Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability


Abstract
1. Introduction
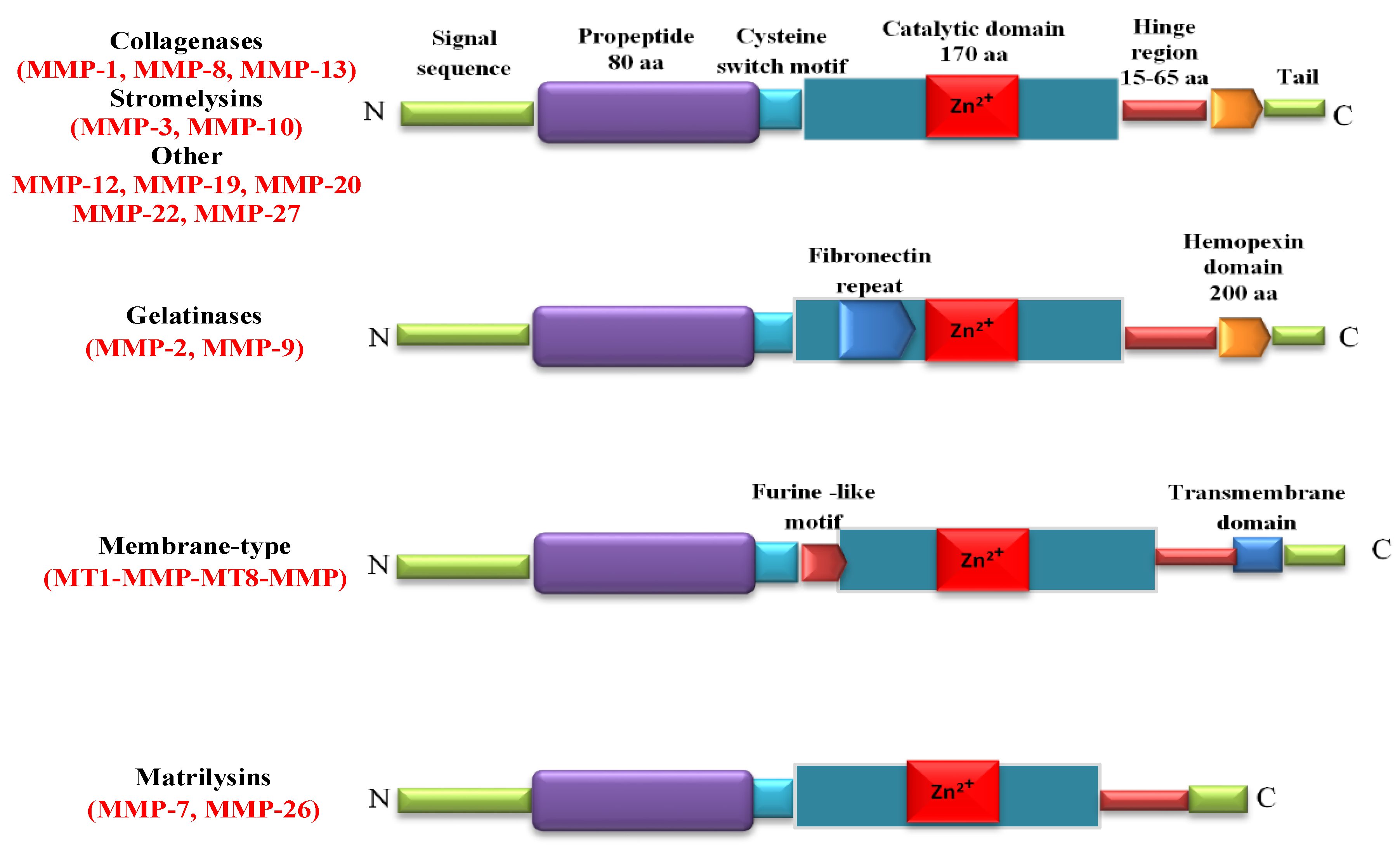
2. MMPs Structure and Tissue Distribution
3. Extracellular Vesicles as MMPs Carriers
4. Biological Inhibitors of MMPs
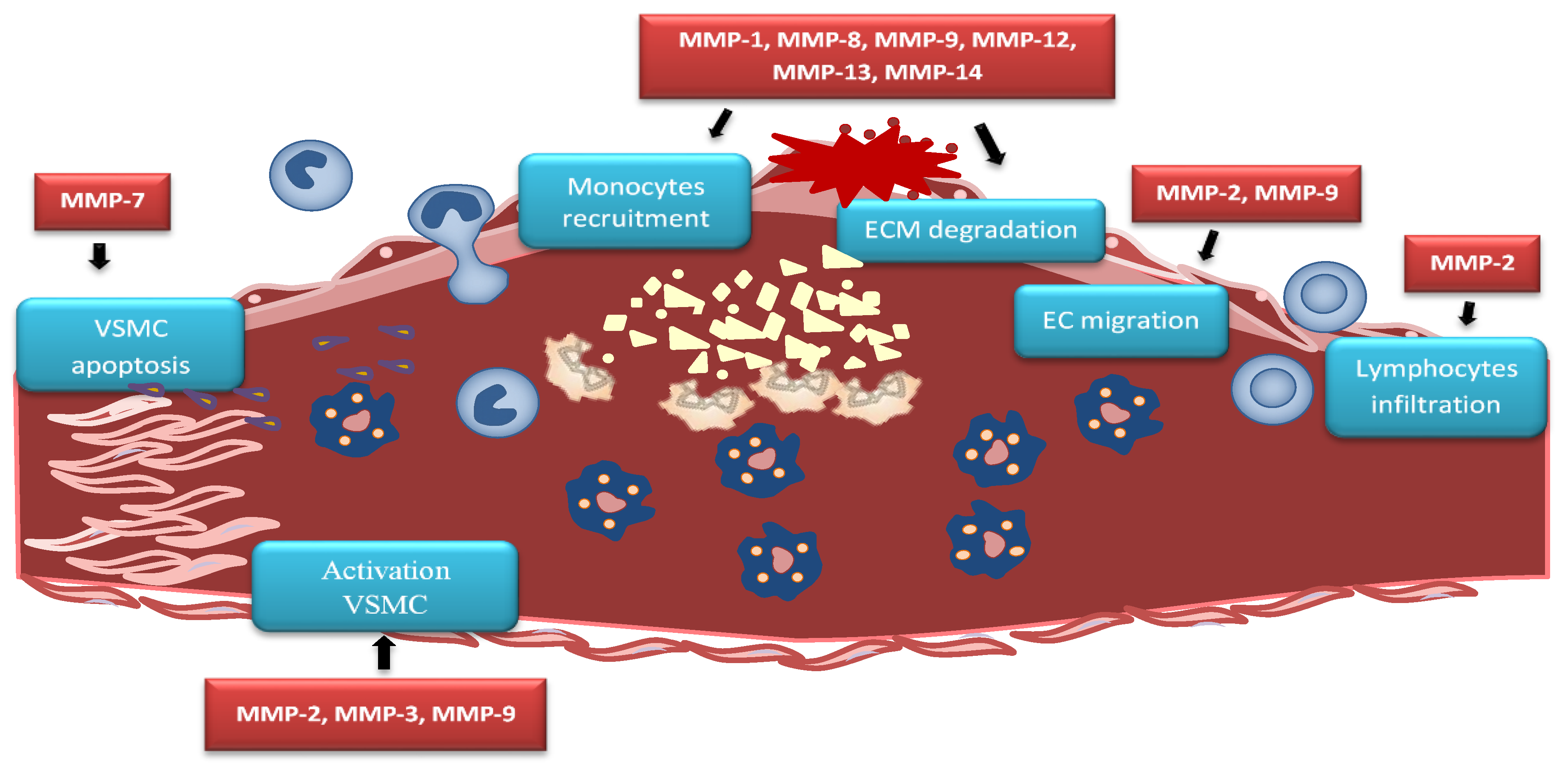
5. MMPs in Pathogenesis of Atherosclerosis
5.1. MMPs in Vascular Inflammation and Recruitment of Immune Cells
5.2. MMPs in Lipid Accumulation
5.3. MMPs in Endothelial Dysfunction
5.4. MMPs in Migration of VSMCs
5.5. MMPs in Plaque Neovascularization
5.6. MMPs in Plaque Calcification
5.7. MMPs in Plaque Activation and Destabilization
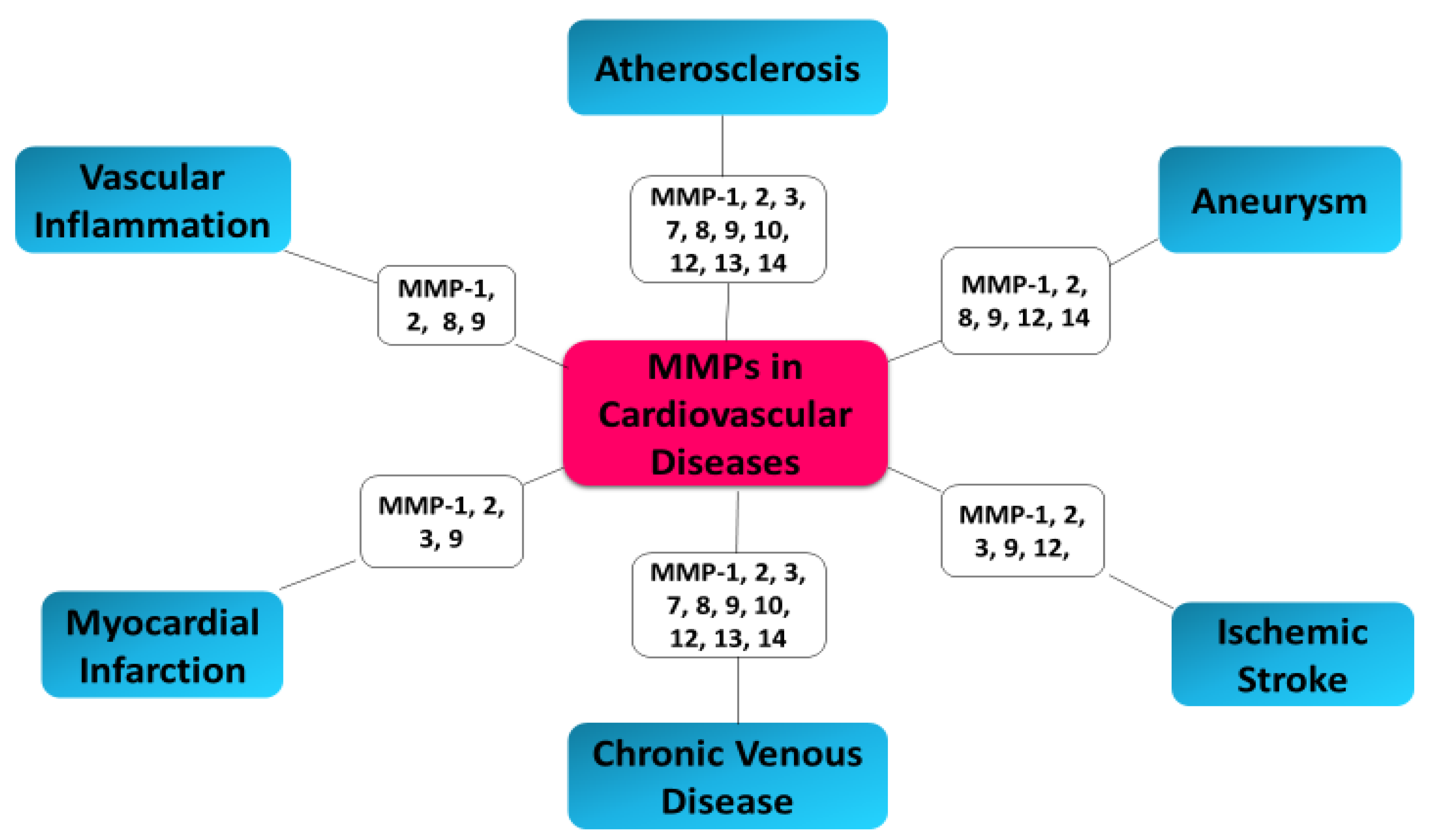
6. MMPs as Biomarkers in Cardiovascular Diseases
7. Conclusions
Funding
Conflicts of Interest
References
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and timps. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef]
- MacColl, E.; Khalil, R.A. Matrix metalloproteinases as regulators of vein structure and function: Implications in chronic venous disease. J. Pharmacol. Exp. Ther 2015, 355, 410–428. [Google Scholar] [CrossRef]
- Marino-Puertas, L.; Goulas, T.; Gomis-Ruth, F.X. Matrix metalloproteinases outside vertebrates. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2026–2035. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.P. Matrix metalloproteinase inhibitors: Specificity of binding and structure-activity relationships. Exp. Suppl. 2012, 103, v–vi. [Google Scholar] [PubMed]
- Chen, Q.; Jin, M.; Yang, F.; Zhu, J.; Xiao, Q.; Zhang, L. Matrix metalloproteinases: Inflammatory regulators of cell behaviors in vascular formation and remodeling. Mediators Inflamm. 2013, 2013, 928315. [Google Scholar] [CrossRef]
- Qiang, B.; Toma, J.; Fujii, H.; Osherov, A.B.; Nili, N.; Sparkes, J.D.; Fefer, P.; Samuel, M.; Butany, J.; Leong-Poi, H.; et al. Statin therapy prevents expansive remodeling in venous bypass grafts. Atherosclerosis 2012, 223, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Loftus, I.M.; Naylor, A.R.; Bell, P.R.; Thompson, M.M. Matrix metalloproteinases and atherosclerotic plaque instability. Br. J. Surg. 2002, 89, 680–694. [Google Scholar] [CrossRef]
- Myasoedova, V.A.; Chistiakov, D.A.; Grechko, A.V.; Orekhov, A.N. Matrix metalloproteinases in pro-atherosclerotic arterial remodeling. J. Mol. Cell Cardiol. 2018, 123, 159–167. [Google Scholar] [CrossRef]
- Serra, R.; Gallelli, L.; Butrico, L.; Buffone, G.; Calio, F.G.; De Caridi, G.; Massara, M.; Barbetta, A.; Amato, B.; Labonia, M.; et al. From varices to venous ulceration: The story of chronic venous disease described by metalloproteinases. Int. Wound J. 2017, 14, 233–240. [Google Scholar] [CrossRef]
- Volkov, A.M.; Murashov, I.S.; Polonskaya, Y.V.; Savchenko, S.V.; Kazanskaya, G.M.; Kliver, E.E.; Ragino, Y.I.; AM, C.H. Changes of content of matrix metalloproteinases and their tissue expression in various types of atherosclerotic plaques. Kardiologiia 2018, 10, 12–18. [Google Scholar] [CrossRef]
- Abbas, A.; Aukrust, P.; Russell, D.; Krohg-Sorensen, K.; Almas, T.; Bundgaard, D.; Bjerkeli, V.; Sagen, E.L.; Michelsen, A.E.; Dahl, T.B.; et al. Matrix metalloproteinase 7 is associated with symptomatic lesions and adverse events in patients with carotid atherosclerosis. PLoS ONE 2014, 9, e84935. [Google Scholar] [CrossRef] [PubMed]
- Ezhov, M.; Safarova, M.; Afanasieva, O.; Mitroshkin, M.; Matchin, Y.; Pokrovsky, S. Matrix metalloproteinase 9 as a predictor of coronary atherosclerotic plaque instability in stable coronary heart disease patients with elevated lipoprotein(a) levels. Biomolecules 2019, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Lahdentausta, L.; Leskela, J.; Winkelmann, A.; Tervahartiala, T.; Sorsa, T.; Pesonen, E.; Pussinen, P.J. Serum MMP-9 diagnostics, prognostics, and activation in acute coronary syndrome and its recurrence. J. Cardiovasc. Transl. Res. 2018, 11, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Liu, Y.; Li, W.; Deng, F.; Liu, X.; Wang, X.; Gui, Y.; Qin, L.; Hu, C.; Chen, L. Associations of matrix metalloproteinase-9 and monocyte chemoattractant protein-1 concentrations with carotid atherosclerosis, based on measurements of plaque and intima-media thickness. Atherosclerosis 2014, 232, 199–203. [Google Scholar] [CrossRef]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix metalloproteinase-9: Many shades of function in cardiovascular disease. Physiology 2013, 28, 391–403. [Google Scholar] [CrossRef]
- Brown, B.A.; Williams, H.; George, S.J. Evidence for the involvement of matrix-degrading metalloproteinases (MMPs) in atherosclerosis. Prog. Mol. Biol. Transl. Sci. 2017, 147, 197–237. [Google Scholar]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and biological attributes of matrix metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar]
- Wang, J.; Tan, G.J.; Han, L.N.; Bai, Y.Y.; He, M.; Liu, H.B. Novel biomarkers for cardiovascular risk prediction. J. Geriatr. Cardiol. 2017, 14, 135–150. [Google Scholar]
- Cauwe, B.; Van den Steen, P.E.; Opdenakker, G. The biochemical, biological, and pathological kaleidoscope of cell surface substrates processed by matrix metalloproteinases. Crit. Rev. Biochem. Mol. Biol. 2007, 42, 113–185. [Google Scholar] [CrossRef]
- Amin, M.; Pushpakumar, S.; Muradashvili, N.; Kundu, S.; Tyagi, S.C.; Sen, U. Regulation and involvement of matrix metalloproteinases in vascular diseases. Front. Biosci. 2016, 21, 89–118. [Google Scholar]
