
Lipid Nanoparticle Technology for Delivering Biologically Active Fatty Acids and Monoglycerides


 ,
, 
Abstract
1. Introduction
2. Why Nano?
2.1. Liposome
2.2. Bicelle
2.3. Solid Lipid Nanoparticle
2.4. Nanostructured Lipid Carrier
2.5. Nanoemulsion
2.6. Lipid Nanocapsule
2.7. Polymeric Micelle
3. Application Examples
3.1. Infectious Diseases
3.1.1. Antibacterial
3.1.2. Antiviral
3.2. Cancer
3.3. Inflammation
3.4. Drug Delivery
4. Conclusions and Outlook
4.1. Engineering Optimization
4.2. Ocular Applications
4.3. Translational Opportunities
4.4. Regulatory Considerations
4.5. Closing Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Greber, U.F.; Tsai, B. Virus and host mechanics support membrane penetration and cell entry. J. Virol. 2016, 90, 3802–3805. [Google Scholar] [CrossRef]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta Biomembr. 2016, 1858, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Cassano, R.; Trombino, S.; Calviello, G. Nanomedicine-based formulations containing ω-3 polyunsaturated fatty acids: Potential application in cardiovascular and neoplastic diseases. Int. J. Nanomed. 2019, 14, 2809–2828. [Google Scholar] [CrossRef] [PubMed]
- Bompard, J.; Rosso, A.; Brizuela, L.; Mebarek, S.; Blum, L.J.; Trunfio-Sfarghiu, A.-M.; Lollo, G.; Granjon, T.; Girard-Egrot, A.; Maniti, O. Membrane fluidity as a new means to selectively target cancer cells with fusogenic lipid carriers. Langmuir 2020, 36, 5134–5144. [Google Scholar] [CrossRef] [PubMed]
- Sunshine, H.; Iruela-Arispe, M.L. Membrane lipids and cell signaling. Curr. Opin. Lipidol. 2017, 28, 408–413. [Google Scholar] [CrossRef]
- Sharma, V.K.; Mamontov, E.; Tyagi, M. Effects of NSAIDs on the nanoscopic dynamics of lipid membrane. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183100. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Jeon, W.-Y.; Sut, T.N.; Cho, N.-J.; Jackman, J.A. Stopping membrane-enveloped viruses with nanotechnology strategies: Toward antiviral drug development and pandemic preparedness. ACS Nano 2021, 15, 125–148. [Google Scholar] [CrossRef]
- Wang, C.-H.; Hsieh, Y.-H.; Powers, Z.M.; Kao, C.-Y. Defeating antibiotic-resistant bacteria: Exploring alternative therapies for a post-antibiotic era. Int. J. Mol. Sci. 2020, 21, 1061. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.A.; Shi, P.-Y.; Cho, N.-J. Targeting the Achilles heel of mosquito-borne viruses for antiviral therapy. ACS Infect. Dis. 2019, 5, 4–8. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Ghimire, J.; Wu, E.; Wimley, W.C. Mechanistic landscape of membrane-permeabilizing peptides. Chem. Rev. 2019, 119, 6040–6085. [Google Scholar] [CrossRef]
- Katdare, A.; Thakkar, S.; Dhepale, S.; Khunt, D.; Misra, M. Fatty acids as essential adjuvants to treat various ailments and their role in drug delivery: A review. Nutrition 2019, 65, 138–157. [Google Scholar] [CrossRef]
- Churchward, C.P.; Alany, R.G.; Snyder, L.A.S. Alternative antimicrobials: The properties of fatty acids and monoglycerides. Crit. Rev. Microbiol. 2018, 44, 561–570. [Google Scholar] [CrossRef]
- Desbois, A.P.; Smith, V.J. Antibacterial free fatty acids: Activities, mechanisms of action and biotechnological potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Valle-González, E.R.; Cho, N.-J. Antibacterial free fatty acids and monoglycerides: Biological activities, experimental testing, and therapeutic applications. Int. J. Mol. Sci. 2018, 19, 1114. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Ahmad, J.; Zafar, S.; Warsi, M.H.; Abdel-Wahab, B.A.; Akhter, S.; Alam, M.A. Omega-3 fatty acids as adjunctive therapeutics: Prospective of nanoparticles in its formulation development. Ther. Deliv. 2020, 11, 851–868. [Google Scholar] [CrossRef]
- Zhang, Q.; Wu, W.; Zhang, J.; Xia, X. Antimicrobial lipids in nano-carriers for antibacterial delivery. J. Drug Target. 2020, 28, 271–281. [Google Scholar] [CrossRef]
- Jackman, J.A.; Yoon, B.K.; Li, D.; Cho, N.-J. Nanotechnology formulations for antibacterial free fatty acids and monoglycerides. Molecules 2016, 21, 305. [Google Scholar] [CrossRef] [PubMed]
- Igyártó, B.Z.; Jacobsen, S.; Ndeupen, S. Future considerations for the mRNA-lipid nanoparticle vaccine platform. Curr. Opin. Virol. 2021, 48, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Thorn, C.R.; Thomas, N.; Boyd, B.J.; Prestidge, C.A. Nano-fats for bugs: The benefits of lipid nanoparticles for antimicrobial therapy. Drug Deliv. Transl. Res. 2021, 11, 1598–1624. [Google Scholar] [CrossRef]
- Gadekar, V.; Borade, Y.; Kannaujia, S.; Rajpoot, K.; Anup, N.; Tambe, V.; Kalia, K.; Tekade, R.K. Nanomedicines accessible in the market for clinical interventions. J. Control. Release 2021, 330, 372–397. [Google Scholar] [CrossRef] [PubMed]
- Shetab Boushehri, M.A.; Dietrich, D.; Lamprecht, A. Nanotechnology as a platform for the development of injectable parenteral formulations: A comprehensive review of the know-hows and state of the art. Pharmaceutics 2020, 12, 510. [Google Scholar] [CrossRef]
- das Neves, J.; Sverdlov Arzi, R.; Sosnik, A. Molecular and cellular cues governing nanomaterial–mucosae interactions: From nanomedicine to nanotoxicology. Chem. Soc. Rev. 2020, 49, 5058–5100. [Google Scholar] [CrossRef]
- Ghasemiyeh, P.; Mohammadi-Samani, S. Potential of nanoparticles as permeation enhancers and targeted delivery options for skin: Advantages and disadvantages. Drug Des. Dev. Ther. 2020, 14, 3271–3289. [Google Scholar] [CrossRef]
- Babadi, D.; Dadashzadeh, S.; Osouli, M.; Daryabari, M.S.; Haeri, A. Nanoformulation strategies for improving intestinal permeability of drugs: A more precise look at permeability assessment methods and pharmacokinetic properties changes. J. Control. Release 2020, 321, 669–709. [Google Scholar] [CrossRef]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid nanoparticles for mRNA delivery. Nat. Rev. Mater. 2021. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Dhawan, V.; Holm, R.; Nagarsenker, M.S.; Perrie, Y. Liposomes: Advancements and innovation in the manufacturing process. Adv. Drug Deliv. Rev. 2020, 154–155, 102–122. [Google Scholar] [CrossRef]
