Abstract
Phthalates have been widely studied for their reprotoxic effects in male rodents and in particular on testosterone production, for which reference doses were established. The female rodent brain can also represent a target for exposure to these environmental endocrine disruptors. Indeed, a large range of behaviors including reproductive behaviors, mood-related behaviors, and learning and memory are regulated by sex steroid hormones. Here we review the experimental studies addressing the effects and mechanisms of phthalate exposure on these behaviors in female rodents, paying particular attention to the experimental conditions (period of exposure, doses, estrous stage of analyses etc.). The objective of this review is to provide a clear picture of the consistent effects that can occur in female rodents and the gaps that still need to be filled in terms of effects and mode(s) of action for a better risk assessment for human health.
1. Introduction
Phthalates are among the most abundant organic pollutants in the environment [1], due to their extensive use in the plastic industry. Several phthalates are listed by the European Chemical Agency (ECHA) as substances of very high concern due to their toxicity for reproduction (butyl benzyl phthalate BBP, dibutyl phthalate DBP, dicyclohexyl phthalate DCHP, di(2-ethylhexyl)phthalate DEHP, di-isobutyl phthalate DIBP, diisohexyl phthalate DIHP, di-isopentyl phthalate DIPP, dipentyl phthalate DPP…), and endocrine disrupting properties for human health (BBP, DBP, DCHP, DEHP, DIBP) and the environment (DEHP). Based on their ability to reduce fetal testosterone levels in males, a tolerable daily intake dose of 50 µg/kg/d was initially established by the European Food Safety Authority [2], and this was recently updated and confirmed for BBP, DBP, DEHP, DINP, and DIDP for use in food contact materials [3]. In recent studies, we showed that the adult rodent nervous system is also vulnerable to exposure to DEHP on its own or in a phthalate mixture. The observed effects on reproductive behavior were linked to an endocrine mode of action involving the neural alteration of androgen receptors in males and progesterone receptors in females [4,5]. Interestingly, these effects were observed at doses equivalent to or below the tolerable daily intake (TDI) dose of 50 µg/kg/d in both male and female mice. Moreover, the females appeared more sensitive to phthalate exposure than males since, under comparable experimental conditions, several components of sexual behavior were affected in females [4].
Therefore, here we aim to review the experimental studies addressing the behavioral effects and underlying mechanisms of phthalate exposure in female rodents, with a particular focus on behaviors known to be induced or modulated by sex steroid hormones (reproductive and mood-related behaviors and learning and memory). Sex steroid hormones play a key role in the organization of the neural structures that underlie these behaviors during developmental and pubertal periods of life (Figure 1). During adulthood, they are also necessary for the activation of these circuitries, leading to the expression of behavior. The brain undergoes developmental and adult neuroplasticity, regulated by several factors including sex steroid hormones. Changes in hormonal levels or their signaling pathways following exposure to endocrine disrupting compounds (EDCs) can lead to long-term or transient modifications in these processes and consequently in behavior. In this context, previous reviews addressed the effects of exposure to EDCs, including phthalates, on social and reproductive behaviors [6,7,8], or effects of exposure to phthalates on hippocampal plasticity [9] or the nervous system [10]. The present review, in addition to being interested in the effects of female exposure to phthalates on behaviors known to be modulated by sex steroids, aims also to shed light on the experimental conditions used for such studies (phthalate doses, period of exposure, stage of the estrous cycle, etc.). Our ultimate goal is to provide a clear picture of the consistent effects that can occur in female rodents and the gaps that still need to be filled in terms of effects and mode(s) of action for a better risk assessment for human health.
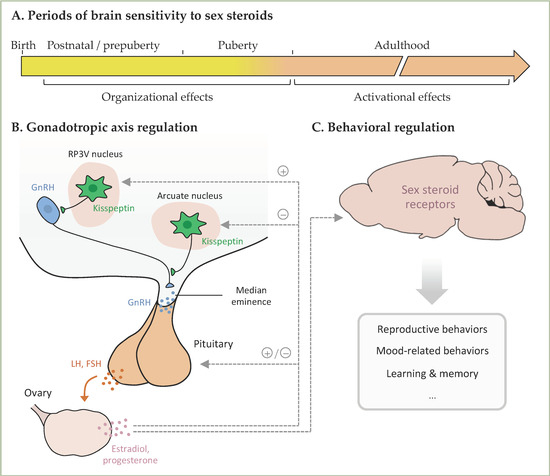
Figure 1.
(A). In females, the ovaries start synthesizing and liberating sex steroids from postnatal day 7. During the postnatal/prepubertal and pubertal periods, sex steroids play an important role in the organization of neural structures involved in the regulation of the gonadotropic axis and behaviors. During adulthood, sex steroids play an activation role. (B). Kisspeptin neurons from the RP3V and arcuate nucleus project on GnRH neurons, thereby stimulating GnRH liberation, which acts on the pituitary to stimulate the liberation of LH and FSH and consequently the ovarian secretion of gonadal hormones. In turn, sex steroids exert positive or negative feedbacks on kisspeptin neurons and the pituitary. (C). Ovarian sex steroids act on the brain to activate the neural structures underlying the mentioned behaviors. RP3V, rostral periventricular area of the third ventricle; GnRH, Gonadotropin-Releasing Hormone; LH, Luteinizing Hormone; FSH, Follicle Stimulating Hormone.
2. Hormonal Regulation of Female Behavior
2.1. Reproductive Behaviors
Sexual behavior. In rodents, female sexual behavior is restricted to a short period of receptivity that coincides with ovulation. During this period, females emit sexual pheromones, which provide information on their hormonal and receptivity states. During the precopulatory phase, both sexual partners engage in mutual olfactory investigation, which is important in the activation of the following behavioral sequence in both sexes. Once stimulated by female pheromones, the male emits ultrasonic vocalizations and exhibits urinary marking (for review [11]). Thereafter, during the copulatory phase, the male displays several mount episodes with pelvic thrusts and intromissions to which the female responds with a lordosis posture, which consists of having all four paws grounded, the hind region lifted, and the back arched. In mice, copulation ends with male ejaculation, whereas rats only reach satiety after several copulations.
At the neuroanatomical level, male pheromones stimulate the female olfactory bulb, which transmits olfactory information to the medial amygdala and to the bed nucleus of the stria terminalis. These regions send signals to the principal facilitatory system of lordosis behavior, i.e., the ventromedial hypothalamus and to the inhibitory system, which includes the lateral septum, medial preoptic area, and arcuate nuclei. These two systems send projections to the periaqueductal grey regions, which relay information to spinal motoneurons innervating muscles involved in the lordosis posture [12].
These circuitries are tightly regulated by ovarian hormones, estradiol and progesterone. The preovulatory surge of estradiol that occurs during proestrus triggers both the ovulatory surge of LH by the pituitary gland and the upregulation of progesterone receptors (PR) in the neural pathway that underlies female sexual behavior. In rodents, progesterone released after ovarian stimulation by LH acts on neural receptors to induce female receptivity. This hormonal sequence activates the facilitatory system and lifts the inhibition exerted by the inhibitory system to ensure lordosis behavior. Estradiol acts primarily through ERα given that in [13], neural ERα deletion greatly reduced the number of PR-immunoreactive neurons in the ventromedial hypothalamus and inhibited lordosis behavior, impaired kisspeptin expression, and arrested cyclicity [13], while in [14], neural ERβ deletion had no effect on the adult expression of sexual behavior or regulation of the gonadotropic axis.
Maternal behavior. Female rodents engage in intensive nest building as early as the middle of gestation, with an increased aggression towards intruders. After birth, dams show strong interest in pups and become highly sensitive to olfactory cues from newborns. Nursing behavior is further established by tactile stimulation of the mother by the pups that then gain access to the nipples to suckle. Pups also emit ultrasonic vocalizations; in response, dams exhibit a retrieving behavior to return the pups safely to the nest [15].
In parturient females, olfactory cues integrated by the amygdala are transmitted to the medial preoptic area to promote pup attractivity. The maternal brain is also sensitive to tactile and auditory cues from the pups, which stimulate the bed nucleus of stria terminalis via the cortex. The medial preoptic area and bed nucleus of stria terminalis stimulate other regions, ensuring the establishment and maintenance of maternal behavior, including the ventral tegmental area, nucleus accumbens, and paraventricular nucleus of the hypothalamus [7,16].
