
Integrated Analysis of miR-430 on Steroidogenesis-Related Gene Expression of Larval Rice Field Eel Monopterus albus

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
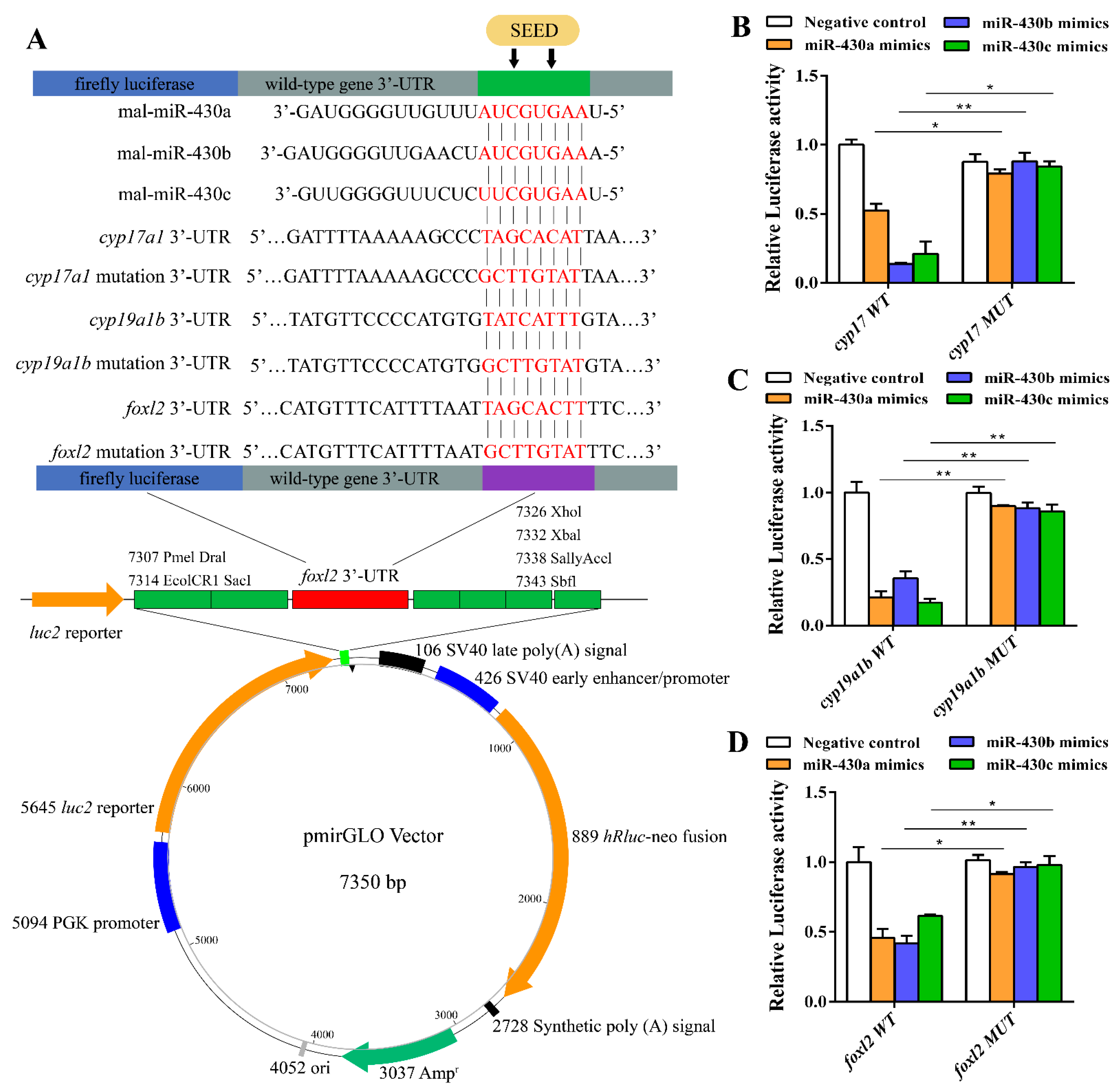{kind=link}
Abstract
1. Introduction
2. Results
2.1. Embryonic and Postembryonic Development of Monopterus albus
2.2. Transcriptome Sequencing Analysis
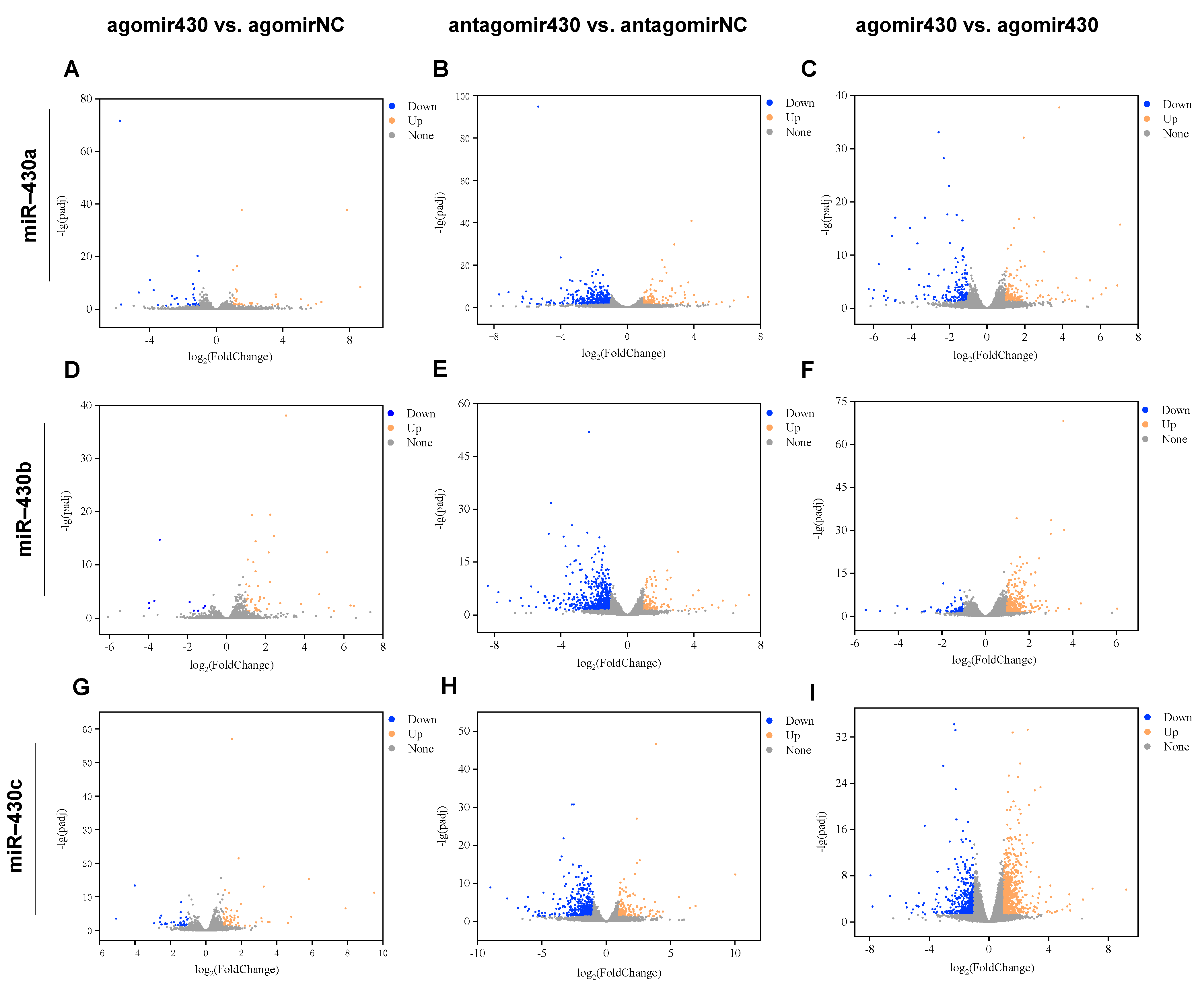
2.2.1. miR430 Expression Level after miR-430 Microinjection of M. albus
2.2.2. Identification and Enrichment Analysis of DEGs
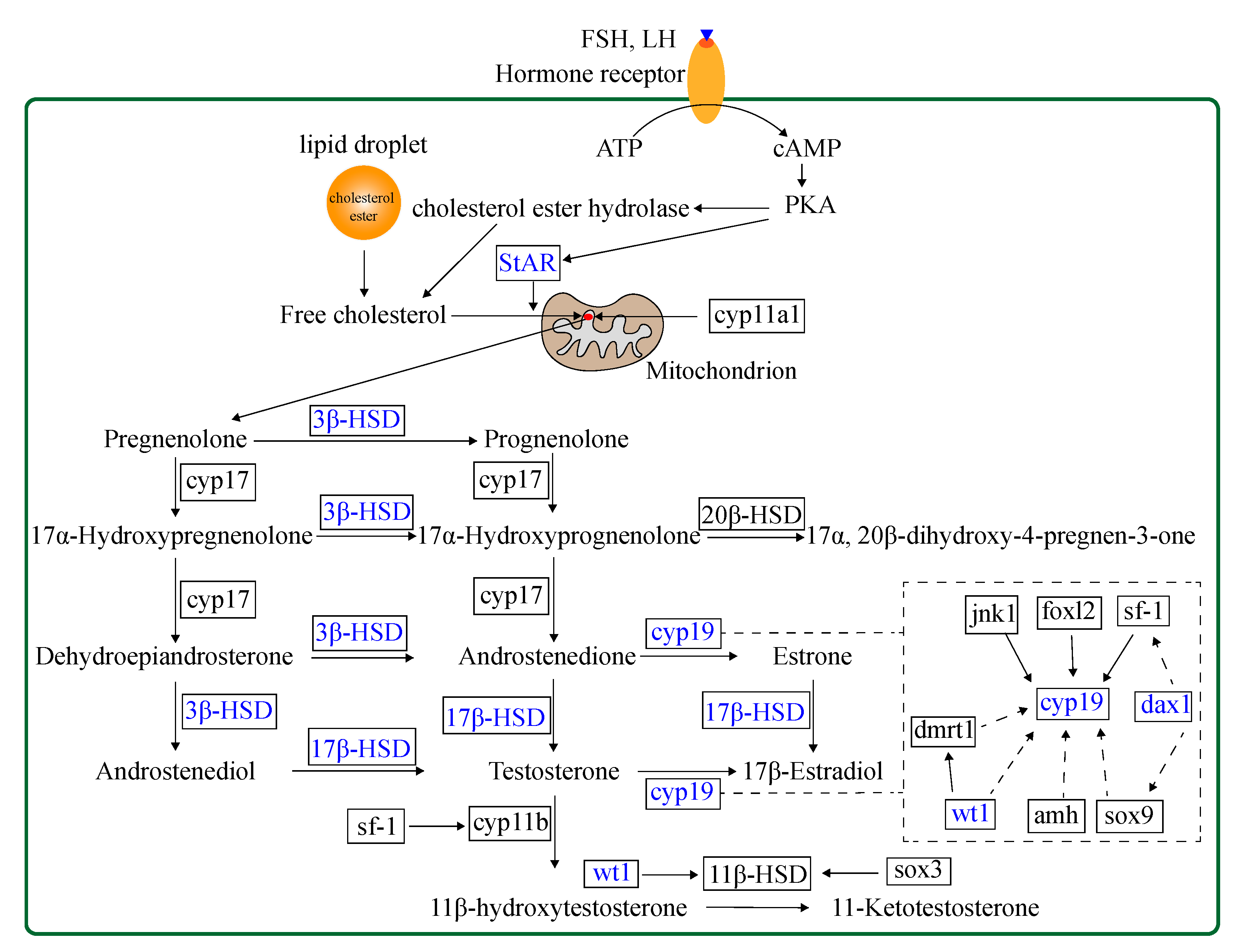
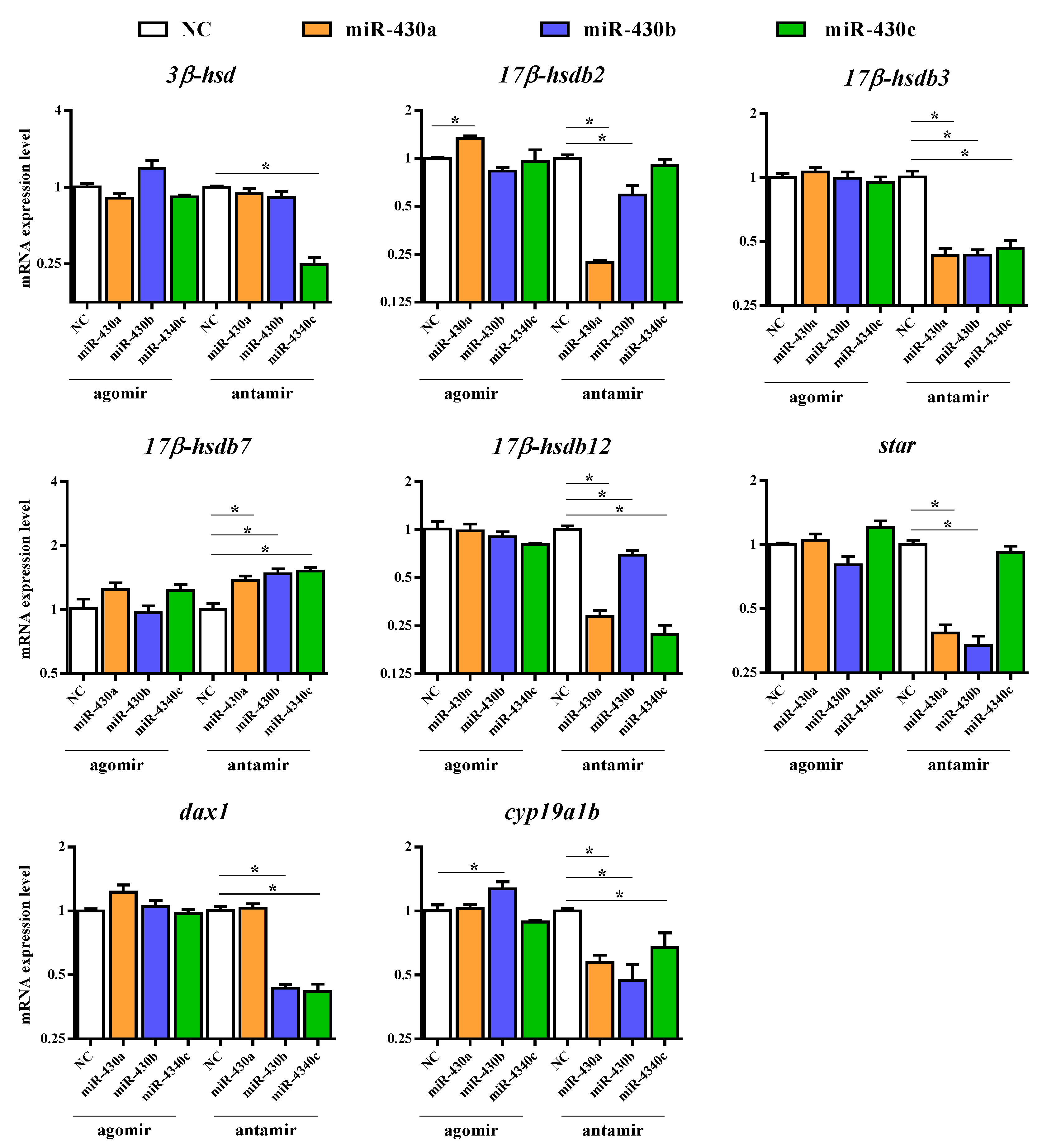
2.2.3. Expression of Steroidogenesis-Related Genes in M. albus
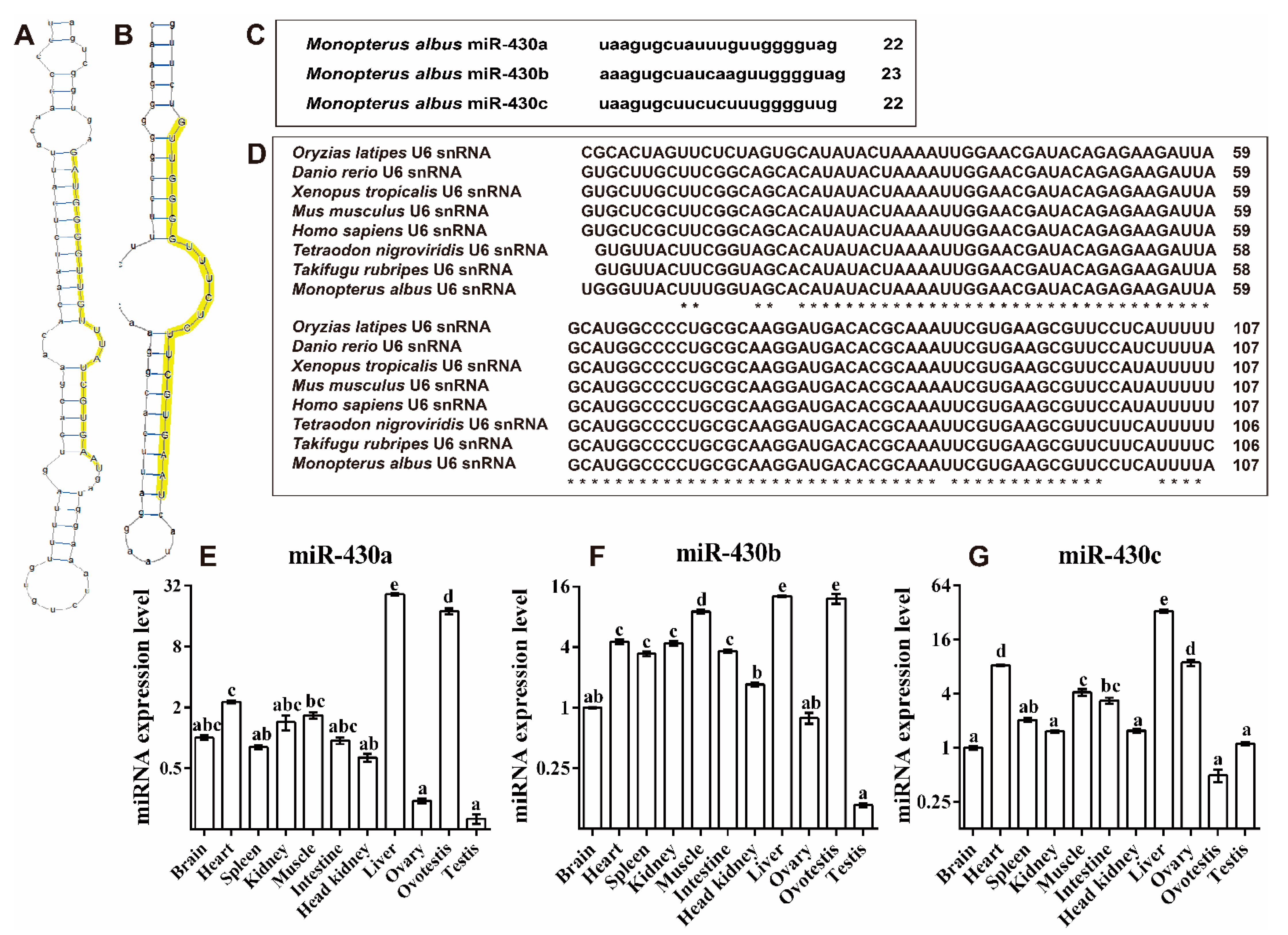
2.3. Molecular Function Identification of miR-430 Family Genes in M. ablus
2.4. Expressions of miR-430 Target Genes Cytochrome P450 Family 17(cyp17), Cytochrome P450 Family 19 Subfamily a (cyp19a1b), and Forkhead Box L2 (foxl2)
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Chemical Reagents
4.3. Microinjection of miRNA Mimics and Inhibitors
4.4. Determination of miRNA Expression by qRT-PCR
4.5. RNA-Seq
4.6. RNA-Seq Result Validation through Quantitative Real-Time PCR
4.7. miR-430 Molecular Structure Analysis and Its Target Gene Verification
4.7.1. Prediction of Secondary Structure of microRNA Precursors in M. albus
4.7.2. Expression Levels of miR-430 in Various Tissues during Different Developmental Stages
4.7.3. Verification of Target Genes of miR-430s
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Omura, T.; Morohashi, K.-I. Gene regulation of steroidogenesis. J. Steroid Biochem. Mol. Biol. 1995, 53, 19–25. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kajiura-Kobayashi, H.; Nagahama, Y. Induction of XY sex reversal by estrogen involves altered gene expression in a teleost, tilapia. Cytogenet. Genome Res. 2003, 101, 289–294. [Google Scholar] [CrossRef]
