Adjustment of Photosynthetic and Antioxidant Activities to Water Deficit Is Crucial in the Drought Tolerance of Lolium multiflorum/Festuca arundinacea Introgression Forms

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Selection of L. multiflorum/F. arundinacea Introgression Forms with Distinct Levels of Drought Tolerance in the Field Conditions
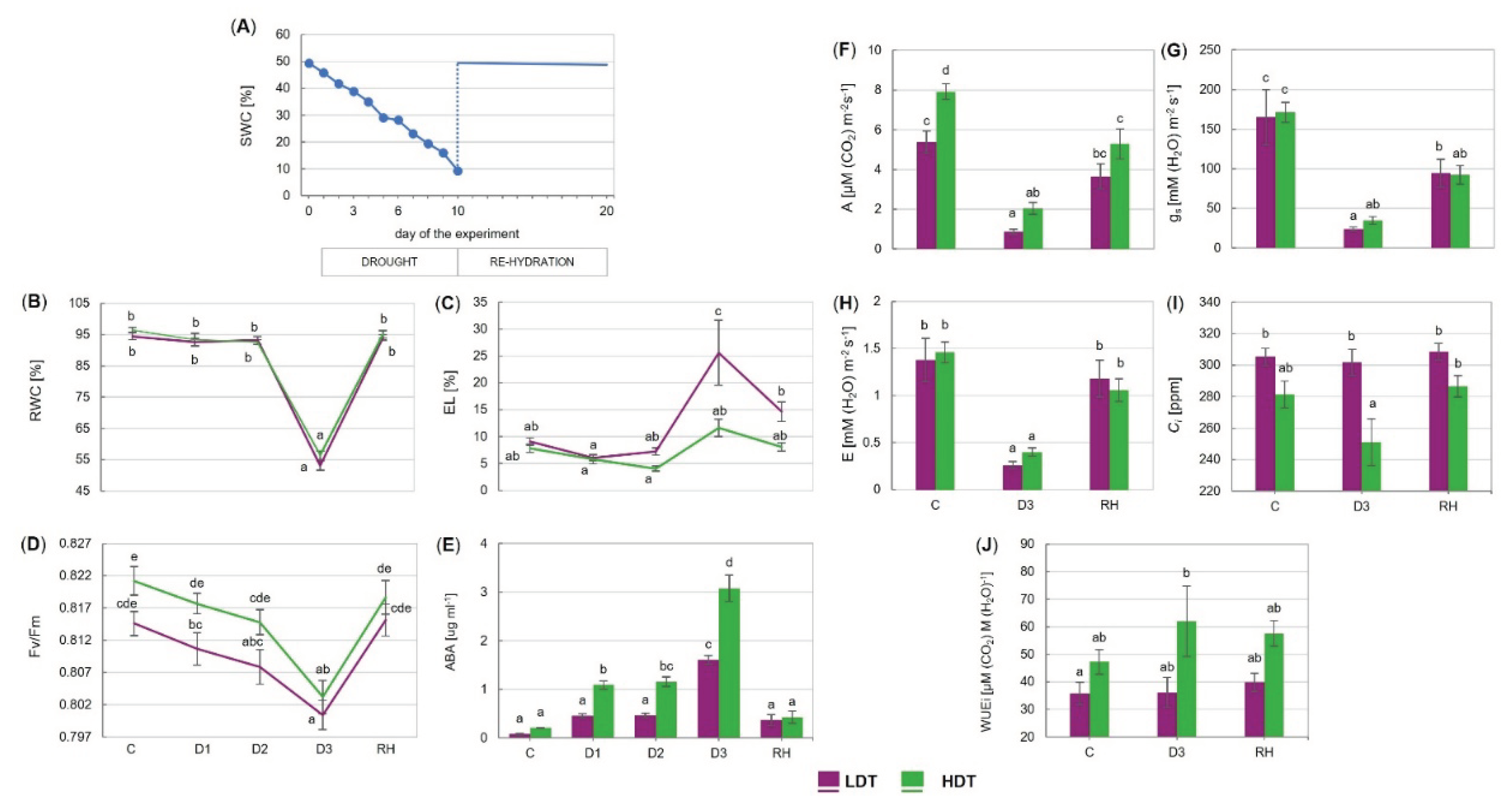
2.2. Physiological Performance of the LDT and HDT Introgression Forms under Drought and Re-Watering Conditions in the Environmental Chamber
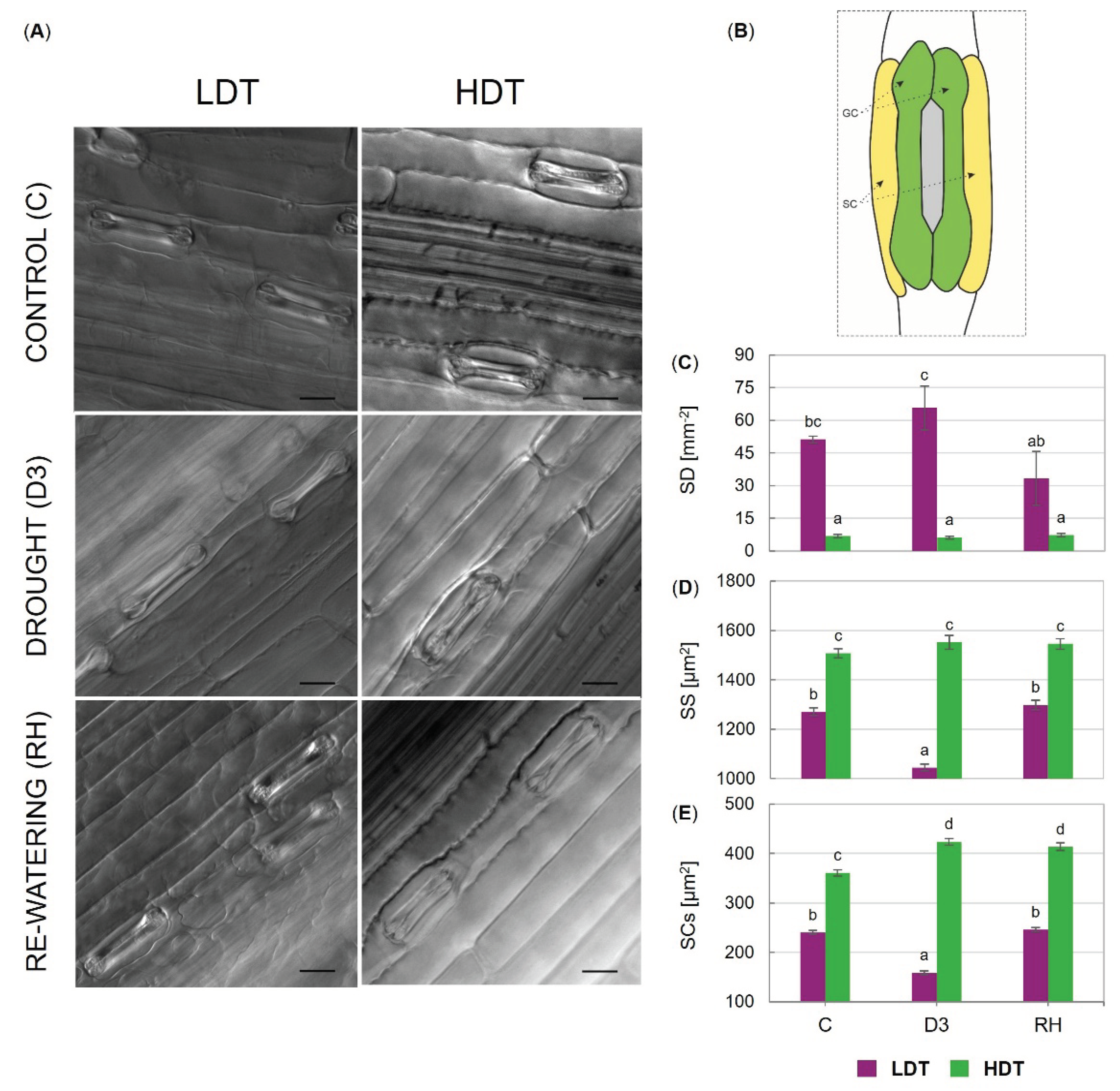
2.3. Stomatal Characteristics
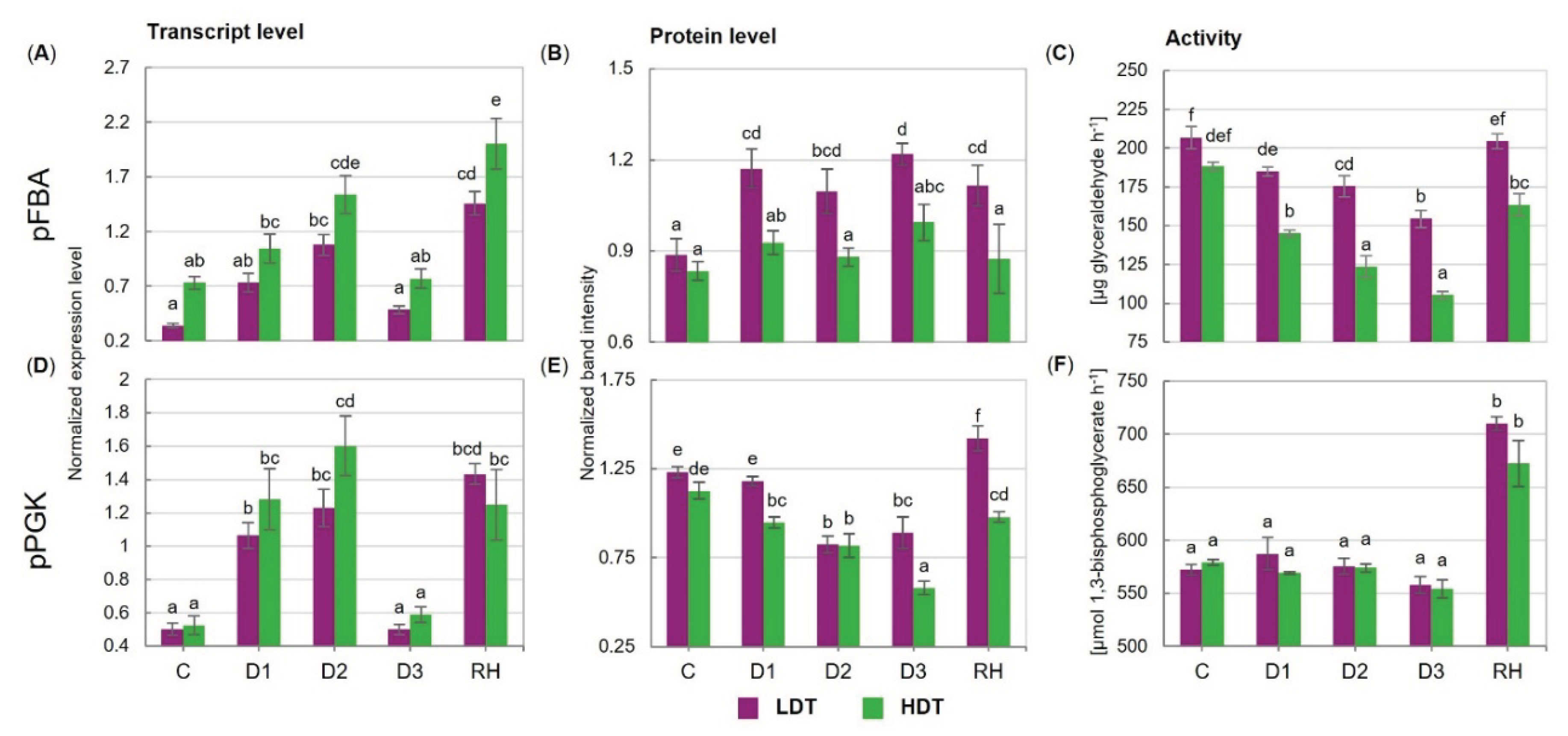
2.4. Expression of Genes Encoding the Calvin Cycle Enzymes at Transcript and Protein Levels
2.5. Activities of the Calvin Cycle Enzymes
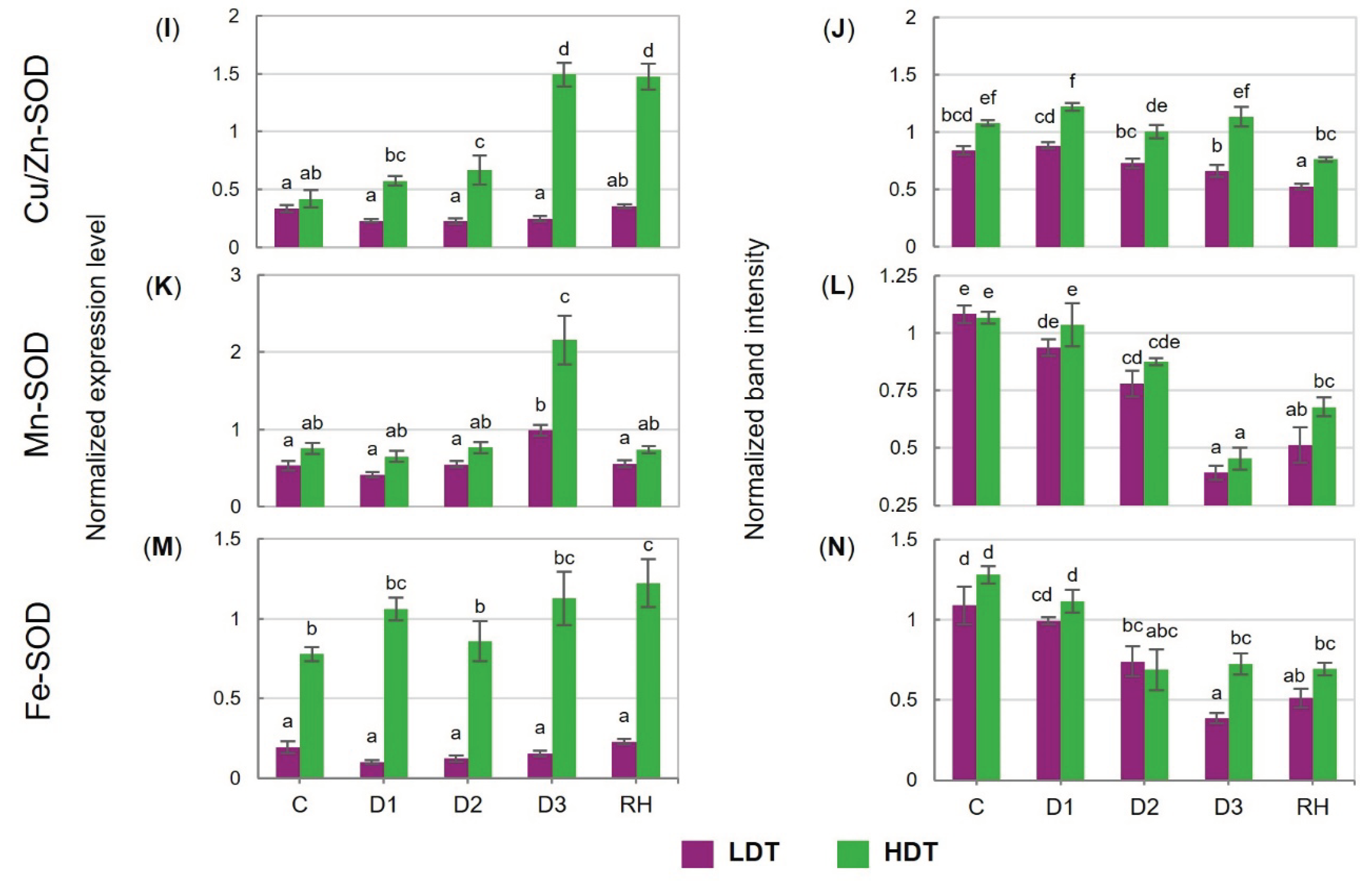
2.6. Expression of Genes Encoding Antioxidant Enzymes at Transcript and Protein Levels
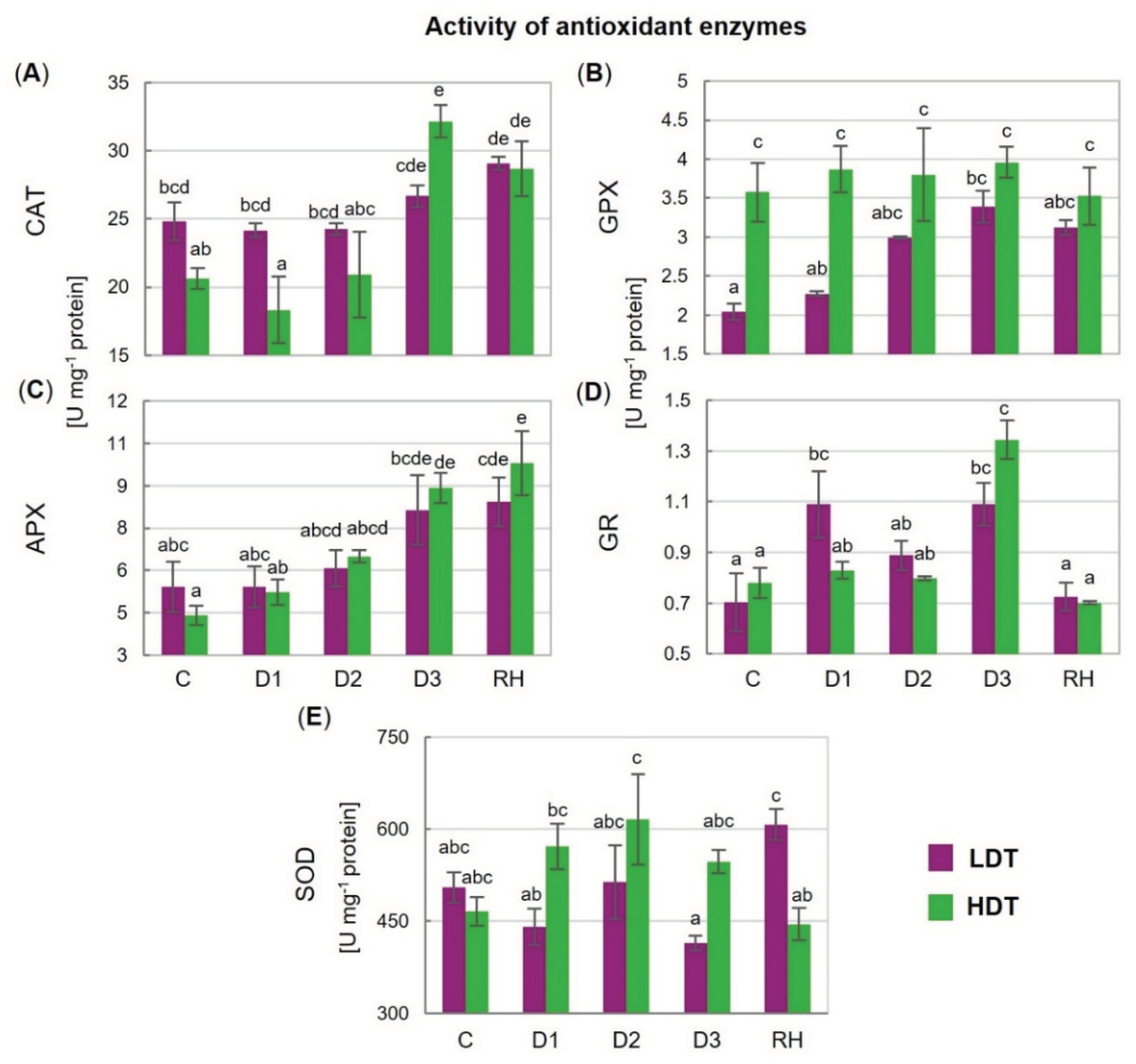
2.7. Activities of Antioxidant Enzymes
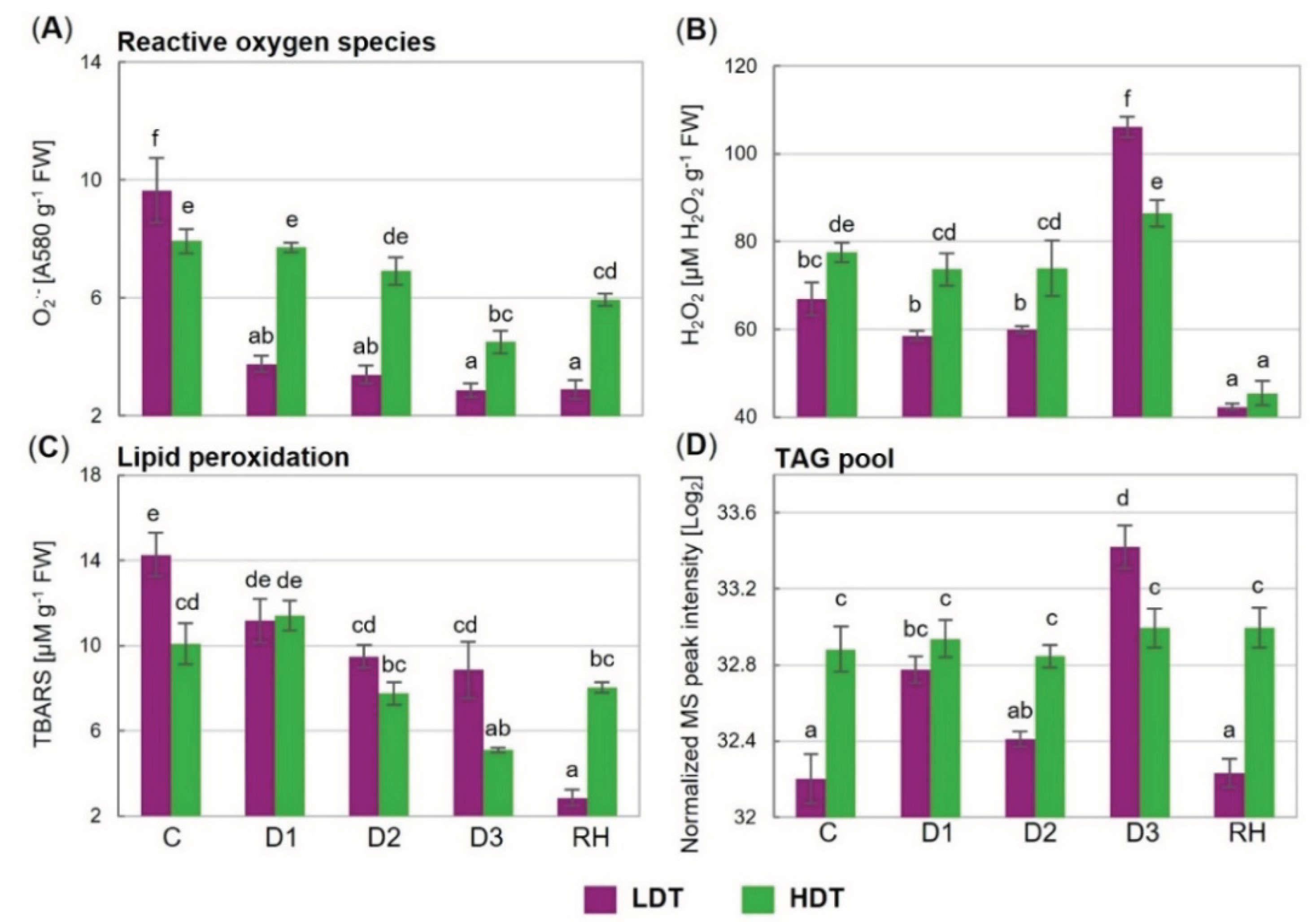
2.8. Superoxide Anion Radical and Hydrogen Peroxide Content
2.9. Lipid Peroxidation Level
2.10. Triacylglycerol Lipid Accumulation
3. Discussion
3.1. Plant Selection Following Prolonged Drought and Subsequent Re-Watering under Field Conditions
3.2. Physiological Response to Short-Term Drought
3.3. Gas Exchange, Stomata, and Photosynthetic Performance
3.4. Reactive Oxygen Species and Antioxidant Capacity
4. Materials and Methods
4.1. Plant Materials
4.2. Plant Selection with Respect to Drought Tolerance under Field Conditions
4.3. Short-Term Drought Conditions in the Environmental Chamber
4.4. Physiological Parameters
4.5. Anatomy and Density of Stomata
4.6. RT-qPCR of the Calvin Cycle and Antioxidant Enzymes
4.7. Western Blot of the Calvin Cycle and Antioxidant Enzymes
4.8. Activity of the Calvin Cycle Enzymes
4.9. Activity of Antioxidant Enzymes
4.10. TBARS Content
4.11. Superoxide Anion Radical and Hydrogen Peroxide Content
4.12. TAG Accumulation
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A | CO2 assimilation |
| ABA | abscisic acid |
| APX | L-ascorbate peroxidase |
| CAT | catalase |
| Ci | internal CO2 concentration |
| Cu/Zn-SOD | copper/zinc superoxide dismutase |
| E | transpiration |
| EL | electrolyte leakage |
| FA | fatty acids |
| Fe-SOD | iron superoxide dismutase |
| Fv/Fm | maximum quantum efficiency of PSII photochemistry |
| FW | fresh weight |
| GC | guard cells |
| gs | stomatal conductance |
| GPX | glutathione peroxidase |
| GR | glutathione reductase |
| HDT | high drought tolerance |
| LDT | low drought tolerance |
| Mn-SOD | manganese superoxide dismutase |
| SC | subsidiary cells |
| SD | stomatal density |
| SOD | superoxide dismutase |
| SS | stomatal size |
| SWC | soil water capacity |
| pFBA | plastid fructose-1,6-bisphosphate aldolase |
| pPGK | plastid phosphoglycerate kinase |
| PSII | photosystem II |
| ROS | reactive oxygen species |
| RWC | relative water content |
| TAG TBARS | triacylglycerol thiobarbituric-reactive substances |
| WUEi | intrinsic water use efficiency |
References
- Spinoni, J.; Vogt, J.V.; Naumann, G.; Barbosa, P.; Dosio, A. Will Drought Events Become More Frequent and Severe in Europe? Int. J. Climatol. 2018, 38, 1718–1736. [Google Scholar] [CrossRef]