- Taguchi, S.; Kang, B.-S.; Suga, K.; Okamoto, Y.; Jung, H.-S.; Umakoshi, H. A novel method of vesicle preparation by simple dilution of bicelle solution. Biochem. Eng. J. 2020, 162, 107725. [Google Scholar] [CrossRef]
- Sut, T.N.; Park, S.; Yoon, B.K.; Jackman, J.A.; Cho, N.-J. Supported lipid bilayer formation from phospholipid-fatty acid bicellar mixtures. Langmuir 2020, 36, 5021–5029. [Google Scholar] [CrossRef] [PubMed]
- Sut, T.N.; Park, S.; Yoon, B.K.; Jackman, J.A.; Cho, N.-J. Optimal formation of uniform-phase supported lipid bilayers from phospholipid–monoglyceride bicellar mixtures. J. Ind. Eng. Chem. 2020, 88, 285–291. [Google Scholar] [CrossRef]
- Dufourc, E.J. Bicelles and nanodiscs for biophysical chemistry. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183478. [Google Scholar] [CrossRef]
- Kolahdouzan, K.; Jackman, J.A.; Yoon, B.K.; Kim, M.C.; Johal, M.S.; Cho, N.-J. Optimizing the formation of supported lipid bilayers from bicellar mixtures. Langmuir 2017, 33, 5052–5064. [Google Scholar] [CrossRef]
- Gordillo-Galeano, A.; Mora-Huertas, C.E. Solid lipid nanoparticles and nanostructured lipid carriers: A review emphasizing on particle structure and drug release. Eur. J. Pharm. Biopharm. 2018, 133, 285–308. [Google Scholar] [CrossRef]
- Subramaniam, B.; Siddik, Z.H.; Nagoor, N.H. Optimization of nanostructured lipid carriers: Understanding the types, designs, and parameters in the process of formulations. J. Nanoparticle Res. 2020, 22, 1–29. [Google Scholar] [CrossRef]
- Solans, C.; Izquierdo, P.; Nolla, J.; Azemar, N.; Garcia-Celma, M.J. Nano-emulsions. Curr. Opin. Colloid Interface Sci. 2005, 10, 102–110. [Google Scholar] [CrossRef]
- Anton, N.; Benoit, J.-P.; Saulnier, P. Design and production of nanoparticles formulated from nano-emulsion templates—A review. J. Control. Release 2008, 128, 185–199. [Google Scholar] [CrossRef]
- Huynh, N.T.; Passirani, C.; Saulnier, P.; Benoit, J.P. Lipid nanocapsules: A new platform for nanomedicine. Int. J. Pharm. 2009, 379, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Owen, S.C.; Chan, D.P.Y.; Shoichet, M.S. Polymeric micelle stability. Nano Today 2012, 7, 53–65. [Google Scholar] [CrossRef]
- Hwang, D.; Ramsey, J.D.; Kabanov, A.V. Polymeric micelles for the delivery of poorly soluble drugs: From nanoformulation to clinical approval. Adv. Drug Deliv. Rev. 2020, 156, 80–118. [Google Scholar] [CrossRef]
- Kabara, J.J.; Swieczkowski, D.M.; Conley, A.J.; Truant, J.P. Fatty acids and derivatives as antimicrobial agents. Antimicrob. Agents Chemother. 1972, 2, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Schlievert, P.M.; Peterson, M.L. Glycerol monolaurate antibacterial activity in broth and biofilm cultures. PLoS ONE 2012, 7, e40350. [Google Scholar] [CrossRef]
- Yang, D.; Pornpattananangkul, D.; Nakatsuji, T.; Chan, M.; Carson, D.; Huang, C.-M.; Zhang, L. The antimicrobial activity of liposomal lauric acids against Propionibacterium acnes. Biomaterials 2009, 30, 6035–6040. [Google Scholar] [CrossRef]
- Pornpattananangkul, D.; Fu, V.; Thamphiwatana, S.; Zhang, L.; Chen, M.; Vecchio, J.; Gao, W.; Huang, C.-M.; Zhang, L. In vivo treatment of Propionibacterium acnes infection with liposomal lauric acids. Adv. Healthc. Mater. 2013, 2, 1322–1328. [Google Scholar] [CrossRef] [PubMed]
- Madan, S.; Nehate, C.; Barman, T.K.; Rathore, A.S.; Koul, V. Design, preparation, and evaluation of liposomal gel formulations for treatment of acne: In vitro and in vivo studies. Drug Dev. Ind. Pharm. 2019, 45, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-M.; Chen, C.-H.; Pornpattananangkul, D.; Zhang, L.; Chan, M.; Hsieh, M.-F.; Zhang, L. Eradication of drug resistant Staphylococcus aureus by liposomal oleic acids. Biomaterials 2011, 32, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Obonyo, M.; Zhang, L.; Thamphiwatana, S.; Pornpattananangkul, D.; Fu, V.; Zhang, L. Antibacterial activities of liposomal linolenic acids against antibiotic-resistant Helicobacter pylori. Mol. Pharm. 2012, 9, 2677–2685. [Google Scholar] [CrossRef]
- Jung, S.W.; Thamphiwatana, S.; Zhang, L.; Obonyo, M. Mechanism of antibacterial activity of liposomal linolenic acid against Helicobacter pylori. PLoS ONE 2015, 10, e0116519. [Google Scholar] [CrossRef] [PubMed]
- Thamphiwatana, S.; Gao, W.; Obonyo, M.; Zhang, L. In vivo treatment of Helicobacter pylori infection with liposomal linolenic acid reduces colonization and ameliorates inflammation. Proc. Natl. Acad. Sci. USA 2014, 111, 17600–17605. [Google Scholar] [CrossRef]
- Li, X.-X.; Shi, S.; Rong, L.; Feng, M.-Q.; Zhong, L. The impact of liposomal linolenic acid on gastrointestinal microbiota in mice. Int. J. Nanomed. 2018, 13, 1399–1409. [Google Scholar] [CrossRef]
- Sut, T.N.; Valle-González, E.R.; Yoon, B.K.; Park, S.; Jackman, J.A.; Cho, N.-J. Engineered lipid bicelle nanostructures for membrane-disruptive antibacterial applications. Appl. Mater. Today 2021, 22, 100947. [Google Scholar] [CrossRef]
- Tran, T.-Q.-M.; Hsieh, M.-F.; Chang, K.-L.; Pho, Q.-H.; Nguyen, V.-C.; Cheng, C.-Y.; Huang, C.-M. Bactericidal effect of lauric acid-loaded PCL-PEG-PCL nano-sized micelles on skin commensal Propionibacterium acnes. Polymers 2016, 8, 321. [Google Scholar] [CrossRef]
- Taylor, E.N.; Kummer, K.M.; Dyondi, D.; Webster, T.J.; Banerjee, R. Multi-scale strategy to eradicate Pseudomonas aeruginosa on surfaces using solid lipid nanoparticles loaded with free fatty acids. Nanoscale 2014, 6, 825–832. [Google Scholar] [CrossRef]
- Umerska, A.; Cassisa, V.; Matougui, N.; Joly-Guillou, M.-L.; Eveillard, M.; Saulnier, P. Antibacterial action of lipid nanocapsules containing fatty acids or monoglycerides as co-surfactants. Eur. J. Pharm. Biopharm. 2016, 108, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Umerska, A.; Cassisa, V.; Bastiat, G.; Matougui, N.; Nehme, H.; Manero, F.; Eveillard, M.; Saulnier, P. Synergistic interactions between antimicrobial peptides derived from plectasin and lipid nanocapsules containing monolaurin as a cosurfactant against Staphylococcus aureus. Int. J. Nanomed. 2017, 12, 5687–5699. [Google Scholar] [CrossRef] [PubMed]