The onset of maternal behavior depends on several hormonal events that occur during gestation and parturition. In rodents, estradiol levels are relatively low during the first days of gestation and rise in the last days prior to parturition. Inversely, progesterone levels are high at the beginning and middle of gestation and then drop. The increased ratio of estradiol/progesterone levels at the end of gestation is crucial for uterine preparation for labor. This results in the increased expression of oxytocin and oxytocin receptors, which induce uterine contractions and are also involved in the expression of maternal behavior and milk ejection [17,18]. Among the other hormones involved in maternal behavior, prolactin is secreted through surges by the pituitary at the beginning of gestation and then before parturition. This complex hormonal environment is thus important for maternal responsiveness to pups at parturition. Indeed, the hormonal stimulation of the medial preoptic area and the bed nucleus of stria terminalis is thought to lift the pup avoidance induced by the periaqueductal grey region. Moreover, it activates the dopaminergic system that increases maternal behavior [19]. While the onset of maternal behavior relies on hormones, rodents will progressively become less dependent on hormones and more dependent on sensory and auditory cues from pups that are essential for the maintenance of maternal behavior.
2.2. Mood-Related Behaviors
Anxiety- and depressive-like behaviors are also modulated by estrogens, with a diminution of these behaviors during proestrus in both mice and rats [14,20,21,22]. The main brain regions involved in these behaviors are the raphe, amygdala, hippocampus, and prefrontal cortex, with an emerging role of the bed nucleus of stria terminalis [23]. In these structures, the serotonergic system is a strong modulator of mood-related behavior. Cellular bodies of serotoninergic neurons are localized in the raphe nuclei of the brain stem. Tryptophan hydroxylases (TPH) 1 and 2 are the limiting enzymes for serotonin production; TPH2 being the major isoform in neural cells [24]. Serotoninergic neurons send projections to the hippocampus, prefrontal cortex, and amygdala, which all express postsynaptic 5-HT1A receptors [25]. Extracellular levels of serotonin are regulated by serotonin transporters (SERT) that uptake serotonin after its liberation in the synaptic cleft. Dysfunctions in serotonin transmission have been associated with anxiety and depression in both humans [26,27] and rodents [28,29]. In [30,31], estrogens, through ERβ, were shown to mediate the estrogen-mediated modulation of the serotoninergic system, especially by increasing the expression levels of THP2 in the dorsal raphe.
Other neuroendocrine systems are also involved, such as the hypothalamus-pituitary-adrenal (HPA) axis. A constant activation of this axis by chronic stress may lead to its dysregulation and can cause anxiety-related disorders. Activation of the HPA axis in response to stress is modulated by the estrous cycle and estradiol levels [32,33]. Estradiol may also regulate mood-related behaviors through oxytocin and arginine-vasopressin, which were shown to reduce the anxiety- and depression-state levels in [34,35]. In particular, mice deleted for neural ERβ display increased anxiety- and depressive-like behaviors that are associated with reduced transcript levels of oxytocin and arginine-vasopressin in the bed nucleus of stria terminalis [20].
2.3. Learning and Memory
Learning and memory define the processes by which individuals acquire knowledge, store it, and later retrieve the learned information. Different types of learning and memory processes rely on various synaptic plasticity mechanisms and depend on several brain areas, among which, in [36,37], the hippocampus was shown to play a key role in spatial memory, nonspatial memory such as visual object recognition, and temporal processing of information. Behavioral tests assessing these different aspects of learning and memory are widely used in rodents including in the field of neurotoxicology [38].
Learning and memory are sensitive to gonadal hormones, which exert modulatory effects on these processes throughout the lifespan of the rat. The modulatory effects of estrogens on learning and memory and on underlying processes were extensively studied in females and the underlying mechanisms detailed in previous reviews [39,40,41]. Briefly, in [42], it was shown that estradiol facilitates the establishment of LTP, and in [43,44,45], it was shown that the type and number of dendritic spines are regulated by sex steroid hormones during the estrous cycle. These effects are mediated in part by the classical nuclear ERα and ERβ, given that females globally knocked out for the genes encoding these receptors exhibited impaired spatial learning in [46,47] and reduced hippocampal long-term potentiation (LTP) and plasticity in [48,49]. Non-classical signaling initiated at the cell membrane and inducing rapid modulation of hippocampal plasticity through the activation of second messenger pathways were also described for estradiol [50].
3. Behavioral and Neural Effects of Phthalate Exposure
Here we describe the data collected for the effects of exposure to phthalates on reproductive and mood-related behaviors and learning and memory in female rodents. The publications were gathered using the search engine Pubmed, with the keywords “Phthalate and nervous system”, “Phthalate and brain” and “Phthalate and female behavior” up to 30 October 2021.
Information on the gathered articles in terms of studied species, period of exposure, studied phthalates and doses, and age at analyses is presented in Figure 2. The behavioral data are reported in Table 1 for reproductive behaviors, Table 2 for mood behaviors, and Table 3 for learning and memory. Details about the behavioral tests used in the assessment of learning and memory are indicated in Table S3. When neuroanatomical or neuroendocrine observations were made in the same studies, they were reported in the same Table 1, Table 2 and Table 3. Otherwise, in the absence of behavioral analyses, this information can be found in Table S1 for the hypothalamus and Table S2 for brain regions related to cognitive processes. Given the estrogen-mediated regulation of female behavior and neuroendocrine responses, indications were made in the tables with respect to the identification or not of the stage of the estrous cycle in adult females.
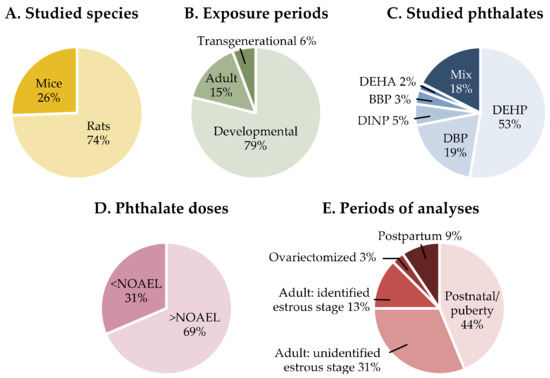
Figure 2.
Summary information on the experimental conditions used in the studies discussed in the present review. DEHP, Di-(2-ethylexyl) phthalate; DBP, Dibutyl phthalate; DINP, Diisononyl phthalate; BBP, Benzyl butyl phthalate; DEHA, Di-(2-ethylhexyl) adipate; NOAEL, non-observed adverse effect level.
3.1. Effects of Phthalate Exposure on Reproductive Behaviors
3.1.1. Sexual Behavior
Developmental exposure. Three studies addressed the effects of developmental exposure to high doses of DBP [51,52,53] or DINP [53] on the expression of female sexual behavior in rats (Table 1). One study reported a lower lordosis quotient at all tested doses [53] while the two other studies found no effect [51,52]. These three studies had not only the use of very high phthalate doses in common but also the assessment of lordosis behavior in intact females at the proestrus or estrus stage. Given the great variability in estradiol levels during these two stages, which may then induce differences in the receptivity levels between intact females, sexual behavior is generally measured under normalized hormonal levels through ovariectomy and priming with estradiol and progesterone.
Disruption of sexual behavior can occur either through changes in estrogen levels, progesterone levels, or both or in the expression levels of their receptors. In the study of Lee et al. [53], the behavioral changes were associated with transient reduced hormonal levels at PND7 as no modifications were seen at adulthood as in the study by Guerra et al. [51].
Table S1 shows that in other neuroendocrine and neuroanatomical studies where no behavioral studies were conducted, changes in the levels of LH at 5 or 30 mg/kg/d of DEHP [54,55] and of estradiol at 5 mg/kg/bw of DBP or progesterone at 500 mg/kg/d of DEHP [55,56] were observed in postnatal animals exposed during the prenatal/postnatal or prepubertal/pubertal period. Similarly, changes were seen in neural aromatase activity for exposure to at least 15 µg/kg/d of DEHP [57], or in the expression of ER for exposure to 0.5 mg/kg/d of DBP and of PR at the highest DINP dose tested [56,58]. Modifications in the GnRH/kisspeptin system, which governs the gonadotropic axis, were seen from 5 mg/kg/d for DBP and 1 mg/kg/d of DEHP [56,59]. Whether these changes that impact the developing brain trigger adult reproductive dysfunctions later on requires further studies. Longer estrous cycles were described [55,56] depending on the exposure period [56].