- Wilhelm, D.; Palmer, S.; Koopman, P. Sex Determination and Gonadal Development in Mammals. Physiol. Rev. 2007, 87, 1–28. [Google Scholar] [CrossRef]
- Asiabi, P.; Leonel, E.; Marbaix, E.; Dolmans, M.M.; A Amorim, C. Immunodetection and quantification of enzymatic markers in theca cells: The early process of ovarian steroidogenesis. Biol. Reprod. 2019, 102, 145–155. [Google Scholar] [CrossRef]
- Trant, J.M.; Thomas, P. Isolation of a novel maturation-inducing steroid produced in vitro by ovaries of Atlantic croaker. Gen. Comp. Endocrinol. 1989, 75, 397–404. [Google Scholar] [CrossRef]
- Sreenivasulu, G.; Senthilkumaran, B.; Sridevi, P.; Rajakumar, A.; Rasheeda, M. Expression and immunolocalization of 20β-hydroxysteroid dehydrogenase during testicular cycle and after hCG induction, in vivo in the catfish, Clarias gariepinus. Gen. Comp. Endocrinol. 2012, 175, 48–54. [Google Scholar] [CrossRef]
- Zhang, X.; Yu, R.M.K.; Jones, P.D.; Lam, G.K.W.; Newsted, J.L.; Gracia, T.; Hecker, M.; Hilscherová, K.; Sanderson, J.T.; Wu, R.S.S.; et al. Quantitative RT-PCR methods for evaluating toxicant-induced effects on steroidogenesis using the H295R cell line. Environ. Sci. Technol. 2005, 39, 2777–2785. [Google Scholar] [CrossRef]
- Gracia, T.; Hilscherova, K.; Jones, P.; Newsted, J.L.; Zhang, X.; Hecker, M.; Higley, E.B.; Sanderson, J.; Yu, R.M.; Wu, R.S.; et al. The H295R system for evaluation of endocrine-disrupting effects. Ecotoxicol. Environ. Saf. 2006, 65, 293–305. [Google Scholar] [CrossRef]
- Hu, Q.; Guo, W.; Gao, Y.; Tang, R.; Li, D. Molecular cloning and analysis of gonadal expression of Foxl2 in the rice-field eel Monopterus albus. Sci. Rep. 2015, 4, 6884. [Google Scholar] [CrossRef]