- Rozenbaum, R.T.; Su, L.; Umerska, A.; Eveillard, M.; Håkansson, J.; Mahlapuu, M.; Huang, F.; Liu, J.; Zhang, Z.; Shi, L.; et al. Antimicrobial synergy of monolaurin lipid nanocapsules with adsorbed antimicrobial peptides against Staphylococcus aureus biofilms in vitro is absent in vivo. J. Control. Release 2019, 293, 73–83. [Google Scholar] [CrossRef]
- Seabra, C.L.; Nunes, C.; Gomez-Lazaro, M.; Correia, M.; Machado, J.C.; Gonçalves, I.C.; Reis, C.A.; Reis, S.; Martins, M.C.L. Docosahexaenoic acid loaded lipid nanoparticles with bactericidal activity against Helicobacter pylori. Int. J. Pharm. 2017, 519, 128–137. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Kao, M.C.; Fang, J.-Y.; Zouboulis, C.C.; Zhang, L.; Gallo, R.L.; Huang, C.-M. Antimicrobial property of lauric acid against Propionibacterium acnes: Its therapeutic potential for inflammatory acne vulgaris. J. Invest Dermatol. 2009, 129, 2480–2488. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-C.; Tsai, T.-H.; Chuang, L.-T.; Li, Y.-Y.; Zouboulis, C.C.; Tsai, P.-J. Anti-bacterial and anti-inflammatory properties of capric acid against Propionibacterium acnes: A comparative study with lauric acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Farkuh, L.; Hennies, P.T.; Nunes, C.; Reis, S.; Barreiros, L.; Segundo, M.A.; Oseliero Filho, P.L.; Oliveira, C.L.P.; Cassago, A.; Portugal, R.V.; et al. Characterization of phospholipid vesicles containing lauric acid: Physicochemical basis for process and product development. Heliyon 2019, 5, e02648. [Google Scholar] [CrossRef]
- Das, U.N. Can bioactive lipids inactivate coronavirus (COVID-19)? Arch. Med. Res. 2020, 51, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, N.F.; Meredith, L.W.; Tidswell, E.L.; Bryden, S.R.; Gonçalves-Carneiro, D.; Chaudhry, Y.; Shannon-Lowe, C.; Folan, M.A.; Lefteri, D.A.; Pingen, M.; et al. A novel antiviral formulation inhibits a range of enveloped viruses. J. Gen. Virol. 2020, 101, 1090–1102. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.A.; Costa, V.V.; Park, S.; Real, A.L.C.V.; Park, J.H.; Cardozo, P.L.; Ferhan, A.R.; Olmo, I.G.; Moreira, T.P.; Bambirra, J.L.; et al. Therapeutic treatment of Zika virus infection using a brain-penetrating antiviral peptide. Nat. Mater. 2018, 17, 971–977. [Google Scholar] [CrossRef] [PubMed]
- Camargos, V.N.; Foureaux, G.; Medeiros, D.C.; da Silveira, V.T.; Queiroz-Junior, C.M.; Matosinhos, A.L.B.; Figueiredo, A.F.A.; Sousa, C.D.F.; Moreira, T.P.; Queiroz, V.F.; et al. In-depth characterization of congenital Zika syndrome in immunocompetent mice: Antibody-dependent enhancement and an antiviral peptide therapy. EBioMedicine 2019, 44, 516–529. [Google Scholar] [CrossRef] [PubMed]
- Hadian, Z.; Sahari, M.A.; Moghimi, H.R.; Barzegar, M. Formulation, characterization and optimization of liposomes containing eicosapentaenoic and docosahexaenoic acids; a methodology approach. Iran J. Pharm. Res. 2014, 13, 393–404. [Google Scholar] [PubMed]
- Chang, H.-I.; Yeh, M.-K. Clinical development of liposome-based drugs: Formulation, characterization, and therapeutic efficacy. Int. J. Nanomed. 2012, 7, 49–60. [Google Scholar]
- Tanaka, Y.; Goto, K.; Matsumoto, Y.; Ueoka, R. Remarkably high inhibitory effects of docosahexaenoic acid incorporated into hybrid liposomes on the growth of tumor cells along with apoptosis. Int. J. Pharm. 2008, 359, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Tanaka, Y.; Matsumoto, Y.; Ueoka, R. Induction of apoptosis of human tumor cells by hybrid liposomes including docosahexaenoic acid. Bioorg. Med. Chem. Lett. 2008, 18, 1880–1883. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Zako, K.; Komizu, Y.; Ueoka, R. Inhibitory effects of hybrid liposomes composed of phosphatidylcholine and docosahexaenoic acid on the growth of colon cancer cells along with apoptosis and differentiation. Chem. Lett. 2010, 40, 90–92. [Google Scholar] [CrossRef]
- Ichihara, H.; Zako, K.; Komizu, Y.; Goto, K.; Ueoka, R. Therapeutic effects of hybrid liposomes composed of phosphatidylcholine and docosahexaenoic acid on the hepatic metastasis of colon carcinoma along with apoptosis in vivo. Biol. Pharm. Bull. 2011, 34, 901–905. [Google Scholar] [CrossRef]
- Skibinski, C.G.; Das, A.; Chen, K.M.; Liao, J.; Manni, A.; Kester, M.; El-Bayoumy, K. A novel biologically active acid stable liposomal formulation of docosahexaenoic acid in human breast cancer cell lines. Chem.-Biol. Interact. 2016, 252, 1–8. [Google Scholar] [CrossRef]
- Jung, S.; Lee, S.; Lee, H.; Yoon, J.; Lee, E.K. Oleic acid-embedded nanoliposome as a selective tumoricidal agent. Colloids Surf. B 2016, 146, 585–589. [Google Scholar] [CrossRef] [PubMed]
- Mussi, S.V.; Sawant, R.; Perche, F.; Oliveira, M.C.; Azevedo, R.B.; Ferreira, L.A.M.; Torchilin, V.P. Novel nanostructured lipid carrier co-loaded with doxorubicin and docosahexaenoic acid demonstrates enhanced in vitro activity and overcomes drug resistance in MCF-7/Adr cells. Pharm. Res. 2014, 31, 1882–1892. [Google Scholar] [CrossRef]
- Fernandes, R.S.; Silva, J.O.; Mussi, S.V.; Lopes, S.C.A.; Leite, E.A.; Cassali, G.D.; Cardoso, V.N.; Townsend, D.M.; Colletti, P.M.; Ferreira, L.A.M.; et al. Nanostructured lipid carrier co-loaded with doxorubicin and docosahexaenoic acid as a theranostic agent: Evaluation of biodistribution and antitumor activity in experimental model. Mol. Imaging Biol. 2018, 20, 437–447. [Google Scholar] [CrossRef]
- Lages, E.B.; Fernandes, R.S.; Silva, J.d.O.; de Souza, Â.M.; Cassali, G.D.; de Barros, A.L.B.; Miranda Ferreira, L.A. Co-delivery of doxorubicin, docosahexaenoic acid, and α-tocopherol succinate by nanostructured lipid carriers has a synergistic effect to enhance antitumor activity and reduce toxicity. Biomed. Pharmacother. 2020, 132, 110876. [Google Scholar] [CrossRef]
