Abstract
Mitochondria play a central role in a plethora of processes related to the maintenance of cellular homeostasis and genomic integrity. They contribute to preserving the optimal functioning of cells and protecting them from potential DNA damage which could result in mutations and disease. However, perturbations of the system due to senescence or environmental factors induce alterations of the physiological balance and lead to the impairment of mitochondrial functions. After the description of the crucial roles of mitochondria for cell survival and activity, the core of this review focuses on the “mitochondrial switch” which occurs at the onset of neuronal degeneration. We dissect the pathways related to mitochondrial dysfunctions which are shared among the most frequent or disabling neurodegenerative diseases such as Alzheimer’s, Parkinson’s, and Huntington’s, Amyotrophic Lateral Sclerosis, and Spinal Muscular Atrophy. Can mitochondrial dysfunctions (affecting their morphology and activities) represent the early event eliciting the shift towards pathological neurobiological processes? Can mitochondria represent a common target against neurodegeneration? We also review here the drugs that target mitochondria in neurodegenerative diseases.
1. Introduction: The Peculiarity of Mitochondria
Classically, mitochondria were considered the “powerhouse” of the cell, the energetic core generating ATP for cell activities [1]. Intensive research on their morphology and functions showed that mitochondria play several different roles.
Interestingly, mitochondria possess their own genome, the mitochondrial DNA (mtDNA), packed in nucleoids in the mitochondrial matrix in close association with the mitochondrial inner membrane [2]. Mutations at the mtDNA level are causative for many human diseases which are generally defined as mitochondrial disorders [3]. Moreover, mitochondria are vital and dynamic organelles able to modify their shape and size in order to respond to the cellular needs and, therefore, to maintain cellular homeostasis [4]. Mitochondria are the site of the oxidation of metabolites, for example through Krebs’ cycle, and of the β-oxidation of fatty acids [5]. Mitochondria are also the main generators of the reactive oxygen species (ROS) [6] and they participate to cell proliferation by maintaining a proper redox state and by recycling oxidized electron carriers. Importantly, mitochondria buffer calcium ions regulate, in turn, calcium homeostasis [7,8]. This is a crucial mitochondrial function impacting many cellular pathways such as the neurotransmitters’ release from neurons and glial cells [9].
In order to maintain the homeostasis of the cell, mitochondria play a crucial role in the choice of cell fate since they are also able to control cellular programmed cell death [10]. Indeed, mitochondria can induce apoptosis via caspase-dependent or independent mechanisms, the first by the activation of pro-apoptotic members of the B-cell lymphoma 2 (Bcl-2) family, the latter by the release of toxic mitochondrial proteins occurring after mitochondrial loss of function [11].
Mitochondria are extremely sensitive to every subtle change perturbing the homeostasis of the cell and, in turn, can modify their shape and number. Indeed, the processes of fusion and fission are fundamental, respectively, to repair a damaged mitochondrion or to increment their number, as in case of an increased demand of energy or to facilitate their removal when damaged in order to protect cellular integrity [12].
Additionally, the number and the size of cristae, which are dynamic bioenergetic compartments of the inner mitochondrial membrane where the respiratory chain occurs, adapt to the needs. Their plasticity guarantees a constant turnover to assure the balance between regeneration, biogenesis, and elimination of damaged mitochondria [13]. On the other hand, a regressive event occurring to mitochondria consists in “mitophagy”, i.e., the process of mitochondria degradation by autophagy [14], aiming to maintain the correct turnover of mitochondria [15].
Finally, mitochondria are also able to protect cell integrity by preventing the damage induced by viral infection [16]; indeed, they can also stimulate the innate immune response against these insults [17].
2. The “Mitochondrial Switch” and Its Impact on Neurodegeneration
Mitochondria are extremely sensitive to the insults that occur and accumulate in the cell, directly impacting on their function, consequently promoting disease development and progression. Indeed, besides human hereditary diseases caused by mutations of the mtDNA or in nuclear genes responsible for mitochondrial deregulation [18], mitochondrial dysfunctions are among the first events which occur in a vast number of pathological conditions ranking from diabetes [19], inflammatory diseases (such as multiple sclerosis [20]), to cancer [21] and neurodegenerative diseases [22]. We review here the “mitochondrial switch” which occurs during neurodegeneration. Neurodegenerative diseases consist of a group of heterogeneous disorders, but, nevertheless, they are all characterized by the progressive loss of specific neuronal populations and circuits in the central nervous system (CNS) triggered by mitochondria dysfunctions [23,24]. Another feature of the majority of neurodegenerative diseases is the progressive and highly disabling motor decline. Indeed, besides neuronal cells, also muscle cells are particularly enriched in mitochondria, since they need a high level of energy to function, and are heavily impacted by mitochondrial dysfunctions. There is growing evidence in the field exploring mitochondrial defects occurring also in the peripheral nervous system (PNS) and peripheral cells. Due to the low potential of neural regeneration, mitochondrial damage results in detrimental effects for neuron survival [25], while in peripheral cells the regeneration capacity is higher and clinical symptoms are evident only later in the disease.
Globally, in neurodegenerative diseases there is a “switch” in mitochondrial function which contributes importantly to the transition from a normal physiological to a degenerative condition. The accumulation of different stresses and the parallel impairment of a number of cell protective processes elicits neurodegeneration (Figure 1).
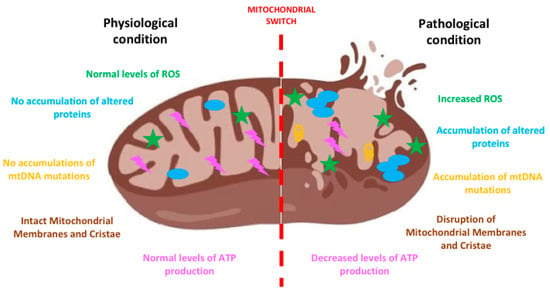
Figure 1.
The mitochondrial switch. The sketch shows the appearance and the features of a mitochondrion before and after the accumulation of damage responsible for its impairment. At the onset of the neurodegenerative process the increase of reactive oxygen species (ROS), mitochondrial DNA (mtDNA) mutations and altered proteins determines the swelling of mitochondria and the disruption of their membranes and cristae. Such an altered morphology heavily impacts on function determining, among others, decreases ATP production, increased ROS, leading to neuronal death. Created with BioRender software.
During neurodegeneration oxidative stress increases and intracellular ROS are formed, inducing to mtDNA mutations [1] and also to the disruption of mitochondrial membranes and cristae with a decrease in ATP production. Among the mechanisms eliciting oxidative stress, it is worth mentioning the process of deuteronation. Deuterium is a hydrogen isotope, present in nature in a well-defined ratio with hydrogen (1:6600) [26]: in body fluids, the probability of ATP synthase deuteronation process is 1/15,000 and the increase of this ratio can negatively interfere with ATP synthesis. Moreover, it has also been demonstrated that deuteronation can enhance formation of free radicals, slowing the electron trafficking in the mitochondrial electron transport chain (ETC) [27]. This mechanism has been evaluated in several conditions or pathologies (i.e., cancer, diabetes, aging, depression) and it can be reverted replacing heavy water with deuterium depleted water (DDW). Moreover, reduction of deuterium content by DDW ingestion can rescue long-term memory in Wistar rats restoring altered deuterium/hydrogen ratio and enhancing synaptic activity [28]. Indeed, DDW is able to promote antioxidant enzymes’ expression (SOD) and restore glutathione (GSH) levels mitigating, in turn, ROS formation and preventing fatty acid oxidation [29].
Moreover, the accumulation of altered proteins occurring in many neurodegenerative diseases can determine a decline in mitochondrial biogenesis, impacting on mitochondrial turnover and inducing structural and functional changes in cells. Different studies showed that various proteins involved in the above-mentioned processes and in mitophagy are affected in neurodegenerative diseases determining a dysregulation of these processes [30]. Indeed, mitophagy is required to remove damaged mitochondria and this process is finely tuned to maintain an equilibrate turnover of mitochondria and, from a bigger picture, proper cellular development and differentiation. Described for the first time in 1998 [31], mitophagy has gained considerable interest in the field of neurodegeneration in the last years; however, because of its fine tuning, it is a difficult target for designing new therapies against neurodegeneration. Some authors hypothesized a relation between mitophagy dysregulation and neurodegenerative diseases, based on the impairment in autophagosome–lysosome fusion and defects in lysosomal acidification, often observed in neurodegeneration [32].
Altogether, these events can trigger the process of mitochondrial membrane permeabilization and, when severely injured mitochondria are not appropriately removed, they release their contents into the cytosol and the extracellular environment: this represents a starting point of both apoptosis and necrosis. This process is called generation of damage-associated molecular patterns (DAMPs) and mitochondria represent the major site of DAMP formation. By triggering cell death, mitochondria-derived ROS, calcium, ATP, and the other signaling molecules also lead to neuroinflammation. The process of neuroinflammation refers to the proliferation and activation of microglia and/or astrocytes (microgliosis, astrogliosis) and, to note, neurodegenerative diseases all show evidence of neuroinflammation; the specific disease-related peculiarities are described later on in the sections dedicated to each pathology. Interestingly, neuroinflammation is closely related to mitochondria [33]: indeed, mitochondria can regulate inflammatory signaling and can modulate the innate immunity by generating ROS and, as a consequence, triggering neurodegeneration [34]. Closely related to ROS and DAMP formation, another largely studied mechanism by which mitochondria drive neuroinflammation is their ability to activate the NOD-, LRR-, and pyrin domain-containing protein 3 (NLRP3) inflammasome [35], in turn leading to the release of proinflammatory cytokines [36]. Interestingly, NLRP3 localizes in the cytosol and once activated it translocates into mitochondria and mitochondria-associated membranes (MAMs) [37]. Another mechanism by which mitochondria trigger and sustain neuroinflammation is closely related to their dynamism: indeed, the inhibition/stimulation of fission induces/reduces NLRP3 inflammasome assembly and activation, respectively [38].