- Xu, Z.; Zhou, G. Responses of Leaf Stomatal Density to Water Status and its Relationship with Photosynthesis in a Grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J.; Niinemets, Ü.; Gallé, A.; Barbour, M.M.; Centritto, M.; Diaz-Espejo, A.; Douthe, C.; Galmés, J.; Ribas-Carbo, M.; Rodriguez, P.L.; et al. Diffusional Conductances to Co2 as a Target for Increasing Photosynthesis and Photosynthetic Water-Use Efficiency. Photosynth. Res. 2013, 117, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of Drought Stress on Photosynthesis and Photosynthetic Electron Transport Chain in Young Apple Tree Leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef] [PubMed]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under Drought and Salt Stress: Regulation Mechanisms from Whole Plant to Cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Soares-Cordeiro, A.S.; Driscoll, S.P.; Pellny, T.K.; Olmos, E.; Arrabaça, M.C.; Foyer, C.H. Variations in the Dorso-Ventral Organization of Leaf Structure and Kranz Anatomy Coordinate the Control of Photosynthesis and Associated Signalling at the Whole Leaf Level in Monocotyledonous Species. Plant. Cell Environ. 2009, 32, 1833–1844. [Google Scholar] [CrossRef]
- Pena-Rojas, K.; Aranda, X.; Fleck, I. Stomatal Limitation to CO2 Assimilation and Down-Regulation of Photosynthesis in Quercus Ilex Resprouts in Response to Slowly Imposed Drought. Tree Physiol. 2004, 24, 813–822. [Google Scholar] [CrossRef]
- Lawlor, D.W. Limitation to Photosynthesis in Water-Stressed Leaves: Stomata vs. Metabolism and the Role of ATP. Ann. Bot. 2002, 89, 871–885. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic Carbon Assimilation and Associated Metabolism in Relation to Water Deficits in Higher Plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef]
- Morison, J.I.L.; Lawson, T. Does Lateral Gas Diffusion in Leaves Matter? Plant Cell Environ. 2007, 30, 1072–1085. [Google Scholar] [CrossRef]
- Lake, J.A.; Woodward, F.I. Response of Stomatal Numbers to CO2 and Humidity: Control by Transpiration Rate and Abscisic Acid. New Phytol. 2008, 179, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Sekiya, N.; Yano, K. Stomatal Density of Cowpea Correlates with Carbon Isotope Discrimination in Different Phosphorus, Water and Co2 Environments. New Phytol. 2008, 179, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Fraser, L.H.; Greenall, A.; Carlyle, C.; Turkington, R.; Friedman, C.R. Adaptive Phenotypic Plasticity of Pseudoroegneria Spicata: Response of Stomatal Density, Leaf Area and Biomass to Changes in Water Supply and Increased Temperature. Ann. Bot. 2009, 103, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, J.A.; Cabrera, H.M.; Queirolo, C.; Hinojosa, L.F. Variability of Water Relations and Photosynthesis in Eucryphia Cordifolia cav. (Cunoniaceae) over the Range of its Latitudinal and Altitudinal Distribution in Chile. Tree Physiol. 2010, 30, 574–585. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yan, F.; Sun, Y.; Song, F.; Liu, F. Differential Responses of Stomatal Morphology to Partial Root-Zone Drying and Deficit Irrigation in Potato Leaves Under Varied Nitrogen Rates. Sci. Hortic. (Amsterdam) 2012, 145, 76–83. [Google Scholar] [CrossRef]
- Rudall, P.J.; Chen, E.D.; Cullen, E. Evolution and Development of Monocot Stomata. Am. J. Bot. 2017, 104, 1122–1141. [Google Scholar] [CrossRef]
- Hepworth, C.; Caine, R.S.; Harrison, E.L.; Sloan, J.; Gray, J.E. Stomatal Development: Focusing on the Grasses. Curr. Opin. Plant Biol. 2018, 41, 1–7. [Google Scholar] [CrossRef]
- McKown, K.H.; Bergmann, D.C. Grass Stomata. Curr. Biol. 2018, 28, R814–R816. [Google Scholar] [CrossRef]
- Nunes, T.D.G.; Zhang, D.; Raissig, M.T. Form, Development and Function of Grass Stomata. Plant J. 2020, 101, 780–799. [Google Scholar] [CrossRef]