- Hu, Q.; Guo, W.; Gao, Y.; Tang, R.; Li, D. Molecular cloning and characterization of amh and dax1 genes and their expression during sex inversion in rice-field eel Monopterus albus. Sci. Rep. 2015, 5, 16667. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, D.; Hu, Q.; Li, D. Foxl3, a Target of miR-9, Stimulates Spermatogenesis in Spermatogonia During Natural Sex Change in Monopterus albus. Endocrinology 2016, 157, 4388–4399. [Google Scholar] [CrossRef] [PubMed]
- Guiguen, Y.; Fostier, A.; Piferrer, F.; Chang, C.-F. Ovarian aromatase and estrogens: A pivotal role for gonadal sex differentiation and sex change in fish. Gen. Comp. Endocrinol. 2010, 165, 352–366. [Google Scholar] [CrossRef]
- Gardner, L.; A Anderson, T.; Place, A.; Dixon, B.; Elizur, A. Sex change strategy and the aromatase genes. J. Steroid Biochem. Mol. Biol. 2005, 94, 395–404. [Google Scholar] [CrossRef]
- Park, M.; Shin, E.; Won, M. FOXL2 interacts with steroidogenicfactor-1 (SF-1) and represses SF-1-induced CYP17 transcription in granulosa cells. Mol. Endocrinol. 2010, 24, 1024–1036. [Google Scholar] [CrossRef]
- Thomas, M.; Lieberman, J.; Lal, A. Desperately seeking microRNA targets. Nat. Struct. Mol. Biol. 2010, 17, 1169–1174. [Google Scholar] [CrossRef]
- Xu, S.; Linher-Melville, K.; Yang, B.B.; Wu, D.; Li, J. Micro-RNA378 (miR-378) regulates ovarian estradiol production by targeting aromatase. Endocrinology 2011, 152, 3941–3951. [Google Scholar] [CrossRef]
- Klinge, C.M. miRNAs and estrogen action. Trends Endocrinol. Metab. 2012, 23, 223–233. [Google Scholar] [CrossRef]
- Nusrin, S.; Tong, S.K.; Chaturvedi, G.; Wu, R.S.; Giesy, J.P.; Kong, R.Y. Regulation of CYP11B1 and CYP11B2 steroidogenic genes by hypoxia-inducible miR-10b in H295R cells. Mar. Pollut. Bull. 2014, 85, 344–351. [Google Scholar] [CrossRef]
- Sirotkin, A.V.; Ovcharenko, D.; Grossmann, R.; Lauková, M.; Mlynček, M. Identification of MicroRNAs controlling human ovarian cell steroidogenesis via a genome-scale screen. J. Cell. Physiol. 2009, 219, 415–420. [Google Scholar] [CrossRef]
