
The Complexity of the Ovine and Caprine Keratin-Associated Protein Genes

 , , ,
, , , 
Abstract
1. Introduction
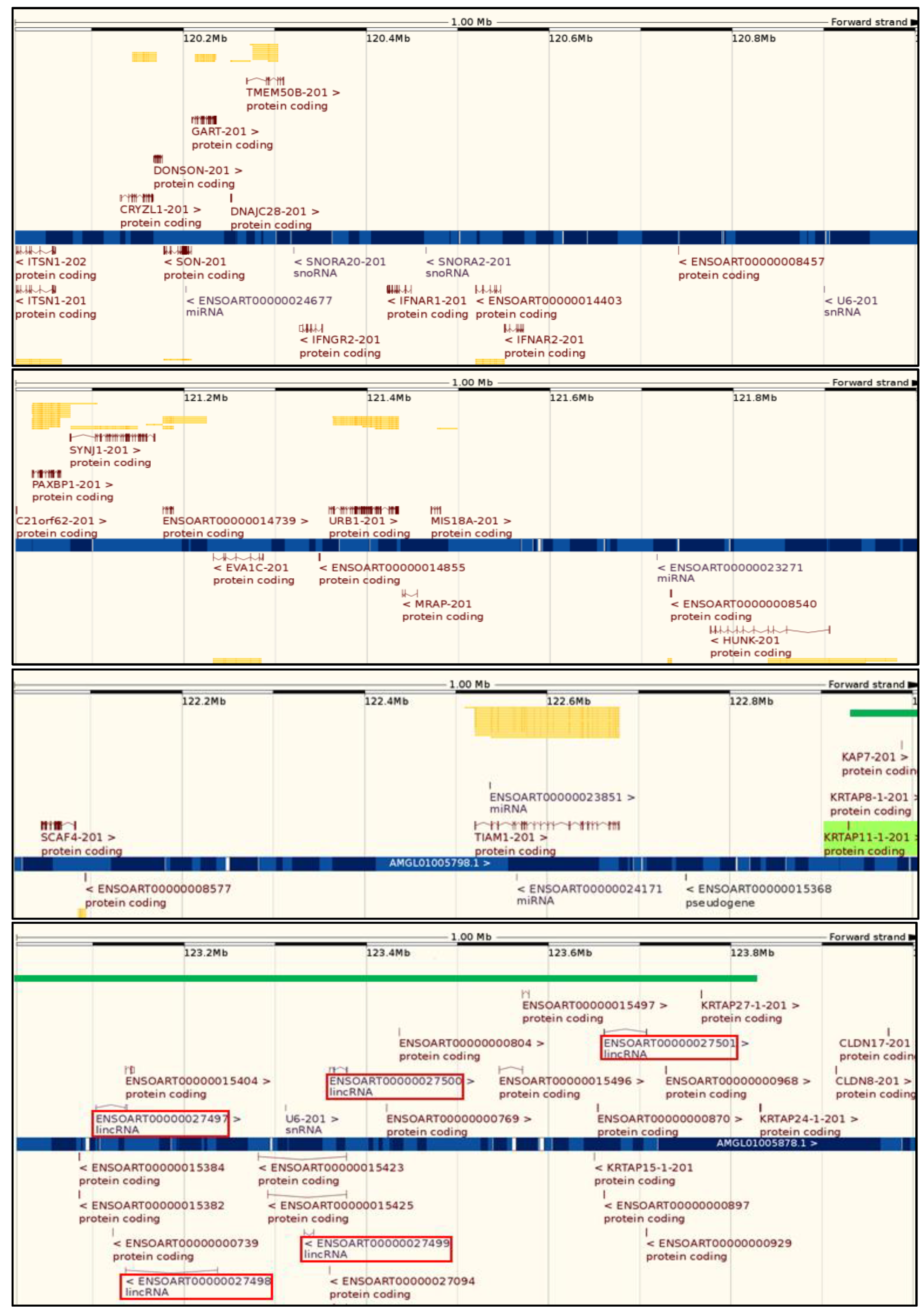
2. Keratin-Associated Proteins and the Genes That Encode Them
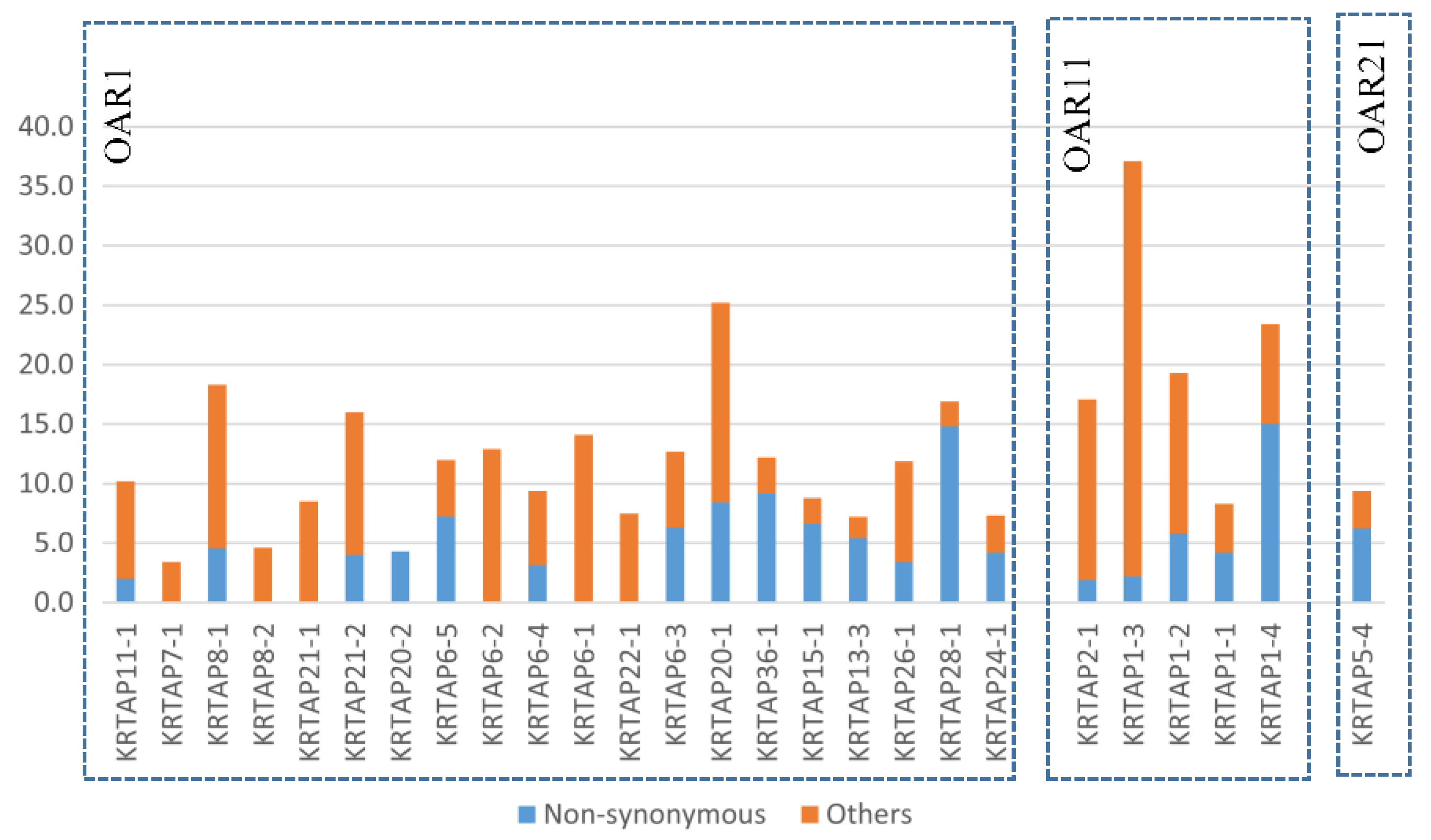
3. Variation in the KRTAPs
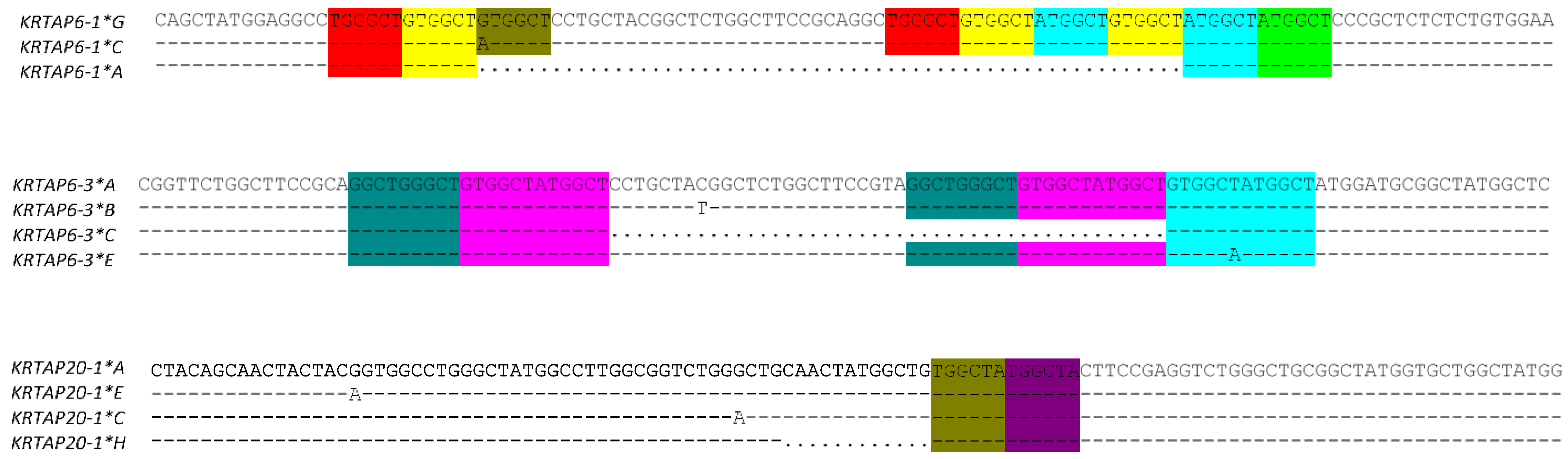
4. Mechanisms for the Generation of KRTAP Variation
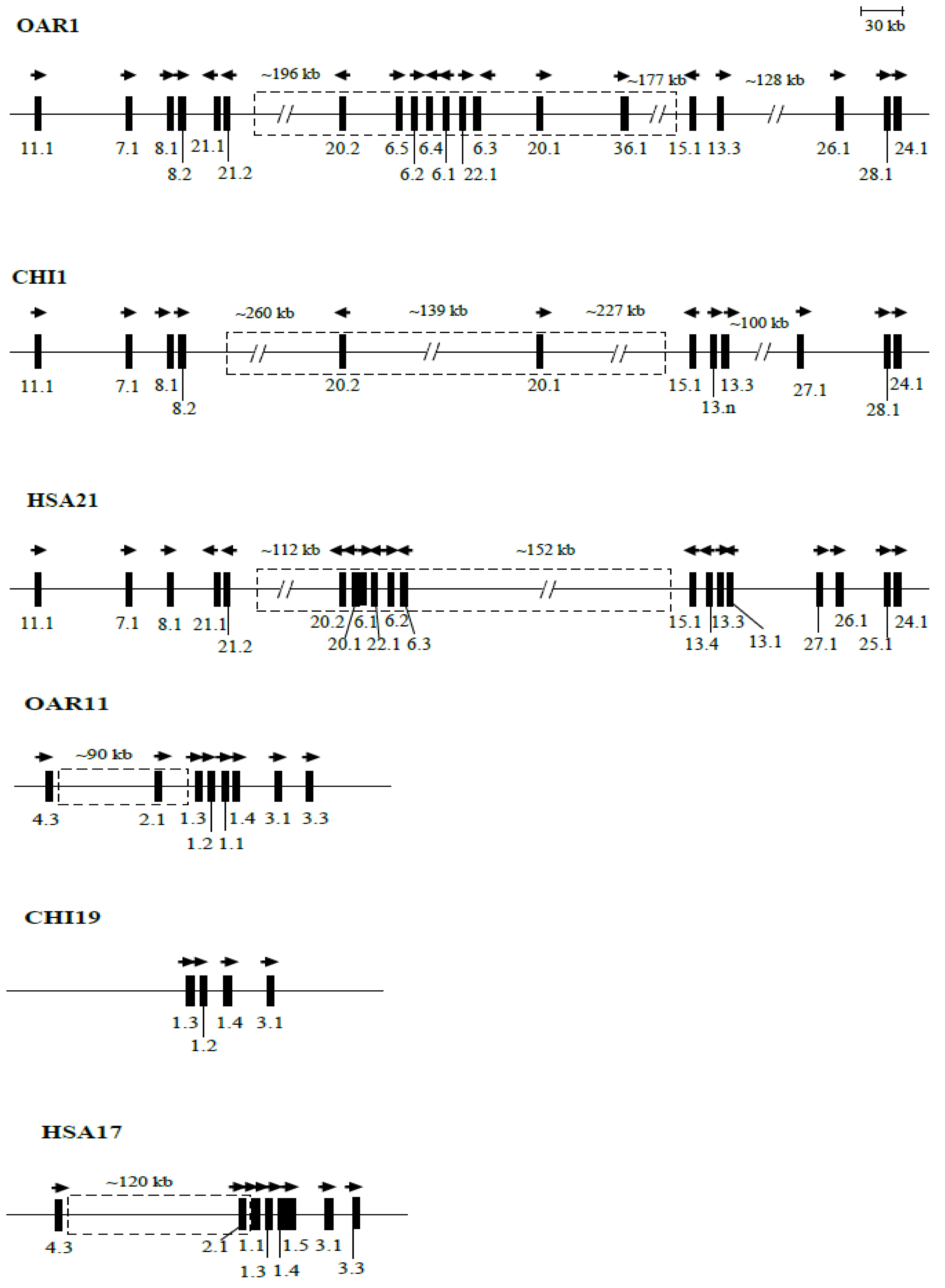
5. The Chromosomal Clustering of KRTAPs and Evolution
6. The Effect of KRTAP Variation
7. Concluding Remarks and Future Research Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Powell, B.C.; Rogers, G.E. The Role of Keratin Proteins and Their Genes in the Growth, Structure and Properties of Hair. In Formation and Structure of Human Hair; Birkhäuser: Basel, Switzerland, 1997; pp. 59–148. [Google Scholar]
- Fujikawa, H.; Fujimoto, A.; Farooq, M.; Ito, M.; Shimomura, Y. Characterisation of the human hair keratin-associated protein 2 (KRTAP2) gene family. J. Invest. Dermatol. 2012, 132, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Plowman, J.E.; Miller, R.E.; Thomas, A.; Grosvenor, A.J.; Harland, D.P.; Deb-Choudhury, S. A detailed mapping of the readily accessible disulphide bonds in the cortex of wool fibres. Proteins 2021, 89, 708–720. [Google Scholar] [CrossRef] [PubMed]
- Haylett, T.; Swart, L. Studies on the high-sulfur proteins of reduced merino wool part III: The amino-acid sequence of protein SCMKB-IIIB2. Text. Res. J. 1968, 39, 917–929. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Winter, H.; Schweizer, J. Human hair keratin-associated proteins (KAPs). Int. Rev. Cytol. 2006, 251, 209–263. [Google Scholar]
- Gong, H.; Zhou, H.; Wang, J.; Li, S.; Luo, Y.; Hickford, J.G. Characterisation of an ovine keratin associated protein (KAP) gene, which would produce a protein rich in glycine and tyrosine, but lacking in cysteine. Genes 2019, 10, 848. [Google Scholar] [CrossRef] [PubMed]
- McGaughey, G.B.; Gagné, M.; Rappé, A.K. π-stacking interactions alive and well in proteins. J. Biol. Chem. 1998, 273, 15458–15463. [Google Scholar] [CrossRef]
- Levitt, M.; Perutz, M.F. Aromatic rings act as hydrogen bond acceptors. J. Mol. Biol. 1988, 201, 751–754. [Google Scholar] [CrossRef]
- Fraser, R.B.; Parry, D.A. Filamentous structure of hard β-keratins in the epidermal appendages of birds and reptiles. Subcell. Biochem. 2017, 82, 231–252. [Google Scholar]
- Rogers, M.A.; Schweizer, J. Human KAP genes, only the half of it? Extensive size polymorphisms in hair keratin-associated protein genes. J. Invest. Dermatol. 2005, 124, 8–9. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Winter, H.; Langbein, L.; Wollschläger, A.; Praetzel-Wunder, S.; Jave-Suarez, L.F.; Schweizer, J. Characterization of human KAP24.1, a cuticular hair keratin-associated protein with unusual amino-acid composition and repeat structure. J. Invest. Dermatol. 2007, 127, 1197–1204. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Praetzel Wunder, S.; Giehl, K. Characterization and expression analysis of the hair keratin associated protein KAP26.1. Br. J. Dermatol. 2008, 159, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Aoki, N.; Ito, K.; Ito, M. Hair follicle has a novel anagen-specific protein, mKAP13. J. Invest. Dermatol. 1998, 111, 804–809. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kuhn, F.; Lassing, C.; Range, A.; Mueller, M.; Hunziker, T.; Ziemiecki, A.; Andres, A.C. Pmg-1 and pmg-2 constitute a novel family of KAP genes differentially expressed during skin and mammary gland development. Mech. Dev. 1999, 86, 193–196. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Winter, H.; Ehmann, C.; Praetzel, S.; Schweizer, J. Characterization of a first domain of human high glycine-tyrosine and high sulfur keratin-associated protein (KAP) genes on chromosome 21q22.1. J. Biol. Chem. 2002, 277, 48993–49002. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Forrest, R.H.; Li, S.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G. Wool keratin-associated protein genes in sheep—a review. Genes 2016, 7, 24. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, X.; Chen, H.; Lan, X.Y.; Guo, Y.K.; Li, J.Y.; Wei, T.B.; Jing, Y.J.; Liu, S.Q.; Zhang, M.H.; et al. The PCR-SSCP and DNA sequencing methods detecting a large deletion mutation at KAP6.2 locus in the cashmere goat. Small Rumin. Res. 2008, 75, 243–246. [Google Scholar] [CrossRef]
- Yin, J.; Hu, T.M.; Li, J.Q.; Zhang, C.l.; Guo, Z.Z.; Zhou, H.M. Construction of a skin cDNA library of cashmere goat and cloning of KAP6-2 full-length cDNA. Zool. Res. 2004, 25, 166–171. [Google Scholar]
- Rogers, G.R.; Hickford, J.G.H.; Bickerstaffe, R. Polymorphism in two genes for B2 high sulfur proteins of wool. Anim. Genet. 1994, 25, 407–415. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Yu, Z.; Dyer, J.; Plowman, J.E.; Hickford, J. Identification of the ovine keratin-associated protein KAP1-2 gene (KRTAP1-2). Exp. Dermatol. 2011, 20, 815–819. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Hodge, S.; Dyer, J.M.; Hickford, J.G. Association of wool traits with variation in the ovine KAP1-2 gene in Merino cross lambs. Small Rumin. Res. 2015, 124, 24–29. [Google Scholar] [CrossRef]
- Itenge-Mweza, T.O.; Forrest, R.H.; McKenzie, G.W.; Hogan, A.; Abbott, J.; Amoafo, O.; Hickford, J.G. Polymorphism of the KAP1.1, KAP1.3 and K33 genes in Merino sheep. Mol. Cell. Probes 2007, 21, 338–342. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Hickford, J.G. Polymorphism of the ovine keratin-associated protein 1-4 gene (KRTAP1-4). Mol. Biol. Rep. 2010, 37, 3377–3380. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, H.; Hickford, J.G.; Luo, Y.; Gong, H.; Hu, J.; Liu, X.; Li, S.; Song, Y.; Ke, N. Identification of the ovine keratin-associated protein 2-1 gene and its sequence variation in four Chinese sheep breeds. Genes 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, M.J.; Powell, B.C.; Ward, K.A.; Sleigh, M.J.; Rogers, G.E. The keratin BIIIB gene family: Isolation of cDNA clones and structure of a gene and a related pseudogene. Genomics 1989, 4, 182–191. [Google Scholar] [CrossRef]