- Merilo, E.; Jõesaar, I.; Brosché, M.; Kollist, H. To Open or to Close: Species-Specific Stomatal Responses to Simultaneously Applied Opposing Environmental Factors. New Phytol. 2014, 202, 499–508. [Google Scholar] [CrossRef]
- McAusland, L.; Vialet-Chabrand, S.; Davey, P.; Baker, N.R.; Brendel, O.; Lawson, T. Effects of Kinetics of Light-Induced Stomatal Responses on Photosynthesis and Water-Use Efficiency. New Phytol. 2016, 211, 1209–1220. [Google Scholar] [CrossRef]
- Haworth, M.; Scutt, C.P.; Douthe, C.; Marino, G.; Gomes, M.T.G.; Loreto, F.; Flexas, J.; Centritto, M. Allocation of the Epidermis to Stomata Relates to Stomatal Physiological Control: Stomatal Factors Involved in the Evolutionary Diversification of the Angiosperms and Development of Amphistomaty. Environ. Exp. Bot. 2018, 151, 55–63. [Google Scholar] [CrossRef]
- Franks, P.J.; Farquhar, G.D. The Mechanical Diversity of Stomata and its Significance in Gas-Exchange Control. Plant Physiol. 2007, 143, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, N.; Maierhofer, T.; Herrmann, J.; Jørgensen, M.E.; Lind, C.; von Meyer, K.; Lautner, S.; Fromm, J.; Felder, M.; Hetherington, A.M.; et al. A Tandem Amino Acid Residue Motif in Guard Cell Slac1 Anion Channel of Grasses Allows for the Control of Stomatal Aperture by Nitrate. Curr. Biol. 2018, 28, 1370–1379.e5. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.J.; Beerling, D.J. Maximum Leaf Conductance Driven by CO2 Effects on Stomatal Size and Density over Geologic Time. PNAS 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Badger, M.R. Photoprotection in Plants: A New Light on Photosystem II Damage. Trends Plant Sci. 2011, 16, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Marchin, R.M.; Turnbull, T.L.; Deheinzelin, A.I.; Adams, M.A. Does Triacylglycerol (TAG) Serve a Photoprotective Function in Plant Leaves? An Examination of Leaf Lipids under Shading and Drought. Physiol. Plant. 2017, 161, 400–413. [Google Scholar] [CrossRef]
- Gołębiowska-Pikania, G.; Kopeć, P.; Surówka, E.; Janowiak, F.; Krzewska, M.; Dubas, E.; Nowicka, A.; Kasprzyk, J.; Ostrowska, A.; Malaga, S.; et al. Changes in Protein Abundance and Activity Induced by Drought during Generative Development of Winter Barley (Hordeum vulgare L.). J. Proteomics 2017, 169, 73–86. [Google Scholar] [CrossRef]
- Lee, H.G.; Park, M.E.; Park, B.Y.; Kim, H.U.; Seo, P.J. The Arabidopsis MYB96 Transcription Factor Mediates Aba-Dependent Triacylglycerol Accumulation in Vegetative Tissues under Drought Stress Conditions. Plants 2019, 8, 296. [Google Scholar] [CrossRef]
- Lu, J.; Xu, Y.; Wang, J.; Singer, S.D.; Chen, G. The Role of Triacylglycerol in Plant Stress Response. Plants 2020, 9, 472. [Google Scholar] [CrossRef]
- Mueller, S.P.; Unger, M.; Guender, L.; Fekete, A.; Mueller, M.J. Diacylglycerol Acyltransferase-Mediated Triacylglyerol Synthesis Augments Basal Thermotolerance. Plant Physiol. 2017, 175, 486–497. [Google Scholar] [CrossRef] [PubMed]
- Perlikowski, D.; Kierszniowska, S.; Sawikowska, A.; Krajewski, P.; Rapacz, M.; Eckhardt, Ä.; Kosmala, A. Remodeling of Leaf Cellular Glycerolipid Composition under Drought and Re-Hydration Conditions in Grasses from the Lolium-Festuca Complex. Front. Plant Sci. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Perlikowski, D.; Augustyniak, A.; Skirycz, A.; Pawłowicz, I.; Masajada, K.; Michaelis, Ä.; Kosmala, A. Efficient Root Metabolism Improves Drought Resistance of Festuca arundinacea. Plant Cell Physiol. 2020, 61, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yu, L.; Xu, C. A central role for Triacylglycerol in Membrane Lipid Breakdown, Fatty Acid β-Oxidation, and Plant Survival under Extended Darkness. Plant Physiol. 2017, 174, 1517–1530. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Benning, C. Functions of Triacylglycerols during Plant Development and Stress. Curr. Opin. Biotechnol. 2018, 49, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, M.W.; Canter, P.J.; Thomas, H.M. Advances in Introgression Technologies for Precision Breeding within the Lolium—Festuca Complex. Ann. Appl. Biol. 2003, 143, 1–10. [Google Scholar] [CrossRef]
