
Microfluidic Electroceuticals Platform for Therapeutic Strategies of Intervertebral Disc Degeneration: Effects of Electrical Stimulation on Human Nucleus Pulposus Cells under Inflammatory Conditions


Abstract
:1. Introduction
2. Results
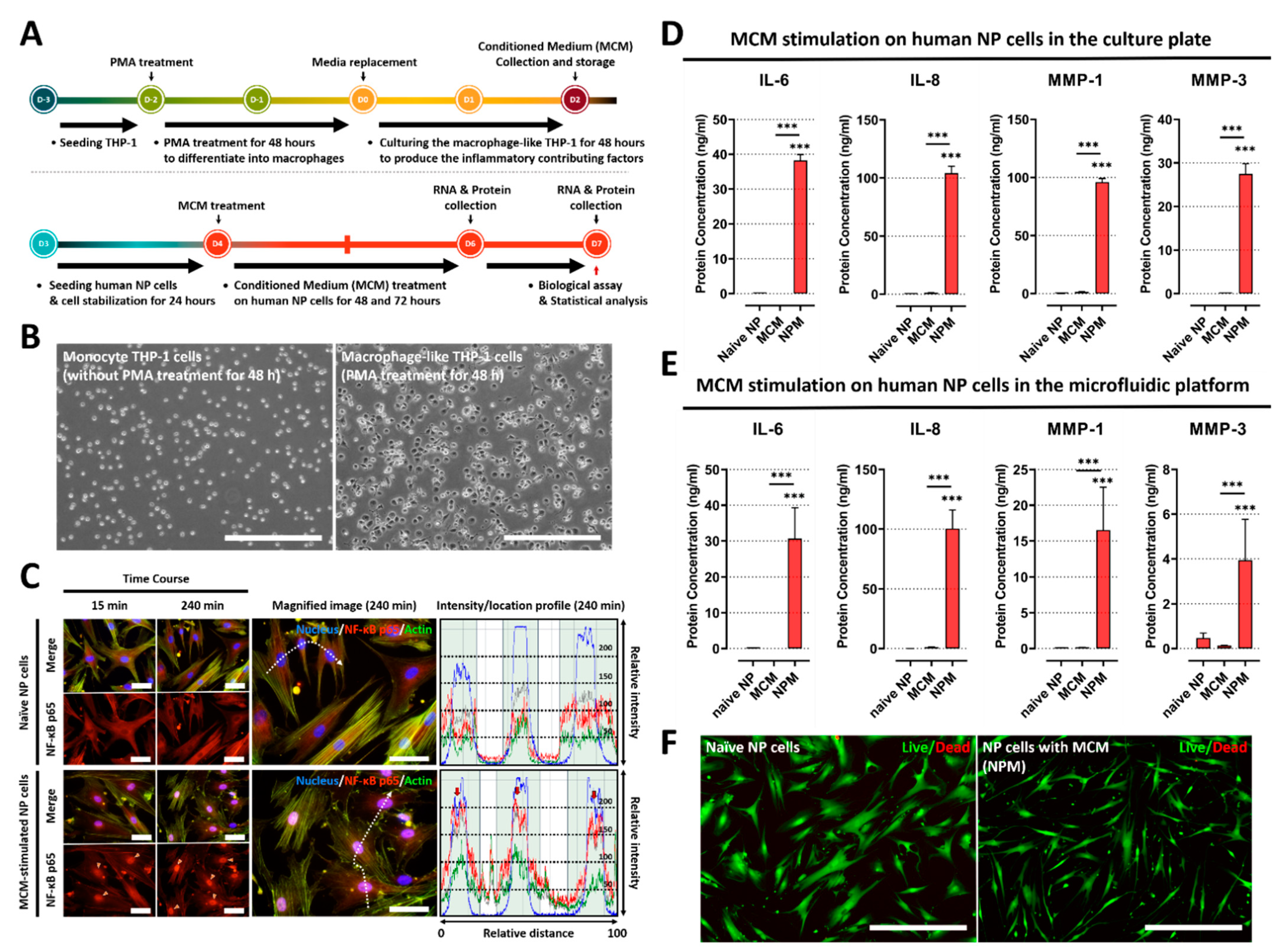
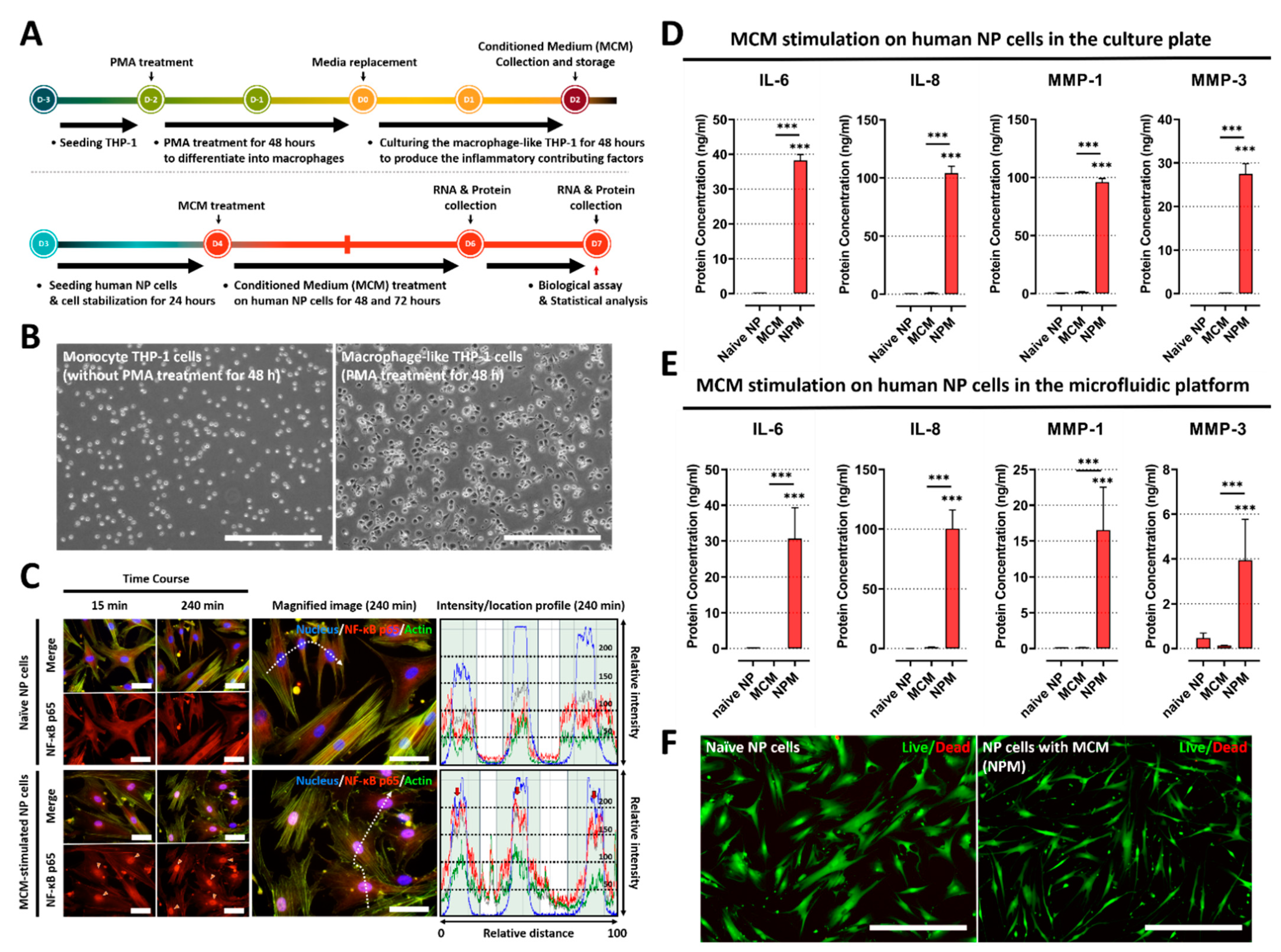
2.1. Conditioned Medium from Macrophage THP-1 Cells Induces the Inflammatory Response by Modulating Protein Expression of Inflammatory Mediators and ECM-Modifying Enzymes in Human NP Cells
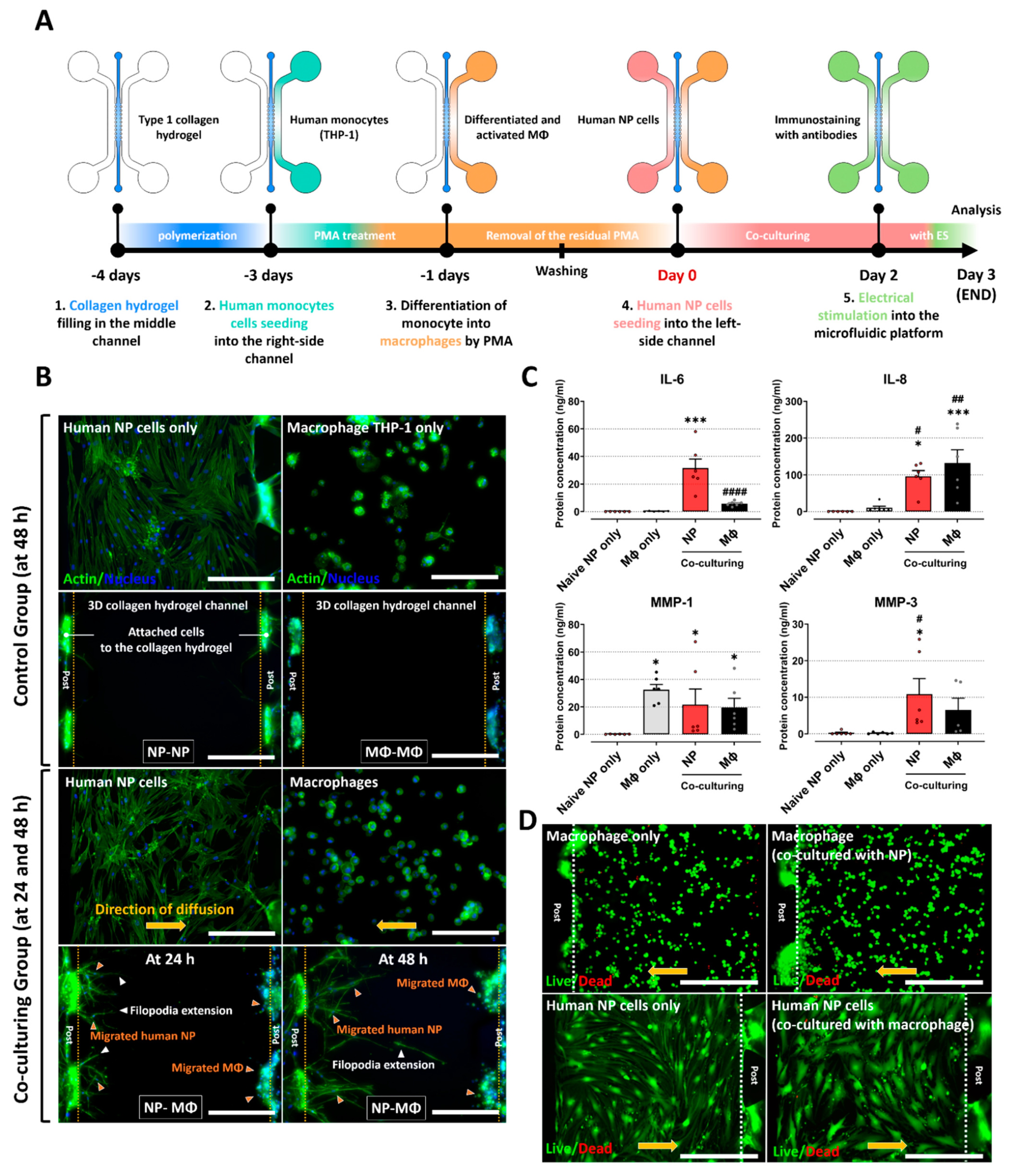
2.2. Inflammatory Responses between Human NP and Macrophage THP-1 Cells through 3-Dimensional Migration in Microfluidic Platform
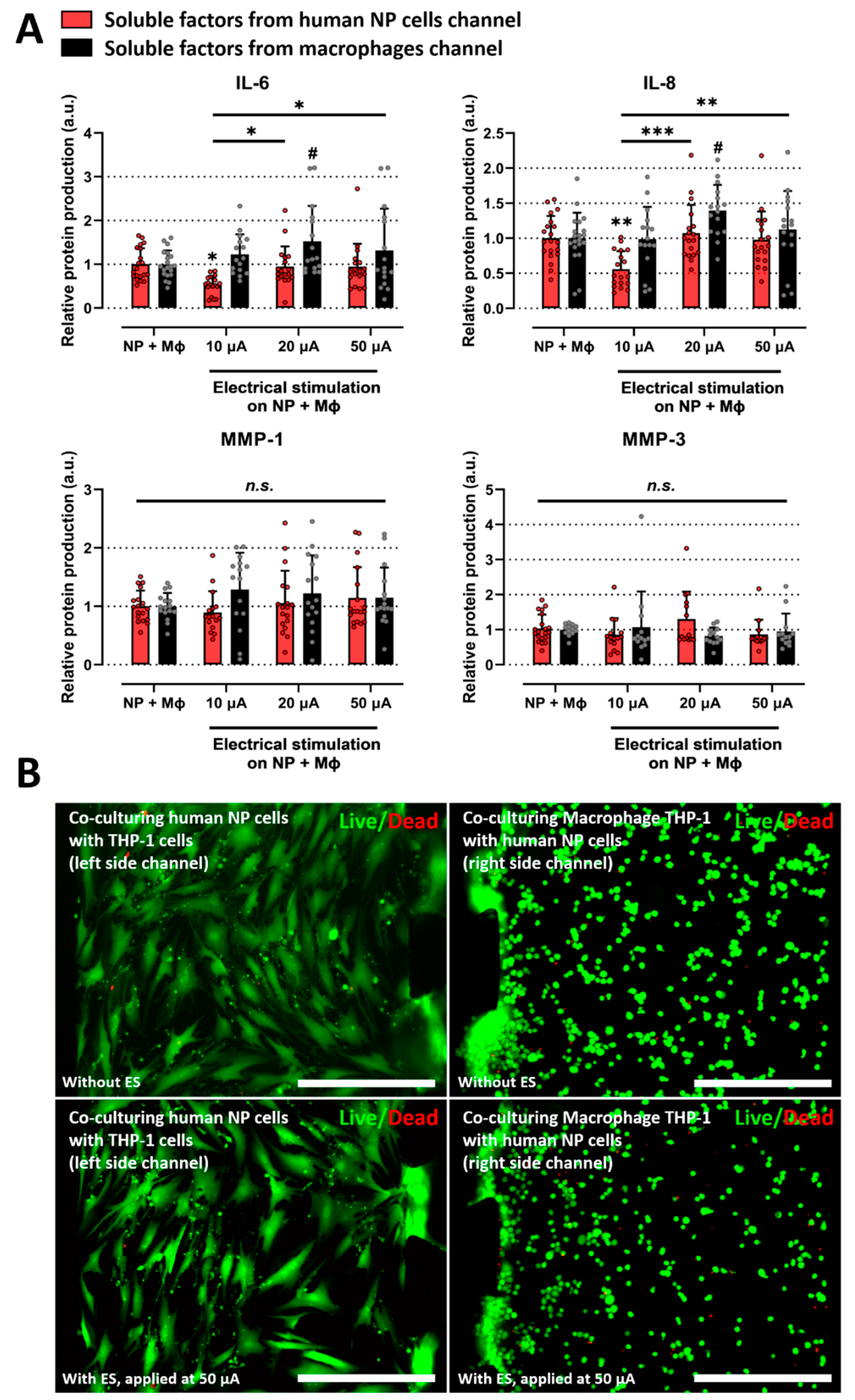
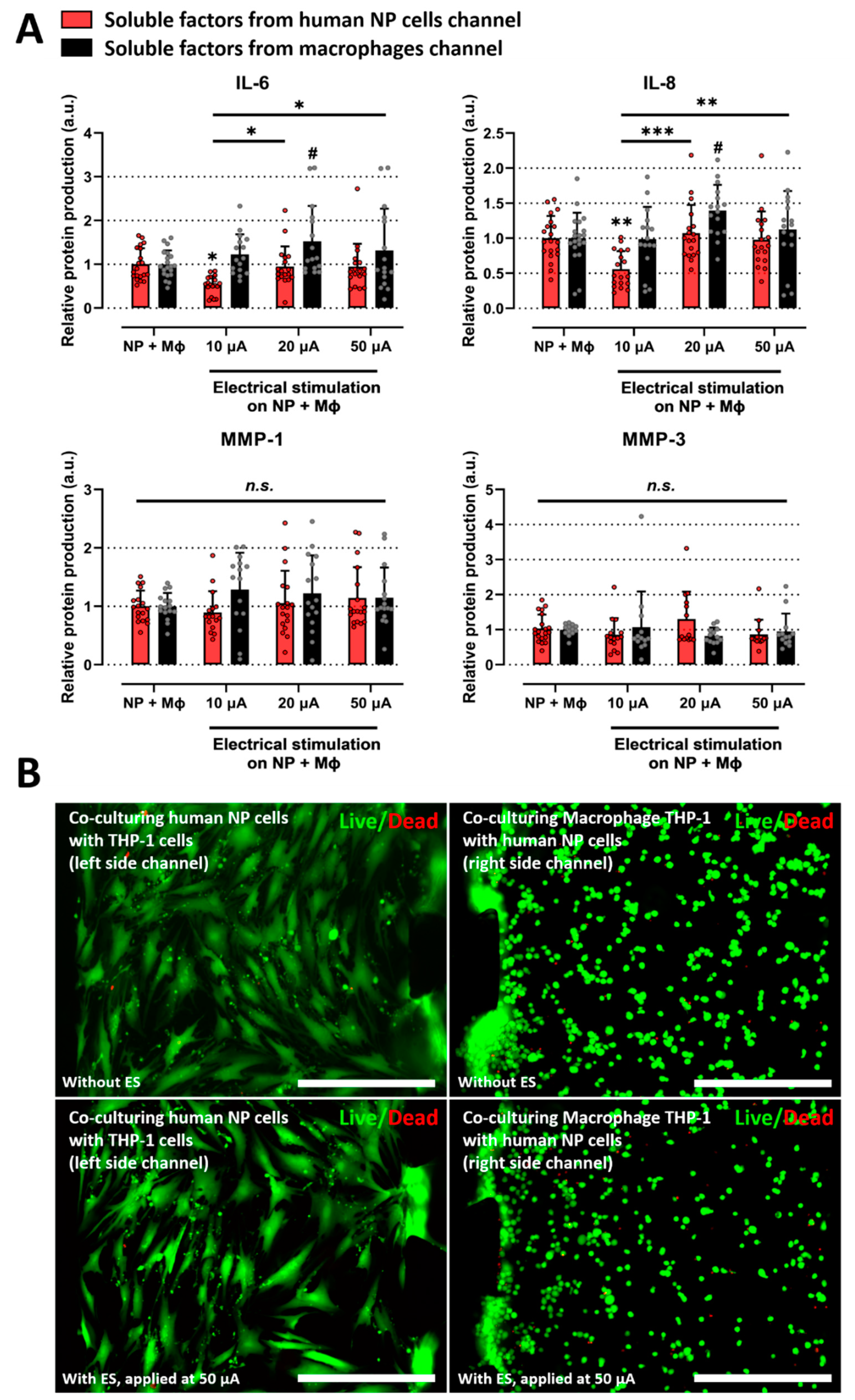
2.3. Electrical Stimulation Alleviates the Inflammatory Response Induced by Interaction between Human NP and Macrophages on Microfluidic Electrode Platform
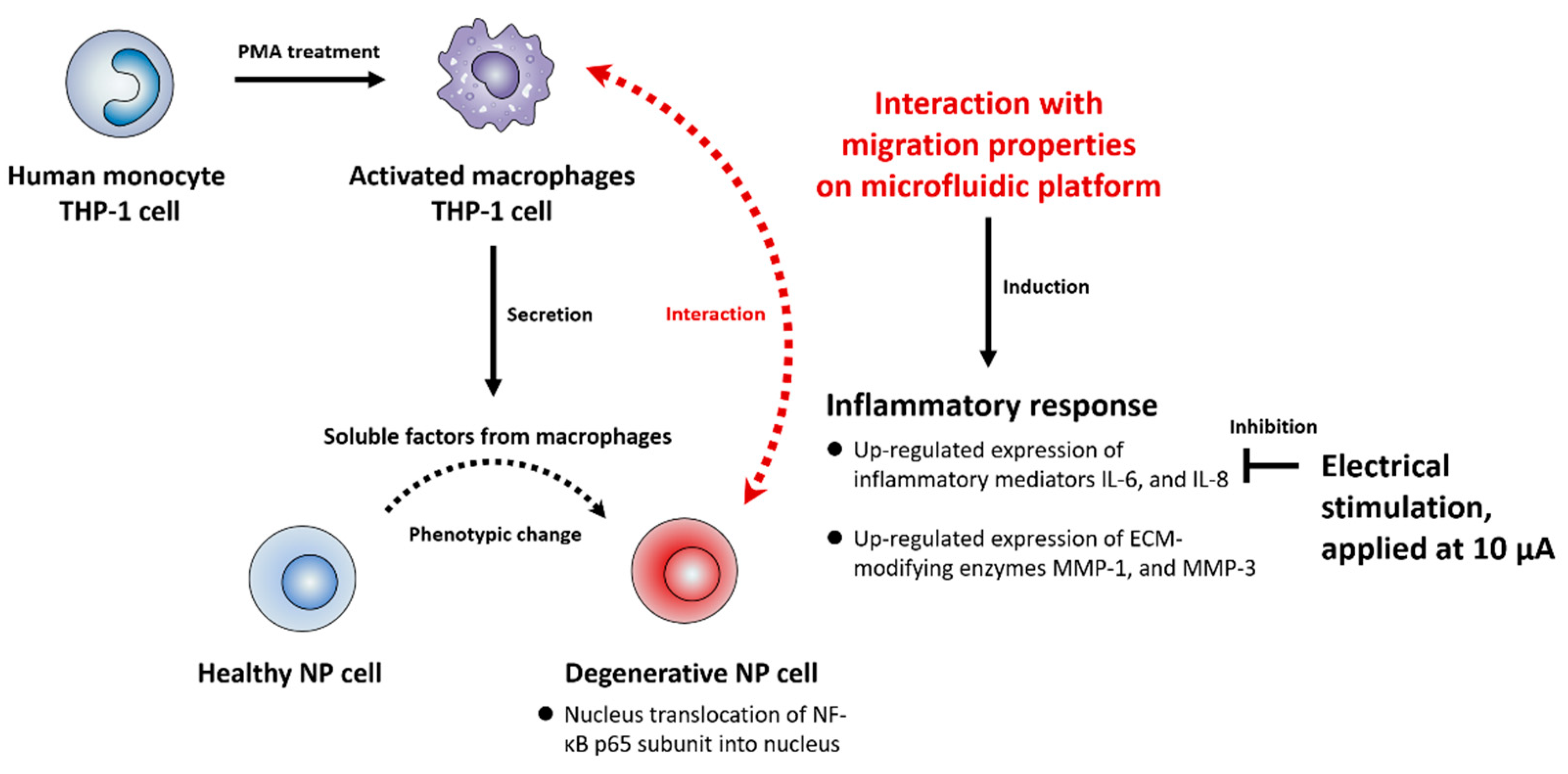
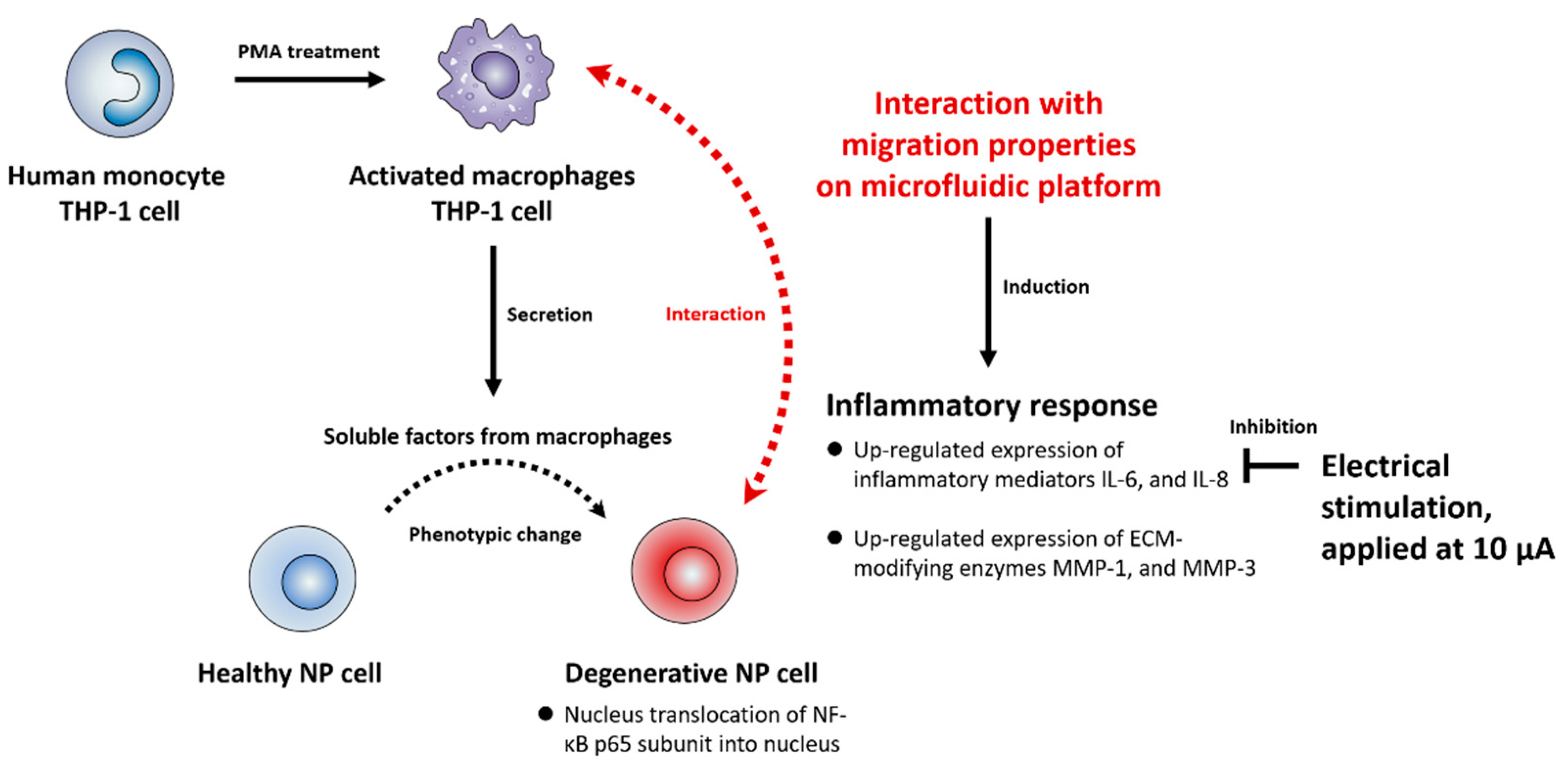
3. Discussion
4. Materials and Methods
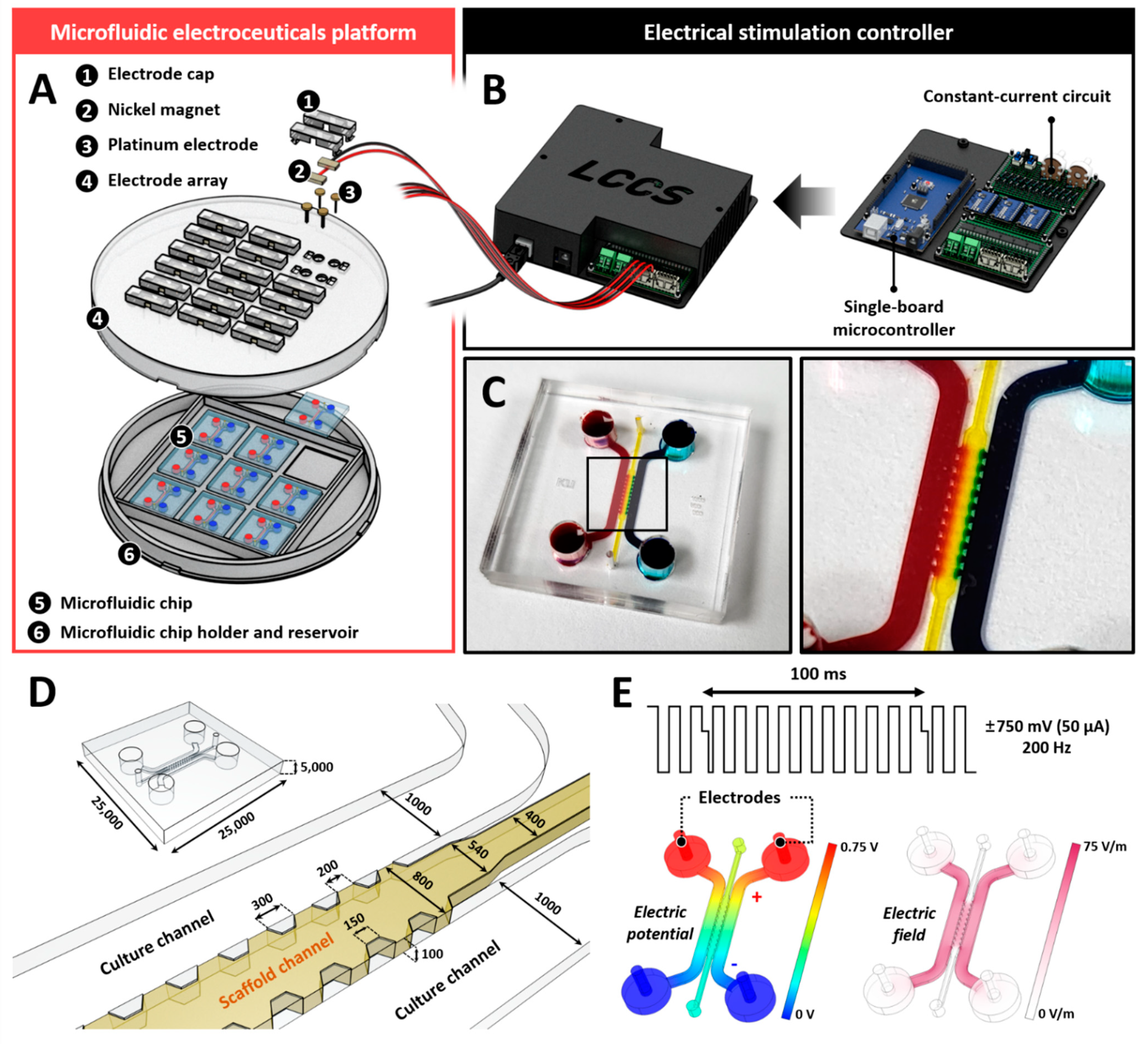
4.1. Fabrication of Microfluidic Electrode Platform
4.2. Culture of Primary Human NP Cells
4.3. Culture of Immortalized Human Monocyte THP-1 Cells and Production of Conditioned Medium
4.4. Generation of Paracrine Signal in the Microfluidic Platform
4.5. Coculturing of Human NP Cells with THP-1 Cells in Microfluidic Platform
4.6. Immunofluorescence Staining
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Cell Cytotoxicity and Lactate Dehydrogenase Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dowdell, J.; Erwin, M.; Choma, T.; Vaccaro, A.; Iatridis, J.; Cho, S.K. Intervertebral Disk Degeneration and Repair. Neurosurgery 2017, 80, S46–S54. [Google Scholar] [CrossRef]
