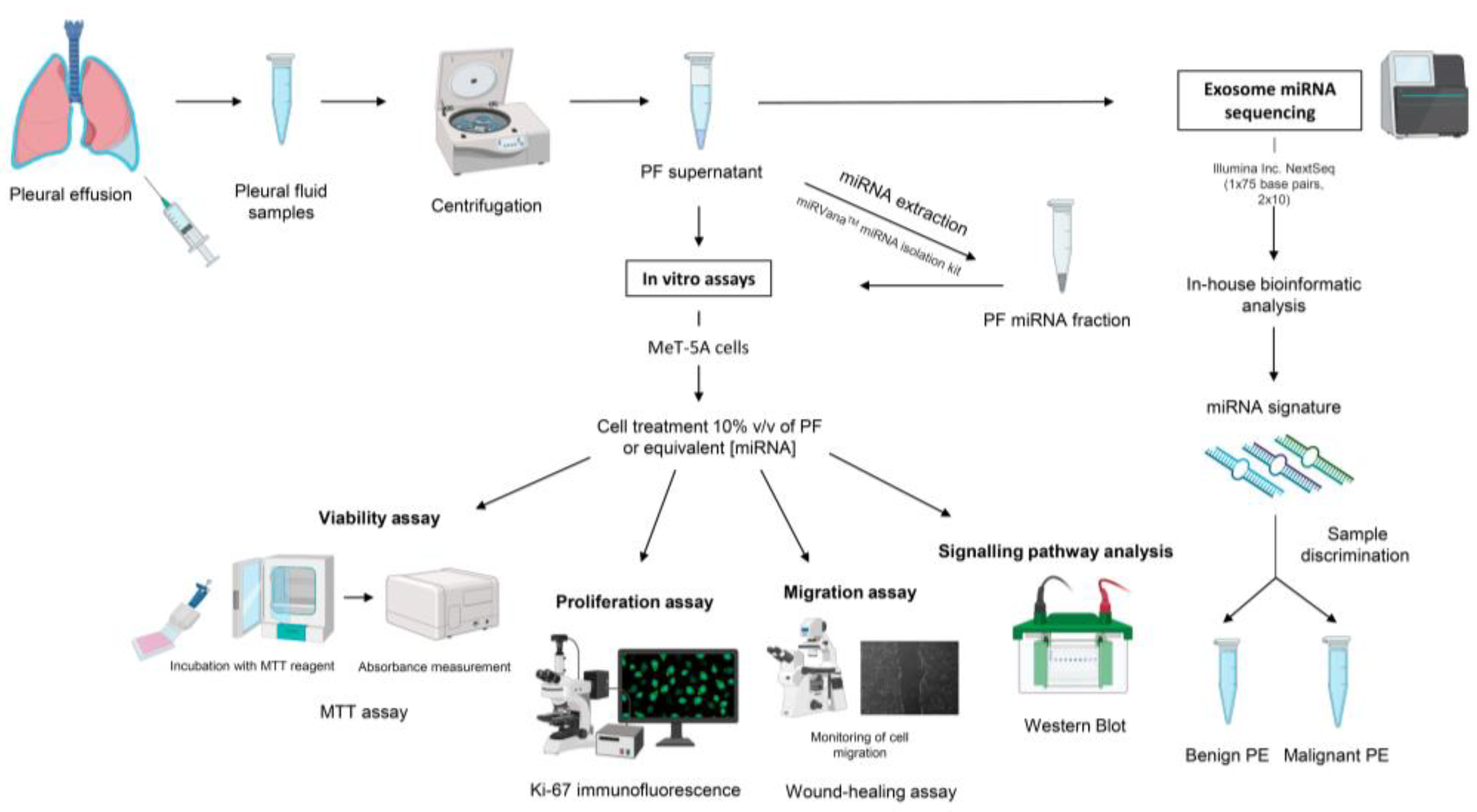
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
4.2. Pleural Fluid Samples
4.3. Total miRNA Isolation
4.4. Functional Assays and Protein Expression Analysis by Western Blotting
4.5. Exosome-miRNA Sequencing
4.6. Real Time-Quantitative Polymerase Chain Reaction
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Psallidas, I.; Kalomenidis, I.; Porcel, J.M.; Robinson, B.W.; Stathopoulos, G.T. Malignant pleural effusion: From bench to bedside. Eur. Respir. Rev. 2016, 25, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Porcel, J.M.; Esquerda, A.; Vives, M.; Bielsa, S. Etiology of pleural effusions: Analysis of more than 3000 consecutive thoracenteses. Arch. Bronconeumol. 2014, 50, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Massague, J.; Obenauf, A.C. Metastatic colonization by circulating tumour cells. Nature 2016, 529, 298–306. [Google Scholar] [CrossRef]
- Stathopoulos, G.T.; Kalomenidis, I. Malignant pleural effusion: Tumor-host interactions unleashed. Am. J. Respir. Crit. Care Med. 2012, 186, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Stathopoulos, G.T.; Zhu, Z.; Everhart, M.B.; Kalomenidis, I.; Lawson, W.E.; Bilaceroglu, S.; Peterson, T.E.; Mitchell, D.; Yull, F.E.; Light, R.W.; et al. Nuclear factor-kappaB affects tumor progression in a mouse model of malignant pleural effusion. Am. J. Respir. Cell Mol. Biol. 2006, 34, 142–150. [Google Scholar] [CrossRef]
- Stathopoulos, G.T.; Kollintza, A.; Moschos, C.; Psallidas, I.; Sherrill, T.P.; Pitsinos, E.N.; Vassiliou, S.; Karatza, M.; Papiris, S.A.; Graf, D.; et al. Tumor necrosis factor-alpha promotes malignant pleural effusion. Cancer Res. 2007, 67, 9825–9834. [Google Scholar] [CrossRef]
- Hooper, C.E.; Elvers, K.T.; Welsh, G.I.; Millar, A.B.; Maskell, N.A. VEGF and sVEGFR-1 in malignant pleural effusions: Association with survival and pleurodesis outcomes. Lung Cancer 2012, 77, 443–449. [Google Scholar] [CrossRef]
- Thomas, R.; Cheah, H.M.; Creaney, J.; Turlach, B.A.; Lee, Y.C. Longitudinal Measurement of Pleural Fluid Biochemistry and Cytokines in Malignant Pleural Effusions. Chest 2016, 149, 1494–1500. [Google Scholar] [CrossRef]
- Chen, Y.M.; Yang, W.K.; Whang-Peng, J.; Kuo, B.I.; Perng, R.P. Elevation of interleukin-10 levels in malignant pleural effusion. Chest 1996, 110, 433–436. [Google Scholar] [CrossRef]
- Yeh, H.H.; Lai, W.W.; Chen, H.H.; Liu, H.S.; Su, W.C. Autocrine IL-6-induced Stat3 activation contributes to the pathogenesis of lung adenocarcinoma and malignant pleural effusion. Oncogene 2006, 25, 4300–4309. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA signatures in human cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Esquela-Kerscher, A.; Slack, F.J. Oncomirs—microRNAs with a role in cancer. Nat. Rev. Cancer 2006, 6, 259–269. [Google Scholar] [CrossRef]