- Wang, S.; Li, H.; Lin, C. Physiological, Biochemical and Growth Responses of Italian Ryegrass to Butachlor Exposure. Pestic. Biochem. Physiol. 2013, 106, 21–27. [Google Scholar] [CrossRef]
- Cyriac, D.; Hofmann, R.W.; Stewart, A.; Sathish, P.; Winefield, C.S.; Moot, D.J. Intraspecific Differences in Long-Term Drought Tolerance in Perennial Ryegrass. PLoS ONE 2018, 13, e0194977. [Google Scholar] [CrossRef]
- Li, M.; Sheng, G.; Wu, Y.; Yu, Z.; Bañuelos, G.S.; Yu, H. Enhancement of Nitrogen and Phosphorus Removal from Eutrophic Water by Economic Plant Annual Ryegrass (Lolium multiflorum) with Ion Implantation. Environ. Sci. Pollut. Res. 2014, 21, 9617–9625. [Google Scholar] [CrossRef]
- Perlikowski, D.; Augustyniak, A.; Masajada, K.; Skirycz, A.; Soja, A.M.; Michaelis, Ä.; Wolter, G.; Kosmala, A. Structural and Metabolic Alterations in Root Systems under Limited Water Conditions in Forage Grasses of Lolium-Festuca Complex. Plant Sci. 2019, 283, 211–223. [Google Scholar] [CrossRef]
- Franca, A.; Loi, A.; Davies, W. Selection of Annual Ryegrass for Adaptation to Semi-Arid Conditions. Eur. J. Agron. 1998, 9, 71–78. [Google Scholar] [CrossRef]
- Humphreys, M.W.; Thomas, H. Improved Drought Resistance in Introgression Lines Derived from Lolium multiflorum x Festuca arundinacea Hybrids. Plant Breed. 1993, 111, 155–161. [Google Scholar] [CrossRef]
- Kemesyte, V.; Statkeviciute, G.; Brazauskas, G. Perennial Ryegrass Yield Performance under Abiotic Stress. Crop Sci. 2017, 57, 1935–1940. [Google Scholar] [CrossRef]
- Kosmala, A.; Perlikowski, D.; Pawłowicz, I.; Rapacz, M. Changes in the Chloroplast Proteome Following Water Deficit and Subsequent Watering in a High- and a Low-Drought-Tolerant Genotype of Festuca arundinacea. J. Exp. Bot. 2012, 63, 6161–6172. [Google Scholar] [CrossRef]
- Lechowicz, K.; Pawłowicz, I.; Perlikowski, D.; Arasimowicz-Jelonek, M.; Majka, J.; Augustyniak, A.; Rapacz, M.; Kosmala, A. Two Festuca species—F. arundinacea and F. glaucescens—Differ in the Molecular Response to Drought, while their Physiological Response is Similar. Int. J. Mol. Sci. 2020, 21, 3174. [Google Scholar] [CrossRef]
- Perlikowski, D.; Kosmala, A.; Rapacz, M.; Kościelniak, J.; Pawlowicz, I.; Zwierzykowski, Z. Influence of Short-Term Drought Conditions and Subsequent Re-Watering on the Physiology and Proteome of Lolium multiflorum/Festuca arundinacea Introgression Forms, with Contrasting Levels of Tolerance to Long-Term Drought. Plant Biol. 2014, 16, 385–394. [Google Scholar] [CrossRef]
- Perlikowski, D.; Czyżniejewski, M.; Marczak, Ł.; Augustyniak, A.; Kosmala, A. Water Deficit Affects Primary Metabolism Differently in Two Lolium multiflorum/Festuca arundinacea Introgression Forms with a Distinct Capacity for Photosynthesis and Membrane Regeneration. Front. Plant Sci. 2016, 7, 1–16. [Google Scholar] [CrossRef]
- Kosmala, A.; Zwierzykowski, Z.; Zwierzykowska, E.; Łuczak, M.; Rapacz, M.; Gasior, D.; Humphreys, M. Introgression Mapping of Genes For Winter Hardiness and Frost Tolerance Transferred from Festuca arundinacea into Lolium multiflorum. J. Hered. 2007, 98, 311–316. [Google Scholar] [CrossRef]