- García-Cosamalón, J.; del Valle, M.E.; Calavia, M.G.; García-Suárez, O.; López-Muñiz, A.; Otero, J.; Vega, J.A. Intervertebral disc, sensory nerves and neurotrophins: Who is who in discogenic pain? J. Anat. 2010, 217, 1–15. [Google Scholar] [CrossRef]
- Hwang, M.H.; Son, H.-G.; Kim, J.; Choi, H. In vitro model of distinct catabolic and inflammatory response patterns of endothelial cells to intervertebral disc cell degeneration. Sci. Rep. 2020, 10, 20596. [Google Scholar] [CrossRef]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; O’Brien, J.; Jayson, M.I. Nerve ingrowth into diseased intervertebral disc in chronic back pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Freemont, A.J.; Watkins, A.; Le Maitre, C.; Baird, P.; Jeziorska, M.; Knight, M.T.; Ross, E.R.; O’Brien, J.P.; Hoyland, J.A. Nerve growth factor expression and innervation of the painful intervertebral disc. J. Pathol. 2002, 197, 286–292. [Google Scholar] [CrossRef]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef]
- Ye, F.; Lyu, F.-J.; Wang, H.; Zheng, Z. The involvement of immune system in intervertebral disc herniation and degeneration. JOR Spine 2022, 5, e1196. [Google Scholar] [CrossRef]
- Hwang, M.H.; Son, H.G.; Lee, J.W.; Yoo, C.M.; Shin, J.H.; Nam, H.G.; Lim, H.J.; Baek, S.M.; Park, J.H.; Kim, J.H. Phototherapy suppresses inflammation in human nucleus pulposus cells for intervertebral disc degeneration. Lasers Med. Sci. 2018, 33, 1055–1064. [Google Scholar] [CrossRef]
- Hwang, M.H.; Son, H.G.; Lee, J.W.; Yoo, C.M.; Shin, J.H.; Nam, H.G.; Lim, H.J.; Baek, S.M.; Park, J.H.; Kim, J.H.; et al. Photobiomodulation of extracellular matrix enzymes in human nucleus pulposus cells as a potential treatment for intervertebral disk degeneration. Sci. Rep. 2018, 8, 11654. [Google Scholar] [CrossRef] [Green Version]