- Calin, G.A.; Liu, C.G.; Sevignani, C.; Ferracin, M.; Felli, N.; Dumitru, C.D.; Shimizu, M.; Cimmino, A.; Zupo, S.; Dono, M.; et al. MicroRNA profiling reveals distinct signatures in B cell chronic lymphocytic leukemias. Proc. Natl. Acad. Sci. USA 2004, 101, 11755–11760. [Google Scholar] [CrossRef] [PubMed]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef]
- Roldo, C.; Missiaglia, E.; Hagan, J.P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G.A.; Volinia, S.; Liu, C.G.; Scarpa, A.; et al. MicroRNA expression abnormalities in pancreatic endocrine and acinar tumors are associated with distinctive pathologic features and clinical behavior. J. Clin. Oncol. 2006, 24, 4677–4684. [Google Scholar] [CrossRef]
- Sorolla, M.A.; Sorolla, A.; Parisi, E.; Salud, A.; Porcel, J.M. Diving into the Pleural Fluid: Liquid Biopsy for Metastatic Malignant Pleural Effusions. Cancers 2021, 13, 2798. [Google Scholar] [CrossRef]
- Zhai, K.; Shi, X.Y.; Yi, F.S.; Huang, Z.Y.; Wu, X.Z.; Dong, S.F.; Wang, W.; Wu, M.T.; Shi, H.Z. IL-10 promotes malignant pleural effusion by regulating TH 1 response via an miR-7116-5p/GPR55/ERK pathway in mice. Eur. J. Immunol. 2020, 50, 1798–1809. [Google Scholar] [CrossRef]
- Qian, Q.; Sun, W.; Zhu, W.; Liu, Y.; Ge, A.; Ma, Y.; Zhang, Y.; Zeng, X.; Huang, M. The role of microRNA-93 regulating angiopoietin2 in the formation of malignant pleural effusion. Cancer Med. 2017, 6, 1036–1048. [Google Scholar] [CrossRef]
- Lv, M.; Xu, Y.; Tang, R.; Ren, J.; Shen, S.; Chen, Y.; Liu, B.; Hou, Y.; Wang, T. miR141-CXCL1-CXCR2 signaling-induced Treg recruitment regulates metastases and survival of non-small cell lung cancer. Mol. Cancer Ther. 2014, 13, 3152–3162. [Google Scholar] [CrossRef]
- Porcel, J.M. Diagnosis and characterization of malignant effusions through pleural fluid cytological examination. Curr. Opin. Pulm. Med. 2019, 25, 362–368. [Google Scholar] [CrossRef]
- Vazquez, F.; Ramaswamy, S.; Nakamura, N.; Sellers, W.R. Phosphorylation of the PTEN tail regulates protein stability and function. Mol. Cell Biol. 2000, 20, 5010–5018. [Google Scholar] [CrossRef]
- Asciak, R.; Kanellakis, N.I.; Yao, X.; Abd Hamid, M.; Mercer, R.M.; Hassan, M.; Bedawi, E.O.; Dobson, M.; Fsadni, P.; Montefort, S.; et al. Pleural Fluid Has Pro-Growth Biological Properties Which Enable Cancer Cell Proliferation. Front. Oncol. 2021, 11, 658395. [Google Scholar] [CrossRef]