- Lanna, E.G.; Siqueira, R.P.; Machado, M.G.C.; de Souza, A.; Trindade, I.C.; Branquinho, R.T.; Mosqueira, V.C.F. Lipid-based nanocarriers co-loaded with artemether and triglycerides of docosahexaenoic acid: Effects on human breast cancer cells. Biomed. Pharmacother. 2021, 134, 111114. [Google Scholar] [CrossRef] [PubMed]
- Mussi, S.V.; Silva, R.C.; Oliveira, M.C.d.; Lucci, C.M.; Azevedo, R.B.d.; Ferreira, L.A.M. New approach to improve encapsulation and antitumor activity of doxorubicin loaded in solid lipid nanoparticles. Eur. J. Pharm. Sci. 2013, 48, 282–290. [Google Scholar] [CrossRef]
- Serini, S.; Cassano, R.; Corsetto, P.A.; Rizzo, A.M.; Calviello, G.; Trombino, S. Omega-3 PUFA loaded in resveratrol-based solid lipid nanoparticles: Physicochemical properties and antineoplastic activities in human colorectal cancer cells in vitro. Int. J. Mol. Sci. 2018, 19, 586. [Google Scholar] [CrossRef]
- Yutaka, K.; Taiki, I.; Koichi, G.; Yoko, M.; Ryuichi, U. Marked inhibitory effects of extracts from barley-koji miso encapsulated in hybrid liposomes on the growth of human stomach tumor cells in vitro. Chem. Lett. 2005, 34, 370–371. [Google Scholar]
- Bégin, M.E.; Das, U.N.; Ells, G.; Horrobin, D.F. Selective killing of human cancer cells by polyunsaturated fatty acids. Prostaglandins Leukot. Med. 1985, 19, 177–186. [Google Scholar] [CrossRef]
- Das, U.N.; Begin, M.E.; Ells, G.; Huang, Y.S.; Horrobin, D.F. Polyunsaturated fatty acids augment free radical generation in tumor cells in vitro. Biochem. Biophys. Res. Commun. 1987, 145, 15–24. [Google Scholar] [CrossRef]
- Sofias, A.M.; Combes, F.; Koschmieder, S.; Storm, G.; Lammers, T. A paradigm shift in cancer nanomedicine: From traditional tumor targeting to leveraging the immune system. Drug Discov. Today 2021, 26, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Sheth, V.; Wang, L.; Bhattacharya, R.; Mukherjee, P.; Wilhelm, S. Strategies for delivering nanoparticles across tumor blood vessels. Adv. Funct. Mater. 2021, 31, 2007363. [Google Scholar] [CrossRef]
- Calder, P.C.; Waitzberg, D.L.; Klek, S.; Martindale, R.G. Lipids in parenteral nutrition: Biological aspects. J. Parenter. Enter. Nutr. 2020, 44, S21–S27. [Google Scholar] [CrossRef] [PubMed]
- Rasti, B.; Jinap, S.; Mozafari, M.R.; Yazid, A.M. Comparative study of the oxidative and physical stability of liposomal and nanoliposomal polyunsaturated fatty acids prepared with conventional and Mozafari methods. Food Chem. 2012, 135, 2761–2770. [Google Scholar] [CrossRef]
- Sahari, M.A.; Moghimi, H.R.; Hadian, Z.; Barzegar, M.; Mohammadi, A. Improved physical stability of docosahexaenoic acid and eicosapentaenoic acid encapsulated using nanoliposome containing α-tocopherol. Int. J. Food Sci. Technol. 2016, 51, 1075–1086. [Google Scholar] [CrossRef]
- Sahari, M.A.; Moghimi, H.R.; Hadian, Z.; Barzegar, M.; Mohammadi, A. Physicochemical properties and antioxidant activity of α-tocopherol loaded nanoliposome’s containing DHA and EPA. Food Chem. 2017, 215, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Alaarg, A.; Jordan, N.Y.; Verhoef, J.J.; Metselaar, J.M.; Storm, G.; Kok, R.J. Docosahexaenoic acid liposomes for targeting chronic inflammatory diseases and cancer: An in vitro assessment. Int. J. Nanomed. 2016, 11, 5027–5040. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Cassano, R.; Facchinetti, E.; Amendola, G.; Trombino, S.; Calviello, G. Anti-irritant and anti-inflammatory effects of DHA encapsulated in resveratrol-based solid lipid nanoparticles in human keratinocytes. Nutrients 2019, 11, 1400. [Google Scholar] [CrossRef] [PubMed]
- de Castro Leão, M.; Raffin Pohlmann, A.; de Cristo Soares Alves, A.; Helena Poliselli Farsky, S.; Klimuk Uchiyama, M.; Araki, K.; Sandri, S.; Stanisçuaski Guterres, S.; Alves Castro, I. Docosahexaenoic acid nanoencapsulated with anti-PECAM-1 as co-therapy for atherosclerosis regression. Eur. J. Pharm. Biopharm. 2021, 159, 99–107. [Google Scholar] [CrossRef]
- Zhang, M.S.; Tran, P.M.; Wolff, A.J.; Tremblay, M.M.; Fosdick, M.G.; Houtman, J.C.D. Glycerol monolaurate induces filopodia formation by disrupting the association between LAT and SLP-76 microclusters. Sci. Signal. 2018, 11, eaam9095. [Google Scholar] [CrossRef]
- Marwah, H.; Garg, T.; Goyal, A.K.; Rath, G. Permeation enhancer strategies in transdermal drug delivery. Drug Deliv. 2016, 23, 564–578. [Google Scholar] [CrossRef]
- Bunchongprasert, K.; Shao, J. Cytotoxicity and permeability enhancement of Capmul®MCM in nanoemulsion formulation. Int. J. Pharm. 2019, 561, 289–295. [Google Scholar] [CrossRef]
- Hossain, Z.; Kurihara, H.; Hosokawa, M.; Takahashi, K. Docosahexaenoic acid and eicosapentaenoic acid-enriched phosphatidylcholine liposomes enhance the permeability, transportation and uptake of phospholipids in Caco-2 cells. Mol. Cell. Biochem. 2006, 285, 155–163. [Google Scholar] [CrossRef]
- Eckert, G.P.; Chang, S.; Eckmann, J.; Copanaki, E.; Hagl, S.; Hener, U.; Müller, W.E.; Kögel, D. Liposome-incorporated DHA increases neuronal survival by enhancing non-amyloidogenic APP processing. Biochim. Biophys. Acta Biomembr. 2011, 1808, 236–243. [Google Scholar] [CrossRef]
- Narvekar, M.; Xue, H.Y.; Tran, N.T.; Mikhael, M.; Wong, H.L. A new nanostructured carrier design including oil to enhance the pharmaceutical properties of retinoid therapy and its therapeutic effects on chemo-resistant ovarian cancer. Eur. J. Pharm. Biopharm. 2014, 88, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Si, M.; Wu, D.; Xue, H.Y.; Hu, W.; Wong, H.L. Incorporation of docosahexaenoic acid (DHA) enhances nanodelivery of antiretroviral across the blood-brain barrier for treatment of HIV reservoir in brain. J. Control. Release 2020, 328, 696–709. [Google Scholar] [CrossRef]
- Jackman, J.A.; Cho, N.-J. Supported lipid bilayer formation: Beyond vesicle fusion. Langmuir 2020, 36, 1387–1400. [Google Scholar] [CrossRef] [PubMed]
- Thid, D.; Benkoski, J.J.; Svedhem, S.; Kasemo, B.; Gold, J. DHA-induced changes of supported lipid membrane morphology. Langmuir 2007, 23, 5878–5881. [Google Scholar] [CrossRef]