- Balint, R.; Cassidy, N.J.; Cartmell, S.H. Electrical stimulation: A novel tool for tissue engineering. Tissue Eng. Part. B Rev. 2013, 19, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Bai, X.; Ding, Y.; Lee, I.-S. Electrical stimulation as a novel tool for regulating cell behavior in tissue engineering. Biomater. Res. 2019, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gordon, T.; English, A.W. Strategies to promote peripheral nerve regeneration: Electrical stimulation and/or exercise. Eur. J. Neurosci. 2016, 43, 336–350. [Google Scholar] [CrossRef] [PubMed]
- Hiemer, B.; Krogull, M.; Bender, T.; Ziebart, J.; Krueger, S.; Bader, R.; Jonitz-Heincke, A. Effect of electric stimulation on human chondrocytes and mesenchymal stem cells under normoxia and hypoxia. Mol. Med. Rep. 2018, 18, 2133–2141. [Google Scholar] [CrossRef]
- Love, M.R.; Palee, S.; Chattipakorn, S.C.; Chattipakorn, N. Effects of electrical stimulation on cell proliferation and apoptosis. J. Cell. Physiol. 2018, 233, 1860–1876. [Google Scholar] [CrossRef]
- Vaca-González, J.J.; Guevara, J.M.; Moncayo, M.A.; Castro-Abril, H.; Hata, Y.; Garzón-Alvarado, D.A. Biophysical stimuli: A review of electrical and mechanical stimulation in hyaline cartilage. Cartilage 2019, 10, 157–172. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Z.; Zhang, G.; Clark, C.C.; Brighton, C.T. Up-regulation of chondrocyte matrix genes and products by electric fields. Clin. Orthop. Relat. Res. 2004, 427, S163–S173. [Google Scholar] [CrossRef]
- Zhao, S.; Mehta, A.S.; Zhao, M. Biomedical applications of electrical stimulation. Cell. Mol. Life Sci. 2020, 77, 2681–2699. [Google Scholar] [CrossRef]
- Kubelt, C.; Molkewehrum, H.; Lucius, R.; Synowitz, M.; Held-Feindt, J.; Helmers, A.K. Influence of Simulated Deep Brain Stimulation on the Expression of Inflammatory Mediators by Human Central Nervous System Cells In Vitro. Neuromolecular. Med. 2022, 24, 169–182. [Google Scholar] [CrossRef]