Among the five studies that analyzed the impact of prenatal/postnatal or prepubertal/pubertal exposure in adult females [55,60,61,62,63,64], only two studies identified the stage of the estrous cycle [55,61]. Roepke et al. [61] reported the unchanged expression of Kiss1, ERα, and ERβ in the arcuate nucleus of proestrus females exposed to 500 mg/kg/d of DEHP. Yu et al. [55] confirmed the unaffected Kiss1 expression in the arcuate nucleus in estrus females exposed to DEHP at 500 or 5 mg/kg/d during the prepubertal/pubertal period, and showed that these two doses induced opposite effects with either higher or lower hypothalamic GnRH expression, Kiss1 mRNAs levels in the AVPV, and serum levels of LH, estradiol, and progesterone. Altogether, these observations indicate that neuroendocrine changes can occur in the female brain areas that underlie the regulation of the gonadotropic axis following developmental exposure to moderate to low doses of phthalates. Whether these modifications impair the adult expression of sexual behavior still needs further investigations.
Adult exposure. To our knowledge, only one study investigated the effects of chronic adult exposure to phthalates on female sexual behavior [4]. This analysis was performed on ovariectomized females, which were supplemented with estradiol and primed with progesterone before tests to normalize their hormonal levels. It was shown that exposure to low doses of DEHP alone (5 and 50 µg/kg/d) or in an environmental phthalate mixture disrupted both the precopulatory and copulatory phases of sexual behavior. Females exposed to DEHP alone or in a phthalate mixture were less attractive to males, and those exposed to DEHP at 50 µg/kg/d or to the phthalate mixture were unable to discriminate between male and female pheromonal cues. Moreover, the lordosis quotient was diminished while the rejection behavior towards male mounts was increased.
These behavioral alterations were associated with a selective reduced number of PR-immunoreactive neurons in the neural circuitry that underlies sexual behavior (arcuate nucleus, preoptic area, ventromedial hypothalamus, medial amygdala, and bed nucleus of the stria terminalis) [4]. In these regions, no effects were found on the number of ERα-immunoreactive neurons. This suggests that, under normalized hormonal levels, exposure to DEHP alone or in a mixture directly targeted neural structures to lower the expression of female sexual behavior through an endocrine mode of action involving the PR.
Two other studies assessed the effects of acute or sub-chronic exposure to BBP or DEHP in ovariectomized or intact females, respectively [65,66]. Acute exposure of hormonally depleted rats to 10 mg of BBP increased PR expression in the preoptic area only [65], while Liu et al. [66] reported increased hypothalamic GnRH expression and lower hormonal levels following sub-chronic exposure to DEHP at 300 to 3000 mg/kg/d in females at an unidentified estrous stage (Table S1). Apart from the fact that these treatments affected hypothalamic gene expression or hormonal levels, their physiological meaning in the context of gonadotropic axis needs further investigation.

Table 1.
Effects of phthalate exposure on female reproductive behaviors. Avp, arginine-vasopressin; BBP, benzyl butyl phthalate; Cyp19a1, cytochrome P450 family 19 subfamily member 1; DBP, dibutyl phthalate; DEHA, di-(2-ethylhexyl) adipate; DEHP, Di-(2-ethylexyl) phthalate; DEP, diethyl phthalate; DiBP, diisobutyl phthalate; DINP, diisononyl phthalate; E2, estradiol; Esr1 and 2, estrogen receptor 1 and 2; FSH, follicle stimulating hormone; GD, gestational day; GnRH1, Gonadotropin Releasing Hormone 1; Kiss1, kisspeptine 1; LH, luteinizing hormone; Oxt, oxytocin; OVX, ovariectomy; P, progesterone; PND, postnatal day; PNW, postnatal week; PPD, postpartum day; ppm, parts per million.
3.1.2. Maternal Behavior
Developmental exposure. One study assessed the effects of perinatal exposure to DEHP at 10 mg/kg/d on pup-retrieving behavior of F1 rats [67]. Exposure increased the latency to retrieve the first pup and lowered the total number of pups retrieved and the time spent licking the pups but did not affect nursing. These behavioral effects were associated with lower hypothalamic levels of oxytocin receptor expression, increased oxytocin receptor methylation, and lower plasmatic oxytocin [67], suggesting that DEHP may reduce maternal behavior by impacting hypothalamic oxytocin signaling.
Adult exposure. Four studies investigated the effects of adult phthalate exposure on maternal behavior, with exposure beginning during gestation and lasting until the lactation period [60,68,69,70]. Lee et al. [70] reported that dams exposed to DBP (50 or 100 mg/kg/d) exhibited a higher latency to retrieve the second pup and created a poor-quality nest in the DBP-50 group. In contrast, no effects of exposure to high doses of DINP (300–900 mg/kg/d) or to a phthalate mixture (0.2 to 1 mg/kg/d) were found on pup retrieval or spontaneous maternal behavior in rats [60,68]. Similarly, exposure to DEHP (5 to 400 µg/kg/d) had no effect on the spontaneous maternal behavior of F0 or F2 mouse dams [69].
3.2. Effects of Phthalate Exposure on Mood-Related Behaviors
Developmental exposure. Eight studies (five on rats, three on mice) investigated the effects of prenatal/postnatal or prepubertal/pubertal exposure to DEHP or a phthalate mixture on anxiety-like behavior of F1 offspring, using mainly the elevated plus maze alone or in combination with the open-field (Table 2). Among these studies, seven analyzed these effects on cyclic females [69,71,72,73,74,75,76] and one in postpartum females [67]. Four out of seven studies found increased anxiety-state levels in PND30-35 mice exposed to 5 or 40 µg/kg/d of DEHP [69], in PND42 and adult females at the diestrus or estrus stage exposed to DEHP above 10 mg/kg/d [74,75], and in adult females of unknown estrous stage exposed to DEHP from 1 mg/kg/d [73]. In the study of Xu et al. [75], increased depressive-like behavior was also observed in PND42 and adult females at the diestrus stage. In these studies, increased anxiety-related behavior was associated with impaired HPA axis as evidenced by higher ACTH and lower corticosterone levels together with elevated hypothalamic amounts of GR [73], and lower amounts of hippocampal and striatal ERβ and dopamine receptor 2 [75], with no changes in circulating levels of estradiol [74,75]. The remaining three of seven studies reported no effects in pubertal (PND30, PND42 and PND45) or adult females of unknown estrous stage, which were exposed prenatally or postnatally to DEHP at 30 mg/kg/d [71] or 200 mg/kg/d [76] or to a phthalate mixture at 0.2 or 1 mg/kg/d [72]. Whether this discrepancy is due to differences in the female estrous stage or to other experimental conditions needs further investigation.
No effects of developmental exposure were observed in postpartum females perinatally exposed to 50 or 100 mg/kg/d of DBP [67]. In studies analyzing the transgenerational effects of DEHP exposure in the F3 generation [77,78], no effects were reported in PND30-35 and adult females of unknown estrous stage except at the high dose of 750 mg/kg/d, which seemed to induce a higher time spent in the open arms of the elevated plus maze [77].
Table 2.
Effects of phthalate exposure on mood-related behaviors. ACTH, adrenocorticotropic hormone; Akt, protein kinase B; AR, androgen receptor; BBP, benzyl butyl phthalate; BDNF, brain-derived neurotrophic factor; CREB, cAMP response element-binding protein; CRH, corticotropin-releasing hormone; D1R and D2R, dopamine receptor 1 and 2; DBP, dibutyl phthalate; DEHP, Di-(2-ethylexyl) phthalate, DEP, diethyl phthalate; DiBP, diisobutyl phthalate; DINP, diisononyl phthalate; DNMT, DNA methyltransferase; E2, estradiol; Esr1 and 2, estrogen receptor 1 and 2; ERβ, estrogen receptor β; ERK, extracellular signal-regulated kinases; FSH, follicle stimulating hormone; GD, gestational day; GR, glucocorticoid receptor; LH, luteinizing hormone; MeCP2, methyl-CpG binding protein 2; MR, mineralocorticoid receptor; NR43A1 and 3, Nuclear Receptor Subfamily 4 Group A Member 1 and 3; P, progesterone; PND, postnatal day; PPD, postpartum day.
Adult exposure. One study examined the effects of adult exposure to DBP on depressive-like behavior during the lactational period in mice [70]. Exposure of dams to DBP at 50 and 100 mg/kg/d had no effect on the forced swim and tail suspension tests.