- Flynn, K.R.; Martin, L.L.; Ackland, M.L.; Torriero, A.A.J. Real-time quartz crystal microbalance monitoring of free docosahexaenoic acid interactions with supported lipid bilayers. Langmuir 2016, 32, 11717–11727. [Google Scholar] [CrossRef]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Cho, N.-J. Spectrum of membrane morphological responses to antibacterial fatty acids and related surfactants. Langmuir 2015, 31, 10223–10232. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, L.M.; Yoon, B.K.; Jackman, J.A.; Knoll, W.; Weiss, P.S.; Cho, N.-J. Understanding how sterols regulate membrane remodeling in supported lipid bilayers. Langmuir 2017, 33, 14756–14765. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Jackman, J.A.; Kim, M.C.; Sut, T.N.; Cho, N.-J. Correlating membrane morphological responses with micellar aggregation behavior of capric acid and monocaprin. Langmuir 2017, 33, 2750–2759. [Google Scholar] [CrossRef] [PubMed]
- Valle-González, E.R.; Jackman, J.A.; Yoon, B.K.; Park, S.; Sut, T.N.; Cho, N.-J. Characterizing how acidic pH conditions affect the membrane-disruptive activities of lauric acid and glycerol monolaurate. Langmuir 2018, 34, 13745–13753. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Jackman, J.A.; Park, S.; Mokrzecka, N.; Cho, N.-J. Characterizing the membrane-disruptive behavior of dodecylglycerol using supported lipid bilayers. Langmuir 2019, 35, 3568–3575. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Kim, M.C.; Jackman, J.A.; Cho, N.-J. Dynamic remodeling of giant unilamellar vesicles induced by monoglyceride nano-micelles: Insights into supramolecular organization. Appl. Mater. Today 2021, 24, 101099. [Google Scholar] [CrossRef]
- Yoon, B.K.; Park, S.; Ma, G.J.; Kolahdouzan, K.; Zhdanov, V.P.; Jackman, J.A.; Cho, N.-J. Competing interactions of fatty acids and monoglycerides trigger synergistic phospholipid membrane remodeling. J. Phys. Chem. Lett. 2020, 11, 4951–4957. [Google Scholar] [CrossRef]
- Yoon, B.K.; Park, S.; Jackman, J.A.; Cho, N.-J. Supported lipid bilayer platform for characterizing the optimization of mixed monoglyceride nano-micelles. Appl. Mater. Today 2020, 19, 100598. [Google Scholar] [CrossRef]
- Zhdanov, V.P.; Jackman, J.A. Analysis of the initiation of viral infection under flow conditions with applications to transmission in feed. Biosystems 2020, 196, 104184. [Google Scholar] [CrossRef]
- Jackman, J.A.; Hakobyan, A.; Zakaryan, H.; Elrod, C.C. Inhibition of African swine fever virus in liquid and feed by medium-chain fatty acids and glycerol monolaurate. J. Anim. Sci. Biotechnol. 2020, 11, 114. [Google Scholar] [CrossRef]
- Hsia, C.-Y.; Chen, L.; Singh, R.R.; DeLisa, M.P.; Daniel, S. A molecularly complete planar bacterial outer membrane platform. Sci. Rep. 2016, 6, 32715. [Google Scholar] [CrossRef]
- Mohamed, Z.; Shin, J.-H.; Ghosh, S.; Sharma, A.K.; Pinnock, F.; Bint E Naser Farnush, S.; Dörr, T.; Daniel, S. Clinically relevant bacterial outer membrane models for antibiotic screening applications. ACS Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Singh, M.; Bharadwaj, S.; Lee, K.E.; Kang, S.G. Therapeutic nanoemulsions in ophthalmic drug administration: Concept in formulations and characterization techniques for ocular drug delivery. J. Control. Release 2020, 328, 895–916. [Google Scholar] [CrossRef]
- Suay-García, B.; Pérez-Gracia, M.T. Future prospects for Neisseria gonorrhoeae treatment. Antibiotics 2018, 7, 49. [Google Scholar] [CrossRef]
- Churchward, C.P.; Alany, R.G.; Kirk, R.S.; Walker, A.J.; Snyder, L.A.S. Prevention of ophthalmia neonatorum caused by Neisseria gonorrhoeae using a fatty acid-based formulation. mBio 2017, 8, e00534-17. [Google Scholar] [CrossRef] [PubMed]
- Churchward, C.P.; Calder, A.; Snyder, L.A.S. Mutations in Neisseria gonorrhoeae grown in sub-lethal concentrations of monocaprin do not confer resistance. PLoS ONE 2018, 13, e0195453. [Google Scholar] [CrossRef]
- Churchward, C.P.; Al-Kinani, A.A.; Abdelkader, H.; Swinden, J.; Siwoku, O.; Varnakulasingam, T.; Alany, R.G.; Snyder, L.A.S. Monocaprin eye drop formulation to combat antibiotic resistant gonococcal blindness. Sci. Rep. 2020, 10, 12010. [Google Scholar] [CrossRef]
- Butt, U.; ElShaer, A.; Snyder, L.A.S.; Al-Kinani, A.A.; Le Gresley, A.; Alany, R.G. Fatty acid based microemulsions to combat ophthalmia neonatorum caused by Neisseria gonorrhoeae and Staphylococcus aureus. Nanomaterials 2018, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Jurišić Dukovski, B.; Juretić, M.; Bračko, D.; Randjelović, D.; Savić, S.; Crespo Moral, M.; Diebold, Y.; Filipović-Grčić, J.; Pepić, I.; Lovrić, J. Functional ibuprofen-loaded cationic nanoemulsion: Development and optimization for dry eye disease treatment. Int. J. Pharm. 2020, 576, 118979. [Google Scholar] [CrossRef] [PubMed]
- Varela-Fernández, R.; Díaz-Tomé, V.; Luaces-Rodríguez, A.; Conde-Penedo, A.; García-Otero, X.; Luzardo-Álvarez, A.; Fernández-Ferreiro, A.; Otero-Espinar, F.J. Drug delivery to the posterior segment of the eye: Biopharmaceutic and pharmacokinetic considerations. Pharmaceutics 2020, 12, 269. [Google Scholar] [CrossRef]
- Walenga, R.L.; Babiskin, A.H.; Zhang, X.; Absar, M.; Zhao, L.; Lionberger, R.A. Impact of vehicle physicochemical properties on modeling-based predictions of cyclosporine ophthalmic emulsion bioavailability and tear film breakup time. J. Pharm. Sci. 2019, 108, 620–629. [Google Scholar] [CrossRef]
- Gan, L.; Wang, J.; Jiang, M.; Bartlett, H.; Ouyang, D.; Eperjesi, F.; Liu, J.; Gan, Y. Recent advances in topical ophthalmic drug delivery with lipid-based nanocarriers. Drug Discov. Today 2013, 18, 290–297. [Google Scholar] [CrossRef]
- dos Santos Ramos, M.A.; de Toledo, L.G.; Spósito, L.; Marena, G.D.; de Lima, L.C.; Fortunato, G.C.; Araújo, V.H.S.; Bauab, T.M.; Chorilli, M. Nanotechnology-based lipid systems applied to resistant bacterial control: A review of their use in the past two decades. Int. J. Pharm. 2021, 603, 120706. [Google Scholar] [CrossRef] [PubMed]