- Wang, Z.; Hutton, W.C.; Yoon, S.T. The effect of capacitively coupled (CC) electrical stimulation on human disc nucleus pulposus cells and the relationship between CC and BMP-7. Eur. Spine J. 2017, 26, 240–247. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, H.; Suh, M.J.; Shin, J.H.; Hwang, M.H.; Lee, H.-M. Effect of biphasic electrical current stimulation on IL-1β–stimulated annulus fibrosus cells using in vitro microcurrent generating chamber system. Spine 2013, 38, E1368–E1376. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.; Hwang, M.; Back, S.; Nam, H.; Yoo, C.; Park, J.; Son, H.; Lee, J.; Lim, H.; Lee, K. Electrical impulse effects on degenerative human annulus fibrosus model to reduce disc pain using micro-electrical impulse-on-a-chip. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Teixeira, G.Q.; Ribeiro-Machado, C.; Pereira, C.L.; Ferreira, J.R.; Molinos, M.; Santos, S.G.; Barbosa, M.A.; Goncalves, R.M. Modulation of the In Vivo Inflammatory Response by Pro- Versus Anti-Inflammatory Intervertebral Disc Treatments. Int. J. Mol. Sci. 2020, 21, 1730. [Google Scholar] [CrossRef] [PubMed]
- Doita, M.; Kanatani, T.; Ozaki, T.; Matsui, N.; Kurosaka, M.; Yoshiya, S. Influence of macrophage infiltration of herniated disc tissue on the production of matrix metalloproteinases leading to disc resorption. Spine 2001, 26, 1522–1527. [Google Scholar] [CrossRef]
- Peng, B.; Hao, J.; Hou, S.; Wu, W.; Jiang, D.; Fu, X.; Yang, Y. Possible pathogenesis of painful intervertebral disc degeneration. Spine 2006, 31, 560–566. [Google Scholar] [CrossRef]
- Ghajar, C.M.; George, S.C.; Putnam, A. Matrix metalloproteinase control of capillary morphogenesis. Crit. Rev. Eukaryot. Gene Expr. 2008, 18, 251–278. [Google Scholar] [CrossRef]