- Sansilvestri-Morel, P.; Fioretti, F.; Rupin, A.; Senni, K.; Fabiani, J.N.; Godeau, G.; Verbeuren, T.J. Comparison of extracellular matrix in skin and saphenous veins from patients with varicose veins: Does the skin reflect venous matrix changes? Clin. Sci. 2007, 112, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Pradhan-Palikhe, P.; Vikatmaa, P.; Lajunen, T.; Palikhe, A.; Lepantalo, M.; Tervahartiala, T.; Salo, T.; Saikku, P.; Leinonen, M.; Pussinen, P.J.; et al. Elevated MMP-8 and decreased myeloperoxidase concentrations associate significantly with the risk for peripheral atherosclerosis disease and abdominal aortic aneurysm. Scand. J. Immunol. 2010, 72, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Seizer, P.; May, A.E. Platelets and matrix metalloproteinases. Thromb. Haemost. 2013, 110, 903–909. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Tang, T.; Hou, Y.; Zeng, Q.; Wang, Y.; Fan, W.; Qu, S. Extracellular vesicles in atherosclerosis. Clin. Chim. Acta 2019, 495, 109–117. [Google Scholar] [CrossRef]
- Sluijter, J.P.G.; Davidson, S.M.; Boulanger, C.M.; Buzas, E.I.; de Kleijn, D.P.V.; Engel, F.B.; Giricz, Z.; Hausenloy, D.J.; Kishore, R.; Lecour, S.; et al. Extracellular vesicles in diagnostics and therapy of the ischaemic heart: Position paper from the working group on cellular biology of the heart of the european society of cardiology. Cardiovasc. Res. 2018, 114, 19–34. [Google Scholar] [CrossRef]
- van Niel, G.; D'Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Sinha, S.; Hoshino, D.; Hong, N.H.; Kirkbride, K.C.; Grega-Larson, N.E.; Seiki, M.; Tyska, M.J.; Weaver, A.M. Cortactin promotes exosome secretion by controlling branched actin dynamics. J. Cell Biol. 2016, 214, 197–213. [Google Scholar] [CrossRef]
- Clark, E.S.; Whigham, A.S.; Yarbrough, W.G.; Weaver, A.M. Cortactin is an essential regulator of matrix metalloproteinase secretion and extracellular matrix degradation in invadopodia. Cancer Res. 2007, 67, 4227–4235. [Google Scholar] [CrossRef]
- Wiesner, C.; El Azzouzi, K.; Linder, S. A specific subset of rabgtpases controls cell surface exposure of MT1-MMP, extracellular matrix degradation and three-dimensional invasion of macrophages. J. Cell Sci. 2013, 126, 2820–2833. [Google Scholar] [CrossRef]
- Bachmann, A.; Straube, A. Kinesins in cell migration. Biochem. Soc. Trans. 2015, 43, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Clancy, J.W.; Sedgwick, A.; Rosse, C.; Muralidharan-Chari, V.; Raposo, G.; Method, M.; Chavrier, P.; D'Souza-Schorey, C. Regulated delivery of molecular cargo to invasive tumour-derived microvesicles. Nat. Commun. 2015, 6, 6919. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.; Shah, N.; Zanetti, B.R.; Maugeri, M.; Silvestre, R.N.; Fatima, F.; Neder, L.; Valadi, H. Extracellular vesicles and matrix remodeling enzymes: The emerging roles in extracellular matrix remodeling, progression of diseases and tissue repair. Cells 2018, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Bobryshev, Y.V.; Killingsworth, M.C.; Orekhov, A.N. Increased shedding of microvesicles from intimal smooth muscle cells in athero-prone areas of the human aorta: Implications for understanding of the predisease stage. Pathobiology 2013, 80, 24–31. [Google Scholar] [CrossRef]
- Taraboletti, G.; D'Ascenzo, S.; Borsotti, P.; Giavazzi, R.; Pavan, A.; Dolo, V. Shedding of the matrix metalloproteinases mmp-2, mmp-9, and mt1-mmp as membrane vesicle-associated components by endothelial cells. Am. J. Pathol 2002, 160, 673–680. [Google Scholar] [CrossRef]
- Lozito, T.P.; Tuan, R.S. Endothelial cell microparticles act as centers of matrix metalloproteinsase-2 (MMP-2) activation and vascular matrix remodeling. J. Cell Physiol. 2012, 227, 534–549. [Google Scholar] [CrossRef]
- Laghezza Masci, V.; Taddei, A.R.; Gambellini, G.; Giorgi, F.; Fausto, A.M. Microvesicles shed from fibroblasts act as metalloproteinase carriers in a 3-d collagen matrix. J. Circ. Biomark 2016, 5, 1849454416663660. [Google Scholar] [CrossRef]
- Moreno, P.R.; Purushothaman, K.R.; Fuster, V.; Echeverri, D.; Truszczynska, H.; Sharma, S.K.; Badimon, J.J.; O'Connor, W.N. Plaque neovascularization is increased in ruptured atherosclerotic lesions of human aorta: Implications for plaque vulnerability. Circulation 2004, 110, 2032–2038. [Google Scholar] [CrossRef]
- Shimoda, M.; Khokha, R. Metalloproteinases in extracellular vesicles. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 1989–2000. [Google Scholar] [CrossRef]
- Cheng, X.W.; Song, H.; Sasaki, T.; Hu, L.; Inoue, A.; Bando, Y.K.; Shi, G.P.; Kuzuya, M.; Okumura, K.; Murohara, T. Angiotensin type 1 receptor blocker reduces intimal neovascularization and plaque growth in apolipoprotein e-deficient mice. Hypertension 2011, 57, 981–989. [Google Scholar] [CrossRef]
- Badimon, L.; Suades, R.; Arderiu, G.; Pena, E.; Chiva-Blanch, G.; Padro, T. Microvesicles in atherosclerosis and angiogenesis: From bench to bedside and reverse. Front. Cardiovasc. Med. 2017, 4, 77. [Google Scholar] [CrossRef] [PubMed]
- Bakhshian Nik, A.; Hutcheson, J.D.; Aikawa, E. Extracellular vesicles as mediators of cardiovascular calcification. Front. Cardiovasc. Med. 2017, 4, 78. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Gao, W.; Yao, K.; Ge, J. Roles of exosomes derived from immune cells in cardiovascular diseases. Front. Immunol 2019, 10, 648. [Google Scholar] [CrossRef] [PubMed]
- Bailey, G.; Meadows, J.; Morrison, A.R. Imaging atherosclerotic plaque calcification: Translating biology. Curr. Atheroscler Rep. 2016, 18, 51. [Google Scholar] [CrossRef]
- Kapustin, A.N.; Shanahan, C.M. Emerging roles for vascular smooth muscle cell exosomes in calcification and coagulation. J. Physiol. 2016, 594, 2905–2914. [Google Scholar] [CrossRef]
- Lu, X. The role of exosomes and exosome-derived micrornas in atherosclerosis. Curr. Pharm. Des. 2017, 23, 6182–6193. [Google Scholar] [CrossRef]
- Alique, M.; Ramirez-Carracedo, R.; Bodega, G.; Carracedo, J.; Ramirez, R. Senescent microvesicles: A novel advance in molecular mechanisms of atherosclerotic calcification. Int. J. Mol. Sci. 2018, 19, 2003. [Google Scholar] [CrossRef]
- Newby, A.C. Matrix metalloproteinase inhibition therapy for vascular diseases. Vascul Pharmacol. 2012, 56, 232–244. [Google Scholar] [CrossRef]