- Dai, A.; Sun, H.; Fang, T.; Zhang, Q.; Wu, S.; Jiang, Y.; Ding, L.; Yan, G.; Hu, Y. MicroRNA-133b stimulates ovarian estradiol synthesis by targeting Foxl2. FEBS Lett. 2013, 587, 2474–2482. [Google Scholar] [CrossRef]
- Yu, R.M.; Chaturvedi, G.; Tong, S.K.; Nusrin, S.; Giesy, J.P.; Wu, R.S.; Kong, R.Y. Evidence for microRNA-mediated regulation of steroidogenesis by hypoxia. Environ. Sci. Technol. 2015, 49, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, W.; Liu, Q.; Li, B.; An, L.; Hao, R.; Zhao, J.; Liu, S.; Song, J. Coordinated microRNA and messenger RNA ex-pression profiles for understanding sexual dimorphism of gonads and the potential roles of microRNA in the steroidogenesis pathway in Nile tilapia (Oreochromis niloticus). Theriogenology 2016, 85, 970–978. [Google Scholar] [CrossRef]
- Mishima, Y.; Giraldez, A.; Takeda, Y.; Fujiwara, T.; Sakamoto, H.; Schier, A.F.; Inoue, K. Differential regulation of germline mRNAs in soma and germ Cells by zebrafish miR-430. Curr. Biol. 2006, 16, 2135–2142. [Google Scholar] [CrossRef]
- Mei, J.; Yue, H.M.; Li, Z.; Chen, B.; Zhong, J.X.; Dan, C.; Zhou, L.; Gui, J.F. C1q-like factor, a target of miR-430, regulates pri-mordial germ cell development in early embryos of Carassius auratus. Int. J. Biol. Sci. 2014, 10, 15. [Google Scholar] [CrossRef]
- Giraldez, A.J.; Cinalli, R.M.; Glasner, M.E.; Enright, A.; Thomson, J.M.; Baskerville, S.; Hammond, S.M.; Bartel, D.P.; Schier, A.F. MicroRNAs regulate brain morphogenesis in zebrafish. Science 2005, 308, 833–838. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, P.; Flemr, M. The role of miRNAs and endogenous siRNAs in maternal-to-zygotic reprogramming and the estab-lishment of pluripotency. EMBO Rep. 2010, 11, 590–597. [Google Scholar] [CrossRef]