- Moon, H.J.; Yurube, T.; Lozito, T.P.; Pohl, P.; Hartman, R.A.; Sowa, G.A.; Kang, J.D.; Vo, N. Effects of secreted factors in culture medium of annulus fibrosus cells on microvascular endothelial cells: Elucidating the possible pathomechanisms of matrix degradation and nerve in-growth in disc degeneration. Osteoarthr. Cartil. 2014, 22, 344–354. [Google Scholar] [CrossRef]
- Kim, J.H.; Studer, R.K.; Sowa, G.A.; Vo, N.V.; Kang, J.D. Activated macrophage-like THP-1 cells modulate anulus fibrosus cell production of inflammatory mediators in response to cytokines. Spine 2008, 33, 2253–2259. [Google Scholar] [CrossRef]
- Li, X.-C.; Luo, S.-J.; Fan, W.; Zhou, T.-L.; Tan, D.-Q.; Tan, R.-X.; Xian, Q.-Z.; Li, J.; Huang, C.-M.; Wang, M.-S. Macrophage polarization regulates intervertebral disc degeneration by modulating cell proliferation, inflammation mediator secretion, and extracellular matrix metabolism. Front. Immunol. 2022, 13, 1508. [Google Scholar] [CrossRef]
- Nakazawa, K.R.; Walter, B.A.; Laudier, D.M.; Krishnamoorthy, D.; Mosley, G.E.; Spiller, K.L.; Iatridis, J.C. Accumulation and localization of macrophage phenotypes with human intervertebral disc degeneration. Spine J. 2018, 18, 343–356. [Google Scholar] [CrossRef]
- Yang, C.; Cao, P.; Gao, Y.; Wu, M.; Lin, Y.; Tian, Y.; Yuan, W. Differential expression of p38 MAPK α, β, γ, δ isoforms in nucleus pulposus modulates macrophage polarization in intervertebral disc degeneration. Sci. Rep. 2016, 6, 22182. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.; Silva, A.J.; Pereira, P.; Vaz, R.; Gonçalves, R.M.; Barbosa, M.A. The inflammatory response in the regression of lumbar disc herniation. Arthritis Res. Ther. 2018, 20, 251. [Google Scholar] [CrossRef] [PubMed]
- Navone, S.; Peroglio, M.; Guarnaccia, L.; Beretta, M.; Grad, S.; Paroni, M.; Cordiglieri, C.; Locatelli, M.; Pluderi, M.; Rampini, P. Mechanical loading of intervertebral disc modulates microglia proliferation, activation, and chemotaxis. Osteoarthr. Cartil. 