- Cerisano, G.; Buonamici, P.; Valenti, R.; Sciagra, R.; Raspanti, S.; Santini, A.; Carrabba, N.; Dovellini, E.V.; Romito, R.; Pupi, A.; et al. Early short-term doxycycline therapy in patients with acute myocardial infarction and left ventricular dysfunction to prevent the ominous progression to adverse remodelling: The tiptop trial. Eur. Heart J. 2014, 35, 184–191. [Google Scholar] [CrossRef]
- Schulze, C.J.; Castro, M.M.; Kandasamy, A.D.; Cena, J.; Bryden, C.; Wang, S.H.; Koshal, A.; Tsuyuki, R.T.; Finegan, B.A.; Schulz, R. Doxycycline reduces cardiac matrix metalloproteinase-2 activity but does not ameliorate myocardial dysfunction during reperfusion in coronary artery bypass patients undergoing cardiopulmonary bypass. Crit. Care Med. 2013, 41, 2512–2520. [Google Scholar] [CrossRef]
- Bench, T.J.; Jeremias, A.; Brown, D.L. Matrix metalloproteinase inhibition with tetracyclines for the treatment of coronary artery disease. Pharmacol. Res. 2011, 64, 561–566. [Google Scholar] [CrossRef]
- Rodriguez-Granillo, G.A.; Rodriguez-Granillo, A.; Milei, J. Effect of doxycycline on atherosclerosis: From bench to bedside. Recent Pat. Cardiovasc. Drug Discov 2011, 6, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Kormi, I.; Alfakry, H.; Tervahartiala, T.; Pussinen, P.J.; Sinisalo, J.; Sorsa, T. The effect of prolonged systemic doxycycline therapy on serum tissue degrading proteinases in coronary bypass patients: A randomized, double-masked, placebo-controlled clinical trial. Inflamm Res. 2014, 63, 329–334. [Google Scholar] [CrossRef]
- Rival, Y.; Beneteau, N.; Chapuis, V.; Taillandier, T.; Lestienne, F.; Dupont-Passelaigue, E.; Patoiseau, J.F.; Colpaert, F.C.; Junquero, D. Cardiovascular drugs inhibit MMP-9 activity from human THP-1 macrophages. DNA Cell Biol. 2004, 23, 283–292. [Google Scholar] [CrossRef]
- Cevik, C.; Otahbachi, M.; Nugent, K.; Warangkana, C.; Meyerrose, G. Effect of 3-hydroxy-3-methylglutaryl coenzyme a reductase inhibition on serum matrix metalloproteinase-13 and tissue inhibitor matrix metalloproteinase-1 levels as a sign of plaque stabilization. J. Cardiovasc. Med. (Hagerstown) 2008, 9, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Dorecka, M.; Francuz, T.; Garczorz, W.; Siemianowicz, K.; Romaniuk, W. The influence of elastin degradation products, glucose and atorvastatin on metalloproteinase-1, -2, -9 and tissue inhibitor of metalloproteinases-1, -2, -3 expression in human retinal pigment epithelial cells. Acta Biochim. Pol. 2014, 61, 265–270. [Google Scholar] [CrossRef]
- Luan, Z.; Chase, A.J.; Newby, A.C. Statins inhibit secretion of metalloproteinases-1, -2, -3, and -9 from vascular smooth muscle cells and macrophages. Arterioscler Thromb. Vasc Biol. 2003, 23, 769–775. [Google Scholar] [CrossRef] [PubMed]
- Komukai, K.; Kubo, T.; Kitabata, H.; Matsuo, Y.; Ozaki, Y.; Takarada, S.; Okumoto, Y.; Shiono, Y.; Orii, M.; Shimamura, K.; et al. Effect of atorvastatin therapy on fibrous cap thickness in coronary atherosclerotic plaque as assessed by optical coherence tomography: The easy-fit study. J. Am. Coll. Cardiol. 2014, 64, 2207–2217. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.L.; Yang, Y.; Yang, S.L.; Luo, J.P.; Li, H.; Jing, L.M.; Shen, Z.Q. Administration of a loading dose of atorvastatin before percutaneous coronary intervention prevents inflammation and reduces myocardial injury in stemi patients: A randomized clinical study. Clin. Ther. 2013, 35, 261–272. [Google Scholar] [CrossRef]
- Li, D.D.; Pang, H.G.; Song, J.N.; Huang, H.; Zhang, M.; Zhao, Y.L.; Sun, P.; Zhang, B.F.; Ma, X.D. The rapid lipopolysaccharide-induced release of matrix metalloproteinases 9 is suppressed by simvastatin. Cell Biol. Int. 2015, 39, 788–798. [Google Scholar] [CrossRef]
- Ichihara, S.; Noda, A.; Nagata, K.; Obata, K.; Xu, J.; Ichihara, G.; Oikawa, S.; Kawanishi, S.; Yamada, Y.; Yokota, M. Pravastatin increases survival and suppresses an increase in myocardial matrix metalloproteinase activity in a rat model of heart failure. Cardiovasc. Res. 2006, 69, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Sun, X.; He, Z.; Jiang, Y.; Zhang, X. Role of matrix metalloproteinase-9 in apoptosis of hippocampal neurons in rats during early brain injury after subarachnoid hemorrhage. Neurol. Sci. 2010, 31, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X.; Li, L.; Yao, H.; Jiang, Y.; Zhang, J. Effects of combination of ezetimibe and rosuvastatin on coronary artery plaque in patients with coronary heart disease. Heart Lung Circ. 2016, 25, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Sapienza, P.; Mingoli, A.; Borrelli, V.; Brachini, G.; Biacchi, D.; Sterpetti, A.V.; Grande, R.; Serra, R.; Tartaglia, E. Inflammatory biomarkers, vascular procedures of lower limbs, and wound healing. Int. Wound J. 2019, 16, 716–723. [Google Scholar] [CrossRef]
- Sapienza, P.; Mingoli, A.; Borrelli, V.; Grande, R.; Sterpetti, A.V.; Biacchi, D.; Ferrer, C.; Rubino, P.; Serra, R.; Tartaglia, E. Different inflammatory cytokines release after open and endovascular reconstructions influences wound healing. Int. Wound J. 2019, 16, 1034–1044. [Google Scholar] [CrossRef]
- Wolf, D.; Ley, K. Immunity and inflammation in atherosclerosis. Circ. Res. 2019, 124, 315–327. [Google Scholar] [CrossRef]
- Olejarz, W.; Lacheta, D.; Gluszko, A.; Migacz, E.; Kukwa, W.; Szczepanski, M.J.; Tomaszewski, P.; Nowicka, G. RAGE and TLRs as key targets for antiatherosclerotic therapy. Biomed. Res. Int. 2018, 2018, 7675286. [Google Scholar] [CrossRef]
- Olejarz, W.; Gluszko, A.; Cyran, A.; Bednarek-Rajewska, K.; Proczka, R.; Smith, D.F.; Ishman, S.L.; Migacz, E.; Kukwa, W. TLRs and RAGE are elevated in carotid plaques from patients with moderate-to-severe obstructive sleep apnea syndrome. Sleep Breath 2020, 1–8. [Google Scholar] [CrossRef]
- Wu, M.Y.; Li, C.J.; Hou, M.F.; Chu, P.Y. New insights into the role of inflammation in the pathogenesis of atherosclerosis. Int. J. Mol. Sci. 2017, 18, 2034. [Google Scholar] [CrossRef]