3.3. Effects of Phthalate Exposure on Learning and Memory and Related Processes
Developmental exposure. Five studies (three on rats, two on mice) analyzed the effects of prenatal/postnatal exposure to DBP, DINP, and DEHP at high doses ranging from 10 mg/kg/d to more than 1 g/kg/d on spatial memory using the Morris water maze [76,79,80,81] or both Morris water and radial arm mazes [68] as shown in Table 3. No treatment effects were reported on the learning of prepubertal and adult female mice [79] or prepubertal or pubertal rats [68,76,80], while performance during the test day seemed to change in three of these studies [68,76,80]. A lower number of neurons in the three hippocampal regions, associated with a higher apoptotic cell death and caspase-3 activity, were observed at PND5 and PND21, but not in adulthood, in rats exposed to DBP at 500 mg/kg/d during the perinatal period [80]. This suggested that the behavioral changes were possibly caused by neurotoxic effects of such high phthalate doses.
Four other studies assessed the effects of exposure to lower doses of phthalates with BBP at 10 µg/kg/d, DEHP at 33 mg/kg/d, or a phthalate mixture at 0.2 to 1 mg/kg/d on aversive stimulus associated learning [82], object recognition memory [83], and cognitive flexibility [72,84], respectively. In mice, the novel object recognition test indicated a higher preference for the novel object in mice exposed to 200 mg/kg/d [83]. Cognitive flexibility tests showed a lower percentage of correct answers and higher percentage of perseveration errors after exposure to a phthalate mixture at 1 mg/kg/d. The cognitive flexibility texts showed a higher percentage of omission errors in rats exposed during the prenatal/postnatal period [84] but not in those exposed during the prepubertal/pubertal period [72]. In particular, the behavioral effects induced by prenatal/postnatal exposure to this phthalate mixture were associated with a lower number of neurons and synapses in the prefrontal cortex and a reduced volume of this brain region in adult females [84]. This phthalate mixture also induced earlier changes given that reduced neurogenesis and increased apoptosis in the cortex of PND5 and PND10 rats at 200–1000 µg/kg/d [85] and the reduced expression of arginine-vasopressin receptor 1B in the prefrontal cortex of PND10 rats at 0.2 mg/kg/d without changing the expression of other signaling pathways on PND10 or adulthood were reported by this laboratory [60].
In the aversive-stimulus associative test, a shorter duration of freezing during the interval period and the tone period was observed in rats exposed to BBP at 10 µg/kg/d [82]. Analyses of the hippocampus and amygdala, the two regions involved in this behavior, showed increased amounts of synapsin-1 in the dorsal hippocampus and lower amounts of ERα and MeCP2 protein in the amygdala [82]. These studies suggest that developmental exposure to even relatively low doses of phthalates can alter morphology and cognitive function related to the hippocampus, cortex, and amygdala.
Table S2 summarizes the data from other studies assessing the effects of developmental exposure to phthalates on brain morphology and key signaling pathways that underlie hippocampal and cortical functions. Three studies assessing the effects of gestational and lactational exposure to DEHP at high doses ranging from 30 to 750 mg/kg/d did not report any effect on hippocampal structural neuroplasticity and protein markers of functional plasticity, or cortical signaling pathways in the hippocampus on PND7, PND14, PND21 [86,87,88], or LTP in adult female rats [86]. Postnatal exposure of rats to 10 mg/kg/d of DEHP affected neither hippocampal neurogenesis nor plasticity in PND26 animals [89,90]. In contrast, modifications were observed in the dopaminergic signaling pathway in the cortex of PND15 mice perinatally exposed to DBP at 50 mg/kg/d [70]. The number of dopaminergic neurons in the substantia nigra and ventral tegmental area of adult females postnatally exposed to 10–20 mg/kg/d of DEHP was also affected [91]. It would have been interesting to perform all these morphological and molecular analyses in parallel in both the developing and adult brain to see whether the reported changes last over adulthood and whether they can explain the behavioral modifications presented above.
Table 3.
Effects of phthalate exposure on learning and memory. BBP, benzyl butyl phthalate; DBP, dibutyl phthalate; DEHP, di-(2-ethylexyl) phthalate; DEP, diethyl phthalate; DiBP, diisobutyl phthalate; DINP, diisononyl phthalate; E2, estradiol; ERα, estrogen receptor α; GD, gestational day; MeCP2, methyl-CpG binding protein 2; mPFC, medial prefrontal cortex; LIMK, LIM domain kinase; NMDA, N-methyl-D-aspartate; NR1 and NR2B, NMDA receptor 1 and 2b; PAK, P21 activated kinase; PND, postnatal day; PPW, postpartum week; ROCK, Rho Associated Coiled-Coil Containing Protein Kinase; Rac, Rac Family Small GTPase; Ube3a, ubiquitin protein ligase E3A.
Adult exposure. A study performed in ICR mice showed that sub-chronic exposure to DBP at doses ranging from 10 to 250 mg/kg/d did not affect spatial memory measured in the Morris water maze, despite molecular changes reported in the hippocampus [92].
4. Discussion
Beyond the reduced testosterone production reported for phthalate exposure and used to establish the reference doses for these molecules, the female brain can also represent a sensitive target for these environmental chemicals. It appears, from the present review, that the exposure of rodents to phthalates, in particular at doses in the range of or lower than the no-observed adverse effect level (NOAEL of 5 mg/kg bw/d) or TDI doses may affect reproductive and mood-related behaviors and learning and memory. In some cases, behavioral modifications were associated with an endocrine mode of action involving alterations in neural PR immunoreactivity [4], the amounts of hippocampal and striatal ERβ [75], the amounts of ERα protein in the amygdala [82], oxytocin signaling [65], HPA axis [73], or the expression of arginine-vasopressin receptor 1B in the prefrontal cortex [60]. However, several aspects still need further attention to achieve a better view on the potential effects of environmental exposure to phthalates on female brain and behavior and underlying endocrine mode(s) of action. First, in the experimental studies addressing the female behaviors presented in this review, the endocrine mode(s) of action that underlie the observed effects are not systematically characterized. Indeed, while measurement of circulating levels of sex steroids and brain expression of their receptors is generally performed in parallel in the studies assessing reproductive behaviors, this is not always the case for the other behaviors. Only 50% of the total studies analyzing mood-related behavior and 20% of those analyzing learning and memory addressed this aspect. Second, a majority of available studies used phthalate doses in the mg to g range, higher than the NOAEL established for these substances (Figure 2). This could be barely relevant for the doses estimated for the environmental exposure to phthalates, which are in general at the microgram or nanogram range [93]. Moreover, the endocrine mode(s) of action characterized for an adverse effect induced by high doses of a substance can be secondary to other general effects; this point is not specific to phthalates or females but concerns many other substances and both sexes. Third, several analyses of pubertal or adult females did not report the stage of the estrous cycle (Figure 2). This greatly hampers the correct interpretation of data given the modulation of several reproductive and non-reproductive behaviors by sex steroids. In this context, it is important to mention that it is not necessary to use four groups of females with each corresponding to a stage of the estrous cycle. The estrous cycle can be monitored in cyclic females during the period of analyses, and females can be divided into two groups with one corresponding to the estrus-proestrus phase (high estrogen levels) and the other one to the diestrus-estrus stage (low estrogen levels) as previously reported for analyses of the anxiety- or depressive-related behaviors [14,20]. Finally, adulthood is still an underestimated period of exposure with only a few studies analyzing the effects induced by phthalate exposure in adult females (Figure 2). Previous studies showed that the adult brain can also be impacted by endocrine disruptors including phthalates in both sexes [4,5,94,95,96,97,98]. Given the critical role of several brain regions in the regulation of female behaviors and the potential transgenerational effects of many chemical substances including phthalates on descendants, adulthood should be studied as much as the developmental periods.
In humans, epidemiological studies reported associations between exposure to phthalates and low interest in sexual activity in women [99]. Previous systematic reviews of the literature also support an association between prenatal exposure to phthalates and adverse cognitive outcomes and neurological disorders [100,101]. Other studies described an association between urinary phthalates and anxiety or depressive symptoms in young adults [102] or the elderly population [103]. It is possible that these effects in humans occur through disruption of endocrine systems including those related to sex steroids. Indeed, it is largely known that sex steroids are not only mandatory for sexual function but that they also influence anxiety state level and cognition [104,105,106]. Therefore, the adverse effects and mode(s) of action reported in rodents following environmental exposure to phthalates might be also relevant for humans.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms23052559/s1.