In particular, mitochondrial dysfunctions (both affecting neuronal survival and triggering neuroinflammation) have been reported in Alzheimer’s (AD), Parkinson’s (PD), and Huntington’s (HD) diseases, Amyotrophic Lateral Sclerosis (ALS), and Spinal Muscular Atrophy (SMA), thus representing a potential biomarker of the diseases, as one major unifying basic mechanism involved in aging and neurodegeneration [39,40]. In the subparagraphs below we describe the mitochondrial dysfunctions which are peculiar or shared among the above-mentioned diseases.
2.1. Mitochondrial Dysfunctions in AD
AD is the most common and devastating neurodegenerative disease expected to affect more than 81 million people worldwide by 2040 [41,42]. The first evidence of pathogenic mechanisms in AD has been the progressive accumulation of beta-amyloid peptide (Aβ) extracellularly in the brain and of neurofibrillary tangles of hyperphosphorylated tau inside neurons [43], determining the progressive loss of cortical and hippocampal neurons and inducing brain atrophy, consequent cognitive and memory loss. More recently, advanced neuroimaging techniques showed evident metabolic alterations in the brain of AD patients at a very early stage of the disease [44]. Indeed, the degenerative process probably starts 20–30 years before the clinical onset and one of the major goals of AD research is the early detection before symptom onset to prevent the disease outbreak. Oxidative stress-induced damage occurs even before amyloid and tau deposition, and mitochondrial dysfunctions are early events which precede neurodegeneration [45]. In AD there is also evidence of neuroinflammation; indeed, microglia and reactive astrocytes are closely associated with amyloid plaques in patients’ brains [46].
Mitochondria appear morphologically and functionally altered, impacting on many processes such as excessive ROS formation, and resulting in the decrease of brain energy because of the reduction of ATP [47], alteration of calcium homeostasis, and of apoptosis induction [48]: indeed, altered levels of apoptotic markers such as Bcl-2, Bax, and caspases have been observed in models of AD [49]. Moreover, alterations in mitochondrial dynamics appear early in AD development, contributing to impair neuronal autophagy because of defective mitophagy [50]. There is a vast literature on the detrimental impact of the accumulation of damage and misfolded proteins inducing dysfunctions at the cellular and mitochondrial level not only in the NS but also in peripheral tissues such as patients fibroblasts [51] which show mitochondrial calcium dysregulation similar to those occurring in the brain [52]. Moreover, AD-related proteins, such as the Amyloid Precursor Protein (APP) and Presenilins (PSs), have been recently described as involved in peripheral processes associated to muscle trophism, neuromuscular junction (NMJ) formation and mitochondrial morphology [53,54,55]. Mitochondrial dysfunction in peripheral cells could represent a new target for early and more accessible diagnosis and, eventually, therapy for AD.
2.2. Mitochondrial Dysfunctions in PD
PD, the second most common neurodegenerative disease in aging, is characterized by the reduction of dopamine (DA) levels in the striatum due to the degeneration of dopaminergic neurons in the SNpc. Patients show a progressive muscle rigidity and tremors due to decrease in dopaminergic modulation on striatal neurons altering motor systems [56,57]. Inclusions of Lewy bodies and presence of immunoreactive α-synuclein and ubiquitin are also hallmarks of the disease [58]. Moreover, neuroinflammation plays a role in PD; reactive microglia in the substantia nigra together with positron emission tomography (PET) imaging studies of PD brains provided early evidence for the role of neuroinflammation in PD [59,60].
The first evidence of mitochondrial dysfunction in PD came in 1983, when Langston described chronic parkinsonism effects in young humans after the use of 1-methyl-4phenyl-1,2,3,6-tetrahydropyridine (MPTP), which is a prodrug to the neurotoxin 1-methyl-4-phenylpyridinium (MPP+); the latter interferes with the activity of the complex I of the electron transport chain [61] whose functioning is fundamental for dopaminergic neuron survival. Decades of research in the field confirmed that mitochondrial dysfunctions are a common feature in PD and are present in animal models, in induced pluripotent stem cell (iPSCs)-derived neurons and in patients’ brains [62,63,64,65]. Mitochondrial dysfunctions are related to both sporadic and familial PD and are associated to disturbances of mitochondrial function, morphology, and dynamics (for a detailed review see Bose and Beal [66]). High levels of ROS, due to the high metabolic demand, determine accumulation of toxic oxidative species and structural alterations of complex I, affecting mitochondria functionality in patient brains and mouse models [67,68]. Moreover, α-synuclein oligomers accumulation induces the permeabilization of the mitochondrial membrane and direct toxicity by increasing ROS production and consequently leading to neuronal death [69]. Disturbances in calcium homeostasis and calcium overload could force the opening of the mitochondrial permeabilization transition pore, resulting in ROS formation, Cyt C release, and apoptosis activation [70].
Mitochondrial dysfunctions in PD are also a result of changes in mitochondria biogenesis caused by the dysregulation of transcription factors such as the peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α), as demonstrated both in mice and humans [71,72]. Furthermore, mitochondrial fragmentation, which occurs rapidly after the loss of membrane potential, has been observed in PD: indeed, many proteins involved in fusion and fission (such as the dynamin-related protein 1 (DRP1)) are altered in a model of familial PD [73]. Notably, genes such as PINK1 and Parkin, which are related to familial-PD forms, are involved in the control of mitochondrial dynamics [74] and several other genetic mutations, including PINK1, Parkin, DJ-1, LRRK2, and α-Syn, have been linked to familial PD and the corresponding gene products are also involved in mitophagy. Additionally, also in PD, mitochondrial dysfunctions are not limited to CNS, but also involve peripheral tissues as it has been described in skeletal muscles and platelets [62,75].
2.3. Mitochondrial Dysfunctions in HD
HD is an autosomal dominant neurodegenerative disorder, caused by the expansion of CAG repeats in exon 1 of the huntingtin gene (HTT), leading to the specific loss of GABAergic striatal medium spiny neurons. Patients show impairments in behavior, muscle coordination, and a progressive mental decline leading to death [76]. The huntingtin protein (HTT) is involved in many processes such as axonal transport, signal transduction, and autophagy; its misfolding due to HTT mutation determines the disruption of the above-mentioned biological processes and of many others [77]. Mitochondrial dysfunctions play a central role in HD progression. Indeed, abnormal oxidative stress and ROS production, calcium imbalance, and decreased enzymatic activity of the respiratory chain complexes resulting in the alteration of lactate production have been observed both in mouse models of HD and in patients’ brains [78,79,80]. Several studies have revealed reduced mitophagy in the brain of HD patients [81]. Moreover, accumulation of mtDNA defects occurs early in HD to the point of being suggested as a potential biomarker of the disease [82,83]. Altogether, these mitochondria-derived defects recall pro-inflammatory activated innate immune cells; indeed, in HD, a remarkable neuroinflammation has been observed in brains from HD patients and PET imaging showed that microglia activation correlates with the pathology progression (for an extensive review on the subject see [84]).
As for the majority of neurodegenerative disorders, the approach in HD research field has always been to focus on the neurological symptoms and phenotypes. However, the HTT gene is ubiquitously expressed [85] and in HD there are important dysfunctions occurring outside the brain. In this regard, impaired uptake and levels of glucose and cardiac dysfunctions have been shown in HD mouse models [86,87]. Moreover, HD patients show a high predisposition to diabetes and muscle wasting, actually before neurodegeneration [88,89]. Interestingly, these dysfunctions are closely connected to mitochondrial activity and to tissues with high mitochondrial density. Moreover, mitochondrial alterations are accompanied by oxidative stress in skin fibroblasts of patients [90]; additionally, the enzymatic activity of the Aco2, which catalyzes the conversion of citrate to isocitrate in the TCA cycle, is impaired in PBMCs of HD patients and PreHD carriers [91]. Authors suggest it as a potential biomarker to assess the disease status of both patients and carriers, being an easy and affordable non-invasive blood test in the clinical routine.
2.4. Mitochondrial Dysfunctions in ALS
ALS is a neurodegenerative disorder due to the loss of upper and lower motor neurons (MNs) which entails muscle atrophy, progressive weakness, and respiratory failure leading to death [92]. Most cases are sporadic, whereas 10% are familial, with mutations mainly occurring on superoxide dismutase 1 (SOD1), FUS, TDP43 genes causing the accumulation of the corresponding mutant proteins [93,94,95]. However, the etiology of the disease is not yet understood: it includes excitotoxicity, protein misfolding and aggregation, dysregulation of RNA metabolism, and neuroinflammation [96]. Inflammatory changes have been largely described in ALS patients: astrogliosis is evident within the spinal cord (both ventral and dorsal horns) and brain (cortical gray matter and subcortical white matter), whereas microgliosis in the spinal cord ventral horns, corticospinal tract, and motor cortex [97,98].
For sure, mitochondrial-dependent dysfunctions such as ATP deprivation, oxidative stress, impaired cell signaling together with altered mitochondrial morphology/dynamics are also correlated to the pathogenesis of ALS and represent very early phenomena [99]. More in details, (i) a high amount of ROS, free radicals, and other toxic species such as peroxynitrite (ONOO−) [100] and (ii) an alteration of the levels of expression of proteins involved in mitochondrial fusion and fission (i.e., OPA1, Mitofusin for fusion, and Fis1 and DRP1 for fission) [100], were observed and (iii) apoptotic signaling is impacted in different ALS models. The latter triggers cell death, pivotal to keep the organism healthy by eliminating cells which cannot be rescued from damage, and mitochondria are fundamental for this. Indeed, as previously introduced, mitochondria induce apoptosis either via caspase-dependent or independent mechanisms. In ALS, the apoptotic cascade is activated [101] and the mutant SOD1 is playing a major role in this cascade by interacting with Bcl-2 [102]. Indeed, therapeutic strategies targeting mitochondrial dysfunction to prevent ALS progression represent a possible treatment option [103].