- Wang, X.; Khalil, R.A. Matrix metalloproteinases, vascular remodeling, and vascular disease. Adv. Pharmacol. 2018, 81, 241–330. [Google Scholar]
- Kowara, M.; Cudnoch-Jedrzejewska, A.; Opolski, G.; Wlodarski, P. MicroRNA regulation of extracellular matrix components in the process of atherosclerotic plaque destabilization. Clin. Exp. Pharmacol. Physiol. 2017, 44, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Gistera, A.; Hansson, G.K. The immunology of atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Falk, E. Pathogenesis of atherosclerosis. J. Am. Coll. Cardiol. 2006, 47, C7–C12. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Peng, W.; Xia, X.; Li, R.; Wang, Y.; Wei, D. Endothelial-to-mesenchymal transition: A potential mechanism for atherosclerosis plaque progression and destabilization. DNA Cell Biol. 2017, 36, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Nissinen, L.; Kahari, V.M. Matrix metalloproteinases in inflammation. Biochim. Biophys. Acta 2014, 1840, 2571–2580. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Maeda, T.; Zhang, H.; Berry, G.J.; Zeisbrich, M.; Brockett, R.; Greenstein, A.E.; Tian, L.; Goronzy, J.J.; Weyand, C.M. Mmp (matrix metalloprotease)-9-producing monocytes enable t cells to invade the vessel wall and cause vasculitis. Circ. Res. 2018, 123, 700–715. [Google Scholar] [CrossRef]
- Uzui, H.; Harpf, A.; Liu, M.; Doherty, T.M.; Shukla, A.; Chai, N.N.; Tripathi, P.V.; Jovinge, S.; Wilkin, D.J.; Asotra, K.; et al. Increased expression of membrane type 3-matrix metalloproteinase in human atherosclerotic plaque: Role of activated macrophages and inflammatory cytokines. Circulation 2002, 106, 3024–3030. [Google Scholar] [CrossRef]
- Raffetto, J.D.; Khalil, R.A. Matrix metalloproteinases and their inhibitors in vascular remodeling and vascular disease. Biochem. Pharmacol. 2008, 75, 346–359. [Google Scholar] [CrossRef]
- Di Gregoli, K.; George, S.J.; Jackson, C.L.; Newby, A.C.; Johnson, J.L. Differential effects of tissue inhibitor of metalloproteinase (TIMP)-1 and TIMP-2 on atherosclerosis and monocyte/macrophage invasion. Cardiovasc. Res. 2016, 109, 318–330. [Google Scholar] [CrossRef]
- Nam, S.I.; Yu, G.I.; Kim, H.J.; Park, K.O.; Chung, J.H.; Ha, E.; Shin, D.H. A polymorphism at -1607 2g in the matrix metalloproteinase-1 (MMP-1) increased risk of sudden deafness in korean population but not at -519a/g in mmp-1. Laryngoscope 2011, 121, 171–175. [Google Scholar] [CrossRef]
- Akiba, S.; Kumazawa, S.; Yamaguchi, H.; Hontani, N.; Matsumoto, T.; Ikeda, T.; Oka, M.; Sato, T. Acceleration of matrix metalloproteinase-1 production and activation of platelet-derived growth factor receptor beta in human coronary smooth muscle cells by oxidized ldl and 4-hydroxynonenal. Biochim. Biophys. Acta 2006, 1763, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. Metalloproteinase production from macrophages—A perfect storm leading to atherosclerotic plaque rupture and myocardial infarction. Exp. Physiol. 2016, 101, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, Q.; Zhu, J.; Xiao, Q.; Zhang, L. Reactive oxygen species: Key regulators in vascular health and diseases. Br. J. Pharmacol. 2018, 175, 1279–1292. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. Metalloproteinase expression in monocytes and macrophages and its relationship to atherosclerotic plaque instability. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 2108–2114. [Google Scholar] [CrossRef]
- Valentin, F.; Bueb, J.L.; Kieffer, P.; Tschirhart, E.; Atkinson, J. Oxidative stress activates MMP-2 in cultured human coronary smooth muscle cells. Fundam Clin. Pharmacol. 2005, 19, 661–667. [Google Scholar] [CrossRef]
- Ayuk, S.M.; Abrahamse, H.; Houreld, N.N. The role of matrix metalloproteinases in diabetic wound healing in relation to photobiomodulation. J. Diabetes Res. 2016, 2016, 2897656. [Google Scholar] [CrossRef]
- Hernandez-Anzaldo, S.; Brglez, V.; Hemmeryckx, B.; Leung, D.; Filep, J.G.; Vance, J.E.; Vance, D.E.; Kassiri, Z.; Lijnen, R.H.; Lambeau, G.; et al. Novel role for matrix metalloproteinase 9 in modulation of cholesterol metabolism. J. Am. Heart Assoc. 2016, 5, 1–16. [Google Scholar] [CrossRef]
- Johnson, J.L. Metalloproteinases in atherosclerosis. Eur J. Pharmacol. 2017, 816, 93–106. [Google Scholar] [CrossRef]
- de Franciscis, S.; Serra, R. Matrix metalloproteinases and endothelial dysfunction: The search for new prognostic markers and for new therapeutic targets for vascular wall imbalance. Thromb. Res. 2015, 136, 5–6. [Google Scholar] [CrossRef]
- Carmona-Rivera, C.; Zhao, W.; Yalavarthi, S.; Kaplan, M.J. Neutrophil extracellular traps induce endothelial dysfunction in systemic lupus erythematosus through the activation of matrix metalloproteinase-2. Ann. Rheum Dis. 2015, 74, 1417–1424. [Google Scholar] [CrossRef]
- Amato, B.; Compagna, R.; Amato, M.; Grande, R.; Butrico, L.; Rossi, A.; Naso, A.; Ruggiero, M.; de Franciscis, S.; Serra, R. Adult vascular wall resident multipotent vascular stem cells, matrix metalloproteinases, and arterial aneurysms. Stem Cells Int. 2015, 2015, 434962. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. Dual role of matrix metalloproteinases (matrixins) in intimal thickening and atherosclerotic plaque rupture. Physiol. Rev. 2005, 85, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L. Matrix metalloproteinases: Influence on smooth muscle cells and atherosclerotic plaque stability. Expert Rev. Cardiovasc. Ther 2007, 5, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Kong, L.; Kang, J.; Vaughn, D.M.; Bush, G.D.; Walding, A.L.; Grigorian, A.A.; Robinson, J.S., Jr.; Nakayama, D.K. Interleukin-lbeta induces migration of rat arterial smooth muscle cells through a mechanism involving increased matrix metalloproteinase-2 activity. J. Surg. Res. 2011, 169, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Auge, N.; Maupas-Schwalm, F.; Elbaz, M.; Thiers, J.C.; Waysbort, A.; Itohara, S.; Krell, H.W.; Salvayre, R.; Negre-Salvayre, A. Role for matrix metalloproteinase-2 in oxidized low-density lipoprotein-induced activation of the sphingomyelin/ceramide pathway and smooth muscle cell proliferation. Circulation 2004, 110, 571–578. [Google Scholar] [CrossRef]