- Augustyniak, A.; Perlikowski, D.; Rapacz, M.; Kościelniak, J.; Kosmala, A. Insight into Cellular Proteome of Lolium multiflorum/Festuca arundinacea Introgression Forms to Decipher Crucial Mechanisms of Cold Acclimation in Forage Grasses. Plant Sci. 2018, 272, 22–31. [Google Scholar] [CrossRef]
- Płażek, A.; Pociecha, E.; Augustyniak, A.; Masajada, K.; Dziurka, M.; Majka, J.; Perlikowski, D.; Pawłowicz, I.; Kosmala, A. Dissection of Resistance to Microdochium nivale in Lolium multiflorum/Festuca arundinacea Introgression Forms. Plant Physiol. Biochem. 2018, 123, 43–53. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Y.; He, Q.; Li, H.; Zhang, X.; Zhang, F. Comparative Proteomics Illustrates the Complexity of Drought Resistance Mechanisms in Two Wheat (Triticum aestivum L.) Cultivars under Dehydration and Rehydration. BMC Plant Biol. 2016, 16, 188. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Meng, C.; Wang, J.; Ma, X.; Fan, X.; Yang, Z.; Zhou, M.; Zhang, X. Integrated Omics Data of Two Annual Ryegrass (Lolium multiflorum L.) Genotypes Reveals Core Metabolic Processes under Drought Stress. BMC Plant Biol. 2018, 18, 26. [Google Scholar] [CrossRef]
- Loggini, B.; Scartazza, A.; Brugnoli, E.; Navari-Izzo, F. Antioxidative Defense System, Pigment Composition, and Photosynthetic Efficiency in Two Wheat Cultivars Subjected to Drought. Plant Physiol. 1999, 113, 1091–1100. [Google Scholar] [CrossRef] [PubMed]
- Bian, S.; Jiang, Y. Reactive Oxygen Species, Antioxidant Enzyme Activities and Gene Expression Patterns in Leaves and Roots of Kentucky Bluegrass in Response to Drought Stress and Recovery. Sci. Hortic. (Amsterdam) 2009, 120, 264–270. [Google Scholar] [CrossRef]
- Shao, H.B.; Liang, Z.S.; Shao, M.A.; Wang, B.C. Changes of Anti-Oxidative Enzymes and Membrane Peroxidation for Soil Water Deficits among 10 Wheat Genotypes at Seedling Stage. Colloids Surf. B Biointerfaces 2005, 42, 107–113. [Google Scholar] [CrossRef]
- Ahmed, I.M.; Nadira, U.A.; Bibi, N.; Cao, F.; He, X.; Zhang, G.; Wu, F. Secondary Metabolism and Antioxidants are Involved in the Tolerance to Drought and Salinity, Separately and Combined, in Tibetan Wild Barley. Environ. Exp. Bot. 2015, 111, 1–12. [Google Scholar] [CrossRef]
- Beechey-Gradwell, Z.; Cooney, L.; Winichayakul, S.; Andrews, M.; Hea, S.Y.; Crowther, T.; Roberts, N. Storing Carbon in Leaf Lipid Sinks Enhances Perennial Ryegrass Carbon Capture Especially under High N and Elevated CO2. J. Exp. Bot. 2020, 71, 2351–2361. [Google Scholar] [CrossRef]
- Paul, M.J.; Eastmond, P.J. Turning Sugar into Oil: Making Photosynthesis Blind to Feedback Inhibition. J. Exp. Bot. 2020, 71, 2216–2218. [Google Scholar] [CrossRef]
- Doheny-Adams, T.; Hunt, L.; Franks, P.J.; Beerling, D.J.; Gray, J.E. Genetic Manipulation of Stomatal Density Influences Stomatal Size, Plant Growth and Tolerance to Restricted Water Supply across a Growth Carbon Dioxide Gradient. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 547–555. [Google Scholar] [CrossRef]
- Hughes, J.; Hepworth, C.; Dutton, C.; Dunn, J.A.; Hunt, L.; Stephens, J.; Waugh, R.; Cameron, D.D.; Gray, J.E. Reducing Stomatal Density in Barley Improves Drought Tolerance without Impacting on Yield. Plant Physiol. 2017, 174, 776–787. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with Reduced Stomatal Density Conserves Water and Has Improved Drought Tolerance under Future Climate Conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Raissig, M.T.; Matos, J.L.; Gil, M.X.A.; Kornfeld, A.; Bettadapur, A.; Abrash, E.; Allison, H.R.; Badgley, G.; Vogel, J.P.; Berry, J.A.; et al. Mobile MUTE Specifies Subsidiary Cells to Build Physiologically Improved Grass Stomata. Science 2017, 355, 1215–1218. [Google Scholar] [CrossRef] [PubMed]