- Yu, Z.; Gordon, S.W.; Nixon, A.J.; Bawden, C.S.; Rogers, M.A.; Wildermoth, J.E.; Maqbool, N.J.; Pearson, A.J. Expression patterns of keratin intermediate filament and keratin associated protein genes in wool follicles. Differentiation 2009, 77, 307–316. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, P.; Powell, B.; Rogers, G. Structure and expression of genes for a class of cysteine-rich proteins of the cuticle layers of differentiating wool and hair follicles. J. Cell Biol. 1990, 111, 2587–2600. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Plowman, J.E.; Dyer, J.M.; Hickford, J.G. Analysis of variation in the ovine ultra-high sulphur keratin-associated protein KAP5-4 gene using PCR-SSCP technique. Electrophoresis 2010, 31, 3545–3547. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, H.; Li, S.; Luo, Y.; Hickford, J.G.H. A 57-bp deletion in the ovine KAP6-1 gene affects wool fibre diameter. J. Anim. Breed Genet. 2015, 132, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Zhou, H.; Gong, H.; Yang, Z.; Ma, Q.; Cheng, L.; Ding, W.; Li, Y.; Hickford, J.G. Variation in the KAP6-1 gene in Chinese Tan sheep and associations with variation in wool traits. Small Rumin. Res. 2017, 154, 129–132. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, H.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G.H. Identification of four new gene members of the KAP6 gene family in sheep. Sci. Rep. 2016, 6, 24074. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Hickford, J.G.H. Diversity of the glycine/tyrosine-rich keratin-associated protein 6 gene (KAP6) family in sheep. Mol. Biol. Rep. 2011, 38, 31–35. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Luo, Y.; Hickford, J.G. Variation in the ovine KAP6-3 gene (KRTAP6-3) is associated with variation in mean fibre diameter-associated wool traits. Genes 2017, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Zhou, H.; Plowman, J.E.; Dyer, J.M.; Hickford, J.G.H. Search for variation in the ovine KAP7-1 and KAP8-1 genes using polymerase chain reaction–single-stranded conformational polymorphism screening. DNA Cell Biol. 2012, 31, 367–370. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G. The sheep KAP8-2 gene, a new KAP8 family member that is absent in humans. SpringerPlus 2014, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ullah, F.; Jamal, S.; Zhou, H.; Hickford, J.G.H. Variation in ovine KRTAP8-2 and its association with wool characteristics in Pakistani sheep. Small Rumin. Res. 2021. (Under review). [Google Scholar]
- Gong, H.; Zhou, H.; Dyer, J.M.; Hickford, J.G. Identification of the ovine KAP11-1 gene (KRTAP11-1) and genetic variation in its coding sequence. Mol. Biol. Rep. 2011, 38, 5429–5433. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Dyer, J.M.; Plowman, J.E.; Hickford, J.G.H. Identification of the keratin-associated protein 13-3 (KAP13-3) gene in sheep. Open J. Genet. 2011, 1, 60–64. [Google Scholar] [CrossRef]
- Li, W.; Gong, H.; Zhou, H.; Wang, J.; Liu, X.; Li, S.; Luo, Y.; Hickford, J.G.H. Variation in the ovine keratin-associated protein 15-1 gene affects wool yield. J. Agric. Sci. 2018, 156, 922–928. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Bai, L.; Li, W.; Li, S.; Wang, J.; Luo, Y.; Hickford, J.G.H. Associations between variation in the ovine high glycine-tyrosine keratin-associated protein gene KRTAP20-1 and wool traits. J. Anim. Sci. 2019, 97, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Gong, H.; Zhou, H.; Tao, J.; Hickford, J.G.H. A nucleotide substitution in the ovine KAP20-2 gene leads to a premature stop codon that affects wool fibre curvature. Anim. Genet. 2018, 49, 357–358. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Liu, X.; Hu, J.; Luo, Y.; Hickford, J.G.H. Identification of the ovine keratin-associated protein 21-1 gene and its association with variation in wool traits. Animals 2019, 9, 450. [Google Scholar] [CrossRef]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Liu, X.; Hu, J.; Luo, Y.; Hickford, J.G.H. The mean staple length of wool fibre is associated with variation in the ovine keratin-associated protein 21-2 gene. Genes 2020, 11, 148. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Liu, X.; Luo, Y.; Hickford, J.G.H. Identification of the ovine keratin-associated protein 22-1 (KAP22-1) gene and its effect on wool traits. Genes 2017, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Gong, H.; Yan, W.; Luo, Y.; Hickford, J.G.H. Identification and sequence analysis of the keratin-associated protein 24-1 (KAP24-1) gene homologue in sheep. Gene 2012, 511, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Hu, J.; Luo, Y.; Hickford, J.G.H. Identification of the ovine keratin-associated protein 26-1 gene and its association with variation in wool traits. Genes 2017, 8, 225. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Wang, J.; Zhou, H.; Gong, H.; Tao, J.; Hickford, J.G.H. Identification of ovine KRTAP28-1 and its association with wool fibre diameter. Animals 2019, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Visser, C.; van Marle-Köster, E. Identification of novel variants for KAP1.1, KAP8.1 and KAP13.3 in South African goats. Small Rumin. Res. 2017, 149, 176–180. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, H.; Luo, Y.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Zhang, K.; Zhen, H.; Hickford, J.G.H. Variation in a newly identified caprine KRTAP gene is associated with raw cashmere fiber weight in Longdong cashmere goats. Genes 2021, 12, 625. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Ganai, T.; Shanaz, S.; Ayaz, A.; Khan, N. Allelic polymorphism of KAP1.3 gene in goats. Indian J. Small Rumin. 2017, 23, 257–260. [Google Scholar] [CrossRef]
- Shah, R.; Ganai, T.; Sheikh, F.; Shanaz, S.; Shabir, M.; Khan, H. Characterization and polymorphism of keratin associated protein 1.4 gene in goats. Gene 2013, 518, 431–442. [Google Scholar] [CrossRef]
- Parris, D.; Swart, L.S. Studies on the high-sulphur proteins of reduced mohair. The isolation and amino acid sequence of protein scmkb-m1.2. Biochem. J. 1975, 145, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, A.; Singh, N.; Ganai, N.A. Comparative sequence analysis of keratin associated protein (KAP7.1) gene in two indigenous Pashmina goat breeds of India. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 3314–3318. [Google Scholar]
- Liu, H.; Yue, C.W.; Zhang, W.; Zhu, X.; Yang, G.; Jia, Z. Association of the KAP8.1 gene polymorphisms with fibre traits in inner mongolian cashmere goats. Asian-Australas. J. Anim. Sci. 2011, 24, 1341–1347. [Google Scholar] [CrossRef]
- Liu, H.; Li, N.; Jia, C.; Zhu, X.; Jia, Z. Effect of the polymorphisms of keratin associated protein 8.2 gene on fibre traits in Inner Mongolia cashmere goats. Asian-Australas. J. Anim. Sci. 2007, 20, 821–826. [Google Scholar] [CrossRef]
