AKAP12 Supports Blood-Brain Barrier Integrity against Ischemic Stroke

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
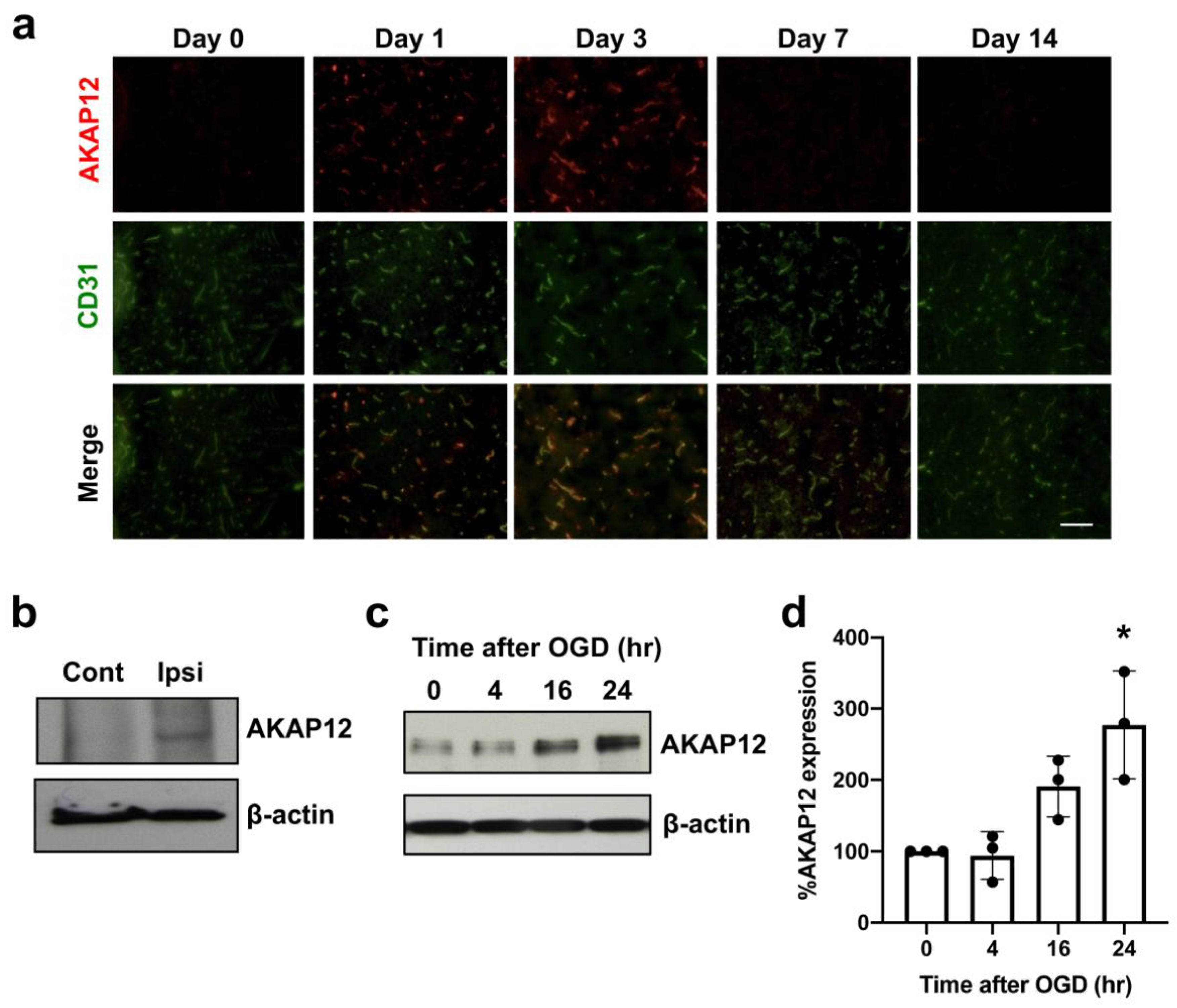
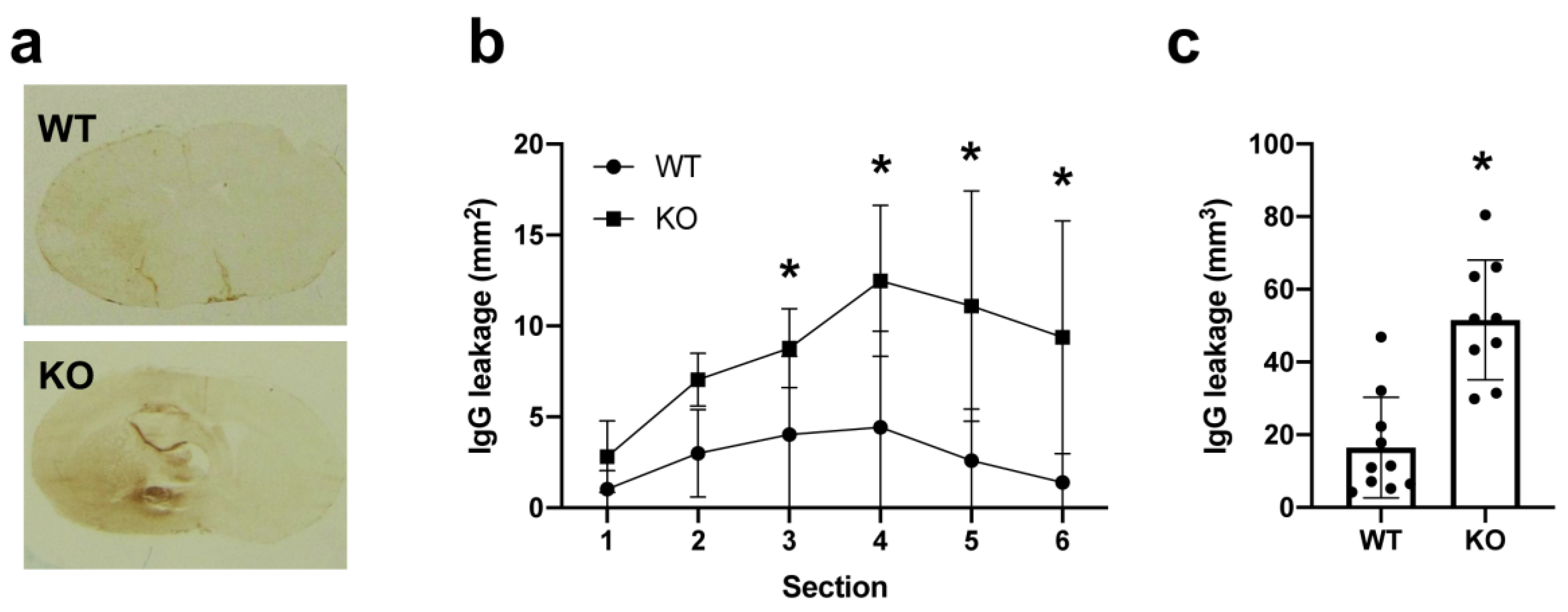
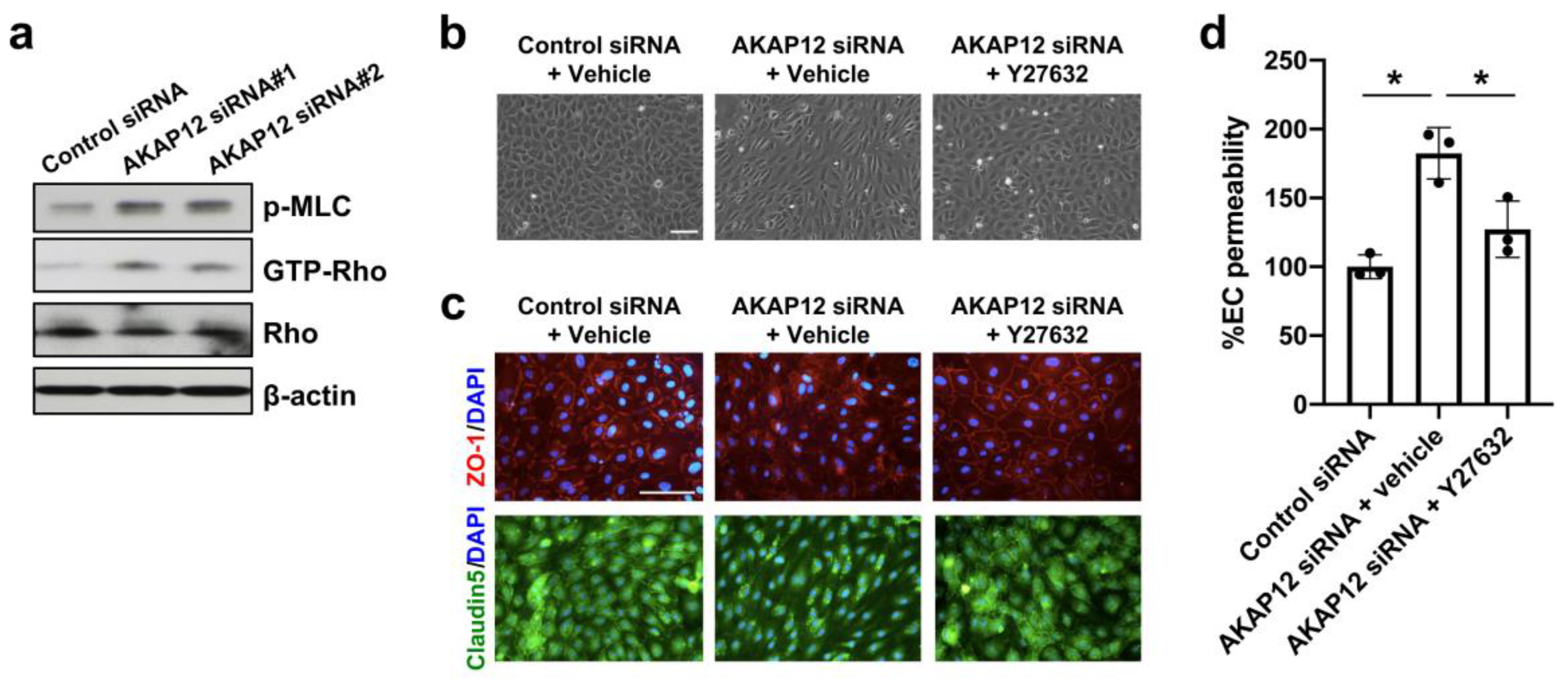
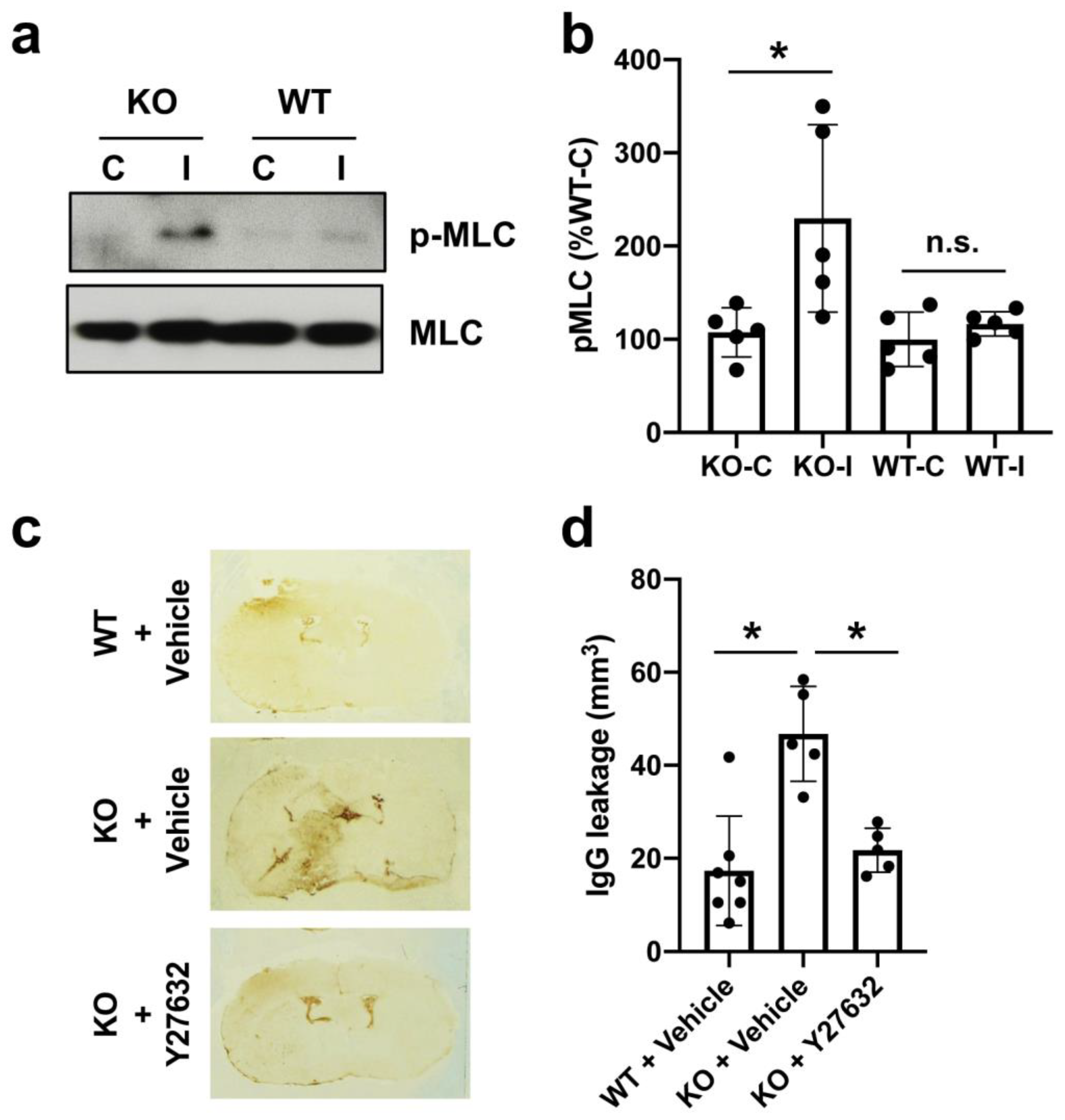
2. Results
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Focal Stroke Model in Mice
4.3. Cell Culture
4.4. In Vitro Stroke Model
4.5. In Vitro Endothelial Permeability Assay
4.6. Y-27632 Treatment
4.7. Western Blot
4.8. Brain Microvessel Isolation
4.9. Rho Pull-Down Assay
4.10. Immunocytochemistry
4.11. Immunohistochemistry
4.12. Fluorescence-Activated Cell Sorting (FACS)
4.13. Data Analysis and Statistics
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AKAP12 | A-kinase anchor protein 12 |
| Ang-1 | Angiopoetin-1 |
| BBB | Blood-brain barrier |
| HBMECs | Human brain microvascular endothelial cells |
| HMEC-1 | Human microvascular endothelial cell line 1 |
| NVU | Neurovascular unit |
| MCA | Middle cerebral artery |
| MLC | Myosin light chain |
| OGD | Oxygen-glucose deprivation |
| PKA | Protein kinase A |
| PKC | Protein kinase C |
| PTX3 | Pentraxin 3 |
| TIMP-1 | Tissue inhibitor of metalloproteinase 1 |
References
- Virani, S.S.; Alonso, A.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics-2020 Update: A Report From the American Heart Association. Circulation 2020, 141, e139–e596. [Google Scholar] [CrossRef]
- Van Leyen, K.; Wang, X.; Selim, M.; Lo, E.H. Opening the time window. J. Cereb. Blood Flow Metab. 2019, 39, 2539–2540. [Google Scholar] [CrossRef]
- Shi, L.; Rocha, M.; Leak, R.K.; Zhao, J.; Bhatia, T.N.; Mu, H.; Wei, Z.; Yu, F.; Weiner, S.L.; Ma, F.; et al. A new era for stroke therapy: Integrating neurovascular protection with optimal reperfusion. J. Cereb. Blood Flow Metab. 2018, 38, 2073–2091. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H.; Ning, M. Mechanisms and challenges in translational stroke research. J. Investig. Med. 2016, 64, 827–829. [Google Scholar] [CrossRef] [PubMed]