- Rapacz, M.; Wójcik-Jagła, M.; Fiust, A.; Kalaji, H.M.; Koscielniak, J. Genome-Wide Associations of Chlorophyll Fluorescence Ojip Transient Parameters Connected with Soil Drought Response in Barley. Front. Plant Sci. 2019, 10, 78. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Dong, K.; Ge, P.; Bian, Y.; Dong, L.; Deng, X.; Li, X.; Yan, Y. Identification of Leaf Proteins Differentially Accumulated between Wheat Cultivars Distinct in Their Levels of Drought Tolerance. PLoS ONE 2015, 10, e0125302. [Google Scholar] [CrossRef]
- Das, A.; Eldakak, M.; Paudel, B.; Kim, D.-W.; Hemmati, H.; Basu, C.; Rohila, J.S. Leaf Proteome Analysis Reveals Prospective Drought and Heat Stress Response Mechanisms in Soybean. Biomed. Res. Int. 2016, 2016, 6021047. [Google Scholar] [CrossRef]
- Liu, D.; Li, W.; Cheng, J.; Hou, L. AtPGK2, a Member of PGKs Gene Family in Arabidopsis, has a Positive Role in Salt Stress Tolerance. Plant Cell. Tissue Organ Cult. 2015, 120, 251–262. [Google Scholar] [CrossRef]
- Joshi, R.; Karan, R.; Singla-Pareek, S.L.; Pareek, A. Ectopic Expression of Pokkali Phosphoglycerate Kinase-2 (OsPGK2-P) Improves Yield in Tobacco Plants under Salinity Stress. Plant Cell Rep. 2016, 35, 27–41. [Google Scholar] [CrossRef]
- Wang, X.; Du, T.; Huang, J.; Peng, S.; Xiong, D. Leaf Hydraulic Vulnerability Triggers the Decline in Stomatal and Mesophyll Conductance During Drought in Rice. J. Exp. Bot. 2018, 69, 4033–4045. [Google Scholar] [CrossRef]
- Li, W.; Zhang, S.; Shan, L. Responsibility of Non-Stomatal Limitations for the Reduction of Photosynthesis-Response of Photosynthesis and Antioxidant Enzyme Characteristics in Alfalfa (Medicago sativa L.) Seedlings to Water Stress and Rehydration. Front. Agric. China 2007, 1, 255–264. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive Oxygen Species Homeostasis and Signalling During Drought and Salinity Stresses. Plant, Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, J. Water Stress-Induced Abscisic Acid Accumulation Triggers the Increased Generation of Reactive Oxygen Species and Up-Regulates the Activities of Antioxidant Enzymes in Maize Leaves. J. Exp. Bot. 2002, 53, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Luna, C.M. Drought Controls on H2O2 Accumulation, Catalase (CAT) Activity and CAT Gene Expression in Wheat. J. Exp. Bot. 2004, 56, 417–423. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS Are Good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Huseynova, I.M.; Aliyeva, D.R.; Aliyev, J.A. Subcellular Localization and Responses of Superoxide Dismutase Isoforms in Local Wheat Varieties Subjected to Continuous Soil Drought. Plant Physiol. Biochem. 2014, 81, 54–60. [Google Scholar] [CrossRef]
- Leonowicz, G.; Trzebuniak, K.F.; Zimak-Piekarczyk, P.; Ślesak, I.; Mysliwa-Kurdziel, B. The activity of Superoxide Dismutases (SODs) at the Early Stages of Wheat Deetiolation. PLoS ONE 2018, 13, e0194678. [Google Scholar] [CrossRef]
- Guo, Z.; Ou, W.; Lu, S.; Zhong, Q. Differential Responses of Antioxidative System to Chilling and Drought in Four Rice Cultivars Differing in Sensitivity. Plant Physiol. Biochem. 2006, 44, 828–836. [Google Scholar] [CrossRef]