- Thatcher, E.J.; Bond, J.; Paydar, I.; Patton, J.G. Genomic Organization of Zebrafish microRNAs. BMC Genom. 2008, 9, 253. [Google Scholar] [CrossRef]
- Tani, S.; Kusakabe, R.; Naruse, K.; Sakamoto, H.; Inoue, K. Genomic organization and embryonic expression of miR-430 in medaka (Oryzias latipes): Insights into the post-transcriptional gene regulation in early development. Gene 2010, 449, 41–49. [Google Scholar] [CrossRef]
- Liu, C. Rudimentary hermaphroditism in the symbranchoid eel, Monopterus javanensis. Sinensia 1944, 15, 1–8. [Google Scholar]
- Gao, Y.; Guo, W.; Hu, Q.; Zou, M.; Tang, R.; Chi, W.; Li, D. Characterization and differential expression patterns of conserved microRNAs and mRNAs in three genders of the rice field eel (Monopterus albus). Sex. Dev. 2014, 8, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Chi, W.; Gao, Y.; Hu, Q.; Guo, W.; Li, D. Genome-wide analysis of brain and gonad transcripts reveals changes of key sex re-versal-related genes expression and signaling pathways in three stages of Monopterus albus. PLoS ONE 2017, 12, e0173974. [Google Scholar] [CrossRef]
- Tao, W.; Sun, L.; Shi, H.; Cheng, Y.; Jiang, D.; Fu, B.; Conte, M.A.; Gammerdinger, W.J.; Kocher, T.D.; Wang, D. Integrated analysis of miRNA and mRNA expression profiles in tilapia gonads at an early stage of sex differentiation. BMC Genom. 2016, 17, 328. [Google Scholar] [CrossRef]
- Steffen, P.; Voss, B.; Rehmsmeier, M.; Reeder, J.; Giegerich, R. RNAshapes: An integrated RNA analysis package based on abstract shapes. Bioinformatics 2005, 22, 500–503. [Google Scholar] [CrossRef]
- Nakamura, M.; Kobayashi, T.; Chang, X.-T.; Nagahama, Y. Gonadal sex differentiation in teleost fish. J. Exp. Zoȯl. 1998, 281, 362–372. [Google Scholar] [CrossRef]