- Xing, C.; Arai, K.; Lo, E.H.; Hommel, M. Pathophysiologic cascades in ischemic stroke. Int. J. Stroke 2012, 7, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Zhong, W.; Hu, Z.; Tang, X. A review of the role of cav-1 in neuropathology and neural recovery after ischemic stroke. J. Neuroinflamm. 2018, 15, 348. [Google Scholar] [CrossRef]
- Gelman, I.H. Suppression of tumor and metastasis progression through the scaffolding functions of SSeCKS/Gravin/AKAP12. Cancer Metastasis Rev. 2012, 31, 493–500. [Google Scholar] [CrossRef]
- Wu, X.; Wu, T.; Li, K.; Li, Y.; Hu, T.T.; Wang, W.F.; Qiang, S.J.; Xue, S.B.; Liu, W.W. The Mechanism and Influence of AKAP12 in Different Cancers. Biomed. Environ. Sci. 2018, 31, 927–932. [Google Scholar] [CrossRef]
- Gelman, I.H. Emerging Roles for SSeCKS/Gravin/AKAP12 in the Control of Cell Proliferation, Cancer Malignancy, and Barriergenesis. Genes Cancer 2010, 1, 1147–1156. [Google Scholar] [CrossRef]
- Maki, T.; Choi, Y.K.; Miyamoto, N.; Shindo, A.; Liang, A.C.; Ahn, B.J.; Mandeville, E.T.; Kaji, S.; Itoh, K.; Seo, J.H.; et al. A-Kinase Anchor Protein 12 Is Required for Oligodendrocyte Differentiation in Adult White Matter. Stem Cells 2018. [Google Scholar] [CrossRef]
- Cha, J.H.; Wee, H.J.; Seo, J.H.; Ahn, B.J.; Park, J.H.; Yang, J.M.; Lee, S.W.; Kim, E.H.; Lee, O.H.; Heo, J.H.; et al. AKAP12 mediates barrier functions of fibrotic scars during CNS repair. PLoS ONE 2014, 9, e94695. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.W.; Kim, W.J.; Choi, Y.K.; Song, H.S.; Son, M.J.; Gelman, I.H.; Kim, Y.J.; Kim, K.W. SSeCKS regulates angiogenesis and tight junction formation in blood-brain barrier. Nat. Med. 2003, 9, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Sweeney, M.D.; Sagare, A.P.; Zlokovic, B.V. Blood-brain barrier breakdown in Alzheimer disease and other neurodegenerative disorders. Nat. Rev. Neurol. 2018, 14, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Abbott, N.J.; Patabendige, A.A.; Dolman, D.E.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25. [Google Scholar] [CrossRef]
- Ronaldson, P.T.; Davis, T.P. Regulation of blood-brain barrier integrity by microglia in health and disease: A therapeutic opportunity. J. Cereb. Blood Flow Metab. 2020. [Google Scholar] [CrossRef]
- Zan, L.; Wu, H.; Jiang, J.; Zhao, S.; Song, Y.; Teng, G.; Li, H.; Jia, Y.; Zhou, M.; Zhang, X.; et al. Temporal profile of Src, SSeCKS, and angiogenic factors after focal cerebral ischemia: Correlations with angiogenesis and cerebral edema. Neurochem. Int. 2011, 58, 872–879. [Google Scholar] [CrossRef]
- Choi, Y.K.; Kim, K.W. AKAP12 in astrocytes induces barrier functions in human endothelial cells through protein kinase Czeta. FEBS J. 2008, 275, 2338–2353. [Google Scholar] [CrossRef]
- Gelman, I.H.; Gao, L. SSeCKS/Gravin/AKAP12 metastasis suppressor inhibits podosome formation via RhoA- and Cdc42-dependent pathways. Mol. Cancer Res. 2006, 4, 151–158. [Google Scholar] [CrossRef]
- Shin, H.K.; Salomone, S.; Ayata, C. Targeting cerebrovascular Rho-kinase in stroke. Expert Opin. Ther. Targets 2008, 12, 1547–1564. [Google Scholar] [CrossRef]
- Park, J.C.; Baik, S.H.; Han, S.H.; Cho, H.J.; Choi, H.; Kim, H.J.; Choi, H.; Lee, W.; Kim, D.K.; Mook-Jung, I. Annexin A1 restores Abeta1-42 -induced blood-brain barrier disruption through the inhibition of RhoA-ROCK signaling pathway. Aging Cell 2017, 16, 149–161. [Google Scholar] [CrossRef]