- Li, H.; Liang, J.; Castrillon, D.H.; DePinho, R.A.; Olson, E.N.; Liu, Z.P. Foxo4 regulates tumor necrosis factor alpha-directed smooth muscle cell migration by activating matrix metalloproteinase 9 gene transcription. Mol. Cell Biol. 2007, 27, 2676–2686. [Google Scholar] [CrossRef]
- Guo, R.W.; Yang, L.X.; Wang, H.; Liu, B.; Wang, L. Angiotensin ii induces matrix metalloproteinase-9 expression via a nuclear factor-kappab-dependent pathway in vascular smooth muscle cells. Regul. Pept. 2008, 147, 37–44. [Google Scholar] [CrossRef]
- Dwivedi, A.; Slater, S.C.; George, S.J. MMP-9 and -12 cause n-cadherin shedding and thereby beta-catenin signalling and vascular smooth muscle cell proliferation. Cardiovasc. Res. 2009, 81, 178–186. [Google Scholar] [CrossRef]
- Orbe, J.; Montero, I.; Rodriguez, J.A.; Beloqui, O.; Roncal, C.; Paramo, J.A. Independent association of matrix metalloproteinase-10, cardiovascular risk factors and subclinical atherosclerosis. J. Thromb. Haemost. 2007, 5, 91–97. [Google Scholar] [CrossRef]
- Liu, J.; Khalil, R.A. Matrix metalloproteinase inhibitors as investigational and therapeutic tools in unrestrained tissue remodeling and pathological disorders. Prog. Mol. Biol. Transl. Sci. 2017, 148, 355–420. [Google Scholar]
- Fischer, T.; Senn, N.; Riedl, R. Design and structural evolution of matrix metalloproteinase inhibitors. Chemistry 2019, 25, 7960–7980. [Google Scholar] [CrossRef] [PubMed]
- Mott, J.D.; Werb, Z. Regulation of matrix biology by matrix metalloproteinases. Curr. Opin. Cell Biol. 2004, 16, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Chistiakov, D.A.; Myasoedova, V.A.; Melnichenko, A.A.; Grechko, A.V.; Orekhov, A.N. Calcifying matrix vesicles and atherosclerosis. Biomed. Res. Int. 2017, 2017, 7463590. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, H.; Yan, L.; Du, W.; Zhang, M.; Chen, H.; Zhang, L.; Li, G.; Li, J.; Dong, Y.; et al. MMP-2 and MMP-9 contribute to the angiogenic effect produced by hypoxia/15-hete in pulmonary endothelial cells. J. Mol. Cell Cardiol. 2018, 121, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Butoi, E.; Gan, A.M.; Tucureanu, M.M.; Stan, D.; Macarie, R.D.; Constantinescu, C.; Calin, M.; Simionescu, M.; Manduteanu, I. Cross-talk between macrophages and smooth muscle cells impairs collagen and metalloprotease synthesis and promotes angiogenesis. Biochim. Biophys. Acta 2016, 1863, 1568–1578. [Google Scholar] [CrossRef] [PubMed]
- Kalampogias, A.; Siasos, G.; Oikonomou, E.; Tsalamandris, S.; Mourouzis, K.; Tsigkou, V.; Vavuranakis, M.; Zografos, T.; Deftereos, S.; Stefanadis, C.; et al. Basic mechanisms in atherosclerosis: The role of calcium. Med. Chem. 2016, 12, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Gaubatz, J.W.; Ballantyne, C.M.; Wasserman, B.A.; He, M.; Chambless, L.E.; Boerwinkle, E.; Hoogeveen, R.C. Association of circulating matrix metalloproteinases with carotid artery characteristics: The atherosclerosis risk in communities carotid mri study. Arterioscler Thromb. Vasc Biol. 2010, 30, 1034–1042. [Google Scholar] [CrossRef]
- Sasaki, T.; Nakamura, K.; Sasada, K.; Okada, S.; Cheng, X.W.; Suzuki, T.; Murohara, T.; Sato, K.; Kuzuya, M. Matrix metalloproteinase-2 deficiency impairs aortic atherosclerotic calcification in apoe-deficient mice. Atherosclerosis 2013, 227, 43–50. [Google Scholar] [CrossRef]
- Rattazzi, M.; Bennett, B.J.; Bea, F.; Kirk, E.A.; Ricks, J.L.; Speer, M.; Schwartz, S.M.; Giachelli, C.M.; Rosenfeld, M.E. Calcification of advanced atherosclerotic lesions in the innominate arteries of apoe-deficient mice: Potential role of chondrocyte-like cells. Arterioscler Thromb. Vasc Biol. 2005, 25, 1420–1425. [Google Scholar] [CrossRef]
- Gresele, P.; Falcinelli, E.; Sebastiano, M.; Momi, S. Matrix metalloproteinases and platelet function. Prog. Mol. Biol. Transl. Sci. 2017, 147, 133–165. [Google Scholar]
- Sebastiano, M.; Momi, S.; Falcinelli, E.; Bury, L.; Hoylaerts, M.F.; Gresele, P. A novel mechanism regulating human platelet activation by mmp-2-mediated par1 biased signaling. Blood 2017, 129, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Purroy, A.; Roncal, C.; Orbe, J.; Meilhac, O.; Belzunce, M.; Zalba, G.; Villa-Bellosta, R.; Andres, V.; Parks, W.C.; Paramo, J.A.; et al. Matrix metalloproteinase-10 deficiency delays atherosclerosis progression and plaque calcification. Atherosclerosis 2018, 278, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Newby, A.C. Metalloproteinases and vulnerable atherosclerotic plaques. Trends Cardiovasc. Med. 2007, 17, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Montero, I.; Orbe, J.; Varo, N.; Beloqui, O.; Monreal, J.I.; Rodriguez, J.A.; Diez, J.; Libby, P.; Paramo, J.A. C-reactive protein induces matrix metalloproteinase-1 and -10 in human endothelial cells: Implications for clinical and subclinical atherosclerosis. J. Am. Coll. Cardiol. 2006, 47, 1369–1378. [Google Scholar] [CrossRef]
- Lemaitre, V.; O'Byrne, T.K.; Borczuk, A.C.; Okada, Y.; Tall, A.R.; D'Armiento, J. Apoe knockout mice expressing human matrix metalloproteinase-1 in macrophages have less advanced atherosclerosis. J. Clin. Investig. 2001, 107, 1227–1234. [Google Scholar] [CrossRef]
- Hojo, Y.; Ikeda, U.; Takahashi, M.; Sakata, Y.; Takizawa, T.; Okada, K.; Saito, T.; Shimada, K. Matrix metalloproteinase-1 expression by interaction between monocytes and vascular endothelial cells. J. Mol. Cell Cardiol. 2000, 32, 1459–1468. [Google Scholar] [CrossRef]