- Juretić, M.; Jurišić Dukovski, B.; Krtalić, I.; Reichl, S.; Cetina-Čižmek, B.; Filipović-Grčić, J.; Lovrić, J.; Pepić, I. HCE-T cell-based permeability model: A well-maintained or a highly variable barrier phenotype? Eur. J. Pharm. Sci. 2017, 104, 23–30. [Google Scholar] [CrossRef]
- Pepić, I.; Lovrić, J.; Cetina-Čižmek, B.; Reichl, S.; Filipović-Grčić, J. Toward the practical implementation of eye-related bioavailability prediction models. Drug Discov. Today 2014, 19, 31–44. [Google Scholar] [CrossRef]
- Mattern, K.; Beißner, N.; Reichl, S.; Dietzel, A. DynaMiTES—A dynamic cell culture platform for in vitro drug testing PART 1–Engineering of microfluidic system and technical simulations. Eur. J. Pharm. Biopharm. 2018, 126, 159–165. [Google Scholar] [CrossRef]
- Beiβner, N.; Mattern, K.; Dietzel, A.; Reichl, S. DynaMiTES—A dynamic cell culture platform for in vitro drug testing PART 2–Ocular DynaMiTES for drug absorption studies of the anterior eye. Eur. J. Pharm. Biopharm. 2018, 126, 166–176. [Google Scholar] [CrossRef]
- Ribeiro, L.N.d.M.; de Paula, E.; Rossi, D.A.; Martins, F.A.; de Melo, R.T.; Monteiro, G.P.; Breitkreitz, M.C.; Goulart, L.R.; Fonseca, B.B. Nanocarriers from natural lipids with in vitro activity against Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2021, 10, 571040. [Google Scholar] [CrossRef]
- Souto, E.B.; Baldim, I.; Oliveira, W.P.; Rao, R.; Yadav, N.; Gama, F.M.; Mahant, S. SLN and NLC for topical, dermal, and transdermal drug delivery. Expert Opin. Drug Deliv. 2020, 17, 357–377. [Google Scholar] [CrossRef] [PubMed]
- Jackman, J.A.; Mészáros, T.; Fülöp, T.; Urbanics, R.; Szebeni, J.; Cho, N.-J. Comparison of complement activation-related pseudoallergy in miniature and domestic pigs: Foundation of a validatable immune toxicity model. Nanomed. NBM 2016, 12, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Szente, L.; Fenyvesi, É. Cyclodextrin-lipid complexes: Cavity size matters. Struct. Chem. 2017, 28, 479–492. [Google Scholar] [CrossRef]
- Hădărugă, N.G.; Hădărugă, D.I.; Păunescu, V.; Tatu, C.; Ordodi, V.L.; Bandur, G.; Lupea, A.X. Thermal stability of the linoleic acid/α- and β-cyclodextrin complexes. Food Chem. 2006, 99, 500–508. [Google Scholar] [CrossRef]
- Lee, M.C.; Jiang, X.; Brenna, J.T.; Abbaspourrad, A. Oleogel-structured composite for the stabilization of ω3 fatty acids in fish oil. Food Funct. 2018, 9, 5598–5606. [Google Scholar] [CrossRef] [PubMed]
- Vestland, T.L.; Jacobsen, Ø.; Sande, S.A.; Myrset, A.H.; Klaveness, J. Compactible powders of omega-3 and β-cyclodextrin. Food Chem. 2015, 185, 151–158. [Google Scholar] [CrossRef][Green Version]
- Arima, H.; Higashi, T.; Motoyama, K. Improvement of the bitter taste of drugs by complexation with cyclodextrins: Applications, evaluations and mechanisms. Ther. Deliv. 2012, 3, 633–644. [Google Scholar] [CrossRef]
- Brunaldi, K.; Huang, N.; Hamilton, J.A. Fatty acids are rapidly delivered to and extracted from membranes by methyl-β-cyclodextrin. J. Lipid Res. 2010, 51, 120–131. [Google Scholar]
- Vahedi, A.; Bigdelou, P.; Farnoud, A.M. Quantitative analysis of red blood cell membrane phospholipids and modulation of cell-macrophage interactions using cyclodextrins. Sci. Rep. 2020, 10, 15111. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, H.; Benkovics, G.; Fenyvesi, É.; Farewell, A.; Malanga, M.; Ericson, M.B. Delivery of cyclodextrin polymers to bacterial biofilms—An exploratory study using rhodamine labelled cyclodextrins and multiphoton microscopy. Int. J. Pharm. 2017, 531, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Abdelkader, H.; Fathalla, Z.; Moharram, H.; Ali, T.F.S.; Pierscionek, B. Cyclodextrin enhances corneal tolerability and reduces ocular toxicity caused by diclofenac. Oxid. Med. Cell. Longev. 2018, 2018, 5260976. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Q.; Xia, Q. Development of a hydroxypropyl-β-cyclodextrin complex for improving water solubility and reducing skin sensitivity of salicylic acid. Adv. Mat. Res. 2014, 1004–1005, 919–923. [Google Scholar] [CrossRef]
- Negi, J.S.; Chattopadhyay, P.; Sharma, A.K.; Ram, V. Preparation of gamma cyclodextrin stabilized solid lipid nanoparticles (SLNS) using stearic acid–γ-cyclodextrin inclusion complex. J. Incl. Phenom. Macrocycl. Chem. 2014, 80, 359–368. [Google Scholar] [CrossRef]
- Rajnavölgyi, É.; Laczik, R.; Kun, V.; Szente, L.; Fenyvesi, É. Effects of RAMEA-complexed polyunsaturated fatty acids on the response of human dendritic cells to inflammatory signals. Beilstein J. Org. Chem. 2014, 10, 3152–3160. [Google Scholar] [CrossRef]
- Karginov, V.A. Cyclodextrin derivatives as anti-infectives. Curr. Opin. Pharmacol. 2013, 13, 717–725. [Google Scholar] [CrossRef]
- Pratelli, A.; Colao, V. Role of the lipid rafts in the life cycle of canine coronavirus. J. Gen. Virol. 2015, 96, 331–337. [Google Scholar] [CrossRef]
- Sorice, M.; Misasi, R.; Riitano, G.; Manganelli, V.; Martellucci, S.; Longo, A.; Garofalo, T.; Mattei, V. Targeting lipid rafts as a strategy against coronavirus. Front. Cell Dev. Biol. 2021, 8, 618296. [Google Scholar] [CrossRef]
- Leclercq, L.; Nardello-Rataj, V. How to improve the chemical disinfection of contaminated surfaces by viruses, bacteria and fungus? Eur. J. Pharm. Sci. 2020, 155, 105559. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yan, L.; Wang, X.; Zhu, S.; Chen, C.; Gu, Z.; Zhao, Y. Progress, challenges, and future of nanomedicine. Nano Today 2020, 35, 101008. [Google Scholar] [CrossRef]
- Paliwal, R.; Babu, R.J.; Palakurthi, S. Nanomedicine scale-up technologies: Feasibilities and challenges. AAPS PharmSciTech 2014, 15, 1527–1534. [Google Scholar] [CrossRef] [PubMed]
- Roces, C.B.; Port, E.C.; Daskalakis, N.N.; Watts, J.A.; Aylott, J.W.; Halbert, G.W.; Perrie, Y. Rapid scale-up and production of active-loaded PEGylated liposomes. Int. J. Pharm. 2020, 586, 119566. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Markwalter, C.E.; Tian, C.; Armstrong, M.; Prud’homme, R.K. Translational formulation of nanoparticle therapeutics from laboratory discovery to clinical scale. J. Transl. Med. 2019, 17, 200. [Google Scholar] [CrossRef] [PubMed]