- Amano, M.; Ito, M.; Kimura, K.; Fukata, Y.; Chihara, K.; Nakano, T.; Matsuura, Y.; Kaibuchi, K. Phosphorylation and activation of myosin by Rho-associated kinase (Rho-kinase). J. Biol. Chem. 1996, 271, 20246–20249. [Google Scholar] [CrossRef] [PubMed]
- Maki, T.; Hayakawa, K.; Pham, L.D.; Xing, C.; Lo, E.H.; Arai, K. Biphasic mechanisms of neurovascular unit injury and protection in CNS diseases. CNS Neurol. Disord. Drug Targets 2013, 12, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H. A new penumbra: Transitioning from injury into repair after stroke. Nat. Med. 2008, 14, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A.; Estrada, E.Y.; Dencoff, J.E. Matrix metalloproteinases and TIMPs are associated with blood-brain barrier opening after reperfusion in rat brain. Stroke 1998, 29, 2189–2195. [Google Scholar] [CrossRef]
- Shindo, A.; Maki, T.; Mandeville, E.T.; Liang, A.C.; Egawa, N.; Itoh, K.; Itoh, N.; Borlongan, M.; Holder, J.C.; Chuang, T.T.; et al. Astrocyte-Derived Pentraxin 3 Supports Blood-Brain Barrier Integrity Under Acute Phase of Stroke. Stroke 2016, 47, 1094–1100. [Google Scholar] [CrossRef]
- Alfieri, A.; Srivastava, S.; Siow, R.C.M.; Cash, D.; Modo, M.; Duchen, M.R.; Fraser, P.A.; Williams, S.C.R.; Mann, G.E. Sulforaphane preconditioning of the Nrf2/HO-1 defense pathway protects the cerebral vasculature against blood-brain barrier disruption and neurological deficits in stroke. Free Radic. Biol. Med. 2013, 65, 1012–1022. [Google Scholar] [CrossRef]
- Wang, S.; Cao, C.; Chen, Z.; Bankaitis, V.; Tzima, E.; Sheibani, N.; Burridge, K. Pericytes regulate vascular basement membrane remodeling and govern neutrophil extravasation during inflammation. PLoS ONE 2012, 7, e45499. [Google Scholar] [CrossRef]
- Dalkara, T.; Alarcon-Martinez, L. Cerebral microvascular pericytes and neurogliovascular signaling in health and disease. Brain Res. 2015, 1623, 3–17. [Google Scholar] [CrossRef]