- Yu, H.; Wang, X.; Chen, H.; Wang, M.; Zhao, M.; Lan, X.Y.; Lei, C.Z.; Wang, K.Y.; Lai, X.S.; Wang, X.L. The polymorphism of a novel 30 bp-deletion mutation at KAP9.2 locus in the cashmere goat. Small Rumin. Res. 2008, 80, 111–115. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Z.; Xu, H.; Qu, L.; Zhao, H.; Li, T.; Zhang, Z. Variation and expression of KAP9.2 gene affecting cashmere trait in goats. Mol. Biol. Rep. 2012, 39, 10525–10529. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Cao, Q.; Wang, R.; Piao, J.; Zhao, F.; Piao, J. Molecular characterization and expression pattern of a novel Keratin-associated protein 11.1 gene in the Liaoning cashmere goat (Capra hircus). Asian-Australas. J. Anim. Sci. 2017, 30, 328–337. [Google Scholar] [CrossRef]
- Li, M.; Liu, X.; Wang, J.; Li, S.; Luo, Y. Molecular characterization of caprine KRTAP13-3 in Liaoning cashmere goat in China. J. Appl. Anim. Res. 2014, 42, 140–144. [Google Scholar] [CrossRef]
- Fang, Y.; Liu, W.; Zhang, F.; Shao, Y.; Yu, S. The polymorphism of a novel mutation of KAP13. 1 gene and its associations with cashmere traits on Xinjiang local goat breed in China. Asian J. Anim. Vet. Adv. 2010, 5, 34–42. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, H.; Hickford, J.G.; Gong, H.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Hao, Z.; Luo, Y. Variation in the caprine keratin-associated protein 15-1 (KAP15-1) gene affects cashmere fibre diameter. Arch. Anim. Breed. 2019, 62, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hao, Z.; Zhou, H.; Luo, Y.; Hu, J.; Liu, X.; Li, S.; Hickford, J.G.H. A keratin-associated protein (KAP) gene that is associated with variation in cashmere goat fleece weight. Small Rumin. Res. 2018, 167, 104–109. [Google Scholar] [CrossRef]
- Wang, J.; Che, L.; Hickford, J.G.; Zhou, H.; Hao, Z.; Luo, Y.; Hu, J.; Liu, X.; Li, S. Identification of the caprine keratin-associated protein 20-2 (KAP20-2) gene and its effect on cashmere traits. Genes 2017, 8, 328. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, H.; Luo, Y.; Zhao, M.; Gong, H.; Hao, Z.; Hu, J.; Hickford, J.G.H. Variation in the caprine KAP24-1 gene affects cashmere fibre diameter. Animals 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhou, H.; Luo, Y.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Hao, Z.; Jin, X.; Song, Y. Variation in the caprine keratin-associated protein 27-1 gene is associated with cashmere fiber diameter. Genes 2020, 11, 934. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, H.; Hickford, J.G.; Zhao, M.; Gong, H.; Hao, Z.; Shen, J.; Hu, J.; Liu, X.; Li, S. Identification of caprine KRTAP28-1 and its effect on cashmere fiber diameter. Genes 2020, 11, 121. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Gong, H.; Zhou, H.; Wang, J.; Li, S.; Liu, X.; Luo, Y.; Hickford, J.G. Variation in KRTAP6-1 affects wool fibre diameter in New Zealand Romney ewes. Arch. Anim. Breed. 2019, 62, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Kijas, J.W.; Townley, D.; Dalrymple, B.P.; Heaton, M.P.; Maddox, J.F.; McGrath, A.; Wilson, P.; Ingersoll, R.G.; McCulloch, R.; McWilliam, S. A genome wide survey of SNP variation reveals the genetic structure of sheep breeds. PLoS ONE 2009, 4, e4668. [Google Scholar] [CrossRef]
- Duret, L.; Galtier, N. Biased gene conversion and the evolution of mammalian genomic landscapes. Ann. Rev. Genom. Hum. Genet. 2009, 10, 285–311. [Google Scholar] [CrossRef] [PubMed]