As for the other neurodegenerative diseases, also in ALS the PNS is dramatically affected and mitochondrial dysfunctions occur in peripheral tissues as well. MN degeneration is preceded by NMJ denervation, determining a dying-back retrograde neuropathy. Such defects may cause MN death by predisposing them to calcium-mediated excitotoxicity, increasing ROS generation and initiating the apoptotic pathway [104]. Moreover, features of mitophagy in the presynaptic terminals of ALS’ NMJs have been observed [105], suggesting impaired mitophagy as one of the mechanisms causing degeneration (for a review see Elfawy and Das [106]).
2.5. Mitochondrial Dysfunctions in SMA
SMA affects MNs in children and young adults with a mutation/deletion of the Survival Motor Neuron 1 (SMN1) gene [107]. SMN has a role in the assembly of small ribonucleoprotein particles, that function in pre-mRNA splicing and gene transcription and in mRNA transport in MN axons [108]. The outcome of its genetic alteration is a decrease in the levels of the functional protein which results in motor impairment, muscle atrophy, and premature death [109]. The disease severity in humans depends on SMN2 which is a highly homologous gene of SMN1. However, SMN2 produces only 10% of fully functional SMN; therefore, the degree of compensation relies on patient’s SMN2 copy numbers. Although the genetic cause of SMA has been identified, many aspects of its pathogenesis remain unclear. Interestingly, SMN deficiency has been associated to oxidative stress, mitochondrial dysfunction, and impairment of bioenergetic pathways in different models. SMN silencing in NSC-34 cell lines induces an increase in cytochrome c oxidase activity and mitochondrial membrane potential, generating free radicals [110]; in human SMA iPSCs, axonal mitochondrial transport and mitochondrial number and area are altered [111]. Studies in SMA murine models also show alterations in mitochondrial respiration, mitochondrial membrane potential and mobility, and confirm an increased oxidative stress level and fragmentation [112]. Additionally, proteomic studies show that bioenergetic pathways associated to glycolytic enzymes, such as GAPDH and PGK1, are altered in SMA models [113,114]. Moreover, besides cell-autonomous motoneuronal toxicity, also glial-mediated inflammation seems to negatively impact on neuronal survival and to promote progression and propagation of the degenerative process by the activation of apoptotic cascades [115].
Even if SMA has been classically categorized as a MN disease, SMA pathogenesis is more complex and many mechanisms and districts are involved. Indeed, since SMN protein is ubiquitously expressed, its lack affects not only MNs, but also NMJs, which show dramatic alterations, including immaturity, denervation, neurofilament accumulation, and impaired synaptic functions [116]. Additionally, skeletal [117] and heart muscle cells [118] are also dramatically affected: they require high levels of energy and are, therefore, particularly enriched in mitochondria. Alterations in the process of mitophagy have been described in the muscles of SMA patients [119]. Some researchers suggest that these peripheral abnormalities may be the origin of the detrimental effects observed on MN survival through retrograde signals coming from the muscles/NMJs [120,121]. On the contrary, others support the idea that peripheral dysfunctions derive from MN degeneration which, in turn, results in impairment of the nerve-muscle interplay [122]. This ‘chicken or the egg’ question about where the degeneration starts could help with understanding the important mechanisms of the disease and the development of disease-modifying interventions which could have greater therapeutic impact by preferentially targeting the CNS, the PNS, or peripheral tissues. Above all, one possibility is that mitochondrial impairment, which is an early event in SMA, occurs both at central and peripheral level, probably representing a relevant target independently from the tissue.
3. Are Mitochondria the Red Thread in Neurodegenerative Diseases?
Considering the enormous economic and social impacts, finding a cure for neurodegenerative disorders remains a priority in science. Researchers are focused on identifying the common pathogenic processes shared among these diseases, in order to design new treatments and/or drug combinations and repurposing. Indeed, scientists and pharmaceutical companies aim at the development of common therapies for multiple neurodegenerative diseases. To this aim, understanding the co-occurrence and overall interactions among these diseases is the first step for drug development.
The majority of neurodegenerative disorders have significant genetic components, with genetic heritability such as for AD, PD, HD, ALS, and SMA. To date, the most common and methodologically defined approaches used are based on microarrays, next-generation RNA sequencing, statistical and bioinformatics methods to detect shared genes, single nucleotide polymorphisms (SNPs), and profile networks of pathways underlying neurodegenerative mechanisms [123,124,125,126]. Genome wide association studies, which represent the largest publicly available resource in the genomic domain, reveal the similarities among diseases and can help in the process of drug screening and discovery (for a comprehensive review see Arneson et al. [127]). In addition to genetics, considering that the majority of the forms of neurodegeneration are sporadic, factors such as the ageing of the global population, the lifestyle, and the exposure to chemicals are taken into account too and are nowadays known risk factors for neurodegeneration [128,129].
Neurodegeneration can be defined as a process characterized by the loss of neuronal populations, progressive cognitive decline, and/or motor symptoms. As largely described above, there is overwhelming evidence of impaired mitochondrial functions as a causative factor driving the development of neurodegenerative diseases together with the unavoidable elevation of oxidative stress, the increase of free radicals in the brain, impaired DNA repair capability, and decreased tissue regeneration. Indeed, besides the intrinsic differences among diseases, mitochondria are a shared key crossing point in the biological processes driving neurodegeneration and determining the clinical outcomes. Mitochondrial dysfunctions could be considered a possible red thread in neurodegenerative diseases. This is the consequence of the fact that mitochondria, besides being the powerhouse of eukaryotic cells, are fundamental organelles involved in critical pathways connected to cell growth and differentiation, cellular signaling, apoptosis, and cell cycle control. Together with the fact that mitochondrial dysfunctions appear at the early disease onset and contribute to disease progression, they are a hallmark of neurodegeneration.
Interestingly, the mitochondrial switch driving neurodegeneration occurs indistinctly in every tissue, even in the periphery although neurodegeneration has always been approached and studied focusing on the CNS. To some extent, one could investigate neurodegenerative disorders as affecting mitochondria primarily. In Figure 2 the mitochondrial dysfunctions and effects on the CNS, PNS, and periphery are represented for AD, PD, HD, ALS, and SMA. Therefore, targeting mitochondria can represent new therapeutic approaches for different neurodegenerative diseases. Below, molecules tested for neurodegeneration targeting the main pathways related to mitochondrial-dependent dysfunctions, such as oxidative stress, mitochondrial biogenesis, mitochondrial membrane permeability and dynamics, are listed.
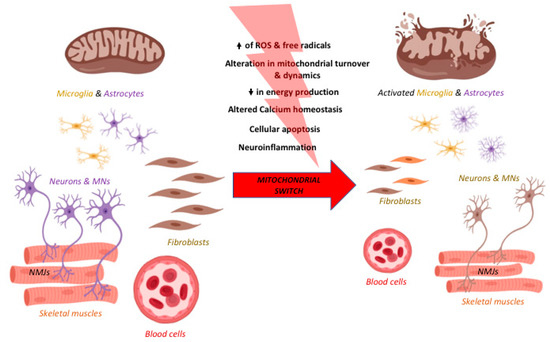
Figure 2.
The process of neurodegeneration in the central nervous system (CNS), in the periphery and across diseases. Neurodegeneration is a progressive process taking place not only in the CNS but also in the periphery. After the accumulation of damage at the cellular and, in particular, mitochondrial level, the switch from physiology to pathology is fast and rarely reversible and it is occurring in many cell types. Indeed, it occurs in the CNS but also in the periphery, in particular the neuromuscular junction (NMJ), the skeletal muscles, blood cells, and fibroblasts. Moreover, the major pathways involved are linked to ROS and free radical formation, alterations in ATP formation, calcium homeostasis, apoptosis, mitochondrial turnover and dynamics, and neuroinflammation in terms of microglia and astrocytes activation. These pathways are affected in all the neurodegenerative diseases discussed in this review. Increase of ROS is represented by the black arrow pointing up and reduction in energy production by the black arrow pointing down. Created with BioRender software.
4. Therapies Targeting Mitochondria
Despite different time of onset, symptoms, and etiology, the above-mentioned neurodegenerative pathologies share the progressive neuronal degeneration [130,131], resulting in the loss of neuronal populations and impairment of neurotransmission (a highly energy-demanding process) [132]. However, even if these neurodegenerative diseases have been studied for decades, up to now, most of them lack of disease-modifying therapies: the available treatments (summarized together with their main effects and limitations in Table 1) are generally symptomatic and they are unable to fully prevent the disease progression [133,134,135,136,137,138].

Table 1.
Available treatments for Alzheimer’s disease (AD), Parkinson’s disease (PD), Huntington’s disease (HD), Amyotrophic Lateral Sclerosis (ALS), and Spinal Muscular Atrophy (SMA). The table summarizes the most common therapies for these neurodegenerative pathologies. Unfortunately, the majority of these treatments are only symptomatic or palliative and cannot stop the disease progression (especially for AD, PD, HD, and ALS) or are less effective in milder and late treated patients (as in the case of SMA).
Nevertheless, in the last years the landscape of the potential drugs has expanded, in the attempt not only to stop the disease progression, but also to prevent the onset of symptoms [139]. To this aim, the identification of an early and possibly common target could represent a turning point in the treatment of these pathologies: in this scenario, mitochondria could be considered promising targets, because (i) their impairment can be detected since the earliest stages, influencing the onset and progression of the diseases, (ii) their dysfunctions are common to all the pathologies described here [140,141]. In the next paragraphs we will focus on the most common therapies that target these organelles in neurodegenerative diseases and that have been object of preclinical studies and/or whose role has provided promising outcomes in clinical trials, classifying them according to the mechanisms of action.
4.1. Antioxidant
As previously mentioned, ROS are produced by mitochondria both in physiological and pathological conditions [149] and can be responsible for onset and progression of many neurodegenerative diseases [150,151]. Furthermore, as discussed above, one of the major events related to neurodegenerative diseases, driven by the redox status, is microglial activation-derived neuroinflammation [152]: the induction of the expression of proinflammatory genes leading to the release of cytokines and chemokines can represent a consequence of the uncontrolled ROS production by mitochondria. This chronic inflammatory state is characteristic of many neurodegenerative diseases [153].