Author Contributions
N.A. and S.M.-K. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Sorbonne Université, Centre National de la Recherche Scientifique (CNRS) and Institut National de la Santé et de la Recherche Médicale (Inserm).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gao, D.-W.; Wen, Z.-D. Phthalate Esters in the Environment: A Critical Review of Their Occurrence, Biodegradation, and Removal during Wastewater Treatment Processes. Sci. Total Environ. 2016, 541, 986–1001. [Google Scholar] [CrossRef]
- European Food Safety Authority. Opinion of the Scientific Panel on Food Additives, Flavourings, Processing Aids and Materials in Contact with Food (AFC) Related to Bis(2-Ethylhexyl)Phthalate (DEHP) for Use in Food Contact Materials. EFSA J. 2005, 243, 1–20. [Google Scholar] [CrossRef]
- EFSA Panel on Food Contact Materials, Enzymes and Processing Aids (CEP); Silano, V.; Barat Baviera, J.M.; Bolognesi, C.; Chesson, A.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; et al. Update of the Risk Assessment of Di-Butylphthalate (DBP), Butyl-Benzyl-Phthalate (BBP), Bis(2-Ethylhexyl)Phthalate (DEHP), Di-Isononylphthalate (DINP) and Di-Isodecylphthalate (DIDP) for Use in Food Contact Materials. EFSA J. Eur. Food Saf. Auth. 2019, 17, e05838. [Google Scholar] [CrossRef] [Green Version]
- Adam, N.; Brusamonti, L.; Mhaouty-Kodja, S. Exposure of Adult Female Mice to Low Doses of Di(2-Ethylhexyl) Phthalate Alone or in an Environmental Phthalate Mixture: Evaluation of Reproductive Behavior and Underlying Neural Mechanisms. Environ. Health Perspect. 2021, 129, 017008. [Google Scholar] [CrossRef]
- Dombret, C.; Capela, D.; Poissenot, K.; Parmentier, C.; Bergsten, E.; Pionneau, C.; Chardonnet, S.; Hardin-Pouzet, H.; Grange-Messent, V.; Keller, M.; et al. Neural Mechanisms Underlying the Disruption of Male Courtship Behavior by Adult Exposure to Di(2-Ethylhexyl) Phthalate in Mice. Environ. Health Perspect. 2017, 125, 097001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gore, A.C.; Krishnan, K.; Reilly, M.P. Endocrine-Disrupting Chemicals: Effects on Neuroendocrine Systems and the Neurobiology of Social Behavior. Horm. Behav. 2019, 111, 7–22. [Google Scholar] [CrossRef]
- Keller, M.; Vandenberg, L.N.; Charlier, T.D. The Parental Brain and Behavior: A Target for Endocrine Disruption. Front. Neuroendocrinol. 2019, 54, 100765. [Google Scholar] [CrossRef] [PubMed]
- Mhaouty-Kodja, S.; Naulé, L.; Capela, D. Sexual Behavior: From Hormonal Regulation to Endocrine Disruption. Neuroendocrinology 2018, 107, 400–416. [Google Scholar] [CrossRef] [PubMed]
- Holahan, M.R.; Smith, C.A. Phthalates and Neurotoxic Effects on Hippocampal Network Plasticity. NeuroToxicology 2015, 48, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Safarpour, S.; Ghasemi-Kasman, M.; Safarpour, S.; Darban, Y.M. Effects of Di-2-Ethylhexyl Phthalate on Central Nervous System Functions: A Narrative Review. Curr. Neuropharmacol. 2021. Online ahead of print. [Google Scholar] [CrossRef]
- Mhaouty-Kodja, S. Courtship Vocalizations: A Potential Biomarker of Adult Exposure to Endocrine Disrupting Compounds? Mol. Cell. Endocrinol. 2020, 501, 110664. [Google Scholar] [CrossRef] [PubMed]
- Kow, L.M.; Pfaff, D.W. Physiology of Somatosensory and Estrogenic Control Over the Lordosis Reflex. In Gonadal Steroids and Brain Function; Wuttke, W., Horowski, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1981; Volume 3, pp. 262–273. ISBN 978-3-540-10606-7. [Google Scholar]
- Trouillet, A.-C.; Ducroq, S.; Naulé, L.; Capela, D.; Parmentier, C.; Radovick, S.; Hardin-Pouzet, H.; Mhaouty-Kodja, S. Sex Differential Effects of Neural Estrogen Receptor Alpha Deletion on Reproductive Behavior in Mice. Commun. Biol. Under final revision.
- Naulé, L.; Robert, V.; Parmentier, C.; Martini, M.; Keller, M.; Cohen-Solal, M.; Hardin-Pouzet, H.; Grange-Messent, V.; Franceschini, I.; Mhaouty-Kodja, S. Delayed Pubertal Onset and Prepubertal Kiss1 Expression in Female Mice Lacking Central Oestrogen Receptor Beta. Hum. Mol. Genet. 2015, 24, 7326–7338. [Google Scholar] [CrossRef] [Green Version]
- Numan, M.; Insel, T.R. The Neurobiology of Parental Behavior; Springer: New York, NY, USA, 2003; ISBN 9780387217994. [Google Scholar]
- Sheehan, T.; Paul, M.; Amaral, E.; Numan, M.J.; Numan, M. Evidence That the Medial Amygdala Projects to the Anterior/Ventromedial Hypothalamic Nuclei to Inhibit Maternal Behavior in Rats. Neuroscience 2001, 106, 341–356. [Google Scholar] [CrossRef]
- Neumann, I.D. Brain Oxytocin: A Key Regulator of Emotional and Social Behaviours in Both Females and Males. J. Neuroendocrinol. 2008, 20, 858–865. [Google Scholar] [CrossRef]
- Soloff, M.S.; Alexandrova, M.; Fernstrom, M.J. Oxytocin Receptors: Triggers for Parturition and Lactation? Science 1979, 204, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- Numan, M. Hypothalamic Neural Circuits Regulating Maternal Responsiveness Toward Infants. Behav. Cogn. Neurosci. Rev. 2006, 5, 163–190. [Google Scholar] [CrossRef] [Green Version]
- Dombret, C.; Naulé, L.; Trouillet, A.-C.; Parmentier, C.; Hardin-Pouzet, H.; Mhaouty-Kodja, S. Effects of Neural Estrogen Receptor Beta Deletion on Social and Mood-Related Behaviors and Underlying Mechanisms in Male Mice. Sci. Rep. 2020, 10, 6242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora, S.; Dussaubat, N.; Díaz-Véliz, G. Effects of the Estrous Cycle and Ovarian Hormones on Behavioral Indices of Anxiety in Female Rats. Psychoneuroendocrinology 1996, 21, 609–620. [Google Scholar] [CrossRef]
- Walf, A.A.; Koonce, C.; Manley, K.; Frye, C.A. Proestrous Compared to Diestrous Wildtype, but Not Estrogen Receptor Beta Knockout, Mice Have Better Performance in the Spontaneous Alternation and Object Recognition Tasks and Reduced Anxiety-like Behavior in the Elevated plus and Mirror Maze. Behav. Brain Res. 2009, 196, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Lebow, M.A.; Chen, A. Overshadowed by the Amygdala: The Bed Nucleus of the Stria Terminalis Emerges as Key to Psychiatric Disorders. Mol. Psychiatry 2016, 21, 450–463. [Google Scholar] [CrossRef] [Green Version]
- Walther, D.J.; Bader, M. A Unique Central Tryptophan Hydroxylase Isoform. Biochem. Pharmacol. 2003, 66, 1673–1680. [Google Scholar] [CrossRef]
- Savitz, J.; Lucki, I.; Drevets, W.C. 5-HT1A Receptor Function in Major Depressive Disorder. Prog. Neurobiol. 2009, 88, 17–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, M.W.; Sugden, K.; Huezo-Diaz, P.; Williamson, R.; Sterne, A.; Purcell, S.; Sham, P.C.; Craig, I.W. Association Analysis of Monoamine Genes with Measures of Depression and Anxiety in a Selected Community Sample of Siblings. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2005, 135B, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gainetdinov, R.R.; Beaulieu, J.-M.; Sotnikova, T.D.; Burch, L.H.; Williams, R.B.; Schwartz, D.A.; Krishnan, K.R.R.; Caron, M.G. Loss-of-Function Mutation in Tryptophan Hydroxylase-2 Identified in Unipolar Major Depression. Neuron 2005, 45, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donaldson, Z.R.; Nautiyal, K.M.; Ahmari, S.E.; Hen, R. Genetic Approaches for Understanding the Role of Serotonin Receptors in Mood and Behavior. Curr. Opin. Neurobiol. 2013, 23, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, J.P.R.; Medvedev, I.O.; Caron, M.G. The 5-HT Deficiency Theory of Depression: Perspectives from a Naturalistic 5-HT Deficiency Model, the Tryptophan Hydroxylase 2 Arg 439 His Knockin Mouse. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2444–2459. [Google Scholar] [CrossRef] [Green Version]
- Donner, N.; Handa, R.J. Estrogen Receptor Beta Regulates the Expression of Tryptophan-Hydroxylase 2 MRNA within Serotonergic Neurons of the Rat Dorsal Raphe Nuclei. Neuroscience 2009, 163, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, H.; Barros, R.P.A.; Sugiyama, N.; Krishnan, V.; Yaden, B.C.; Kim, H.-J.; Warner, M.; Gustafsson, J.-Å. Involvement of Estrogen Receptor β in Maintenance of Serotonergic Neurons of the Dorsal Raphe. Mol. Psychiatry 2013, 18, 674–680. [Google Scholar] [CrossRef]
- Oyola, M.G.; Handa, R.J. Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes: Sex Differences in Regulation of Stress Responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef]
- Viau, V.; Meaney, M.J. Variations in the Hypothalamic-Pituitary-Adrenal Response to Stress during the Estrous Cycle in the Rat. Endocrinology 1991, 129, 2503–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duque-Wilckens, N.; Steinman, M.Q.; Laredo, S.A.; Hao, R.; Perkeybile, A.M.; Bales, K.L.; Trainor, B.C. Inhibition of Vasopressin V1a Receptors in the Medioventral Bed Nucleus of the Stria Terminalis Has Sex- and Context-Specific Anxiogenic Effects. Neuropharmacology 2016, 110, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acevedo-Rodriguez, A.; Mani, S.K.; Handa, R.J. Oxytocin and Estrogen Receptor β in the Brain: An Overview. Front. Endocrinol. 2015, 6, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kesner, R.P.; Hunsaker, M.R. The Temporal Attributes of Episodic Memory. Behav. Brain Res. 2010, 215, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Hunsaker, M.R.; Kesner, R.P. Unfolding the Cognitive Map: The Role of Hippocampal and Extra-Hippocampal Substrates Based on a Systems Analysis of Spatial Processing. Neurobiol. Learn. Mem. 2018, 147, 90–119. [Google Scholar] [CrossRef] [PubMed]
- Harry, G.J.; McBride, S.; Witchey, S.K.; Mhaouty-Kodja, S.; Trembleau, A.; Bridge, M.; Bencsik, A. Roadbumps at the Crossroads of Integrating Behavioral and In Vitro Approaches for Neurotoxicity Assessment. Front. Toxicol. Accepted.