2018, 26, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chee, A.; Shi, P.; Adams, S.L.; Markova, D.Z.; Anderson, D.G.; Smith, H.E.; Deng, Y.; Plastaras, C.T.; An, H.S. Intervertebral Disc Cells Produce Interleukins Found in Patients with Back Pain. Am. J. Phys. Med. Rehabil 2016, 95, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.-J.; Cui, H.; Pan, H.; Mc Cheung, K.; Cao, X.; Iatridis, J.C.; Zheng, Z. Painful intervertebral disc degeneration and inflammation: From laboratory evidence to clinical interventions. Bone Res. 2021, 9, 7. [Google Scholar] [CrossRef]
- Weber, K.T.; Alipui, D.O.; Sison, C.P.; Bloom, O.; Quraishi, S.; Overby, M.C.; Levine, M.; Chahine, N.O. Serum levels of the proinflammatory cytokine interleukin-6 vary based on diagnoses in individuals with lumbar intervertebral disc diseases. Arthritis Res. 2016, 18, 3. [Google Scholar] [CrossRef]
- Le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. Localization of degradative enzymes and their inhibitors in the degenerate human intervertebral disc. J. Pathol 2004, 204, 47–54. [Google Scholar] [CrossRef]
- Aripaka, S.S.; Bech-Azeddine, R.; Jørgensen, L.M.; Mikkelsen, J.D. The expression of metalloproteinases in the lumbar disc correlates strongly with Pfirrmann MRI grades in lumbar spinal fusion patients. Brain Spine 2022, 2, 100872. [Google Scholar] [CrossRef]
- Kawamura, K.; Kano, Y. Electrical stimulation induces neurite outgrowth in PC12m3 cells via the p38 mitogen-activated protein kinase pathway. Neurosci Lett 2019, 698, 81–84. [Google Scholar] [CrossRef]
- Konstantinou, E.; Zagoriti, Z.; Pyriochou, A.; Poulas, K. Microcurrent Stimulation Triggers MAPK Signaling and TGF-β1 Release in Fibroblast and Osteoblast-Like Cell Lines. Cells 2020, 9, 1294. [Google Scholar] [CrossRef]