Many synthetic or natural molecules (summarized in Table 2) have been investigated, since they can reduce the ROS-induced effects, among which neuroinflammation, in a similar way due to their molecular structure, through direct or indirect mechanisms.
Table 2.
Antioxidant compounds for neurodegenerative disease treatment. The table lists synthetic and natural antioxidant compounds tested for the treatment of neurodegenerative diseases both in preclinical and clinical studies. The most recent and promising studies are collected here. For clinical trial ID we referred to https://clinicaltrials.gov.
4.1.1. Synthetic Antioxidants
As reviewed by Cenini and Voos [139], some compounds such as MitoQ play a key role in the ETC preventing ROS formation due to the electron leakage to oxygen. It has a lipophilic tail that lets it cross the inner mitochondrial membrane (IMM) and a benzoquinone ring that can be reduced to ubiquinol and oxidized back to ubiquinone, performing the antioxidant activity [154,155]. It induced neuroprotective effects in preclinical studies in vitro on AD cortical neurons and determined an increased lifespan and a milder cognitive decline, due to cholinergic neurons protection and synaptic preservation in AD mice [139,155]; furthermore, MitoQ prevented PD mice dopaminergic cells loss [156,157], increased lifespan and hindlimb strength of SOD1G93A mice [158], and finally mitigated muscle wasting in R6/2 HD mice [159].
As reviewed by Weissig [160], SkQ1 belongs to the quinone derivative group and it is still analyzed only in preclinical studies in OXYS rats providing learning and memory enhancing and behavioral improvement [139,161]. SkQ1 and MitoQ share the TPP+, a lipophilic cation that facilitates molecules targeting to mitochondria; the same residue is also present in Mito-Apo, another compound that displayed its neuroprotective effects in AD and PD both in vitro and in vivo providing, respectively, a reduction of neuronal degeneration [162] and an attenuation of motor deficit in PD mouse models when bound to apocynin, a NADPH oxidase inhibitor that counteracts ROS formation [158,163,164].
Beyond quinone structures, there are many other aromatic molecules that can provide themselves not only redox activity but also lipophilic properties to cross the membrane. For instance, the phenothiazine ring allows Methylene Blue both to reach the mitochondrial matrix and to perform redox reactions reducing ROS formation [165,166]; indeed, this activity is ensured in vivo, increasing attentional functions and preserving dopaminergic neurons in a PD rat model [157,167]. In Szeto-Schiller tetrapeptides the tyrosine or dimethyl tyrosine residues can scavenge ROS but, differently from the previous drugs, they act in the IMM without joining the mitochondrial matrix, as described by Rocha et al. [168]; both in in vitro and in vivo studies they have shown multiple functions deriving from the antioxidant activity, making it a promising compound for AD therapy [139], but they have also provided interesting results in further studies on ALS cell models and in PD and ALS mice, ensuring increased survival and motor performance enhancement due to neuroprotective effects [169].
Not only an aromatic structure can perform antioxidant activity scavenging ROS: thanks to its dithiolane ring, α-Lipoic acid [170] can perform redox reactions promoting a reduction of cytotoxic events in vitro and an amelioration of cognitive functions in AD animal models; the first promising results on AD patients have been already obtained in association with other antioxidant compounds but, unfortunately, there are no clinical trials to evaluate efficacy of isolated α-Lipoic acid [171].
Unlike the aforementioned compounds, inosine plays an indirect antioxidant activity: its derivative, urate, exerts neuroprotective effects mediated by the nuclear factor erythroid 2-related factor 2 (Nrf2), one of the main controllers of the response to oxidative stress; moreover, it seems to be able to increase GSH levels and its release. This was highlighted in different experimental studies in vitro but positive outcomes have also emerged in PD mouse models, encouraging scientists to consider preclinical studies also on ALS models [172,173]. Nowadays, there are some clinical trials that have confirmed safety and efficacy of inosine in increasing urate levels in both pathologies (NCT00833690; [174]; NCT02288091; [175]).
Finally, as reviewed by Reiter et al. [176], melatonin is able to counteract oxidative stress not only directly interacting with free radicals [177], but also by stimulating antioxidant enzymes and upregulating GSH synthesis, through an indirect pathway. For these reasons, its use in the treatment of neurodegenerative diseases has been proposed: in vitro it exerted an antiapoptotic activity and in vivo it improved behavioral and cognitive functions and extended the lifespan of AD and ALS mice and rats [151,178,179,180,181]; furthermore, it enhanced locomotor performances in PD mouse models [182]. Otherwise, meta-analysis and clinical trials on AD and PD patients revealed only an improvement in sleep quality without any ameliorations in terms of cognitive or motor functions and clinical trials testing melatonin in a specific range dose to establish its neuroprotective activity are needed [181,182,183]. Similarly, N-Acetylcysteine (NAC) can exert both a direct and an indirect antioxidant effect, reacting with ROS or restoring GSH levels [184,185,186]. Its efficacy has been already evaluated in vitro and in vivo, where it, respectively, downregulated apoptotic markers in AD and PD cells and enhanced mitochondrial activity (for example increasing brain connections or reducing lipid peroxidation [186]) and preventing the degeneration of MNs derived from SMA iPSCs [118]; furthermore, a mitigation of cognitive and motor impairment has been observed on HD mice [187]. Finally, different clinical trials have examined its efficacy on AD patients showing an amelioration in cognitive and behavioral functions (NCT01320527; [188]) and an increase in GSH brain levels in PD patients, although this outcome has not been obtained in all clinical trials, maybe because of NAC low oral bioavailability [186].
4.1.2. Natural Antioxidants
Beside the synthetic antioxidants, several natural compounds (taken as dietary supplements) can play a direct antioxidant role. Among them, Vitamin C can scavenge free radicals when oxidized to dehydroascorbate and recycled back to ascorbic acid through a radical mechanism [189]; its effectiveness in reducing apoptotic process and in regulating ROS balance and oxygen consumption has been already proven, respectively, on Aβ1-42 peptide-treated human cortical neurons and in APP/PSEN1 and 5XFAD Tg mice [190,191,192]. Even Vitamin E, that includes its tocopherol and tocotrienol derivatives, can regenerate tocopherol structure after lipid peroxidation inhibition, in a mechanism that can involve Vitamin C or GSH, as described by Stahl and Sies [189]. Preclinical studies on aged mice and on APP/PS1 mice showed promising results; indeed, ROS scavenging can recover mitochondrial damage, but also enhance mitochondrial biogenesis and bioenergetics: this in turn has effects on cognitive and behavioral impairment improvement [193,194,195]. Despite these encouraging outcomes in preclinical stages, further clinical trials are needed to investigate the real effects of Vitamin E supplementation in AD in order to consider all the factors that can influence patients’ responsiveness [196]. Even in ALS, Vitamin E intake is debated because the positive outcomes obtained in preclinical trials on ALS onset and progression cannot always be translated to patients [151,197].
Carotenoids (i.e., Astaxanthin) can exert a double antioxidant activity, since they can both quench ROS through a radical mechanism (for an extended review see Fiedor and Burda [198]), or improve the activity of several antioxidant enzymes like SOD, CAT, and GPX [199]; for instance, their efficacy has been already proven on an AD cell model showing a prevention of synaptotoxic events due to ROS production [200].
4.2. Mitochondrial Biogenesis and Permeability
Mitochondrial biogenesis is an important process, since increasing mitochondria number can partially rescue, at least from a quantitative point of view, the low efficiency of the impaired ones, through a compensatory mechanism [201]. In this scenario, PGC-1α represents the main regulator in mitochondrial biogenesis (Table 3). Its reduction has been related to PD onset, because it could indirectly induce alpha-synuclein oligomerization: in fact, it has been proven that PGC-1α restoration can reduce alpha-synuclein toxicity both in cell culture and in transgenic animals [202]. To further maintain mitochondrial functionality, the prevention of the formation of the mitochondrial permeability transition pore (mPTP) could be a good strategy: it can avoid mitochondria swelling, outer membrane disruption, and releasing of apoptotic factors [203], finally resulting in neuronal death. According to this, Olesoxime is a molecule that can bind two proteins of the outer mitochondrial membrane (TSPO and VDAC), in this way preventing the mPTP opening [204]. Following different preclinical investigations in which it provided lifespan prolongation in transgenic mouse models [204], this drug has been considered a promising compound for ALS and SMA patients but, even if clinical trials on ALS have failed [205], a Phase 2 trial on SMA patients has shown an amelioration in motor functions (NCT01302600) that encouraged further investigations [206]. Moreover, some studies on PD and HD models have confirmed both in vitro and in vivo its key role in mitochondria stability resulting in a mitigation of cognitive and behavioral impairment [206].
Table 3.
Compounds acting on mitochondrial biogenesis and permeability. The table shows the promising experimental results obtained in vitro and/or in vivo after the administration of PGC-1α (to enhance mitochondria biogenesis) and Olesoxime (to regulate mitochondria permeability). The latter one already gave promising results in SMA patients. For clinical trial ID we referred to https://clinicaltrials.gov.
4.3. Mitochondrial Bioenergetics
Neurotransmission and metabolism require high energy levels, and mitochondria are the key organelles for ATP production; therefore, acting on impaired mitochondria with compounds that may support neurons in carrying on their activity could be a good strategy in neurodegenerative diseases treatment (Table 4).
Table 4.
Compounds acting on mitochondrial bioenergetics. The table lists compounds involved in mitochondrial bioenergetics enhancement, providing an overview of preclinical and clinical studies. For clinical trial ID we referred to https://clinicaltrials.gov.