- Rubio, M.C.; Gonzalez, E.M.; Minchin, F.R.; Webb, K.J.; Arrese-Igor, C.; Ramos, J.; Becana, M. Effects of Water Stress on Antioxidant Enzymes of Leaves and Nodules of Transgenic Alfalfa Overexpressing Superoxide Dismutases. Physiol. Plant. 2002, 115, 531–540. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive Oxygen Gene Network of Plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.J. The Role of the Plant Antioxidant System in Drought Tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef]
- Fan, J.; Yan, C.; Xu, C. Phospholipid: Diacylglycerol Acyltransferase- Mediated Triacylglycerol Biosynthesis is Crucial for Protection against Fatty Acid-Induced Cell Death in Growing Tissues of Arabidopsis. Plant J. 2013, 76, 930–942. [Google Scholar] [CrossRef]
- Gasulla, F.; Vom Dorp, K.; Dombrink, I.; Zähringer, U.; Gisch, N.; Dörmann, P.; Bartels, D. The Role of Lipid Metabolism in the Acquisition of Desiccation Tolerance in Craterostigma plantagineum: A Comparative Approach. Plant J. 2013, 75, 726–741. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yan, C.; Roston, R.; Shanklin, J.; Xu, C. Arabidopsis lipins, PDAT1 Acyltransferase, and SDP1 Triacylglycerol Lipase Synergistically Direct Fatty Acids toward Β-Oxidation, Thereby Maintaining Membrane Lipid Homeostasis. Plant Cell 2014, 26, 4119–4134. [Google Scholar] [CrossRef] [PubMed]
- Chaffai, R.; Cherif, A. The cadmium-induced changes in the polar and neutral lipid compositions suggest the involvement of triacylglycerol in the defense response in maize. Physiol. Mol. Biol. Plants 2020, 26, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Flexas, J. Genetic Improvement of Leaf Photosynthesis and Intrinsic Water Use Efficiency in C3 Plants: Why So Much Little Success? Plant Sci. 2016, 251, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Hurkman, W.J.; Tanaka, C.K. Solubilization of Plant Membrane Proteins for Analysis by Two-Dimensional Gel Electrophoresis. Plant Physiol. 1986, 81, 802–806. [Google Scholar] [CrossRef]
- Pawłowicz, I.; Kosmala, A.; Rapacz, M. Expression Pattern of the psbO Gene and its Involvement in Acclimation of the Photosynthetic Apparatus during Abiotic Stresses in Festuca arundinacea and F. pratensis. Acta Physiol. Plant. 2012, 34, 1915–1924. [Google Scholar] [CrossRef]
- Sibley, J.A.; Lehninger, A.L. Determination of Aldolase in Animal Tissues. J. Biol. Chem. 1949, 177, 859–872. [Google Scholar]
- Willard, J.M.; Gibbs, M. Role of Aldolase in Photosynthesis. II Demonstration of Aldolase Types in Photosynthetic Organisms. Plant Physiol. 1968, 43, 793–798. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf Senescence: Correlated with Increased Levels of Membrane Permeability and Lipid Peroxidation, and Decreased Levels of Superoxide Dismutase and Catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in Isolated Chloroplasts. I. Kinetics and Stoichiometry of Fatty Acid Peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Doke, N. Involvement of Superoxide Anion Generation in The Hypersensitive Response of Potato Tuber Tissues to Infection with an Incompatible Race of Phytophthora infestans and to the Hyphal Wall Components. Physiol. Plant Pathol. 1983, 23, 345–357. [Google Scholar] [CrossRef]
- Arasimowicz, M.; Floryszak-Wieczorek, J.; Milczarek, G.; Jelonek, T. Nitric Oxide, Induced by Wounding, Mediates Redox Regulation in Pelargonium Leaves. Plant Biol. 2009, 11, 650–663. [Google Scholar] [CrossRef] [PubMed]
- Becana, M.; Aparicio-Tejo, P.; Jose Irigoyen, J.; Sanchez-Diaz, M. Some Enzymes of Hydrogen Peroxide Metabolism in Leaves and Root Nodules of Medicago sativa. Plant Physiol. 1986, 82, 1169–1171. [Google Scholar] [CrossRef] [PubMed]