- Đorđević, S.; Gonzalez, M.M.; Conejos-Sánchez, I.; Carreira, B.; Pozzi, S.; Acúrcio, R.C.; Satchi-Fainaro, R.; Florindo, H.F.; Vicent, M.J. Current hurdles to the translation of nanomedicines from bench to the clinic. Drug Deliv. Transl. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Hua, S.; de Matos, M.B.C.; Metselaar, J.M.; Storm, G. Current trends and challenges in the clinical translation of nanoparticulate nanomedicines: Pathways for translational development and commercialization. Front. Pharmacol. 2018, 9, 790. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Yano, S.; Kawai, T.; Jinbo, Y.; Nonomura, Y. Selective antibacterial activity of palmitoleic acid in emulsions and other formulations. J. Surfactants Deterg. 2021. [Google Scholar] [CrossRef]
- Mueller, E.A.; Schlievert, P.M. Non-aqueous glycerol monolaurate gel exhibits antibacterial and anti-biofilm activity against Gram-positive and Gram-negative pathogens. PLoS ONE 2015, 10, e0120280. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, A.C.; Widdice, L.E.; Hughes, B.L.; Schlievert, P.; Swamy, G.K.; Stockdale, C.K.; Bernstein, D.I.; Winokur, P.L. Five percent monolaurin vaginal gel for the treatment of bacterial vaginosis: A randomized placebo-controlled trial. J. Low. Genit. Tract Dis. 2020, 24, 277–283. [Google Scholar] [CrossRef] [PubMed]



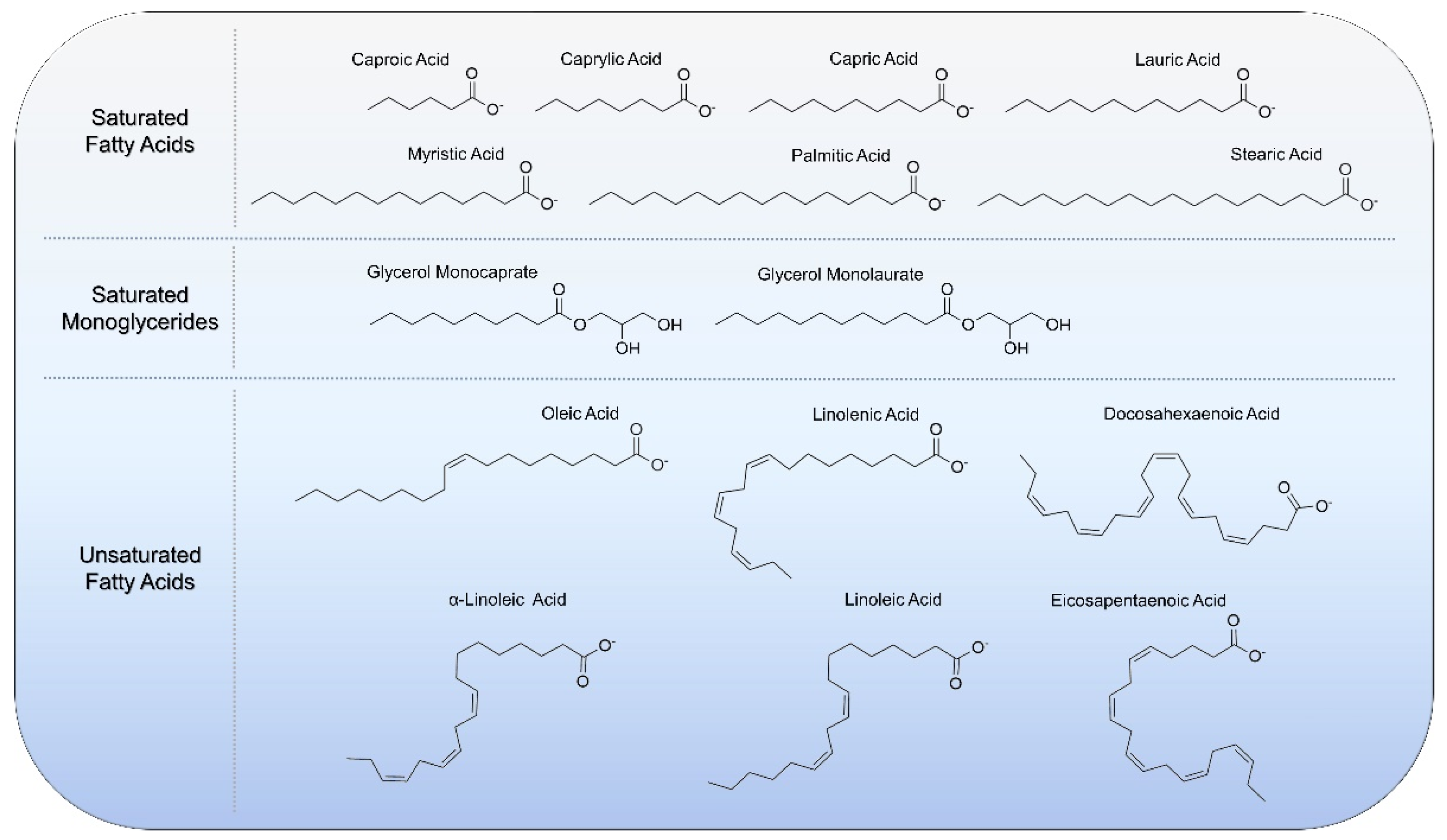{kind=link}
{kind=link}
| Fatty Acid/ Monoglyceride | Bacteria (In Vitro/ In Vivo) | Formulation (Size) | Key Effects | Ref. |
|---|---|---|---|---|
| LA | P. acnes (+) In vitro | Liposomes composed of Egg PC 3, cholesterol, and ~0–69 mol% LA (~120-nm diameter) | - LipoLA was more potent against P. acnes than free LA in vitro, provided there is a critical amount of LA per liposome. - LipoLA fuses with P. acnes bacterial membrane to release LA into bacterial cell membrane. | [42] |
| LA | P. acnes (+) In vivo | Liposomes composed of Egg PC 3, cholesterol, and ~0–69 mol% LA (~120-nm diameter) | - LipoLA showed bactericidal effects against P. acnes in mouse ear model, applied via intradermal injection or topically. - LipoLA did not cause skin irritation commonly caused by acne medications. | [43] |
| LA | P. acnes (+) In vitro, in vivo | Liposomal gel formulation co-loaded with curcumin and LA | - In vitro, curcumin-LA liposomal gel was more potent against P. acnes than curcumin-only gel. - In vivo, curcumin-LA liposomal gel induced greater P. acnes bacterial load reduction than curcumin-only gel in mouse ear model. | [44] |
| OA | S. aureus (+) In vitro, in vivo | Liposomes composed of Egg PC 3, cholesterol, and ~0–61 mol% OA (~80-nm diameter) | - LipoOA was more potent against methicillin-resistant S. aureus than free OA in vitro. - LipoOA had significantly greater bactericidal effects compared to bare liposomes in vivo on mouse skin. | [45] |
| LLA | H. pylori (-) In vitro | Liposomes composed of Egg PC 3, cholesterol, and ~61 mol% LLA (~88-nm diameter) | - LipoLLA killed both spiral and coccoid forms of H. pylori. - LipoLLA treatment had no resistance development while LLA triggered resistant strains to emerge. | [46] |
| LLA, OA, SA | H. pylori (-) In vitro | Liposomes composed of Egg PC 3, cholesterol, and ~50 mol% SA, OA, or LLA (~85-nm diameters) | - LipoLLA was the most potent, followed by LipoOA, while LipoSA showed no antibacterial effect. - LipoLLA caused greater membrane disruption than LipoOA. | [47] |
| LLA | H. pylori (-) In vivo | Liposomes composed of Egg PC 3, cholesterol, and 51 mol% LLA (~106-nm diameter) | - LipoLLA was more potent against H. pylori than free LLA in vitro. - In H. pylori-infected mouse stomach, LipoLLA reduced H. pylori bacterial load significantly compared to conventional antibiotic treatment. | [48] |
| LLA | H. pylori (-) In vivo | Liposomes composed of Egg PC 3, cholesterol, and 51 mol% LLA (~110-nm diameter) | - In H. pylori-infected mouse stomach, LipoLLA caused less gastrointestinal side effects compared to conventional antibiotic treatment. | [49] |
| GML | S. aureus (+) In vitro | Bicelles composed of DOPC 4 loaded with ~29–95 mol% GML (~100–600-nm diameters) | - DOPC/GML bicelles showed membrane-disruptive activity that was distinct from that of free GML, and bicelles with ~80 mol% GML caused pore formation. - Bicelles with ~80 mol% GML were bactericidal against S. aureus at 0.125 mM effective GML concentration. | [50] |
| LA | P. acnes (+) In vitro | Polymeric micelles composed of PCL-PEG-PCL 5 loaded with ~89–96 mol% LA (~30–90 nm diameters) | - LA-loaded micelles were more potent against P. acnes than free LA in vitro. - Efficacy of LA-loaded micelles increased with the amount of loaded LA. | [51] |
| SA, LA, OA | P. aeruginosa (-) In vitro | SLNs loaded with 32 mol% SA + 68 mol% LA; 45 mol% SA + 49 mol% LA + 6% OA; or 35 mol% SA + 59 mol% LA + 6% OA (~150–190 nm diameters) | - All SLN formulations were bactericidal, while the addition of OA enhanced SLN antimicrobial activity. - The SLN formulations could reduce bacterial adhesion on surfaces by ~99%. | [52] |
| COA, CYA, CA, LA, MA, PA, SA, GMC, GML | S. aureus (+), P. aeruginosa (-), A. baumannii (-) In vitro | LNCs 6 loaded with ~20 mol% of various fatty acids (~40–90 nm diameters) | - All fatty acid-LNCs (FA-LNCs) and monoglyceride-LNCs (MG-LNCs), but not LNCs without FA or MG, inhibited Gram-positive S. aureus, while Gram-negative bacteria were more resistant. - MG-LNCs were more potent than FA-LNCs against bacteria. - Longer chain length in antimicrobial lipids increased activity against Gram-positive bacteria and decreased activity against Gram-negative bacteria. | [53] |