- Frick, K.M. Molecular Mechanisms Underlying the Memory-Enhancing Effects of Estradiol. Horm. Behav. 2015, 74, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Galea, L.A.M.; Wainwright, S.R.; Roes, M.M.; Duarte-Guterman, P.; Chow, C.; Hamson, D.K. Sex, Hormones and Neurogenesis in the Hippocampus: Hormonal Modulation of Neurogenesis and Potential Functional Implications. J. Neuroendocrinol. 2013, 25, 1039–1061. [Google Scholar] [CrossRef] [Green Version]
- Hamson, D.K.; Roes, M.M.; Galea, L.A.M. Sex Hormones and Cognition: Neuroendocrine Influences on Memory and Learning. Compr. Physiol. 2016, 6, 1295–1337. [Google Scholar] [CrossRef] [Green Version]
- Córdoba Montoya, D.A.; Carrer, H.F. Estrogen Facilitates Induction of Long Term Potentiation in the Hippocampus of Awake Rats. Brain Res. 1997, 778, 430–438. [Google Scholar] [CrossRef]
- Brusco, J.; Wittmann, R.; de Azevedo, M.S.; Lucion, A.B.; Franci, C.R.; Giovenardi, M.; Rasia-Filho, A.A. Plasma Hormonal Profiles and Dendritic Spine Density and Morphology in the Hippocampal CA1 Stratum Radiatum, Evidenced by Light Microscopy, of Virgin and Postpartum Female Rats. Neurosci. Lett. 2008, 438, 346–350. [Google Scholar] [CrossRef]
- González-Burgos, I.; Alejandre-Gómez, M.; Cervantes, M. Spine-Type Densities of Hippocampal CA1 Neurons Vary in Proestrus and Estrus Rats. Neurosci. Lett. 2005, 379, 52–54. [Google Scholar] [CrossRef] [PubMed]
- Woolley, C.; Gould, E.; Frankfurt, M.; McEwen, B. Naturally Occurring Fluctuation in Dendritic Spine Density on Adult Hippocampal Pyramidal Neurons. J. Neurosci. 1990, 10, 4035–4039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fugger, H.N.; Foster, T.C.; Gustafsson, J.; Rissman, E.F. Novel Effects of Estradiol and Estrogen Receptor Alpha and Beta on Cognitive Function. Brain Res. 2000, 883, 258–264. [Google Scholar] [CrossRef]
- Rissman, E.F.; Heck, A.L.; Leonard, J.E.; Shupnik, M.A.; Gustafsson, J.-A. Disruption of Estrogen Receptor Beta Gene Impairs Spatial Learning in Female Mice. Proc. Natl. Acad. Sci. USA 2002, 99, 3996–4001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Day, M.; Good, M. Ovariectomy-Induced Disruption of Long-Term Synaptic Depression in the Hippocampal CA1 Region in Vivo Is Attenuated with Chronic Estrogen Replacement. Neurobiol. Learn. Mem. 2005, 83, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Spencer-Segal, J.L.; Tsuda, M.C.; Mattei, L.; Waters, E.M.; Romeo, R.D.; Milner, T.A.; McEwen, B.S.; Ogawa, S. Estradiol Acts via Estrogen Receptors Alpha and Beta on Pathways Important for Synaptic Plasticity in the Mouse Hippocampal Formation. Neuroscience 2012, 202, 131–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taxier, L.R.; Gross, K.S.; Frick, K.M. Oestradiol as a Neuromodulator of Learning and Memory. Nat. Rev. Neurosci. 2020, 21, 535–550. [Google Scholar] [CrossRef]
- Guerra, M.T.; Scarano, W.R.; de Toledo, F.C.; Franci, J.A.A.; Kempinas, W.D.G. Reproductive Development and Function of Female Rats Exposed to Di-η-Butyl-Phthalate (DBP) in Utero and during Lactation. Reprod. Toxicol. 2010, 29, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.; Heng, K.; Mann, N.; Anand-Ivell, R.; Ivell, R. Maternal Exposure to Dibutyl Phthalate (DBP) or Diethylstilbestrol (DES) Leads to Long-Term Changes in Hypothalamic Gene Expression and Sexual Behavior. Int. J. Mol. Sci. 2021, 22, 4163. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-C.; Yamanouchi, K.; Nishihara, M. Effects of Perinatal Exposure to Phthalate/Adipate Esters on Hypothalamic Gene Expression and Sexual Behavior in Rats. J. Reprod. Dev. 2006, 52, 343–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, S.; Samaniego, Y.A.; Cutrera, R.; Reynoso, R.; Cardoso, N.; Scacchi, P.; Moguilevsky, J.A.; Ponzo, O.J. Different Effects by Sex on Hypothalamic–Pituitary Axis of Prepubertal Offspring Rats Produced by in Utero and Lactational Exposure to Di-(2-Ethylhexyl) Phthalate (DEHP). NeuroToxicology 2012, 33, 78–84. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, F.; Han, J.; Lu, R.; Li, Q.; Cai, L.; Li, B.; Chen, J.; Wang, K.; Lin, W.; et al. Opposite Effects of High- and Low-Dose Di-(2-Ethylhexyl) Phthalate (DEHP) Exposure on Puberty Onset, Oestrous Cycle Regularity and Hypothalamic Kisspeptin Expression in Female Rats. Reprod. Fertil. Dev. 2020, 32, 610. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Du, G.; Zhang, W.; Huang, H.; Chen, D.; Wu, D.; Wang, X. Short-Term Neonatal/Prepubertal Exposure of Dibutyl Phthalate (DBP) Advanced Pubertal Timing and Affected Hypothalamic Kisspeptin/GPR54 Expression Differently in Female Rats. Toxicology 2013, 314, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.J.M.; Grande, S.W.; Talsness, C.E.; Grote, K.; Chahoud, I. A Dose–Response Study Following in Utero and Lactational Exposure to Di-(2-Ethylhexyl)-Phthalate (DEHP): Non-Monotonic Dose–Response and Low Dose Effects on Rat Brain Aromatase Activity. Toxicology 2006, 227, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Shibutani, M.; Lee, K.-Y.; Masutomi, N.; Fujita, H.; Inoue, K.; Mitsumori, K.; Hirose, M. Impact of Maternal Dietary Exposure to Endocrine-Acting Chemicals on Progesterone Receptor Expression in Microdissected Hypothalamic Medial Preoptic Areas of Rat Offspring. Toxicol. Appl. Pharmacol. 2005, 208, 127–136. [Google Scholar] [CrossRef]
- Shao, P.; Wang, Y.; Zhang, M.; Wen, X.; Zhang, J.; Xu, Z.; Hu, M.; Jiang, J.; Liu, T. The Interference of DEHP in Precocious Puberty of Females Mediated by the Hypothalamic IGF-1/PI3K/Akt/MTOR Signaling Pathway. Ecotoxicol. Environ. Saf. 2019, 181, 362–369. [Google Scholar] [CrossRef]
- Kougias, D.G.; Cortes, L.R.; Moody, L.; Rhoads, S.; Pan, Y.-X.; Juraska, J.M. Effects of Perinatal Exposure to Phthalates and a High-Fat Diet on Maternal Behavior and Pup Development and Social Play. Endocrinology 2018, 159, 1088–1105. [Google Scholar] [CrossRef] [Green Version]