- Sovak, G.; Budgell, B. TENS-like Stimulation Downregulates Inflammatory Cytokines in a PC-12 Cell Line. J. Manip. Physiol. 2017, 40, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Rouabhia, M.; Zhang, Z. Pulsed electrical stimulation benefits wound healing by activating skin fibroblasts through the TGFβ1/ERK/NF-κB axis. Biochim. Biophys. Acta 2016, 1860, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Dailey, D.L.; Rakel, B.A.; Vance, C.G.T.; Liebano, R.E.; Amrit, A.S.; Bush, H.M.; Lee, K.S.; Lee, J.E.; Sluka, K.A. Transcutaneous electrical nerve stimulation reduces pain, fatigue and hyperalgesia while restoring central inhibition in primary fibromyalgia. Pain 2013, 154, 2554–2562. [Google Scholar] [CrossRef]
- Fang, J.F.; Liang, Y.; Du, J.Y.; Fang, J.Q. Transcutaneous electrical nerve stimulation attenuates CFA-induced hyperalgesia and inhibits spinal ERK1/2-COX-2 pathway activation in rats. BMC Complement. Altern Med. 2013, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Bezanilla, F. How membrane proteins sense voltage. Nat. Rev. Mol. Cell Biol. 2008, 9, 323–332. [Google Scholar] [CrossRef]
- Curtze, S.; Dembo, M.; Miron, M.; Jones, D.B. Dynamic changes in traction forces with DC electric field in osteoblast-like cells. J. Cell Sci. 2004, 117, 2721–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor (ng/mL) | Naïve Human NP | Human NP Cells Cocultured with Macrophages | |||
|---|---|---|---|---|---|
| without ES | + ES, 10 μA | + ES, 20 μA | + ES, 50 μA | ||
| IL-6 | 0.02 ± 0.02 | 31.49 ± 6.58 ### | 18.15 ± 6.21 * | 27.83 ± 5.67 | 28.54 ± 7.14 |
| IL-8 | 0.30 ± 0.14 | 96.01 ± 15.45 # | 54.12 ± 14.83 ** | 96.11 ± 13.93 | 89.65 ± 18.38 |
| MMP-1 | 0.13 ± 0.13 | 21.67 ± 11.40 # | 17.69 ± 9.05 | 15.67 ± 6.55 | 18.05 ± 8.48 |
| MMP-3 | 0.23 ± 0.49 | 10.24 ± 2.07 # | 8.76 ± 1.83 | 11.28 ± 1.91 | 8.62 ± 2.0 |
| Factor (ng/mL) | naïve macrophage | Macrophages cocultured with human NP cells | |||
| without ES | + ES, 10 μA | + ES, 20 μA | + ES, 50 μA | ||