- Ronaldson, P.T.; Davis, T.P. Blood-brain barrier integrity and glial support: Mechanisms that can be targeted for novel therapeutic approaches in stroke. Curr. Pharm. Des. 2012, 18, 3624–3644. [Google Scholar] [CrossRef]
- Ballabh, P.; Braun, A.; Nedergaard, M. The blood-brain barrier: An overview: Structure, regulation, and clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [Google Scholar] [CrossRef]
- Beard, D.J.; Brown, L.S.; Sutherland, B.A. The rise of pericytes in neurovascular research. J. Cereb. Blood Flow Metab. 2020. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.K.; Kim, J.H.; Kim, W.J.; Lee, H.Y.; Park, J.A.; Lee, S.W.; Yoon, D.K.; Kim, H.H.; Chung, H.; Yu, Y.S.; et al. AKAP12 regulates human blood-retinal barrier formation by downregulation of hypoxia-inducible factor-1alpha. J. Neurosci. 2007, 27, 4472–4481. [Google Scholar] [CrossRef] [PubMed]
- Andjelkovic, A.V.; Stamatovic, S.M.; Phillips, C.M.; Martinez-Revollar, G.; Keep, R.F. Modeling blood-brain barrier pathology in cerebrovascular disease in vitro: Current and future paradigms. Fluids Barriers CNS 2020, 17, 44. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Guo, S.; Lok, J.; Navaratna, D.; Whalen, M.J.; Kim, K.W.; Lo, E.H. Neurovascular matrix metalloproteinases and the blood-brain barrier. Curr. Pharm. Des. 2012, 18, 3645–3648. [Google Scholar] [CrossRef]
- Hayakawa, K.; Qiu, J.; Lo, E.H. Biphasic actions of HMGB1 signaling in inflammation and recovery after stroke. Ann. N. Y. Acad. Sci. 2010, 1207, 50–57. [Google Scholar] [CrossRef]
- Zhao, B.Q.; Wang, S.; Kim, H.Y.; Storrie, H.; Rosen, B.R.; Mooney, D.J.; Wang, X.; Lo, E.H. Role of matrix metalloproteinases in delayed cortical responses after stroke. Nat. Med. 2006, 12, 441–445. [Google Scholar] [CrossRef]