- Galtier, N.; Duret, L.; Glémin, S.; Ranwez, V. GC-biased gene conversion promotes the fixation of deleterious amino acid changes in primates. Trends Genet. 2009, 25, 1–5. [Google Scholar] [CrossRef] [PubMed]
- 1000 Genomes Project Consortium; Abecasis, G.R.; Altshuler, D.; Auton, A.; Brooks, L.D.; Durbin, R.M.; Gibbs, R.A.; Hurles, M.E.; McVean, G.A. A map of human genome variation from population-scale sequencing. Nature 2010, 467, 1061–1073. [Google Scholar] [PubMed]
- Levinson, G.; Gutman, G.A. Slipped-strand mispairing: A major mechanism for DNA sequence evolution. Mol. Biol. Evol. 1987, 4, 203–221. [Google Scholar] [PubMed]
- Szostak, J.W.; Wu, R. Unequal crossing over in the ribosomal DNA of Saccharomyces cerevisiae. Nature 1980, 284, 426–430. [Google Scholar] [CrossRef]
- Huang, C.R.L.; Burns, K.H.; Boeke, J.D. Active transposition in genomes. Ann. Rev. Genet. 2012, 46, 651–675. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, S.B.; Goode, D.L.; Kvikstad, E.; Albers, C.A.; Zhang, Z.D.; Mu, X.J.; Ananda, G.; Howie, B.; Karczewski, K.J.; Smith, K.S. The origin, evolution, and functional impact of short insertion–deletion variants identified in 179 human genomes. Genome Res. 2013, 23, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Viguera, E.; Canceill, D.; Ehrlich, S.D. Replication slippage involves DNA polymerase pausing and dissociation. EMBO J. 2001, 20, 2587–2595. [Google Scholar] [CrossRef]
- Lovett, S.T.; Drapkin, P.T.; Sutera, V.; Gluckman-Peskind, T.J. A sister-strand exchange mechanism for recA-independent deletion of repeated DNA sequences in Escherichia coli. Genetics 1993, 135, 631–642. [Google Scholar] [CrossRef]
- Ripley, L.S. Model for the participation of quasi-palindromic DNA sequences in frameshift mutation. Proc. Natl. Acad. Sci. USA 1982, 79, 4128–4132. [Google Scholar] [CrossRef]
- Kiktev, D.A.; Sheng, Z.; Lobachev, K.S.; Petes, T.D. GC content elevates mutation and recombination rates in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2018, 115, E7109–E7118. [Google Scholar] [CrossRef]
- Tian, D.; Wang, Q.; Zhang, P.; Araki, H.; Yang, S.; Kreitman, M.; Nagylaki, T.; Hudson, R.; Bergelson, J.; Chen, J.Q. Single-nucleotide mutation rate increases close to insertions/deletions in eukaryotes. Nature 2008, 455, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Wu, Y.; Yang, H.; Bergelson, J.; Kreitman, M.; Tian, D. Variation in the ratio of nucleotide substitution and indel rates across genomes in mammals and bacteria. Mol. Biol. Evol. 2009, 26, 1523–1531. [Google Scholar] [CrossRef]
- McDonald, M.J.; Wang, W.C.; Huang, H.D.; Leu, J.Y. Clusters of nucleotide substitutions and insertion/deletion mutations are associated with repeat sequences. PLoS Biol. 2011, 9, e1000622. [Google Scholar] [CrossRef]
- Smith, G.R.; Kunes, S.M.; Schultz, D.W.; Taylor, A.; Triman, K.L. Structure of chi hotspots of generalized recombination. Cell 1981, 24, 429–436. [Google Scholar] [CrossRef]
- Cheng, K.C.; Smith, G.R. Recombinational hotspot activity of Chi-like sequences. J. Mol. Biol. 1984, 180, 371–377. [Google Scholar] [CrossRef]
- Rogers, M.A.; Langbein, L.; Winter, H.; Ehmann, C.; Praetzel, S.; Korn, B.; Schweizer, J. Characterization of a cluster of human high/ultrahigh sulfur keratin-associated protein genes embedded in the type I keratin gene domain on chromosome 17q12-21. J. Biol. Chem. 2001, 276, 19440–19451. [Google Scholar] [CrossRef]