As exhaustively explained by Carrera-Juliá et al. [151], NAD+ is a compound involved in many metabolic pathways that are related to neuroprotection and, in turn, its intake could be considered a promising approach to treatment of neurodegenerative diseases. Indeed, acting in glycolysis, Krebs cycle, and oxidative phosphorylation, it can contribute to preserve cognitive functions, representing a promising compound for AD treatment [139,207]. Furthermore, the role of Nicotinamide Riboside, a NAD+ precursor, has been investigated in vivo in AD models and in vitro in PD and ALS mutated cells, providing a wide range of positive outcomes, from mitochondrial biogenesis enhancement to ROS production inhibition, reflected in cognitive improvement due to an increase of synaptic plasticity for what concerns AD mice [64,151,208].
Similarly, triheptanoin can provide several metabolites that powers Krebs cycle, both in CNS and in peripheral tissues [209]. Therefore, its anaplerotic role has been investigated at early stages of HD: however, until now, only one clinical study has confirmed such efficacy, showing an improvement in patients’ brain metabolic profiles (NCT01882062; [210]). Furthermore, thanks to its function, triheptanoin significantly delayed motor neuron loss and motor symptom onset in SOD1G93A mice [211]: for this reason, it has been also considered as a promising treatment to slow ALS progression.
4.4. Compounds Targeting Multiple Mitochondrial Dysfunctions
Besides all the compounds listed until now, there are several other molecules which may act at the same time on many pathways linked to mitochondrial dysfunctions (for a general overview see Table 5). For example, phenylpropanoids are natural molecules derived from the amino acid l-phenylalanine; among them, resveratrol, curcumin, and flavonoids (including epigallocatechin-gallate, quercetin and wogonin) can (i) regulate mitochondrial biogenesis, enhancing the expression of several activators (i.e., PGC-1α, SIRT1, Nrf1, Nrf2, and TFAM), (ii) improve mitochondrial bioenergetics, (iii) enhance ATP production, and (iv) counteract apoptotic pathways (as described in Kolaj et al. [212]). Furthermore, they are able to prevent oxidative stress (i) quenching free radicals through a direct mechanism [213], (ii) upregulating the expression of antioxidative enzymes via the activation of different signaling pathways (i.e., Nrf2), even increasing GSH production [214,215], and (iii) inhibiting ROS-forming enzymes like XO and NOX [216]. Therefore, all these compounds have been investigated as potential therapeutics to treat mitochondrial dysfunctions characterizing many neurodegenerative diseases. When administered in vitro in AD and ALS cell lines, these compounds can significantly increase cell survival, reducing oxidative stress and restoring MMP [139,151,217,218,219]; likewise, preclinical studies in vivo on AD and ALS models confirmed their neuroprotective effects, improving cognitive functions and delaying motor symptom onset and disease progression, respectively [73,139,151,220]. Furthermore, the first promising results on human clinical trials have appeared: in fact, curcumin was effective in lifespan elongation and in decreasing disease progression in ALS patients, although the low oral bioavailability makes new delivery methods necessary [151]; and resveratrol has shown to be safe, tolerable, and effective in mitigating cognitive decline in clinical trials on AD patients (NCT01504854 [221]; NCT00678431 [222]).
Table 5.
Compounds targeting simultaneously different mitochondrial dysfunctions. The table lists the molecules that have been tested in preclinical studies or in clinical trials and that regulate several mitochondrial processes impaired in neurodegenerative diseases. For clinical trial ID we referred to https://clinicaltrials.gov.
Other compounds, derived from drug repurposing studies, are extremely effective in mitigating mitochondrial alterations, although initially employed for different therapeutic purposes and acting on different pathways. Among them, pramipexole, a dopamine receptor agonist approved for PD therapy, seems to have an antioxidant role independent from dopaminergic receptors activation [223]: indeed, it plays a key role in neuroprotection, preventing ROS formation and consequently reducing cell death, as proven on SH-SY5Y cells exposed to MPP+ [157,224]. Moreover, as demonstrated through a patch-clamp experiment on rat liver and brain mitochondria, pramipexole may also inhibit the mPTP opening, preventing mitochondria Ca2+-dependent swelling [225].
Another compound investigated for PD treatment is the Selegiline-analog N-Methyl,N-propynyl-2-phenylethylamine, a MAO-B inhibitor. Besides ameliorating motor impairments of MPTP-treated mice, it was also able to (i) increase complex I activity and UCP-2 expression (reducing oxidative stress) and (ii) to inhibit p53 mitochondrial translocation (maintaining mitochondrial membrane integrity and counteracting apoptosis) [226].
Other molecules, which are not specific for the treatment of neurodegenerative diseases, can counteract mitochondrial alterations, preventing a wide range of altered mechanisms. For instance, 3-N-butylphthalide (NBP), a compound widely employed in the treatment of ischemic stroke in China thanks to its ability in restoring microcirculation [227], can (i) maintain mitochondria permeability and dynamics (preventing mitochondria swelling), (ii) improve mitochondria bioenergetics (increasing complex IV activity), (iii) prevent cytochrome c release and caspase-dependent apoptosis, and (iv) counteract ROS production. Indeed, most of these effects have been observed on in vitro and in vivo PD models, determining neuroprotection and behavioral improvement [228,229] and a randomized controlled trial has demonstrated NBP efficacy in improving motor symptom and sleep quality of PD patients [230]. Further studies on cellular models of AD have confirmed neuroprotection and this positive outcome has been translated also in vivo determining cognitive enhancement [228]; furthermore, increased lifespan and motor performances amelioration have been observed even in ALS mice thanks to a mitigation of motor neuron loss [228].
Finally, several preclinical trials have been performed on ursodeoxycholic acid and tauroursodeoxycholic, two drugs currently approved for the treatment of primary biliary sclerosis [231]. As described by Ackerman and Gerhard [232], although their application in neurodegenerative diseases has to be further investigated, these molecules can (i) exert an antioxidant activity, (ii) stabilize mitochondrial membrane potential, and (iii) downregulate apoptotic markers. Indeed, in vitro they have confirmed their role in apoptosis inhibition and membrane stabilization and in vivo studies have shown behavioral improvement in AD, PD, HD, and ALS models [157,233,234,235]. Nevertheless, at present, positive outcomes in clinical trials have been obtained only in ALS subjects providing a delay in the progression of the pathology (NCT00877604 [236,237]).
5. Conclusions
Besides the recent development of therapies for SMA [238], which still need to be followed for the long-term response, there is no cure for the neurodegenerative diseases that we described in the review. Decades after the discovery of the main pathways and genes involved in the pathophysiological mechanisms of many neurodegenerative processes, researchers and pharmaceutical companies are still investigating a treatment to cure AD, PD, HD, and ALS. We believe that the key for finding new effective targets should be to focus on earlier-phenomena occurring, possibly, before evident degeneration and which could be directed not only to the CNS but also to the PNS and peripheral tissues. Mitochondrial alterations are one of these early and diffuse events. Indeed, mitochondrial dysfunctions appear precociously in neurodegeneration and their accumulation trigger or aggravate the degenerative cascade affecting neuronal tissue, MNs, blood cells, fibroblasts, NMJs, and skeletal muscles. Moreover, mitochondrial dysfunctions represent a common feature in neurodegeneration and should be approached as the crosstalk between AD, PD, HD, ALS, and SMA.
Recent therapeutic strategies ranging from natural or synthetic compounds to stem cell therapy, are targeting mitochondrial pathways which are dysfunctional in the early phases of neurodegeneration. As we described above, many of them seem promising.
Looking forward, we expect that some of these targets of mitochondrial-directed therapies could become also biomarkers of neurodegeneration. On the one hand they could be useful to monitor the progression of the disease, on the other hand they could be an effective read-out to test treatment efficacy. The greatest advantage for both the daily practice of clinicians and for the health systems would be the possibility to validate a diagnosis with a non-invasive and cheap method instead of a time, money, and energy consuming long battery of tests. Moreover, the possibility to develop simple and non-invasive tests i.e., from patients’ fluids, would be much more helpful for the amelioration of patients’ motivation and compliance.