- Sukhova, G.K.; Schonbeck, U.; Rabkin, E.; Schoen, F.J.; Poole, A.R.; Billinghurst, R.C.; Libby, P. Evidence for increased collagenolysis by interstitial collagenases-1 and -3 in vulnerable human atheromatous plaques. Circulation 1999, 99, 2503–2509. [Google Scholar] [CrossRef]
- Molloy, K.J.; Thompson, M.M.; Jones, J.L.; Schwalbe, E.C.; Bell, P.R.; Naylor, A.R.; Loftus, I.M. Unstable carotid plaques exhibit raised matrix metalloproteinase-8 activity. Circulation 2004, 110, 337–343. [Google Scholar] [CrossRef]
- Lee, H.S.; Noh, J.Y.; Shin, O.S.; Song, J.Y.; Cheong, H.J.; Kim, W.J. Matrix metalloproteinase-13 in atherosclerotic plaque is increased by influenza a virus infection. J. Infect. Dis 2020, 221, 256–266. [Google Scholar] [CrossRef]
- Levkau, B.; Kenagy, R.D.; Karsan, A.; Weitkamp, B.; Clowes, A.W.; Ross, R.; Raines, E.W. Activation of metalloproteinases and their association with integrins: An auxiliary apoptotic pathway in human endothelial cells. Cell Death Differ. 2002, 9, 1360–1367. [Google Scholar] [CrossRef]
- Williams, H.; Johnson, J.L.; Jackson, C.L.; White, S.J.; George, S.J. MMP-7 mediates cleavage of n-cadherin and promotes smooth muscle cell apoptosis. Cardiovasc. Res. 2010, 87, 137–146. [Google Scholar] [CrossRef]
- Stoneman, V.E.; Bennett, M.R. Role of apoptosis in atherosclerosis and its therapeutic implications. Clin. Sci. (Lond.) 2004, 107, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Nakazawa, T.; Cho, A.; Dastvan, F.; Shilling, D.; Daum, G.; Reidy, M. Regulation of arterial lesions in mice depends on differential smooth muscle cell migration: A role for sphingosine-1-phosphate receptors. J. Vasc. Surg. 2007, 46, 756–763. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rauch, I.; Iglseder, B.; Paulweber, B.; Ladurner, G.; Strasser, P. MMP-9 haplotypes and carotid artery atherosclerosis: An association study introducing a novel multicolour multiplex realtime pcr protocol. Eur. J. Clin. Investig. 2008, 38, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.T.; Collins, E.T.; Marine, L.A.; Uberti, M.G.; Uchida, H.; Leidenfrost, J.E.; Khan, M.F.; Boc, K.P.; Abendschein, D.R.; Parks, W.C. Matrix metalloproteinase-9 modulation by resident arterial cells is responsible for injury-induced accelerated atherosclerotic plaque development in apolipoprotein e-deficient mice. Arterioscler Thromb. Vasc Biol. 2005, 25, 1020–1025. [Google Scholar] [CrossRef]
- Su, W.; Gao, F.; Lu, J.; Wu, W.; Zhou, G.; Lu, S. Levels of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 mrnas in patients with primary hypertension or hypertension-induced atherosclerosis. J. Int. Med. Res. 2012, 40, 986–994. [Google Scholar] [CrossRef]
- Shah, P.K.; Falk, E.; Badimon, J.J.; Fernandez-Ortiz, A.; Mailhac, A.; Villareal-Levy, G.; Fallon, J.T.; Regnstrom, J.; Fuster, V. Human monocyte-derived macrophages induce collagen breakdown in fibrous caps of atherosclerotic plaques. Potential role of matrix-degrading metalloproteinases and implications for plaque rupture. Circulation 1995, 92, 1565–1569. [Google Scholar]
- Nenseter, M.S.; Narverud, I.; Graesdal, A.; Bogsrud, M.P.; Halvorsen, B.; Ose, L.; Aukrust, P.; Holven, K.B. Elevated serum MMP-9/TIMP-1 ratio in patients with homozygous familial hypercholesterolemia: Effects of LDL-apheresis. Cytokine 2013, 61, 194–198. [Google Scholar] [CrossRef]
- Gu, C.; Wang, F.; Zhao, Z.; Wang, H.; Cong, X.; Chen, X. Lysophosphatidic acid is associated with atherosclerotic plaque instability by regulating nf-kappab dependent matrix metalloproteinase-9 expression via LPA2 in macrophages. Front. Physiol. 2017, 8, 266. [Google Scholar] [CrossRef]
- Giaginis, C.; Zira, A.; Katsargyris, A.; Klonaris, C.; Theocharis, S. Clinical implication of plasma neutrophil gelatinase-associated lipocalin (NGAL) concentrations in patients with advanced carotid atherosclerosis. Clin. Chem. Lab. Med. 2010, 48, 1035–1041. [Google Scholar] [CrossRef]
- Serra, R.; Grande, R.; Montemurro, R.; Butrico, L.; Calio, F.G.; Mastrangelo, D.; Scarcello, E.; Gallelli, L.; Buffone, G.; de Franciscis, S. The role of matrix metalloproteinases and neutrophil gelatinase-associated lipocalin in central and peripheral arterial aneurysms. Surgery 2015, 157, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Sivalingam, Z.; Larsen, S.B.; Grove, E.L.; Hvas, A.M.; Kristensen, S.D.; Magnusson, N.E. Neutrophil gelatinase-associated lipocalin as a risk marker in cardiovascular disease. Clin. Chem. Lab. Med. 2017, 56, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.J.H.; Prince, R.L.; Thompson, P.L.; Thavapalachandran, S.; Ooi, E.; Devine, A.; Lim, E.E.M.; Byrnes, E.; Wong, G.; Lim, W.H.; et al. Association between plasma neutrophil gelatinase-associated lipocalin and cardiac disease hospitalizations and deaths in older women. J. Am. Heart Assoc. 2019, 8, e011028. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.K. Biomarkers of plaque instability. Curr. Cardiol. Rep. 2014, 16, 547. [Google Scholar] [CrossRef] [PubMed]
- Mangge, H.; Almer, G. Immune-mediated inflammation in vulnerable atherosclerotic plaques. Molecules 2019, 24, 3072. [Google Scholar] [CrossRef]
- Romero, J.R.; Vasan, R.S.; Beiser, A.S.; Polak, J.F.; Benjamin, E.J.; Wolf, P.A.; Seshadri, S. Association of carotid artery atherosclerosis with circulating biomarkers of extracellular matrix remodeling: The framingham offspring study. J. Stroke Cerebrovasc. Dis. 2008, 17, 412–417. [Google Scholar] [CrossRef][Green Version]
- Ye, S.; Gale, C.R.; Martyn, C.N. Variation in the matrix metalloproteinase-1 gene and risk of coronary heart disease. Eur. Heart J. 2003, 24, 1668–1671. [Google Scholar] [CrossRef]
- Huang, X.Y.; Han, L.Y.; Huang, X.D.; Guan, C.H.; Mao, X.L.; Ye, Z.S. Association of matrix metalloproteinase-1 and matrix metalloproteinase-3 gene variants with ischemic stroke and its subtype. J. Stroke Cerebrovasc. Dis. 2017, 26, 368–375. [Google Scholar] [CrossRef]