- Arasimowicz-Jelonek, M.; Floryszak-Wieczorek, J.; Gzyl, J.; Chmielowska-Bak, J. Homocysteine Over-Accumulation as the Effect of Potato Leaves Exposure to Biotic Stress. Plant Physiol. Biochem. 2013, 63, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Giavalisco, P.; Li, Y.; Matthes, A.; Eckhardt, A.; Hubberten, H.M.; Hesse, H.; Segu, S.; Hummel, J.; Köhl, K.; Willmitzer, L. Elemental Formula Annotation of Polar and Lipophilic Metabolites Using 13C, 15N and 34S Isotope Labelling, in Combination with High-Resolution Mass Spectrometry. Plant J. 2011, 68, 364–376. [Google Scholar] [CrossRef]
- Bromke, M.A.; Hochmuth, A.; Tohge, T.; Fernie, A.R.; Giavalisco, P.; Burgos, A.; Willmitzer, L.; Brotman, Y. Liquid Chromatography High-Resolution Mass Spectrometry for Fatty acid Profiling. Plant J. 2015, 81, 529–536. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Forward/Reverse | TaqMan Probe | Amplicon Length (bp) |
|---|---|---|---|
| Actin | GTCGAGGGCAACATATGCAA CCAGTGCTGAGCGGGAAT | TTCTCCTTGATGTCACGGAC | 65 |
| Ubiquitin | GCAAGAAGAAGACGTACA GACCTTGTAGAACTGGAG | CTTCACCTTCTTGTCCTTGTGCTT | 86 |
| pFBA | GAGACGTTCTACTACATG GAGGAGCTTGAGAGTGTA | TGTTCCTGTCCTTGCACTCGG | 140 |
| pPGK | CCTTGGTTGAGGAAGATAA CAGCAATGACAACATCAG | CTGGCAACAACTCTCCTGGC | 102 |
| GR | GGGGAGTACGACTACGACCT TCGTAAGTCCACCCAAAGCC | GGCGGCGTCAGGGCCTCGCGCTT | 254 |
| GPX | TCACTCGGCGGCCTGGAGAA TTCACAGTGCGGGCTTACGA | CTACGCCACCGCCGCCACGGAGAA | 212 |
| APX | CTCGTATCGCAGGAGCTCG TTGGGCCACTCGCTAATGTT | CGGCTGCGGCTGGAGATGCGACGGC | 188 |
| Fe-SOD | TCTATCTCGGCGGTTCTCCA CCGTTGTTGTAGGCCTCCTT | GCTCGACACCAGCCCCTTCTACGGCCA | 219 |
| Cu/Zn-SOD | CCAGAGCATCCTCTTCGCC ATTGATGGAGGTGGAAGCCG | TCGCTCCGCCTCGTCTCCGCCCCC | 287 |
| Mn-SOD | TTGACGCCGCTGTCTCTAAGG TTTATCCAACGCCAGCCACA | GCTTCCGCCGTCGTCCAACTCCAGGGC | 266 |
| CAT | GTTCACCTTCCTCTTCGACG AAGTCGAACCTGTCCTCGTG | ACTACCGCCACATGGATGGCTCCG | 297 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lechowicz, K.; Pawłowicz, I.; Perlikowski, D.; Arasimowicz-Jelonek, M.; Blicharz, S.; Skirycz, A.; Augustyniak, A.; Malinowski, R.; Rapacz, M.; Kosmala, A. Adjustment of Photosynthetic and Antioxidant Activities to Water Deficit Is Crucial in the Drought Tolerance of Lolium multiflorum/Festuca arundinacea Introgression Forms. Int. J. Mol. Sci. 2020, 21, 5639. https://doi.org/10.3390/ijms21165639
Lechowicz K, Pawłowicz I, Perlikowski D, Arasimowicz-Jelonek M, Blicharz S, Skirycz A, Augustyniak A, Malinowski R, Rapacz M, Kosmala A. Adjustment of Photosynthetic and Antioxidant Activities to Water Deficit Is Crucial in the Drought Tolerance of Lolium multiflorum/Festuca arundinacea Introgression Forms. International Journal of Molecular Sciences. 2020; 21(16):5639. https://doi.org/10.3390/ijms21165639
Chicago/Turabian StyleLechowicz, Katarzyna, Izabela Pawłowicz, Dawid Perlikowski, Magdalena Arasimowicz-Jelonek, Sara Blicharz, Aleksandra Skirycz, Adam Augustyniak, Robert Malinowski, Marcin Rapacz, and Arkadiusz Kosmala. 2020. "Adjustment of Photosynthetic and Antioxidant Activities to Water Deficit Is Crucial in the Drought Tolerance of Lolium multiflorum/Festuca arundinacea Introgression Forms" International Journal of Molecular Sciences 21, no. 16: 5639. https://doi.org/10.3390/ijms21165639
APA StyleLechowicz, K., Pawłowicz, I., Perlikowski, D., Arasimowicz-Jelonek, M., Blicharz, S., Skirycz, A., Augustyniak, A., Malinowski, R., Rapacz, M., & Kosmala, A. (2020). Adjustment of Photosynthetic and Antioxidant Activities to Water Deficit Is Crucial in the Drought Tolerance of Lolium multiflorum/Festuca arundinacea Introgression Forms. International Journal of Molecular Sciences, 21(16), 5639. https://doi.org/10.3390/ijms21165639