Author Contributions
Conceptualization, original draft preparation, writing, figures and revision S.S.; writing and tables A.C.; critical editing of the entire review M.B. and A.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by by Ministero dell’Istruzione dell’Università e della Ricerca MIUR project “Dipartimenti di Eccellenza 2018–2022” to Department of Neuroscience “Rita Levi Montalcini, by Girotondo/ONLUS and SMArathon-ONLUS foundations granted to AV, and by Fondazione CRT “Richieste Ordinarie 2017” (RF 2017.2052) granted to MB.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Aco2 | Aconitase 2 |
| CAT | Catalase |
| Cyt C | Cytochrome C |
| DAMPs | Damage-associated molecular patterns |
| DDW | Deuterium depleted water |
| ETC | Electron transport chain |
| Fis1 | Mitochondrial fission 1 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GPX | Glutathione peroxidase |
| NLRP3 | NOD-, LRR- and pyrin domain-containing protein 3pyrin domain-containing 3 |
| NOX | NADPH oxidase |
| Nrf1 | Nuclear respiratory factor 1 |
| OPA1 | Protein optic atrophy 1 |
| PBMCs | Peripheral blood mononuclear cells |
| PGK1 | Phosphoglycerate kinase 1 |
| PINK1 | PTEN-induced kinase 1 |
| SIRT1 | Sirtuin1 |
| SNpc | Substantia nigra pars compacta |
| TCA | Tricarboxylic acid |
| TDP43 | TAR DNA-binding protein 43 |
| TFAM | Mitochondrial transcription factor A |
| TPP+ | Alkyl-triphenylphosphonium |
| TSPO | Translocator protein |
| UCP-2 | Mitochondrial uncoupling protein 2 |
| VDAC | Voltage-dependent ion channel |
| XO | Xanthine oxidase |
References
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.A.; Tkachuk, A.N.; Shtengel, G.; Kopek, B.G.; Bogenhagen, D.F.; Hess, H.F.; Clayton, D.A. Superresolution fluorescence imaging of mitochondrial nucleoids reveals their spatial range, limits, and membrane interaction. Mol. Cell Biol. 2011, 31, 4994–5010. [Google Scholar] [CrossRef] [PubMed]
- Steele, H.E.; Horvath, R.; Lyon, J.J.; Chinnery, P.F. Monitoring clinical progression with mitochondrial disease biomarkers. Brain 2017, 140, 2530–2540. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C. Mitochondrial fusion and fission in mammals. Annu. Rev. Cell Dev. Biol. 2006, 22, 79–99. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 5th ed.; Oxford University Press: Oxford, UK, 2015; Volume xxxviii, 905p. [Google Scholar]
- Shadel, G.S.; Horvath, T.L. Mitochondrial ROS signaling in organismal homeostasis. Cell 2015, 163, 560–569. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Vesce, S.; Kirk, L.; Chalmers, S. Interactions between mitochondrial bioenergetics and cytoplasmic calcium in cultured cerebellar granule cells. Cell Calcium 2003, 34, 407–424. [Google Scholar] [CrossRef]
- Giacomello, M.; Drago, I.; Pizzo, P.; Pozzan, T. Mitochondrial Ca2+ as a key regulator of cell life and death. Cell Death Differ. 2007, 14, 1267–1274. [Google Scholar] [CrossRef]
- Drago, I.; De Stefani, D.; Rizzuto, R.; Pozzan, T. Mitochondrial Ca2+ uptake contributes to buffering cytoplasmic Ca2+ peaks in cardiomyocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 12986–12991. [Google Scholar] [CrossRef]
- Wang, X. The expanding role of mitochondria in apoptosis. Genes Dev. 2001, 15, 2922–2933. [Google Scholar]
- Gahl, R.F.; Dwivedi, P.; Tjandra, N. Bcl-2 proteins bid and bax form a network to permeabilize the mitochondria at the onset of apoptosis. Cell Death Dis. 2016, 7, e2424. [Google Scholar] [CrossRef]
- Youle, R.J.; van der Bliek, A.M. Mitochondrial fission, fusion, and stress. Science 2012, 337, 1062–1065. [Google Scholar] [CrossRef] [PubMed]
- Egan, D.F.; Shackelford, D.B.; Mihaylova, M.M.; Gelino, S.; Kohnz, R.A.; Mair, W.; Vasquez, D.S.; Joshi, A.; Gwinn, D.M.; Taylor, R.; et al. Phosphorylation of ULK1 (hATG1) by AMP-activated protein kinase connects energy sensing to mitophagy. Science 2011, 331, 456–461. [Google Scholar] [CrossRef]
- Lemasters, J.J. Selective mitochondrial autophagy, or mitophagy, as a targeted defense against oxidative stress, mitochondrial dysfunction, and aging. Rejuvenation Res. 2005, 8, 3–5. [Google Scholar] [CrossRef]
- Palikaras, K.; Lionaki, E.; Tavernarakis, N. Mechanisms of mitophagy in cellular homeostasis, physiology and pathology. Nat. Cell Biol. 2018, 20, 1013–1022. [Google Scholar] [CrossRef] [PubMed]
- Kirienko, N.V.; Ausubel, F.M.; Ruvkun, G. Mitophagy confers resistance to siderophore-mediated killing by Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2015, 112, 1821–1826. [Google Scholar] [CrossRef] [PubMed]
- West, A.P.; Khoury-Hanold, W.; Staron, M.; Tal, M.C.; Pineda, C.M.; Lang, S.M.; Bestwick, M.; Duguay, B.A.; Raimundo, N.; MacDuff, D.A.; et al. Mitochondrial DNA stress primes the antiviral innate immune response. Nature 2015, 520, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Gorman, G.S.; Chinnery, P.F.; DiMauro, S.; Hirano, M.; Koga, Y.; McFarland, R.; Suomalainen, A.; Thorburn, D.R.; Zeviani, M.; Turnbull, D.M. Mitochondrial diseases. Nat. Rev. Dis. Primers 2016, 2, 16080. [Google Scholar] [CrossRef]
- Kim, J.A.; Wei, Y.; Sowers, J.R. Role of mitochondrial dysfunction in insulin resistance. Circ. Res. 2008, 102, 401–414. [Google Scholar] [CrossRef]
- Barcelos, I.P.; Troxell, R.M.; Graves, J.S. Mitochondrial dysfunction and multiple sclerosis. Biology 2019, 8, 37. [Google Scholar] [CrossRef]
- Porporato, P.E.; Filigheddu, N.; Pedro, J.M.B.; Kroemer, G.; Galluzzi, L. Mitochondrial metabolism and cancer. Cell Res. 2018, 28, 265–280. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, M.; Jiang, J. Mitochondrial dysfunction in neurodegenerative diseases and drug targets via apoptotic signaling. Mitochondrion 2019, 49, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Kandimalla, R.; Manczak, M.; Yin, X.; Wang, R.; Reddy, P.H. Hippocampal phosphorylated tau induced cognitive decline, dendritic spine loss and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Manczak, M.; Reddy, P.H. Mitochondria-targeted molecules MitoQ and SS31 reduce mutant huntingtin-induced mitochondrial toxicity and synaptic damage in Huntington’s disease. Hum. Mol. Genet. 2016, 25, 1739–1753. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, T.L. Mitochondrial trafficking in neurons. Cold Spring Harb. Perspect. Biol. 2013, 5, a011304. [Google Scholar] [CrossRef]
- Somlyai, G.; Jancsó, G.; Jákli, G.; Vass, K.; Barna, B.; Lakics, V.; Gaál, T. Naturally occurring deuterium is essential for the normal growth rate of cells. FEBS Lett. 1993, 317, 1–4. [Google Scholar] [CrossRef]
- Olgun, A. Biological effects of deuteration: ATP synthase as an example. Theor. Biol. Med. Model. 2007, 4, 9. [Google Scholar] [CrossRef]
- Mladin, C.; Ciobica, A.; Lefter, R.; Popescu, A.; Bild, W. Deuterium-depleted water has stimulating effects on long-term memory in rats. Neurosci. Lett. 2014, 583, 154–158. [Google Scholar] [CrossRef]
- Basov, A.; Fedulova, L.; Baryshev, M.; Dzhimak, S. Deuterium-depleted water infulence on the isotope 2H/1H regulation in body and individual adaptation. Nutrients 2019, 11, 1903. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, N.; Lu, B. Mechanisms and roles of mitophagy in neurodegenerative diseases. CNS Neurosci. Ther. 2019, 25, 859–875. [Google Scholar] [CrossRef]
- Scott, S.V.; Klionsky, D.J. Delivery of proteins and organelles to the vacuole from the cytoplasm. Curr. Opin. Cell Biol. 1998, 10, 523–529. [Google Scholar] [CrossRef]
- Berezhnov, A.V.; Soutar, M.P.M.; Fedotova, E.I.; Frolova, M.S.; Plun-Favreau, H.; Zinchenko, V.P.; Abramov, A.Y. Intracellular pH modulates autophagy and mitophagy. J. Biol. Chem. 2016, 291, 8701–8708. [Google Scholar] [CrossRef] [PubMed]
- Missiroli, S.; Genovese, I.; Perrone, M.; Vezzani, B.; Vitto, V.A.M.; Giorgi, C. The role of mitochondria in inflammation: From cancer to neurodegenerative disorders. J. Clin. Med. 2020, 9, 740. [Google Scholar] [CrossRef] [PubMed]
- Mathew, A.; Lindsley, T.A.; Sheridan, A.; Bhoiwala, D.L.; Hushmendy, S.F.; Yager, E.J.; Ruggiero, E.A.; Crawford, D.R. Degraded mitochondrial DNA is a newly identified subtype of the damage associated molecular pattern (DAMP) family and possible trigger of neurodegeneration. J. Alzheimers Dis. 2012, 30, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Kepp, O.; Galluzzi, L.; Kroemer, G. Inflammasomes in carcinogenesis and anticancer immune responses. Nat. Immunol. 2012, 13, 343–351. [Google Scholar] [CrossRef]
- Schroder, K.; Zhou, R.; Tschopp, J. The NLRP3 inflammasome: A sensor for metabolic danger? Science 2010, 327, 296–300. [Google Scholar] [CrossRef]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef]
- Park, S.; Won, J.H.; Hwang, I.; Hong, S.; Lee, H.K.; Yu, J.W. Defective mitochondrial fission augments NLRP3 inflammasome activation. Sci. Rep. 2015, 5, 15489. [Google Scholar] [CrossRef]