- Sapienza, P.; di Marzo, L.; Borrelli, V.; Sterpetti, A.V.; Mingoli, A.; Cresti, S.; Cavallaro, A. Metalloproteinases and their inhibitors are markers of plaque instability. Surgery 2005, 137, 355–363. [Google Scholar] [CrossRef]
- Dabek, J.; Glogowska-Ligus, J.; Szadorska, B. Transcription activity of MMP-2 and MMP-9 metalloproteinase genes and their tissue inhibitor (TIMP-2) in acute coronary syndrome patients. J. Postgrad Med. 2013, 59, 115–120. [Google Scholar]
- Turu, M.M.; Krupinski, J.; Catena, E.; Rosell, A.; Montaner, J.; Rubio, F.; Alvarez-Sabin, J.; Cairols, M.; Badimon, L. Intraplaque MMP-8 levels are increased in asymptomatic patients with carotid plaque progression on ultrasound. Atherosclerosis 2006, 187, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Lenglet, S.; Mach, F.; Montecucco, F. Role of matrix metalloproteinase-8 in atherosclerosis. Mediators Inflamm. 2013, 2013, 659282. [Google Scholar] [CrossRef] [PubMed]
- Okada, Y.; Nakanishi, I. Activation of matrix metalloproteinase 3 (stromelysin) and matrix metalloproteinase 2 ('gelatinase') by human neutrophil elastase and cathepsin g. FEBS Lett. 1989, 249, 353–356. [Google Scholar] [CrossRef]
- Ferry, G.; Lonchampt, M.; Pennel, L.; de Nanteuil, G.; Canet, E.; Tucker, G.C. Activation of MMP-9 by neutrophil elastase in an in vivo model of acute lung injury. FEBS Lett. 1997, 402, 111–115. [Google Scholar] [CrossRef]
- Lenti, M.; Falcinelli, E.; Pompili, M.; de Rango, P.; Conti, V.; Guglielmini, G.; Momi, S.; Corazzi, T.; Giordano, G.; Gresele, P. Matrix metalloproteinase-2 of human carotid atherosclerotic plaques promotes platelet activation. Correlation with ischaemic events. Thromb. Haemost. 2014, 111, 1089–1101. [Google Scholar] [CrossRef]
- Dhillon, O.S.; Khan, S.Q.; Narayan, H.K.; Ng, K.H.; Mohammed, N.; Quinn, P.A.; Squire, I.B.; Davies, J.E.; Ng, L.L. Matrix metalloproteinase-2 predicts mortality in patients with acute coronary syndrome. Clin. Sci. 2010, 118, 249–257. [Google Scholar] [CrossRef]
- Peeters, W.; Moll, F.L.; Vink, A.; van der Spek, P.J.; de Kleijn, D.P.; de Vries, J.P.; Verheijen, J.H.; Newby, A.C.; Pasterkamp, G. Collagenase matrix metalloproteinase-8 expressed in atherosclerotic carotid plaques is associated with systemic cardiovascular outcome. Eur. Heart J. 2011, 32, 2314–2325. [Google Scholar] [CrossRef]
- Seifert, R.; Kuhlmann, M.T.; Eligehausen, S.; Kiefer, F.; Hermann, S.; Schafers, M. Molecular imaging of MMP activity discriminates unstable from stable plaque phenotypes in shear-stress induced murine atherosclerosis. PLoS ONE 2018, 13, e0204305. [Google Scholar] [CrossRef]
- Inokubo, Y.; Hanada, H.; Ishizaka, H.; Fukushi, T.; Kamada, T.; Okumura, K. Plasma levels of matrix metalloproteinase-9 and tissue inhibitor of metalloproteinase-1 are increased in the coronary circulation in patients with acute coronary syndrome. Am. Heart J. 2001, 141, 211–217. [Google Scholar] [CrossRef]
- Kelly, D.; Khan, S.Q.; Thompson, M.; Cockerill, G.; Ng, L.L.; Samani, N.; Squire, I.B. Plasma tissue inhibitor of metalloproteinase-1 and matrix metalloproteinase-9: Novel indicators of left ventricular remodelling and prognosis after acute myocardial infarction. Eur. Heart J. 2008, 29, 2116–2124. [Google Scholar] [CrossRef]
- Goncalves, I.; Bengtsson, E.; Colhoun, H.M.; Shore, A.C.; Palombo, C.; Natali, A.; Edsfeldt, A.; Duner, P.; Fredrikson, G.N.; Bjorkbacka, H.; et al. Elevated plasma levels of MMP-12 are associated with atherosclerotic burden and symptomatic cardiovascular disease in subjects with type 2 diabetes. Arterioscler Thromb. Vasc Biol. 2015, 35, 1723–1731. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wei, G.; Hu, W.; Li, L.; Ye, Y.; Wang, H.; Wan, W.; Li, R.; Li, L.; Ma, L.; et al. Expression of matrix metalloproteinases-12 in st-segment elevation myocardial infarction: A case-control study. Medicine (Baltimore) 2017, 96, e8035. [Google Scholar] [CrossRef] [PubMed]
- Kai, H.; Ikeda, H.; Yasukawa, H.; Kai, M.; Seki, Y.; Kuwahara, F.; Ueno, T.; Sugi, K.; Imaizumi, T. Peripheral blood levels of matrix metalloproteases-2 and -9 are elevated in patients with acute coronary syndromes. J. Am. Coll. Cardiol. 1998, 32, 368–372. [Google Scholar] [CrossRef]
- Blankenberg, S.; Rupprecht, H.J.; Poirier, O.; Bickel, C.; Smieja, M.; Hafner, G.; Meyer, J.; Cambien, F.; Tiret, L.; AtheroGene, I. Plasma concentrations and genetic variation of matrix metalloproteinase 9 and prognosis of patients with cardiovascular disease. Circulation 2003, 107, 1579–1585. [Google Scholar] [CrossRef]
- Cavusoglu, E.; Ruwende, C.; Chopra, V.; Yanamadala, S.; Eng, C.; Clark, L.T.; Pinsky, D.J.; Marmur, J.D. Tissue inhibitor of metalloproteinase-1 (TIMP-1) is an independent predictor of all-cause mortality, cardiac mortality, and myocardial infarction. Am. Heart J. 2006, 151, 1101–1108. [Google Scholar] [CrossRef]
- Lubos, E.; Schnabel, R.; Rupprecht, H.J.; Bickel, C.; Messow, C.M.; Prigge, S.; Cambien, F.; Tiret, L.; Munzel, T.; Blankenberg, S. Prognostic value of tissue inhibitor of metalloproteinase-1 for cardiovascular death among patients with cardiovascular disease: Results from the atherogene study. Eur. Heart J. 2006, 27, 150–156. [Google Scholar] [CrossRef]
- Loftus, I.M.; Naylor, A.R.; Bell, P.R.; Thompson, M.M. Plasma MMP-9—A marker of carotid plaque instability. Eur J. Vasc. Endovasc. Surg. 2001, 21, 17–21. [Google Scholar] [CrossRef]
- Kunte, H.; Amberger, N.; Busch, M.A.; Ruckert, R.I.; Meiners, S.; Harms, L. Markers of instability in high-risk carotid plaques are reduced by statins. J. Vasc. Surg. 2008, 47, 513–522. [Google Scholar] [CrossRef]
- Loftus, I.M.; Naylor, A.R.; Goodall, S.; Crowther, M.; Jones, L.; Bell, P.R.; Thompson, M.M. Increased matrix metalloproteinase-9 activity in unstable carotid plaques. A potential role in acute plaque disruption. Stroke 2000, 31, 40–47. [Google Scholar] [CrossRef]