| IL-6 | 0.20 ± 0.23 | 5.42 ± 1.08 #### | 6.33 ± 0.024 | 7.96 ± 0.49 * | 6.26 ± 0.20 |
| IL-8 | 10.17 ± 11.53 | 143.94 ± 52.95 ## | 106.50 ± 44.01 | 170.55 ± 26.82 * | 118.14 ± 46.39 |
| MMP-1 | 32.53 ± 9.58 | 15.35 ± 7.85 | 23.54 ± 12.94 | 23.81 ± 11.63 | 21.56 ± 8.05 |
| MMP-3 | 0.18 ± 0.22 | 6.57 ± 1.55 | 4.72 ± 1.03 | 5.01 ± 1.15 | 5.69 ± 1.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-W.; Kim, A.-G.; Lee, K.-H.; Hwang, M.-H.; Choi, H. Microfluidic Electroceuticals Platform for Therapeutic Strategies of Intervertebral Disc Degeneration: Effects of Electrical Stimulation on Human Nucleus Pulposus Cells under Inflammatory Conditions. Int. J. Mol. Sci. 2022, 23, 10122. https://doi.org/10.3390/ijms231710122
Kim T-W, Kim A-G, Lee K-H, Hwang M-H, Choi H. Microfluidic Electroceuticals Platform for Therapeutic Strategies of Intervertebral Disc Degeneration: Effects of Electrical Stimulation on Human Nucleus Pulposus Cells under Inflammatory Conditions. International Journal of Molecular Sciences. 2022; 23(17):10122. https://doi.org/10.3390/ijms231710122
Chicago/Turabian StyleKim, Tae-Won, An-Gi Kim, Kwang-Ho Lee, Min-Ho Hwang, and Hyuk Choi. 2022. "Microfluidic Electroceuticals Platform for Therapeutic Strategies of Intervertebral Disc Degeneration: Effects of Electrical Stimulation on Human Nucleus Pulposus Cells under Inflammatory Conditions" International Journal of Molecular Sciences 23, no. 17: 10122. https://doi.org/10.3390/ijms231710122
APA StyleKim, T.-W., Kim, A.-G., Lee, K.-H., Hwang, M.-H., & Choi, H. (2022). Microfluidic Electroceuticals Platform for Therapeutic Strategies of Intervertebral Disc Degeneration: Effects of Electrical Stimulation on Human Nucleus Pulposus Cells under Inflammatory Conditions. International Journal of Molecular Sciences, 23(17), 10122. https://doi.org/10.3390/ijms231710122