- Pannetier, M.; Elzaiat, M.; Thépot, D.; Pailhoux, E. Telling the Story of XX Sex Reversal in the goat: Highlighting the Ssex-crossroad in domestic mammals. Sex. Dev. 2012, 6, 33–45. [Google Scholar] [CrossRef]
- Chen, Y.J.; Hsiao, P.W.; Lee, M.T.; Mason, J.I.; Ke, F.C.; Hwang, J.J. Interplay of PI3K and cAMP/PKA signaling, and rapamycin hypersensitivity in TGFb1 enhancement of FSH-stimulated steroidogenesis in rat ovarian granulosa cells. J. Endocrinol. 2006, 192, 405–419. [Google Scholar] [CrossRef]
- Manna, P.R.; Chandrala, S.P.; Jo, Y.; Stocco, D. cAMP-independent signaling regulates steroidogenesis in mouse Leydig cells in the absence of StAR phosphorylation. J. Mol. Endocrinol. 2006, 37, 81–95. [Google Scholar] [CrossRef] [PubMed]
- Clarke, T.R.; A Bain, P.; Greco, T.L.; Payne, A.H. A novel mouse kidney 3 beta-hydroxysteroid dehydrogenase complementary DNA encodes a 3-ketosteroid reductase instead of a 3 beta-hydroxysteroid dehydrogenase/delta 5-delta 4-isomerase. Mol. Endocrinol. 1993, 7, 1569–1578. [Google Scholar] [CrossRef][Green Version]
- Makker, A.; Goel, M.M.; Mahdi, A.A. PI3K/PTEN/Akt and TSC/mTOR signaling pathways, ovarian dysfunction, and infertility: An update. J. Mol. Endocrinol. 2014, 53, R103–R118. [Google Scholar] [CrossRef]
- Giraldez, A.J.; Mishima, Y.; Rihel, J.; Grocock, R.J.; Van Dongen, S.; Inoue, K.; Enright, A.; Schier, A.F. Zebrafish MiR-430 Promotes Deadenylation and Clearance of Maternal mRNAs. Science 2006, 312, 75–79. [Google Scholar] [CrossRef]
- Hoffmann, J.; Oris, J. Altered gene expression: A mechanism for reproductive toxicity in zebrafish exposed to benzo[a]pyrene. Aquat. Toxicol. 2006, 78, 332–340. [Google Scholar] [CrossRef]
- Zhao, S.; Liu, M.F. Mechanisms of microRNA-mediated gene regulation. Sci. China Ser. C 2009, 52, 1111–1116. [Google Scholar] [CrossRef]