- Wang, L.X.; Lu, S.Z.; Zhang, W.J.; Song, X.T.; Chen, H.; Zhang, L.J. Comparision of high sensitivity c-reactive protein and matrix metalloproteinase 9 in patients with unstable angina between with and without significant coronary artery plaques. Chin. Med. J. (Engl.) 2011, 124, 1657–1661. [Google Scholar]
- Koenig, W.; Khuseyinova, N. Biomarkers of atherosclerotic plaque instability and rupture. Arterioscler Thromb. Vasc Biol. 2007, 27, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.D.; Wu, P.S.; Hou, Y.Q.; Huang, Z.; Zhou, Z.J.; Guo, Z.G.; Xiu, J.C.; Wang, Y.G. Plasma matrix metalloproteinases-2 and -9 levels are elevated in patients with acute coronary syndrome and coronary chronic total occlusion. Nan Fang Yi Ke Da Xue Xue Bao 2009, 29, 1004–1007. [Google Scholar] [PubMed]
- Radosinska, J.; Barancik, M.; Vrbjar, N. Heart failure and role of circulating MMP-2 and MMP-9. Panminerva Med. 2017, 59, 241–253. [Google Scholar] [PubMed]
- Geovanini, G.R.; Libby, P. Atherosclerosis and inflammation: Overview and updates. Clin. Sci. (Lond) 2018, 132, 1243–1252. [Google Scholar] [CrossRef]
- Ridker, P.M.; MacFadyen, J.G.; Thuren, T.; Libby, P. Residual inflammatory risk associated with interleukin-18 and interleukin-6 after successful interleukin-1beta inhibition with canakinumab: Further rationale for the development of targeted anti-cytokine therapies for the treatment of atherothrombosis. Eur. Heart J. 2019, 1–11. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| SUBFAMILY | MMP | Tissue Distribution | Substrates |
|---|---|---|---|
| Collagenases | |||
| Collagenase-1 | MMP-1 | Endothelial cells, VSMCs, vascular wall, platelets, fibroblasts, macrophages | Collagens: I, II, III, VII, VIII, X Gelatins, entactin, aggrecan, link protein |
| Collagenase-2 | MMP-8 | Macrophages, neutrophils | Collagens: I, II, III; Aggrecan link protein |
| Collagenase-3 | MMP-13 | Vascular wall, SMCs, macrophages | Collagens: I, II, III |
| Gelatinases | |||
| Gelatinase-A | MMP-2 | Endothelial cells, VSMCs, adventitia, leukocytes, dermal fibroblasts, platelets | Collagens: I, IV, V, VII, X, XI; Gelatins, elastin, fibronectin, laminin, b-amyloid protein precursor |
| Gelatinase-B | MMP-9 | Endothelial cells, VSMCs, adventitia, vascular wall, macrophages | Collagens: IV, V, XIV; Gelatins, elastin, entactin, vitronectin |
| Stromelysins | |||
| Stromelysin-1 | MMP-3 | Endothelial cells, VSMCs, vascular wall, platelets | Collagens: III, IV, IX, X; Aggrecan, fibronectin, laminin, elastin, gelatins, casein |
| Stromelysin-2 | MMP-10 | Uterus | Collagens: II, IV, V; Aggrecan, fibronectin, gelatins, activate procollagenase |
| Stromelysin-3 | MMP-11 | Uterus, brain | Collagen IV, weakly fibronectin, laminin, aggrecan, gelatins, IGFBP-1, a1-protease inhibitor |
| Matrilysins | |||
| Matrilysin-1 | MMP-7 | Endothelial cells, VSMCs, vascular wall, uterus | Collagen IV, aggrecan fibronectin, laminin, entactin, vitronectin, casein, IGFBP-1 |
| Matrilysin-2 | MMP-26 | Collagen IV, gelatin, fibronectin | |
| Membrane-type MMPs | |||
| MT1-MMP | MMP-14 | Vascular wall, platelets, fibroblasts, uterus, brain | Collagens I, II, III; fibronectin, laminin-1, dermatan sulfate |
| MT2-MMP | MMP-15 | Fibroblasts, leukocytes | Large tenascin-C, fibronectin, laminin, entactin, aggrecan, perlecan |
| MT3-MMP | MMP-16 | Vascular wall, leukocytes | Collagen III, gelatin, casein, fibronectin |
| MT4-MMP | MMP-17 | Brain | Activates MMP2 by cleavage |
| MT5-MMP | MMP-24 | Leukocytes, lung, pancreas, kidney, brain | Activates MMP2 by cleavage |
| MT6-MMP | MMP-25 | Leukocytes | Inactivates alpha-1 proteinase inhibitor |
| Other | |||
| Metalloelastase | MMP-12 | VSMCs, fibroblasts, macrophages, great saphenous vein | Elastin, fibronectin |
| RASI-1 | MMP-19 | Liver | Gelatin |
| Enamelysin | MMP-20 | Tooth enamel | Amelogenin |
| Xenopus-MMP | MMP-21 | Fibroblasts, macrophages, placenta | |
| CA-MMP | MMP-23 | Ovary, testis | Gelatin |
| Epilysin | MMP-28 | Skin, keratinocytes | Casein |
| Vascular Inflammation and Endothelial Dysfunction | |
| Increase of inflammatory cell migration and invasion into the arterial wall | MMP-1, MMP-8, MMP-9, MMP-12, MMP-13, MMP-14 |
| Influence on endothelial dysfunction | MMP-2, MMP-9 |
| Participation in oxLDL effect | MMP-1, MMP-2, MMP-8, MMP-9 |
| Promotion of EC apoptosis | MMP-9 |
| Development of Plaque | |
| Increase of intima–media thickness | MMP-1, MMP-2, MMP-3, MMP-7, MMP-8, MMP-9, MMP-10 |
| Promotion of plaque growth | MMP-2 |
| Promotion of VSCMs migration | MMP-9 |
| Decrease of VSCMs contractility | MMP-2, MMP-9 |
| Neovascularization | |
| Degradation of ECM | MMP-1, MMP-8, MMP-9, MMP-12, MMP-13, MMP-14 |
| Release of growth factors | MMP-2, MMP-9, MMP-13, MMP-14 |
| Calcification | |
| Provoke vascular calcification | MMP-2, MMP-7 |
| Plaque Activation and Destabilization | |
| Apoptosis of VSMCs in the fibrous cap | MMP-7 |
| Collagenolytic activity | MMP-1, MMP-8, MMP-13 |
| Enhances plaque rupture | MMP-1, MMP-2, MMP-3, MMP-7, MMP-8, MMP-9, MMP-12, MMP-13, MMP-14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olejarz, W.; Łacheta, D.; Kubiak-Tomaszewska, G. Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability. Int. J. Mol. Sci. 2020, 21, 3946. https://doi.org/10.3390/ijms21113946
Olejarz W, Łacheta D, Kubiak-Tomaszewska G. Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability. International Journal of Molecular Sciences. 2020; 21(11):3946. https://doi.org/10.3390/ijms21113946
Chicago/Turabian StyleOlejarz, Wioletta, Dominika Łacheta, and Grażyna Kubiak-Tomaszewska. 2020. "Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability" International Journal of Molecular Sciences 21, no. 11: 3946. https://doi.org/10.3390/ijms21113946
APA StyleOlejarz, W., Łacheta, D., & Kubiak-Tomaszewska, G. (2020). Matrix Metalloproteinases as Biomarkers of Atherosclerotic Plaque Instability. International Journal of Molecular Sciences, 21(11), 3946. https://doi.org/10.3390/ijms21113946