- Roepke, T.A.; Yang, J.A.; Yasrebi, A.; Mamounis, K.J.; Oruc, E.; Zama, A.M.; Uzumcu, M. Regulation of Arcuate Genes by Developmental Exposures to Endocrine-Disrupting Compounds in Female Rats. Reprod. Toxicol. 2016, 62, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, D.; Martinez–Arguelles, D.B.; Campioli, E.; Lee, S.; Papadopoulos, V. In Utero Exposure to the Endocrine Disruptor Di(2-Ethylhexyl) Phthalate Targets Ovarian Theca Cells and Steroidogenesis in the Adult Female Rat. Reprod. Toxicol. 2015, 51, 47–56. [Google Scholar] [CrossRef]
- Liu, T.; Jia, Y.; Zhou, L.; Wang, Q.; Sun, D.; Xu, J.; Wu, J.; Chen, H.; Xu, F.; Ye, L. Effects of Di-(2-Ethylhexyl) Phthalate on the Hypothalamus–Uterus in Pubertal Female Rats. Int. J. Environ. Res. Public. Health 2016, 13, 1130. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Wang, Y.; Yang, M.; Shao, P.; Duan, L.; Li, M.; Zhu, M.; Yang, J.; Jiang, J. Di-(2-Ethylhexyl) Phthalate Induces Precocious Puberty in Adolescent Female Rats. Iran. J. Basic Med. Sci. 2018, 21, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Funabashi, T.; Kawaguchi, M.; Kimura, F. The Endocrine Disrupters Butyl Benzyl Phthalate and Bisphenol A Increase the Expression of Progesterone Receptor Messenger Ribonucleic Acid in the Preoptic Area of Adult Ovariectomized Rats. Neuroendocrinology 2001, 74, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Li, N.; Zhu, J.; Yu, G.; Guo, K.; Zhou, L.; Zheng, D.; Qu, X.; Huang, J.; Chen, X.; et al. Effects of Di-(2-Ethylhexyl) Phthalate on the Hypothalamus-Pituitary-Ovarian Axis in Adult Female Rats. Reprod. Toxicol. 2014, 46, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; Lin, H.-T.; Chaudhary, M.A.; Lee, Y.-C.; Wang, D.-C. Effects of Prenatal Phthalate Exposure and Childhood Exercise on Maternal Behaviors in Female Rats at Postpartum: A Role of Oxtr Methylation in the Hypothalamus. Int. J. Mol. Sci. 2021, 22, 9847. [Google Scholar] [CrossRef] [PubMed]
- Boberg, J.; Christiansen, S.; Axelstad, M.; Kledal, T.S.; Vinggaard, A.M.; Dalgaard, M.; Nellemann, C.; Hass, U. Reproductive and Behavioral Effects of Diisononyl Phthalate (DINP) in Perinatally Exposed Rats. Reprod. Toxicol. 2011, 31, 200–209. [Google Scholar] [CrossRef] [Green Version]
- Quinnies, K.M.; Harris, E.P.; Snyder, R.W.; Sumner, S.S.; Rissman, E.F. Direct and Transgenerational Effects of Low Doses of Perinatal Di-(2-Ethylhexyl) Phthalate (DEHP) on Social Behaviors in Mice. PLoS ONE 2017, 12, e0171977. [Google Scholar] [CrossRef]
- Lee, S.M.; Jeon, S.; Jeong, H.J.; Kim, B.-N.; Kim, Y. Dibutyl Phthalate Exposure during Gestation and Lactation in C57BL/6 Mice: Maternal Behavior and Neurodevelopment in Pups. Environ. Res. 2020, 182, 109025. [Google Scholar] [CrossRef] [PubMed]
- Carbone, S.; Ponzo, O.J.; Gobetto, N.; Samaniego, Y.A.; Reynoso, R.; Scacchi, P.; Moguilevsky, J.A.; Cutrera, R. Antiandrogenic Effect of Perinatal Exposure to the Endocrine Disruptor Di-(2-Ethylhexyl) Phthalate Increases Anxiety-like Behavior in Male Rats during Sexual Maturation. Horm. Behav. 2013, 63, 692–699. [Google Scholar] [CrossRef]
- Sellinger, E.P.; Kougias, D.G.; Drzewiecki, C.M.; Juraska, J.M. Behavioral Effects in Adult Rats Exposed to Low Doses of a Phthalate Mixture during the Perinatal or Adolescent Period. Neurotoxicol. Teratol. 2020, 79, 106886. [Google Scholar] [CrossRef]
- Wang, D.-C.; Chen, T.-J.; Lin, M.-L.; Jhong, Y.-C.; Chen, S.-C. Exercise Prevents the Increased Anxiety-like Behavior in Lactational Di-(2-Ethylhexyl) Phthalate-Exposed Female Rats in Late Adolescence by Improving the Regulation of Hypothalamus-Pituitary-Adrenal Axis. Horm. Behav. 2014, 66, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xu, X.; Zhu, Q. Pubertal Exposure to Di-(2-Ethylhexyl) Phthalate Influences Social Behavior and Dopamine Receptor D2 of Adult Female Mice. Chemosphere 2016, 144, 1771–1779. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yang, Y.; Wang, R.; Wang, Y.; Ruan, Q.; Lu, Y. Perinatal Exposure to Di-(2-Ethylhexyl) Phthalate Affects Anxiety- and Depression-like Behaviors in Mice. Chemosphere 2015, 124, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.-L.; Shen, R.; Sun, C.-C.; Zhang, S.-Y.; Wang, B.; Sun, M.-F.; Xu, D.-X. Prenatal Di-(2-Ethylhexyl) Phthalate Maternal Exposure Impairs the Spatial Memory of Adult Mouse Offspring in a Phase- and Gender-Dependent Manner. Environ. Sci. Pollut. Res. 2020, 27, 28267–28275. [Google Scholar] [CrossRef] [PubMed]
- Hatcher, K.M.; Willing, J.; Chiang, C.; Rattan, S.; Flaws, J.A.; Mahoney, M.M. Exposure to Di-(2-Ethylhexyl) Phthalate Transgenerationally Alters Anxiety-like Behavior and Amygdala Gene Expression in Adult Male and Female Mice. Physiol. Behav. 2019, 207, 7–14. [Google Scholar] [CrossRef]
- Quinnies, K.M.; Doyle, T.J.; Kim, K.H.; Rissman, E.F. Transgenerational Effects of Di-(2-Ethylhexyl) Phthalate (DEHP) on Stress Hormones and Behavior. Endocrinology 2015, 156, 3077–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Y.; Yang, Y.; Xu, X.; Hu, Y. Effects of Uterine and Lactational Exposure to Di-(2-Ethylhexyl) Phthalate on Spatial Memory and NMDA Receptor of Hippocampus in Mice. Horm. Behav. 2015, 71, 41–48. [Google Scholar] [CrossRef]
- Li, X.-J.; Jiang, L.; Chen, L.; Chen, H.-S.; Li, X. Neurotoxicity of Dibutyl Phthalate in Brain Development Following Perinatal Exposure: A Study in Rats. Environ. Toxicol. Pharmacol. 2013, 36, 392–402. [Google Scholar] [CrossRef]
- Li, Y.; Zhuang, M.; Li, T.; Shi, N. Neurobehavioral Toxicity Study of Dibutyl Phthalate on Rats Following in Utero and Lactational Exposure. J. Appl. Toxicol. 2009, 29, 603–611. [Google Scholar] [CrossRef]
- DeBartolo, D.; Jayatilaka, S.; Yan Siu, N.; Rose, M.; Ramos, R.L.; Betz, A.J. Perinatal Exposure to Benzyl Butyl Phthalate Induces Alterations in Neuronal Development/Maturation Protein Expression, Estrogen Responses, and Fear Conditioning in Rodents. Behav. Pharmacol. 2016, 27, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Bastos Sales, L.; van Esterik, J.C.J.; Hodemaekers, H.M.; Lamoree, M.H.; Hamers, T.; van der Ven, L.T.M.; Legler, J. Analysis of Lipid Metabolism, Immune Function, and Neurobehavior in Adult C57BL/6JxFVB Mice After Developmental Exposure to Di (2-Ethylhexyl) Phthalate. Front. Endocrinol. 2018, 9, 684. [Google Scholar] [CrossRef] [PubMed]