- Cheah, H.M.; Lansley, S.M.; Varano Della Vergiliana, J.F.; Tan, A.L.; Thomas, R.; Leong, S.L.; Creaney, J.; Lee, Y.C. Malignant pleural fluid from mesothelioma has potent biological activities. Respirology 2017, 22, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Gartel, A.L.; Radhakrishnan, S.K. Lost in transcription: p21 repression, mechanisms, and consequences. Cancer Res. 2005, 65, 3980–3985. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; Wang, G.; He, S.; Shen, G.; Su, C.; Zhang, Y.; Wei, X.; Ye, T.; Li, L.; Yang, S.; et al. Malignant Pleural Effusion and ascites Induce Epithelial-Mesenchymal Transition and Cancer Stem-like Cell Properties via the Vascular Endothelial Growth Factor (VEGF)/Phosphatidylinositol 3-Kinase (PI3K)/Akt/Mechanistic Target of Rapamycin (mTOR) Pathway. J. Biol. Chem. 2016, 291, 26750–26761. [Google Scholar] [CrossRef] [PubMed]
- Gary Lee, Y.C.; Melkerneker, D.; Thompson, P.J.; Light, R.W.; Lane, K.B. Transforming growth factor beta induces vascular endothelial growth factor elaboration from pleural mesothelial cells in vivo and in vitro. Am. J. Respir. Crit. Care Med. 2002, 165, 88–94. [Google Scholar] [CrossRef]
- Maki, Y.; Nishimura, Y.; Toyooka, S.; Soh, J.; Tsukuda, K.; Shien, K.; Furukawa, M.; Muraoka, T.; Ueno, T.; Tanaka, N.; et al. The proliferative effects of asbestos-exposed peripheral blood mononuclear cells on mesothelial cells. Oncol. Lett. 2016, 11, 3308–3316. [Google Scholar] [CrossRef]
- Stathopoulos, G.T.; Sherrill, T.P.; Karabela, S.P.; Goleniewska, K.; Kalomenidis, I.; Roussos, C.; Fingleton, B.; Yull, F.E.; Peebles, R.S., Jr.; Blackwell, T.S. Host-derived interleukin-5 promotes adenocarcinoma-induced malignant pleural effusion. Am. J. Respir. Crit. Care Med. 2010, 182, 1273–1281. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- De Pascalis, C.; Etienne-Manneville, S. Single and collective cell migration: The mechanics of adhesions. Mol. Biol. Cell 2017, 28, 1833–1846. [Google Scholar] [CrossRef] [PubMed]
- Sedgwick, A.E.; D’Souza-Schorey, C. Wnt Signaling in Cell Motility and Invasion: Drawing Parallels between Development and Cancer. Cancers 2016, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Jacobson, K.; Schaller, M.D. MAP kinases and cell migration. J. Cell Sci. 2004, 117, 4619–4628. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, Y.M.; Zou, Y.Q.; Lin, J.; Huang, B.; Liu, J.; Li, J.; Zhang, J.; Yang, W.M.; Min, Q.H.; et al. Identification of differential expressed PE exosomal miRNA in lung adenocarcinoma, tuberculosis, and other benign lesions. Medicine 2017, 96, e8361. [Google Scholar] [CrossRef]