- Nützmann, H.W.; Scazzocchio, C.; Osbourn, A. Metabolic gene clusters in eukaryotes. Ann. Rev. Genet. 2018, 52, 159–183. [Google Scholar] [CrossRef] [PubMed]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2018, 16, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Obayashi, I.; Asakawa, S.; Minoshima, S.; Kudoh, J.; Shimizu, N. A cluster of 21 keratin-associated protein genes within introns of another gene on human chromosome 21q22.3. Genomics 2004, 83, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef]
- Marques-Bonet, T.; Girirajan, S.; Eichler, E.E. The origins and impact of primate segmental duplications. Trends Genet. 2009, 25, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.A.; Church, D.M.; Ventura, M.; Rocchi, M.; Eichler, E.E. Analysis of segmental duplications and genome assembly in the mouse. Genome Res. 2004, 14, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, J.D.; Wei, Y.; Khavari, P.A. The functions and unique features of long intergenic non-coding RNA. Nat. Rev. Mol. Cell Biol. 2018, 19, 143–157. [Google Scholar] [CrossRef]
- Kelley, D.; Rinn, J. Transposable elements reveal a stem cell-specific class of long noncoding RNAs. Genome Biol. 2012, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cerbin, S.; Jiang, N. Duplication of host genes by transposable elements. Cur. Opin. Genet. Dev. 2018, 49, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R.; Sauna, Z.E.; Ambudkar, S.V.; Gottesman, M.M.; Kimchi-Sarfaty, C. Silent (synonymous) SNPs: Should we care about them? Methods Mol. Biol. 2009, 578, 23–39. [Google Scholar]
- Li, S.W.; Ouyang, H.S.; Rogers, G.E.; Bawden, C.S. Characterization of the structural and molecular defects in fibres and follicles of the merino felting lustre mutant. Exp. Dermatol. 2009, 18, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.M.; Plowman, J.E.; Harland, D.P.; Thomas, A.; Kilminster, T.; Scanlon, T.; Milton, J.; Greeff, J.; Oldham, C.; Clerens, S. Influence of feed restriction on the wool proteome: A combined iTRAQ and fiber structural study. J. Proteom. 2014, 103, 170–177. [Google Scholar] [CrossRef]
- He, D.; Chen, L.; Luo, F.; Zhou, H.; Wang, J.; Zhang, Q.; Lu, T.; Wu, S.; Hickford, J.G.H.; Tao, J. Differentially phosphorylated proteins in the crimped and straight wool of Chinese Tan sheep. J. Proteom. 2021, 235, 104115. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
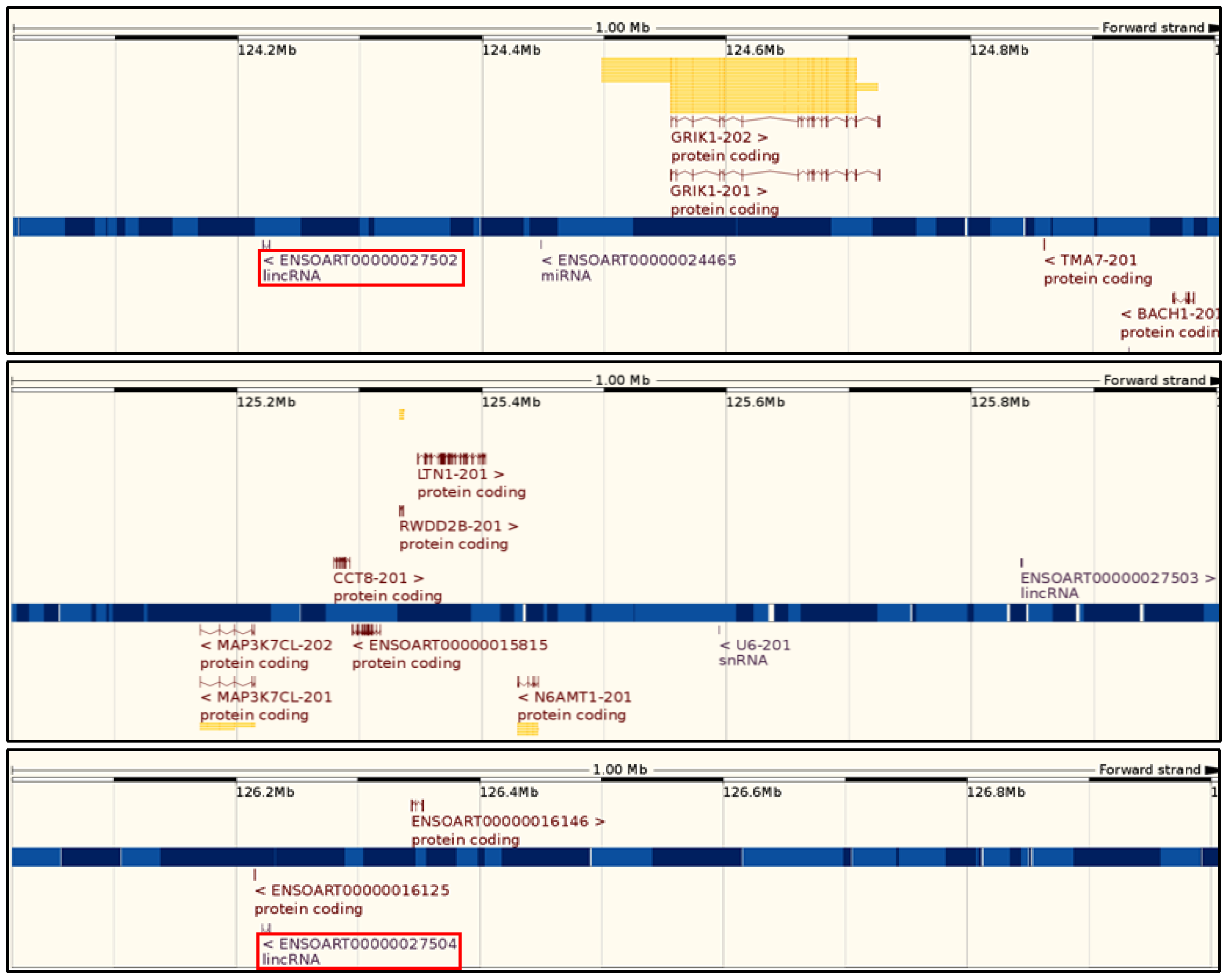{kind=link}
| KRTAP | Number of Variants | Number of SNPs | Length Variation | GenBank Accession Numbers | References |
|---|---|---|---|---|---|
| KRTAP1-1 | 3 | 3 | ± 30-bp repeats | L33885-L33887 | [19] |
| KRTAP1-2 | 11 | 10 | No | HQ897973-HQ897982, KM105941-KM105942 | [20,21] |
| KRTAP1-3 | 9 | 17 | No | AY835589-AY835597 | [22] |
| KRTAP1-4 | 9 | 14 | No | GQ507741-GQ507749 | [23] |
| KRTAP2-1 | 4 | 9 | No | [24] | |
| KRTAP3-1 | Unknown | Unknown | Unknown | M21099 | [25] |
| KRTAP3-3 | Unknown | Unknown | Unknown | N21103 | [25] |
| KRTAP4-3 | Unknown | Unknown | Unknown | EU239778 | [26] |
| KRTAP5-1 | Unknown | Unknown | Unknown | X55294 | [27] |
| KRTAP5-4 | 5 | 6 | ±30-bp repeats | GU255997-GU256001 | [28] |