- Lassiter, C.S.; Linney, E. Embryonic expression and steroid regulation of brain aromatase cyp19a1b in zebrafish (Danio rerio). Zebrafish 2007, 4, 49–58. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, S.; Liu, Z.; Zhang, L.; Zhang, W. Epigenetic modifications during sex change repress gonadotropin stimulation of cyp19a1a in a teleost ricefield eel (Monopterus albus). Endocrinology 2013, 154, 2881–2890. [Google Scholar] [CrossRef]
- Jing, J.; Wu, J.; Liu, W.; Xiong, S.; Ma, W.; Zhang, J.; Wang, W.; Gui, J.-F.; Mei, J. Sex-biased miRNAs in gonad and their potential roles for testis development in yellow catfish. PLoS ONE 2014, 9, e107946. [Google Scholar] [CrossRef]
- Eshel, O.; Shirak, A.; Dor, L.; Band, M.; Zak, T.; Markovich-Gordon, M.; Chalifa-Caspi, V.; Feldmesser, E.; I Weller, J.; Seroussi, E.; et al. Identification of male-specific amh duplication, sexually differentially expressed genes and microRNAs at early embryonic development of Nile tilapia (Oreochromis niloticus). BMC Genom. 2014, 15, 1–18. [Google Scholar] [CrossRef]
- Xiao, J.; Zhong, H.; Zhou, Y.; Yu, F.; Gao, Y.; Luo, Y.J.; Tang, Z.Y.; Guo, Z.B.; Guo, E.Y.; Gan, X.; et al. Identi-fication and characterization of microRNAs in ovary and testis of Nile tilapia (Oreochromis niloticus) by using solexa sequencing technology. PLoS ONE 2014, 9, e86821. [Google Scholar]
- Xiao, Q.; Sun, Y.; Liang, X.; Zhang, L.; Onxayvieng, K.; Li, Z.; Li, D. Visualizing primordial germ cell migration in embryos of rice field eel (Monopterus albus) using fluorescent protein tagged 3′ untranslated regions of nanos3, dead end and vasa. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 235, 62–69. [Google Scholar] [CrossRef]
- Zhao, H.; Soufan, O.; Xia, J.; Tang, R.; Li, L.; Li, D. Transcriptome and physiological analysis reveal alterations in muscle me-tabolisms and immune responses of grass carp (Ctenopharyngodon idellus) cultured at different stocking densities. Aquaculture 2019, 503, 186–197. [Google Scholar] [CrossRef]