- Mitra, A.; Yoshida-Court, K.; Solley, T.N.; Mikkelson, M.; Yeung, C.L.A.; Nick, A.; Lu, K.; Klopp, A.H. Extracellular vesicles derived from ascitic fluid enhance growth and migration of ovarian cancer cells. Sci. Rep. 2021, 11, 9149. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.Y.; Liu, Y.N.; Wu, S.G.; Hsu, C.L.; Chang, T.H.; Tsai, M.F.; Lin, Y.T.; Shih, J.Y. MiR-200c-3p suppression is associated with development of acquired resistance to epidermal growth factor receptor (EGFR) tyrosine kinase inhibitors in EGFR mutant non-small cell lung cancer via a mediating epithelial-to-mesenchymal transition (EMT) process. Cancer Biomark. 2020, 28, 351–363. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, Y.; Li, Q.; Xu, R.; Huang, N. MiR-200c-3p and miR-485-5p overexpression elevates cisplatin sensitivity and suppresses the malignant phenotypes of non-small cell lung cancer cells through targeting RRM2. Thorac. Cancer 2022, 13, 1974–1985. [Google Scholar] [CrossRef]
- Kang, E.; Jung, S.C.; Nam, S.K.; Park, Y.; Seo, S.H.; Park, K.U.; Oh, H.K.; Kim, D.W.; Kang, S.B.; Lee, H.S. Tissue miR-200c-3p and circulating miR-1290 as potential prognostic biomarkers for colorectal cancer. Sci. Rep. 2022, 12, 2295. [Google Scholar] [CrossRef]
- Navarro-Manzano, E.; Luengo-Gil, G.; Gonzalez-Conejero, R.; Garcia-Garre, E.; Garcia-Martinez, E.; Garcia-Torralba, E.; Chaves-Benito, A.; Vicente, V.; Ayala de la Pena, F. Prognostic and Predictive Effects of Tumor and Plasma miR-200c-3p in Locally Advanced and Metastatic Breast Cancer. Cancers 2022, 14, 2390. [Google Scholar] [CrossRef]
- Nersisyan, S.; Gorbonos, A.; Makhonin, A.; Zhiyanov, A.; Shkurnikov, M.; Tonevitsky, A. isomiRTar: A comprehensive portal of pan-cancer 5′-isomiR targeting. PeerJ 2022, 10, e14205. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Z.; Yang, Y.; Luo, M.; Zhang, M.; Wang, X.; Liu, L.; Hou, N.; Guo, Q.; Song, T.; et al. MiR-99b-5p and miR-203a-3p Function as Tumor Suppressors by Targeting IGF-1R in Gastric Cancer. Sci. Rep. 2018, 8, 10119. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Gong, L.; Mou, Y.; Han, Y.; Zheng, S. MicroRNA-203a-3p is a candidate tumor suppressor that targets thrombospondin 2 in colorectal carcinoma. Oncol. Rep. 2019, 42, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Z.; Shao, C.C.; Wang, X.J.; Zhao, X.; Chen, J.Q.; Ouyang, Y.X.; Feng, J.; Zhang, F.; Huang, W.H.; Ying, Q.; et al. circTADA2As suppress breast cancer progression and metastasis via targeting miR-203a-3p/SOCS3 axis. Cell Death Dis. 2019, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Lan, Q.; Huang, Y.; Zhang, Y.; Zeng, Y.; Su, P.; Chu, Z.; Lai, W.; Chu, Z. The mechanisms of colorectal cancer cell mesenchymal-epithelial transition induced by hepatocyte exosome-derived miR-203a-3p. BMC Cancer 2021, 21, 718. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gao, H.; Liang, J.; Qiao, J.; Duan, J.; Shi, H.; Zhen, T.; Li, H.; Zhang, F.; Zhu, Z.; et al. miR-203a-3p promotes colorectal cancer proliferation and migration by targeting PDE4D. Am. J. Cancer Res. 2018, 8, 2387–2401. [Google Scholar] [PubMed]
- Aakko, S.; Straume, A.H.; Birkeland, E.E.; Chen, P.; Qiao, X.; Lonning, P.E.; Kallio, M.J. MYC-Induced miR-203b-3p and miR-203a-3p Control Bcl-xL Expression and Paclitaxel Sensitivity in Tumor Cells. Transl. Oncol. 2019, 12, 170–179. [Google Scholar] [CrossRef]
- Shu, S.; Wu, H.J.; Ge, J.Y.; Zeid, R.; Harris, I.S.; Jovanovic, B.; Murphy, K.; Wang, B.; Qiu, X.; Endress, J.E.; et al. Synthetic Lethal and Resistance Interactions with BET Bromodomain Inhibitors in Triple-Negative Breast Cancer. Mol. Cell 2020, 78, 1096–1113. [Google Scholar] [CrossRef]
- Han, B.; Molins, L.; He, Y.; Vinolas, N.; Sanchez-Lorente, D.; Boada, M.; Guirao, A.; Diaz, T.; Martinez, D.; Ramirez, J.; et al. Characterization of the MicroRNA Cargo of Extracellular Vesicles Isolated from a Pulmonary Tumor-Draining Vein Identifies miR-203a-3p as a Relapse Biomarker for Resected Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2022, 23, 7138. [Google Scholar] [CrossRef]
- Li, J.Z.; Li, J.; Wang, H.Q.; Li, X.; Wen, B.; Wang, Y.J. MiR-141-3p promotes prostate cancer cell proliferation through inhibiting kruppel-like factor-9 expression. Biochem. Biophys. Res. Commun. 2017, 482, 1381–1386. [Google Scholar] [CrossRef]
- Ye, Y.; Li, S.L.; Ma, Y.Y.; Diao, Y.J.; Yang, L.; Su, M.Q.; Li, Z.; Ji, Y.; Wang, J.; Lei, L.; et al. Exosomal miR-141-3p regulates osteoblast activity to promote the osteoblastic metastasis of prostate cancer. Oncotarget 2017, 8, 94834–94849. [Google Scholar] [CrossRef]
- Moon, S.U.; Park, Y.; Park, M.G.; Song, S.K.; Jeong, S.H.; Lee, Y.S.; Heo, H.J.; Jung, W.Y.; Kim, S. Theragnosis by a miR-141-3p molecular beacon: Simultaneous detection and sensitization of 5-fluorouracil resistant colorectal cancer cells through the activation of the TRIM13-associated apoptotic pathway. Chem. Commun. 2019, 55, 7466–7469. [Google Scholar] [CrossRef]