| GML | S. aureus (+) In vitro | LNCs 6 6 co-loaded with ~21 mol% GML and antimicrobial peptides (AMPs) (~40-nm diameter) | - Methicillin-susceptible S. aureus was more sensitive to GML-LNCs than methicillin-resistant S. aureus. - GML-LNCs exerted time-dependent bactericidal effects on S. aureus strains. - AMPs and GML-LNCs acted in synergy against S. aureus, i.e., their combination was more potent than GML-LNCs or AMPs alone. | [54] |
| GML | S. aureus (+) In vivo | LNCs 6 co-loaded with ~21 mol% GML and AMPs (~30–130 nm diameter) | - GML-LNCs showed bactericidal activity against four different S. aureus strains. - GML-LNCs significantly reduced bioluminescent S. aureus bacterial count in an infected mouse wound-healing model. | [55] |
| DHA | H. pylori (-) In vitro | NLCs 7 loaded with ~15–30 mol% DHA (~300-nm diameter) | - DHA-loaded NLCs were more potent against H. pylori than free DHA or unloaded NLCs. - DHA-loaded NLCs act on H. pylori by causing membrane fragmentation and leakage of cytoplasmic contents. | [56] |
| Fatty Acid/ Monoglyceride | Cancer Cell Line (In Vitro/In Vivo) | Formulation (Size) | Key Effects | Ref. |
|---|---|---|---|---|
| OA, LNA, ALA, EPA, DHA, and ethyl esters | Lung carcinoma, colon tumor, stomach tumor cells In vitro | HLs composed of DMPC 2, Tween 80, and ~0–100 μg/mL polyunsaturated fatty acid (PUFA) (~50–200 nm diameters) | - Over 1 month, HL-PUFAs remained stable in size, while blank HLs had aggregation-related size increases. - HL-PUFA inhibited cancer cells more potently than those of control liposomes. | [66] |
| LNA, DHA | Lung carcinoma, colon tumor, stomach tumor cells In vitro | HLs composed of DMPC 2, Tween 80, and ~0–200 μg/mL LNA or DHA (<210-nm diameters) | - HL-LNA inhibited tumor cell growth through necrosis-inducing mechanisms that involved lipid peroxidation. - HL-DHA inhibited tumor cell growth through apoptosis-inducing mechanisms that did not depend on lipid peroxidation. | [67] |
| DHA | Stomach tumor cells In vitro | Liposomes composed of DMPC 2 and 10–60 mol% DHA (~100 nm diameter) | - DMPC/DHA liposomes inhibited cancer cell growth more potently than DMPC liposomes. | [68] |
| DHA | Colon carcinoma cells In vivo | Liposomes composed of DMPC 2 and 50 mol% DHA (~100-nm diameter) | - DMPC/DHA liposomes reduced number of metastatic nodules and induced greater apoptosis in metastatic tumor mouse model than DMPC liposomes. - Longer survival time was observed in mouse model after treatment with DMPC/DHA liposomes than with DMPC liposomes. | [69] |
| DHA | Breast cancer cells In vitro | Liposomes composed of ether and phytanyl PC lipids 3 and 10 mol% DHA (~140-nm diameter) | - Liposomal DHA was stable in the pH range of 1 to 7.4 and less prone to oxidation than free DHA over 1–2 weeks. - Liposomal DHA was more effective than free DHA in inhibiting cancer cell growth and malignant transformation. | [70] |
| OA | Breast and lung cancer cells In vitro | Liposomes composed of DPPC 4 phospholipids, cholesterol, and 40 mol% DHA (~130-nm diameter) | - OA-loaded liposomes were cytotoxic to tumor cells while bare liposomes did not inhibit the tumor cells. | [71] |
| DHA | Breast cancer cells In vitro | NLC 5 loaded with doxorubicin (DOX) and 20 mol% or 40 mol% DHA (~80-nm diameter) | - NLC-DHA/DOX was more cytotoxic than free DOX/DHA for both cancer cells. - NLC-DHA/DOX had higher drug uptake in both cancer cells and penetration in tumor spheroid models than free DHA/DOX. | [72] |
| DHA | Breast cancer cells In vivo | NLC 5 loaded with DOX and ~20 mol% DHA (~70-nm diameter) | - NLC-DHA/DOX treatment significantly inhibited tumor growth compared to blank-NLC or NLC-DOX treatment. | [73] |
| DHA | Breast cancer cells In vitro, in vivo | NLC 5 loaded with DOX, α-tocopherol succinate (TS), and ~20 mol% DHA (~80-nm diameter) | - NLC-DHA/DOX/TS had higher antitumor activity in vitro than NLC-DOX and NLC-TS. - In vivo, NLC-DHA/DOX/TS showed the highest tumor growth inhibition, compared to NLC-DOX and blank NLC. | [74] |
| DHA | Breast cancer cells In vitro | PEG-PLA NCs; NEs; and NLCs 6, all co-loaded artemether (ART) and DHA triglycerides (~150–200 nm diameters) | - All 3 lipid nanocarriers improved ART/DHA selectivity and cytotoxicity towards tumor cells, compared to free-ART/DHA or blank-nanocarriers. - NEs had the highest cellular uptake compared to NCs and NLCs. | [75] |
| DHA | Lung cancer cells In vitro | SLNs 7 loaded with DOX and ~0–4 mol% DHA (~100-nm diameter) | - SLN-DHA/DOX was 3 times more effective against cancer cells than free DHA/DOX. - SLN-DHA/DOX had much higher cellular uptake and DOX accumulation than free DHA/DOX or blank SLN. | [76] |
| DHA, LNA | Colon carcinoma cells In vitro | RV-based SLN 8 loaded with ~0.1 mol% DHA or LNA (~840-nm and ~1000-nm diameters for SLN-DHA and SLN-LNA, respectively) | - SLN encapsulation significantly increased LNA and DHA in cancer cells, compared to free LNA and free DHA. - SLN-DHA and SLN-LNA greatly inhibited cancer cell growth and proliferation compared to free DHA and free LNA. | [77] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, J.Y.B.; Yoon, B.K.; Cho, N.-J.; Lovrić, J.; Jug, M.; Jackman, J.A. Lipid Nanoparticle Technology for Delivering Biologically Active Fatty Acids and Monoglycerides. Int. J. Mol. Sci. 2021, 22, 9664. https://doi.org/10.3390/ijms22189664
Tan JYB, Yoon BK, Cho N-J, Lovrić J, Jug M, Jackman JA. Lipid Nanoparticle Technology for Delivering Biologically Active Fatty Acids and Monoglycerides. International Journal of Molecular Sciences. 2021; 22(18):9664. https://doi.org/10.3390/ijms22189664
Chicago/Turabian StyleTan, Jia Ying Brenda, Bo Kyeong Yoon, Nam-Joon Cho, Jasmina Lovrić, Mario Jug, and Joshua A. Jackman. 2021. "Lipid Nanoparticle Technology for Delivering Biologically Active Fatty Acids and Monoglycerides" International Journal of Molecular Sciences 22, no. 18: 9664. https://doi.org/10.3390/ijms22189664
APA StyleTan, J. Y. B., Yoon, B. K., Cho, N.-J., Lovrić, J., Jug, M., & Jackman, J. A. (2021). Lipid Nanoparticle Technology for Delivering Biologically Active Fatty Acids and Monoglycerides. International Journal of Molecular Sciences, 22(18), 9664. https://doi.org/10.3390/ijms22189664