- Schon, E.A.; Manfredi, G. Neuronal degeneration and mitochondrial dysfunction. J. Clin. Invest. 2003, 111, 303–312. [Google Scholar] [CrossRef]
- Trushina, E.; McMurray, C.T. Oxidative stress and mitochondrial dysfunction in neurodegenerative diseases. Neuroscience 2007, 145, 1233–1248. [Google Scholar] [CrossRef]
- Ferri, C.P.; Prince, M.; Brayne, C.; Brodaty, H.; Fratiglioni, L.; Ganguli, M.; Hall, K.; Hasegawa, K.; Hendrie, H.; Huang, Y.; et al. Global prevalence of dementia: A Delphi consensus study. Lancet 2005, 366, 2112–2117. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Annaert, W.; De Strooper, B. A cell biological perspective on Alzheimer’s disease. Annu. Rev. Cell Dev. Biol. 2002, 18, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Herholz, K. Use of FDG PET as an imaging biomarker in clinical trials of Alzheimer’s disease. Biomark. Med. 2012, 6, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D.; Uryu, K.; Leight, S.; Trojanoswki, J.Q.; Lee, V.M. Increased lipid peroxidation precedes amyloid plaque formation in an animal model of Alzheimer amyloidosis. J. Neurosci. 2001, 21, 4183–4187. [Google Scholar] [CrossRef]
- Medeiros, R.; LaFerla, F.M. Astrocytes: Conductors of the Alzheimer disease neuroinflammatory symphony. Exp. Neurol. 2013, 239, 133–138. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, C. Decoding Alzheimer’s disease from perturbed cerebral glucose metabolism: Implications for diagnostic and therapeutic strategies. Prog. Neurobiol. 2013, 108, 21–43. [Google Scholar] [CrossRef]
- Hroudová, J.; Singh, N.; Fišar, Z. Mitochondrial dysfunctions in neurodegenerative diseases: Relevance to Alzheimer’s disease. Biomed. Res. Int. 2014, 2014, 175062. [Google Scholar] [CrossRef]
- Carvalho, C.; Correia, S.C.; Santos, R.X.; Cardoso, S.; Moreira, P.I.; Clark, T.A.; Zhu, X.; Smith, M.A.; Perry, G. Role of mitochondrial-mediated signaling pathways in Alzheimer disease and hypoxia. J. Bioenerg. Biomembr. 2009, 41, 433–440. [Google Scholar] [CrossRef]
- Kerr, J.S.; Adriaanse, B.A.; Greig, N.H.; Mattson, M.P.; Cader, M.Z.; Bohr, V.A.; Fang, E.F. Mitophagy and Alzheimer’s disease: Cellular and molecular mechanisms. Trends Neurosci. 2017, 40, 151–166. [Google Scholar] [CrossRef]
- Lanni, C.; Nardinocchi, L.; Puca, R.; Stanga, S.; Uberti, D.; Memo, M.; Govoni, S.; D’Orazi, G.; Racchi, M. Homeodomain interacting protein kinase 2: A target for Alzheimer’s beta amyloid leading to misfolded p53 and inappropriate cell survival. PLoS ONE 2010, 5, e10171. [Google Scholar] [CrossRef]
- Pérez, M.J.; Ponce, D.P.; Aranguiz, A.; Behrens, M.I.; Quintanilla, R.A. Mitochondrial permeability transition pore contributes to mitochondrial dysfunction in fibroblasts of patients with sporadic Alzheimer’s disease. Redox Biol. 2018, 19, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Stanga, S.; Zanou, N.; Audouard, E.; Tasiaux, B.; Contino, S.; Vandermeulen, G.; René, F.; Loeffler, J.P.; Clotman, F.; Gailly, P.; et al. APP-dependent glial cell line-derived neurotrophic factor gene expression drives neuromuscular junction formation. FASEB J. 2016, 30, 1696–1711. [Google Scholar] [CrossRef]
- Contino, S.; Porporato, P.E.; Bird, M.; Marinangeli, C.; Opsomer, R.; Sonveaux, P.; Bontemps, F.; Dewachter, I.; Octave, J.N.; Bertrand, L.; et al. Presenilin 2-dependent maintenance of mitochondrial oxidative capacity and morphology. Front. Physiol. 2017, 8, 796. [Google Scholar] [CrossRef]
- Stanga, S.; Brambilla, L.; Tasiaux, B.; Dang, A.H.; Ivanoiu, A.; Octave, J.N.; Rossi, D.; van Pesch, V.; Kienlen-Campard, P. A Role for GDNF and soluble APP as biomarkers of amyotrophic lateral sclerosis pathophysiology. Front. Neurol. 2018, 9, 384. [Google Scholar] [CrossRef] [PubMed]
- Polymeropoulos, M.H.; Lavedan, C.; Leroy, E.; Ide, S.E.; Dehejia, A.; Dutra, A.; Pike, B.; Root, H.; Rubenstein, J.; Boyer, R.; et al. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science 1997, 276, 2045–2047. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Lewis, P.; Revesz, T.; Lees, A.; Paisan-Ruiz, C. The genetics of Parkinson’s syndromes: A critical review. Curr. Opin. Genet. Dev. 2009, 19, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Bender, A.; Krishnan, K.J.; Morris, C.M.; Taylor, G.A.; Reeve, A.K.; Perry, R.H.; Jaros, E.; Hersheson, J.S.; Betts, J.; Klopstock, T.; et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat. Genet. 2006, 38, 515–517. [Google Scholar] [CrossRef]
- McGeer, P.L.; Itagaki, S.; Boyes, B.E.; McGeer, E.G. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology 1988, 38, 1285–1291. [Google Scholar] [CrossRef]
- Gerhard, A.; Pavese, N.; Hotton, G.; Turkheimer, F.; Es, M.; Hammers, A.; Eggert, K.; Oertel, W.; Banati, R.B.; Brooks, D.J. In vivo imaging of microglial activation with [11C](R)-PK11195 PET in idiopathic Parkinson’s disease. Neurobiol. Dis. 2006, 21, 404–412. [Google Scholar] [CrossRef]
- Langston, J.W.; Ballard, P.; Tetrud, J.W.; Irwin, I. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science 1983, 219, 979–980. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Bose, A.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in induced pluripotent stem cell models of Parkinson’s disease. Eur. J. Neurosci. 2019, 49, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Schöndorf, D.C.; Ivanyuk, D.; Baden, P.; Sanchez-Martinez, A.; De Cicco, S.; Yu, C.; Giunta, I.; Schwarz, L.K.; Di Napoli, G.; Panagiotakopoulou, V.; et al. The NAD+ precursor nicotinamide riboside rescues mitochondrial defects and neuronal loss in iPSC and fly models of Parkinson’s disease. Cell Rep. 2018, 23, 2976–2988. [Google Scholar] [CrossRef] [PubMed]
- Mosley, R.L.; Hutter-Saunders, J.A.; Stone, D.K.; Gendelman, H.E. Inflammation and adaptive immunity in Parkinson’s disease. Cold Spring Harb. Perspect. Med. 2012, 2, a009381. [Google Scholar] [CrossRef]
- Bose, A.; Beal, M.F. Mitochondrial dysfunction in Parkinson’s disease. J. Neurochem. 2016, 139, 216–231. [Google Scholar] [CrossRef]
- Keeney, P.M.; Xie, J.; Capaldi, R.A.; Bennett, J.P. Parkinson’s disease brain mitochondrial complex I has oxidatively damaged subunits and is functionally impaired and misassembled. J. Neurosci. 2006, 26, 5256–5264. [Google Scholar] [CrossRef]
- Perier, C.; Bové, J.; Dehay, B.; Jackson-Lewis, V.; Rabinovitch, P.S.; Przedborski, S.; Vila, M. Apoptosis-inducing factor deficiency sensitizes dopaminergic neurons to Parkinsonian neurotoxins. Ann. Neurol. 2010, 68, 184–192. [Google Scholar]
- Parihar, M.S.; Parihar, A.; Fujita, M.; Hashimoto, M.; Ghafourifar, P. Mitochondrial association of alpha-synuclein causes oxidative stress. Cell Mol. Life Sci. 2008, 65, 1272–1284. [Google Scholar] [CrossRef]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef]
- St-Pierre, J.; Drori, S.; Uldry, M.; Silvaggi, J.M.; Rhee, J.; Jäger, S.; Handschin, C.; Zheng, K.; Lin, J.; Yang, W.; et al. Suppression of reactive oxygen species and neurodegeneration by the PGC-1 transcriptional coactivators. Cell 2006, 127, 397–408. [Google Scholar] [CrossRef]
- Zheng, B.; Liao, Z.; Locascio, J.J.; Lesniak, K.A.; Roderick, S.S.; Watt, M.L.; Eklund, A.C.; Zhang-James, Y.; Kim, P.D.; Hauser, M.A.; et al. PGC-1α, a potential therapeutic target for early intervention in Parkinson’s disease. Sci. Transl. Med. 2010, 2, 52ra73. [Google Scholar] [CrossRef] [PubMed]
- Berthet, A.; Margolis, E.B.; Zhang, J.; Hsieh, I.; Hnasko, T.S.; Ahmad, J.; Edwards, R.H.; Sesaki, H.; Huang, E.J.; Nakamura, K. Loss of mitochondrial fission depletes axonal mitochondria in midbrain dopamine neurons. J. Neurosci. 2014, 34, 14304–14317. [Google Scholar] [CrossRef] [PubMed]
- Scarffe, L.A.; Stevens, D.A.; Dawson, V.L.; Dawson, T.M. Parkin and PINK1: Much more than mitophagy. Trends Neurosci. 2014, 37, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.K.; Beal, M.F. Mitochondrial approaches for neuroprotection. Ann. N. Y. Acad. Sci. 2008, 1147, 395–412. [Google Scholar] [CrossRef] [PubMed]
- Vonsattel, J.P.; Myers, R.H.; Stevens, T.J.; Ferrante, R.J.; Bird, E.D.; Richardson, E.P. Neuropathological classification of Huntington’s disease. J. Neuropathol. Exp. Neurol. 1985, 44, 559–577. [Google Scholar] [CrossRef]
- Reddy, P.H.; Charles, V.; Williams, M.; Miller, G.; Whetsell, W.O.; Tagle, D.A. Transgenic mice expressing mutated full-length HD cDNA: A paradigm for locomotor changes and selective neuronal loss in Huntington’s disease. Philos. Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 1035–1045. [Google Scholar] [CrossRef]
- Li, S.; Li, X.J. Multiple pathways contribute to the pathogenesis of Huntington disease. Mol. Neurodegener. 2006, 1, 19. [Google Scholar] [CrossRef][Green Version]
- Browne, S.E.; Beal, M.F. The energetics of Huntington’s disease. Neurochem. Res. 2004, 29, 531–546. [Google Scholar] [CrossRef]