- Masoumi-Dehghi, S.; Babashah, S.; Sadeghizadeh, M. microRNA-141-3p-containing small extracellular vesicles derived from epithelial ovarian cancer cells promote endothelial cell angiogenesis through activating the JAK/STAT3 and NF-kappaB signaling pathways. J. Cell Commun. Signal 2020, 14, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Liu, G.; Sun, Y.; Gao, Y.; Ouyang, X.; Chang, C.; Gong, L.; Yeh, S. Targeting the estrogen receptor alpha (ERalpha)-mediated circ-SMG1.72/miR-141-3p/Gelsolin signaling to better suppress the HCC cell invasion. Oncogene 2020, 39, 2493–2508. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, P.; Kulkarni, P.; Majid, S.; Hashimoto, Y.; Shiina, M.; Shahryari, V.; Bhat, N.S.; Tabatabai, L.; Yamamura, S.; Saini, S.; et al. LncRNA CDKN2B-AS1/miR-141/cyclin D network regulates tumor progression and metastasis of renal cell carcinoma. Cell Death Dis. 2020, 11, 660. [Google Scholar] [CrossRef] [PubMed]
- Phatak, P.; Noe, M.; Asrani, K.; Chesnick, I.E.; Greenwald, B.D.; Donahue, J.M. MicroRNA-141-3p regulates cellular proliferation, migration, and invasion in esophageal cancer by targeting tuberous sclerosis complex 1. Mol. Carcinog. 2021, 60, 125–137. [Google Scholar] [CrossRef]
- Chao, F.; Song, Z.; Wang, S.; Ma, Z.; Zhuo, Z.; Meng, T.; Xu, G.; Chen, G. Novel circular RNA circSOBP governs amoeboid migration through the regulation of the miR-141-3p/MYPT1/p-MLC2 axis in prostate cancer. Clin. Transl. Med. 2021, 11, e360. [Google Scholar] [CrossRef]
- Porcel, J.M.; Light, R.W. Pleural effusions. Dis. Mon. 2013, 59, 29–57. [Google Scholar] [CrossRef]
- Filetti, V.; Lombardo, C.; Loreto, C.; Dounias, G.; Bracci, M.; Matera, S.; Rapisarda, L.; Rapisarda, V.; Ledda, C.; Vitale, E. Small RNA-Seq Transcriptome Profiling of Mesothelial and Mesothelioma Cell Lines Revealed microRNA Dysregulation after Exposure to Asbestos-like Fibers. Biomedicines 2023, 11, 538. [Google Scholar] [CrossRef]
- Yang, D.; Chen, C.; Xia, H.; Chen, J.; Yu, M. Characteristics of transcription profile, adhesion and migration of SETD2-loss Met-5A mesothelial cells exposed with crocidolite. J. Appl. Toxicol. 2023. [Google Scholar] [CrossRef]
- Dell’Anno, I.; Barbarino, M.; Barone, E.; Giordano , A.; Luzzi, L.; Bottaro, M.; Migliore, L.; Agostini, S.; Melani, A.; Melaiu, O.; et al. Catalano C, Cipollini M, Silvestri R, Corrado A, Gemignani F, Landi S. EIF4G1 and RAN as Possible Drivers for Malignant Pleural Mesothelioma. Int. J. Mol. Sci. 2020, 21, 4856. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Dweep, H.; Sticht, C.; Pandey, P.; Gretz, N. miRWalk—database: Prediction of possible miRNA binding sites by "walking" the genes of three genomes. J. Biomed. Inform. 2011, 44, 839–847. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Age (Years) | Sex | Diagnosis | Histological Subtype | PF Cytological Examination | Pleural Biopsy |
|---|---|---|---|---|---|---|
| B1 | 49 | Male | Heart failure | NA | Negative | Negative |
| B2 | 75 | Female | Heart failure | NA | ND | ND |