- Kougias, D.G.; Sellinger, E.P.; Willing, J.; Juraska, J.M. Perinatal Exposure to an Environmentally Relevant Mixture of Phthalates Results in a Lower Number of Neurons and Synapses in the Medial Prefrontal Cortex and Decreased Cognitive Flexibility in Adult Male and Female Rats. J. Neurosci. 2018, 38, 6864–6872. [Google Scholar] [CrossRef] [PubMed]
- Sellinger, E.P.; Riesgo, V.R.; Brinks, A.S.; Willing, J.; Juraska, J.M. Perinatal Phthalate Exposure Increases Developmental Apoptosis in the Rat Medial Prefrontal Cortex. NeuroToxicology 2021, 87, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Fu, H.; Fu, Y.; You, M.; Li, X.; Wang, C.; Leng, K.; Wang, Y.; Chen, J. Maternal Exposure to Di-(2-Ethylhexyl) Phthalate Impairs Hippocampal Synaptic Plasticity in Male Offspring: Involvement of Damage to Dendritic Spine Development. ACS Chem. Neurosci. 2021, 12, 311–322. [Google Scholar] [CrossRef]
- Wang, J.; Li, X.; Wang, C.; Li, Y.; Wang, J.; Fang, R.; Wang, J.; Chen, J.; Dong, J. Exposure to Di-(2-Ethylhexyl) Phthalate Reduces Secretion of GDNF via Interfering with Estrogen Pathway and Downregulating ERK/c-Fos Signaling Pathway in Astrocytes. Food Chem. Toxicol. 2021, 158, 112592. [Google Scholar] [CrossRef]
- You, M.; Dong, J.; Fu, Y.; Cong, Z.; Fu, H.; Wei, L.; Wang, Y.; Wang, Y.; Chen, J. Exposure to Di-(2-Ethylhexyl) Phthalate During Perinatal Period Gender-Specifically Impairs the Dendritic Growth of Pyramidal Neurons in Rat Offspring. Front. Neurosci. 2018, 12, 444. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; MacDonald, A.; Holahan, M.R. Acute Postnatal Exposure to Di(2-Ethylhexyl) Phthalate Adversely Impacts Hippocampal Development in the Male Rat. Neuroscience 2011, 193, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.A.; Holahan, M.R. Reduced Hippocampal Dendritic Spine Density and BDNF Expression Following Acute Postnatal Exposure to Di(2-Ethylhexyl) Phthalate in Male Long Evans Rats. PLoS ONE 2014, 9, e109522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holahan, M.R.; Smith, C.A.; Luu, B.E.; Storey, K.B. Preadolescent Phthalate (DEHP) Exposure Is Associated with Elevated Locomotor Activity and Reward-Related Behavior and a Reduced Number of Tyrosine Hydroxylase Positive Neurons in Post-Adolescent Male and Female Rats. Toxicol. Sci. 2018, 165, 512–530. [Google Scholar] [CrossRef]
- Ding, Y.; Lu, L.; Xuan, C.; Han, J.; Ye, S.; Cao, T.; Chen, W.; Li, A.; Zhang, X. Di-n-Butyl Phthalate Exposure Negatively Influences Structural and Functional Neuroplasticity via Rho-GTPase Signaling Pathways. Food Chem. Toxicol. 2017, 105, 34–43. [Google Scholar] [CrossRef]
- Sangeetha, S.; Vimalkumar, K.; Loganathan, B.G. Environmental Contamination and Human Exposure to Select Endocrine-Disrupting Chemicals: A Review. Sustain. Chem. 2021, 2, 20. [Google Scholar] [CrossRef]
- Picot, M.; Naulé, L.; Marie-Luce, C.; Martini, M.; Raskin, K.; Grange-Messent, V.; Franceschini, I.; Keller, M.; Mhaouty-Kodja, S. Vulnerability of the Neural Circuitry Underlying Sexual Behavior to Chronic Adult Exposure to Oral Bisphenol A in Male Mice. Endocrinology 2014, 155, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capela, D.; Dombret, C.; Poissenot, K.; Poignant, M.; Malbert-Colas, A.; Franceschini, I.; Keller, M.; Mhaouty-Kodja, S. Adult Male Mice Exposure to Nonylphenol Alters Courtship Vocalizations and Mating. Sci. Rep. 2018, 8, 2988. [Google Scholar] [CrossRef] [PubMed]
- Capela, D.; Poissenot, K.; Dombret, C.; Keller, M.; Franceschini, I.; Mhaouty-Kodja, S. Effects of Combined Exposure of Adult Male Mice to Di-(2-Ethylexyl)Phthalate and Nonylphenol on Behavioral and Neuroendocrine Responses. Chemosphere 2019, 221, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Ahmadpour, D.; Mhaouty-Kodja, S.; Grange-Messent, V. Disruption of the Blood-Brain Barrier and Its Close Environment Following Adult Exposure to Low Doses of Di(2-Ethylhexyl)Phthalate Alone or in an Environmental Phthalate Mixture in Male Mice. Chemosphere 2021, 282, 131013. [Google Scholar] [CrossRef]
- Ahmadpour, D.; Mhaouty-Kodja, S.; Grange-Messent, V. Effects and Underlying Cellular Pathway Involved in the Impairment of the Neurovascular Unit Following Exposure of Adult Male Mice to Low Doses of Di(2-Ethylhexyl) Phthalate Alone or in an Environmental Phthalate Mixture. Environ. Res. 2021, 207, 112235. [Google Scholar] [CrossRef]
- Barrett, E.S.; Parlett, L.E.; Wang, C.; Drobnis, E.Z.; Bruce Redmon, J.; Swan, S.H. Environmental Exposure to Di-2-Ethylhexyl Phthalate Is Associated with Low Interest in Sexual Activity in Premenopausal Women. Horm. Behav. 2014, 66, 787–792. [Google Scholar] [CrossRef] [Green Version]
- Hlisníková, H.; Petrovičová, I.; Kolena, B.; Šidlovská, M.; Sirotkin, A. Effects and Mechanisms of Phthalates’ Action on Neurological Processes and Neural Health: A Literature Review. Pharmacol. Rep. 2021, 73, 386–404. [Google Scholar] [CrossRef]
- Ejaredar, M.; Nyanza, E.C.; Ten Eycke, K.; Dewey, D. Phthalate Exposure and Childrens Neurodevelopment: A Systematic Review. Environ. Res. 2015, 142, 51–60. [Google Scholar] [CrossRef]
- Xu, H.; Wu, X.; Liang, C.; Shen, J.; Tao, S.; Wen, X.; Liu, W.; Zou, L.; Yang, Y.; Xie, Y.; et al. Association of Urinary Phthalates Metabolites Concentration with Emotional Symptoms in Chinese University Students. Environ. Pollut. 2020, 262, 114279. [Google Scholar] [CrossRef]
- Kim, K.-N.; Choi, Y.-H.; Lim, Y.-H.; Hong, Y.-C. Urinary Phthalate Metabolites and Depression in an Elderly Population: National Health and Nutrition Examination Survey 2005–2012. Environ. Res. 2016, 145, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Borrow, A.P.; Handa, R.J. Estrogen Receptors Modulation of Anxiety-Like Behavior. In Vitamins and Hormones; Elsevier: Amsterdam, The Netherlands, 2017; Volume 103, pp. 27–52. ISBN 978-0-12-811914-3. [Google Scholar]
- Hsu, C.-M.K.; Ney, L.J.; Honan, C.; Felmingham, K.L. Gonadal Steroid Hormones and Emotional Memory Consolidation: A Systematic Review and Meta-Analysis. Neurosci. Biobehav. Rev. 2021, 130, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Szoeke, C.; Downie, S.J.; Parker, A.F.; Phillips, S. Sex Hormones, Vascular Factors and Cognition. Front. Neuroendocrinol. 2021, 62, 100927. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).