| KRTAP6-1 | 5 | 4 | ± 57-bp | GU319873, GU319875 | [29,30] |
| KRTAP6-2 | 6 | 5 | No | KT725827-KT725832 | [31] |
| KRTAP6-3 | 7 | 5 | ± 45-bp | KT725833-KT725837, GU319876 | [31,32,33] |
| KRTAP6-4 | 3 | 3 | No | KT725838-KT725840 | [31] |
| KRTAP6-5 | 6 | 5 | ± 18-bp | KT725841-KT725846 | [31] |
| KRTAP7-1 | 2 | 1 | No | JN091630, JN091631 | [34] |
| KRTAP8-1 | 5 | 4 | No | JN091632-JN091636 | [34] |
| KRTAP8-2 | 3 | 2 | No | KF220646-KF220647 | [35,36] |
| KRTAP11-1 | 6 | 5 | No | HQ595347-HQ595352 | [37] |
| KRTAP13-3 | 5 | 4 | No | JN377429-JN377433 | [38] |
| KRTAP15-1 | 4 | 4 | No | MH742372-MH742375 | [39] |
| KRTAP20-1 | 8 | 6 | ±12-bp | MH243552-MH243559 | [40] |
| KRTAP20-2 | 2 | 1 | No | MH071391, MH071392 | [41] |
| KRTAP21-1 | 3 | 2 | No | MF143980-MF143983 | [42] |
| KRTAP21-2 | 5 | 4 | No | MF143975-MF143979 | [43] |
| KRTAP22-1 | 3 | 2 | No | KX377616-KX377618 | [44] |
| KRTAP24-1 | 4 | 7 | No | JX112014-JX112017 | [45] |
| KRTAP26-1 | 4 | 7 | No | KX644903–KX644906 | [46] |
| KRTAP28-1 | 6 | 8 | ±2-bp repeats | MN053915-MN053920 | [47] |
| KRTAP36-1 | 3 | 4 | No | MK770620-MK770622 | [6] |
| KRTAP | Number of Variants | Number of SNPs | Length Variation | GenBank Accession Numbers | References |
|---|---|---|---|---|---|
| KRTAP1-1 * | 7 | 5 | No | [48] | |
| KRTAP1-2 | 6 | 5 | ±60-bp and 15-bp | [49] | |
| KRTAP1-3 * | Unknown | Unknown | Unknown | JQ772533 | [50] |
| KRTAP1-4 | 6 | 8 | No | N012101, JN012102, JN000317, JN000318, JQ436929, JQ627657 | [51] |
| KRTAP3-1 | Unknown | Unknown | Unknown | NM_001285774 | [52] |
| KRTAP7-1 | 2 | 1 | No | AY510121 | [53] |
| KRTAP8-1 | 4 | 2 | No | AY510122, EU595394, EU595395 | [48,54] |
| KRTAP8-2 | 3 | 2 | No | AY510123 | [55] |
| KRTAP9-2 | 3 | 1 | ±30-bp | AY510124, EU430080, | [56,57] |
| KRTAP11-1 | Unknown | Unknown | Unknown | JQ795995 | [58] |
| KRTAP13-3 | 18 | 17 | No | JX426138-JX426145 | [48,59] |
| KRTAP13-n | 2 | 1 | No | AY510115 | [60] |
| KRTAP15-1 | 6 | 8 | No | [61] | |
| KRTAP20-1 | 4 | 6 | No | MG742218- MG742221 | [62] |
| KRTAP20-2 | 3 | 4 | No | MF973462-MF973464 | [63] |
| KRTAP24-1 | 4 | 9 | No | MG996011-MG996014 | [64] |
| KRTAP27-1 | 3 | 2 | No | MN934937- MN934939 | [65] |
| KRTAP28-1 | 5 | 8 | ±2-bp repeats | [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, H.; Gong, H.; Wang, J.; Luo, Y.; Li, S.; Tao, J.; Hickford, J.G.H. The Complexity of the Ovine and Caprine Keratin-Associated Protein Genes. Int. J. Mol. Sci. 2021, 22, 12838. https://doi.org/10.3390/ijms222312838
Zhou H, Gong H, Wang J, Luo Y, Li S, Tao J, Hickford JGH. The Complexity of the Ovine and Caprine Keratin-Associated Protein Genes. International Journal of Molecular Sciences. 2021; 22(23):12838. https://doi.org/10.3390/ijms222312838
Chicago/Turabian StyleZhou, Huitong, Hua Gong, Jiqing Wang, Yuzhu Luo, Shaobin Li, Jinzhong Tao, and Jonathan G. H. Hickford. 2021. "The Complexity of the Ovine and Caprine Keratin-Associated Protein Genes" International Journal of Molecular Sciences 22, no. 23: 12838. https://doi.org/10.3390/ijms222312838
APA StyleZhou, H., Gong, H., Wang, J., Luo, Y., Li, S., Tao, J., & Hickford, J. G. H. (2021). The Complexity of the Ovine and Caprine Keratin-Associated Protein Genes. International Journal of Molecular Sciences, 22(23), 12838. https://doi.org/10.3390/ijms222312838