- Murata, Y.; Fujiwara, N.; Seo, J.H.; Yan, F.; Liu, X.; Terasaki, Y.; Luo, Y.; Arai, K.; Ji, X.; Lo, E.H. Delayed inhibition of c-Jun N-terminal kinase worsens outcomes after focal cerebral ischemia. J. Neurosci. 2012, 32, 8112–8115. [Google Scholar] [CrossRef]
- Xing, C.; Hayakawa, K.; Lok, J.; Arai, K.; Lo, E.H. Injury and repair in the neurovascular unit. Neurol. Res. 2012, 34, 325–330. [Google Scholar] [CrossRef]
- Su, B.; Zheng, Q.; Vaughan, M.M.; Bu, Y.; Gelman, I.H. SSeCKS metastasis-suppressing activity in MatLyLu prostate cancer cells correlates with vascular endothelial growth factor inhibition. Cancer Res. 2006, 66, 5599–5607. [Google Scholar] [CrossRef]
- Weissmuller, T.; Glover, L.E.; Fennimore, B.; Curtis, V.F.; MacManus, C.F.; Ehrentraut, S.F.; Campbell, E.L.; Scully, M.; Grove, B.D.; Colgan, S.P. HIF-dependent regulation of AKAP12 (gravin) in the control of human vascular endothelial function. FASEB J. 2014, 28, 256–264. [Google Scholar] [CrossRef]
- Hawkins, B.T.; Davis, T.P. The blood-brain barrier/neurovascular unit in health and disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Qasim, H.; McConnell, B.K. AKAP12 Signaling Complex: Impacts of Compartmentalizing cAMP-Dependent Signaling Pathways in the Heart and Various Signaling Systems. J. Am. Heart Assoc. 2020, 9, e016615. [Google Scholar] [CrossRef] [PubMed]
- Akakura, S.; Huang, C.; Nelson, P.J.; Foster, B.; Gelman, I.H. Loss of the SSeCKS/Gravin/AKAP12 gene results in prostatic hyperplasia. Cancer Res. 2008, 68, 5096–5103. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Pham, L.D.; Katusic, Z.S.; Arai, K.; Lo, E.H. Astrocytic high-mobility group box 1 promotes endothelial progenitor cell-mediated neurovascular remodeling during stroke recovery. Proc. Natl. Acad Sci. USA 2012, 109, 7505–7510. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.H.; Miyamoto, N.; Hayakawa, K.; Pham, L.D.; Maki, T.; Ayata, C.; Kim, K.W.; Lo, E.H.; Arai, K. Oligodendrocyte precursors induce early blood-brain barrier opening after white matter injury. J. Clin. Investig. 2013, 123, 782–786. [Google Scholar] [CrossRef]
- Guo, S.; Zhou, Y.; Xing, C.; Lok, J.; Som, A.T.; Ning, M.; Ji, X.; Lo, E.H. The vasculome of the mouse brain. PLoS ONE 2012, 7, e52665. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seo, J.H.; Maki, T.; Miyamoto, N.; Choi, Y.K.; Chung, K.K.; Hamanaka, G.; Park, J.H.; Mandeville, E.T.; Takase, H.; Hayakawa, K.; et al. AKAP12 Supports Blood-Brain Barrier Integrity against Ischemic Stroke. Int. J. Mol. Sci. 2020, 21, 9078. https://doi.org/10.3390/ijms21239078
Seo JH, Maki T, Miyamoto N, Choi YK, Chung KK, Hamanaka G, Park JH, Mandeville ET, Takase H, Hayakawa K, et al. AKAP12 Supports Blood-Brain Barrier Integrity against Ischemic Stroke. International Journal of Molecular Sciences. 2020; 21(23):9078. https://doi.org/10.3390/ijms21239078
Chicago/Turabian StyleSeo, Ji Hae, Takakuni Maki, Nobukazu Miyamoto, Yoon Kyong Choi, Kelly K. Chung, Gen Hamanaka, Ji Hyun Park, Emiri T. Mandeville, Hajime Takase, Kazuhide Hayakawa, and et al. 2020. "AKAP12 Supports Blood-Brain Barrier Integrity against Ischemic Stroke" International Journal of Molecular Sciences 21, no. 23: 9078. https://doi.org/10.3390/ijms21239078
APA StyleSeo, J. H., Maki, T., Miyamoto, N., Choi, Y. K., Chung, K. K., Hamanaka, G., Park, J. H., Mandeville, E. T., Takase, H., Hayakawa, K., Lok, J., Gelman, I. H., Kim, K.-W., Lo, E. H., & Arai, K. (2020). AKAP12 Supports Blood-Brain Barrier Integrity against Ischemic Stroke. International Journal of Molecular Sciences, 21(23), 9078. https://doi.org/10.3390/ijms21239078