- Li, X.; Cooper, N.G.F.; O’Toole, T.E.; Rouchka, E.C. Choice of library size normalization and statistical methods for differential gene expression analysis in balanced two-group comparisons for RNA-seq studies. BMC Genom. 2020, 21, 1–17. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Fang, S.; Li, S.; Zheng, H.; Zhang, Y.; Ikhwanuddin, M.; Ma, H. mRNA profile provides novel insights into stress adaptation in mud crab megalopa, Scylla paramamosain after salinity stress. BMC Genom. 2020, 21, 1–16. [Google Scholar] [CrossRef]
- A Rupp, R.; Snider, L.; Weintraub, H. Xenopus embryos regulate the nuclear localization of XMyoD. Genes Dev. 1994, 8, 1311–1323. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Yang, Q.; Xu, W.; Wu, Z.; Li, D. Integrated Analysis of miR-430 on Steroidogenesis-Related Gene Expression of Larval Rice Field Eel Monopterus albus. Int. J. Mol. Sci. 2021, 22, 6994. https://doi.org/10.3390/ijms22136994
Zhang L, Yang Q, Xu W, Wu Z, Li D. Integrated Analysis of miR-430 on Steroidogenesis-Related Gene Expression of Larval Rice Field Eel Monopterus albus. International Journal of Molecular Sciences. 2021; 22(13):6994. https://doi.org/10.3390/ijms22136994
Chicago/Turabian StyleZhang, Lihan, Qiushi Yang, Weitong Xu, Zhaojun Wu, and Dapeng Li. 2021. "Integrated Analysis of miR-430 on Steroidogenesis-Related Gene Expression of Larval Rice Field Eel Monopterus albus" International Journal of Molecular Sciences 22, no. 13: 6994. https://doi.org/10.3390/ijms22136994
APA StyleZhang, L., Yang, Q., Xu, W., Wu, Z., & Li, D. (2021). Integrated Analysis of miR-430 on Steroidogenesis-Related Gene Expression of Larval Rice Field Eel Monopterus albus. International Journal of Molecular Sciences, 22(13), 6994. https://doi.org/10.3390/ijms22136994