| B3 | 77 | Male | Heart failure | NA | Negative | ND |
| B4 | 63 | Female | Heart failure | NA | Negative | ND |
| B5 | 83 | Male | Heart failure | NA | Negative | ND |
| B6 | 86 | Female | Heart failure | NA | Negative | Negative |
| M1 | 50 | Female | Breast cancer | Luminal B/HER2- | Positive | ND |
| M2 | 88 | Female | Breast cancer | TNBC | Negative | ND |
| M3 | 70 | Female | Breast cancer | Luminal B/HER2+ | Positive | ND |
| M4 | 72 | Male | NSCLC | Adenocarcinoma | Positive | ND |
| M5 | 65 | Female | NSCLC | Adenocarcinoma | Positive | ND |
| M6 | 56 | Female | NSCLC | Adenocarcinoma | Positive | ND |
| M7 | 69 | Male | Colorectal cancer | Adenocarcinoma | Positive | ND |
| M8 | 60 | Male | Colorectal cancer | Adenocarcinoma | Positive | ND |
| M9 | 84 | Male | Colorectal cancer | Adenocarcinoma | Positive | Negative |
| M10 | 68 | Male | Non-Hodgkin lymphoma | DLBCL | Negative * | ND |
| M11 | 90 | Male | Non-Hodgkin lymphoma | DLBCL | Negative * | Negative |
| M12 | 83 | Female | Non-Hodgkin lymphoma | Follicular large B-cell lymphoma | Negative * | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marqués, M.; Pont, M.; Hidalgo, I.; Sorolla, M.A.; Parisi, E.; Salud, A.; Sorolla, A.; Porcel, J.M. MicroRNAs Present in Malignant Pleural Fluid Increase the Migration of Normal Mesothelial Cells In Vitro and May Help Discriminate between Benign and Malignant Effusions. Int. J. Mol. Sci. 2023, 24, 14022. https://doi.org/10.3390/ijms241814022
Marqués M, Pont M, Hidalgo I, Sorolla MA, Parisi E, Salud A, Sorolla A, Porcel JM. MicroRNAs Present in Malignant Pleural Fluid Increase the Migration of Normal Mesothelial Cells In Vitro and May Help Discriminate between Benign and Malignant Effusions. International Journal of Molecular Sciences. 2023; 24(18):14022. https://doi.org/10.3390/ijms241814022
Chicago/Turabian StyleMarqués, Marta, Mariona Pont, Iván Hidalgo, Maria Alba Sorolla, Eva Parisi, Antonieta Salud, Anabel Sorolla, and José M. Porcel. 2023. "MicroRNAs Present in Malignant Pleural Fluid Increase the Migration of Normal Mesothelial Cells In Vitro and May Help Discriminate between Benign and Malignant Effusions" International Journal of Molecular Sciences 24, no. 18: 14022. https://doi.org/10.3390/ijms241814022
APA StyleMarqués, M., Pont, M., Hidalgo, I., Sorolla, M. A., Parisi, E., Salud, A., Sorolla, A., & Porcel, J. M. (2023). MicroRNAs Present in Malignant Pleural Fluid Increase the Migration of Normal Mesothelial Cells In Vitro and May Help Discriminate between Benign and Malignant Effusions. International Journal of Molecular Sciences, 24(18), 14022. https://doi.org/10.3390/ijms241814022