- Fukui, H.; Moraes, C.T. Extended polyglutamine repeats trigger a feedback loop involving the mitochondrial complex III, the proteasome and huntingtin aggregates. Hum. Mol. Genet. 2007, 16, 783–797. [Google Scholar] [CrossRef]
- Hwang, S.; Disatnik, M.H.; Mochly-Rosen, D. Impaired GAPDH-induced mitophagy contributes to the pathology of Huntington’s disease. EMBO Mol. Med. 2015, 7, 1307–1326. [Google Scholar] [CrossRef]
- Acevedo-Torres, K.; Berríos, L.; Rosario, N.; Dufault, V.; Skatchkov, S.; Eaton, M.J.; Torres-Ramos, C.A.; Ayala-Torres, S. Mitochondrial DNA damage is a hallmark of chemically induced and the R6/2 transgenic model of Huntington’s disease. DNA Repair 2009, 8, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Banoei, M.M.; Houshmand, M.; Panahi, M.S.; Shariati, P.; Rostami, M.; Manshadi, M.D.; Majidizadeh, T. Huntington’s disease and mitochondrial DNA deletions: Event or regular mechanism for mutant huntingtin protein and CAG repeats expansion?! Cell Mol. Neurobiol. 2007, 27, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Crotti, A.; Glass, C.K. The choreography of neuroinflammation in Huntington’s disease. Trends Immunol. 2015, 36, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Duyao, M.P.; Auerbach, A.B.; Ryan, A.; Persichetti, F.; Barnes, G.T.; McNeil, S.M.; Ge, P.; Vonsattel, J.P.; Gusella, J.F.; Joyner, A.L. Inactivation of the mouse Huntington’s disease gene homolog Hdh. Science 1995, 269, 407–410. [Google Scholar] [CrossRef]
- Borovecki, F.; Lovrecic, L.; Zhou, J.; Jeong, H.; Then, F.; Rosas, H.D.; Hersch, S.M.; Hogarth, P.; Bouzou, B.; Jensen, R.V.; et al. Genome-wide expression profiling of human blood reveals biomarkers for Huntington’s disease. Proc. Natl. Acad. Sci. USA 2005, 102, 11023–11028. [Google Scholar] [CrossRef]
- Mihm, M.J.; Amann, D.M.; Schanbacher, B.L.; Altschuld, R.A.; Bauer, J.A.; Hoyt, K.R. Cardiac dysfunction in the R6/2 mouse model of Huntington’s disease. Neurobiol. Dis. 2007, 25, 297–308. [Google Scholar] [CrossRef]
- Farrer, L.A. Diabetes mellitus in Huntington disease. Clin. Genet. 1985, 27, 62–67. [Google Scholar] [CrossRef]
- Busse, M.E.; Hughes, G.; Wiles, C.M.; Rosser, A.E. Use of hand-held dynamometry in the evaluation of lower limb muscle strength in people with Huntington’s disease. J. Neurol. 2008, 255, 1534–1540. [Google Scholar] [CrossRef]
- Jędrak, P.; Mozolewski, P.; Węgrzyn, G.; Więckowski, M.R. Mitochondrial alterations accompanied by oxidative stress conditions in skin fibroblasts of Huntington’s disease patients. Metab. Brain Dis. 2018, 33, 2005–2017. [Google Scholar] [CrossRef]
- Chen, C.M.; Wu, Y.R.; Chang, K.H. Altered aconitase 2 activity in Huntington’s disease peripheral blood cells and mouse model striatum. Int. J. Mol. Sci. 2017, 18, 2480. [Google Scholar] [CrossRef]
- Turner, M.R.; Bowser, R.; Bruijn, L.; Dupuis, L.; Ludolph, A.; McGrath, M.; Manfredi, G.; Maragakis, N.; Miller, R.G.; Pullman, S.L.; et al. Mechanisms, models and biomarkers in amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Frontotemporal Degener. 2013, 14, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Robberecht, W.; Philips, T. The changing scene of amyotrophic lateral sclerosis. Nat. Rev. Neurosci. 2013, 14, 248–264. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.; Rogelj, B.; Hortobágyi, T.; De Vos, K.J.; Nishimura, A.L.; Sreedharan, J.; Hu, X.; Smith, B.; Ruddy, D.; Wright, P.; et al. Mutations in FUS, an RNA processing protein, cause familial amyotrophic lateral sclerosis type 6. Science 2009, 323, 1208–1211. [Google Scholar] [CrossRef]
- Sreedharan, J.; Blair, I.P.; Tripathi, V.B.; Hu, X.; Vance, C.; Rogelj, B.; Ackerley, S.; Durnall, J.C.; Williams, K.L.; Buratti, E.; et al. TDP-43 mutations in familial and sporadic amyotrophic lateral sclerosis. Science 2008, 319, 1668–1672. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, M.; Ferri, A.; Carrì, M.T. Amyotrophic lateral sclerosis: From current developments in the laboratory to clinical implications. Antioxid. Redox Signal. 2008, 10, 405–443. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, D.; Cordera, S.; Cavalla, P.; Migheli, A. Reactive astrogliosis of the spinal cord in amyotrophic lateral sclerosis. J. Neurol. Sci. 1996, 139, 27–33. [Google Scholar] [CrossRef]
- Kawamata, T.; Akiyama, H.; Yamada, T.; McGeer, P.L. Immunologic reactions in amyotrophic lateral sclerosis brain and spinal cord tissue. Am. J. Pathol. 1992, 140, 691–707. [Google Scholar]
- Sasaki, S.; Iwata, M. Mitochondrial alterations in the spinal cord of patients with sporadic amyotrophic lateral sclerosis. J. Neuropathol. Exp. Neurol. 2007, 66, 10–16. [Google Scholar] [CrossRef]
- Smith, E.F.; Shaw, P.J.; De Vos, K.J. The role of mitochondria in amyotrophic lateral sclerosis. Neurosci. Lett. 2019, 710, 132933. [Google Scholar] [CrossRef]
- Shi, P.; Gal, J.; Kwinter, D.M.; Liu, X.; Zhu, H. Mitochondrial dysfunction in amyotrophic lateral sclerosis. Biochim. Biophys. Acta 2010, 1802, 45–51. [Google Scholar] [CrossRef]
- Pasinelli, P.; Belford, M.E.; Lennon, N.; Bacskai, B.J.; Hyman, B.T.; Trotti, D.; Brown, R.H. Amyotrophic lateral sclerosis-associated SOD1 mutant proteins bind and aggregate with Bcl-2 in spinal cord mitochondria. Neuron 2004, 43, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Dupuis, L.; Gonzalez de Aguilar, J.L.; Oudart, H.; de Tapia, M.; Barbeito, L.; Loeffler, J.P. Mitochondria in amyotrophic lateral sclerosis: A trigger and a target. Neurodegener. Dis. 2004, 1, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Pollari, E.; Goldsteins, G.; Bart, G.; Koistinaho, J.; Giniatullin, R. The role of oxidative stress in degeneration of the neuromuscular junction in amyotrophic lateral sclerosis. Front. Cell Neurosci. 2014, 8, 131. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Pasinelli, P.; Trotti, D. Role of mitochondria in mutant SOD1 linked amyotrophic lateral sclerosis. Biochim. Biophys. Acta 2014, 1842, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Elfawy, H.A.; Das, B. Crosstalk between mitochondrial dysfunction, oxidative stress, and age related neurodegenerative disease: Etiologies and therapeutic strategies. Life Sci. 2019, 218, 165–184. [Google Scholar] [CrossRef] [PubMed]
- Lorson, C.L.; Hahnen, E.; Androphy, E.J.; Wirth, B. A single nucleotide in the SMN gene regulates splicing and is responsible for spinal muscular atrophy. Proc. Natl. Acad. Sci. USA 1999, 96, 6307–6311. [Google Scholar] [CrossRef]
- Ripolone, M.; Ronchi, D.; Violano, R.; Vallejo, D.; Fagiolari, G.; Barca, E.; Lucchini, V.; Colombo, I.; Villa, L.; Berardinelli, A.; et al. Impaired muscle mitochondrial biogenesis and myogenesis in spinal muscular atrophy. JAMA Neurol. 2015, 72, 666–675. [Google Scholar] [CrossRef]
- Vaidya, S.; Boes, S. Correction to: Measuring quality of life in children with spinal muscular atrophy: A systematic literature review. Qual. Life Res. 2018, 27, 3095. [Google Scholar] [CrossRef]
- Fuller, H.R.; Gillingwater, T.H.; Wishart, T.M. Commonality amid diversity: Multi-study proteomic identification of conserved disease mechanisms in spinal muscular atrophy. Neuromuscul. Disord. 2016, 26, 560–569. [Google Scholar] [CrossRef]
- Boyd, P.J.; Tu, W.Y.; Shorrock, H.K.; Groen, E.J.N.; Carter, R.N.; Powis, R.A.; Thomson, S.R.; Thomson, D.; Graham, L.C.; Motyl, A.A.L.; et al. Bioenergetic status modulates motor neuron vulnerability and pathogenesis in a zebrafish model of spinal muscular atrophy. PLoS Genet. 2017, 13, e1006744. [Google Scholar] [CrossRef]
- Pagliardini, S.; Giavazzi, A.; Setola, V.; Lizier, C.; Di Luca, M.; DeBiasi, S.; Battaglia, G. Subcellular localization and axonal transport of the survival motor neuron (SMN) protein in the developing rat spinal cord. Hum. Mol. Genet. 2000, 9, 47–56. [Google Scholar] [CrossRef]
- Lotti, F.; Imlach, W.L.; Saieva, L.; Beck, E.S.; Hao, L.T.; Li, D.K.; Jiao, W.; Mentis, G.Z.; Beattie, C.E.; McCabe, B.D.; et al. An SMN-dependent U12 splicing event essential for motor circuit function. Cell 2012, 151, 440–454. [Google Scholar] [CrossRef] [PubMed]
- Boido, M.; Vercelli, A. Neuromuscular junctions as key contributors and therapeutic targets in spinal muscular atrophy. Front. Neuroanat. 2016, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, D.; Le Verche, V.; Jacquier, A.; Ikiz, B.; Przedborski, S.; Re, D.B. Inflammation in ALS and SMA: Sorting out the good from the evil. Neurobiol. Dis. 2010, 37, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Simone, C.; Ramirez, A.; Bucchia, M.; Rinchetti, P.; Rideout, H.; Papadimitriou, D.; Re, D.B.; Corti, S. Is spinal muscular atrophy a disease of the motor neurons only: Pathogenesis and therapeutic implications? Cell Mol. Life Sci. 2016, 73, 1003–1020. [Google Scholar] [CrossRef] [PubMed]
- Dubowitz, V. Muscle disorders in childhood. Major Probl. Clin. Pediatr. 1978, 16, 1–282. [Google Scholar]
- Xu, C.C.; Denton, K.R.; Wang, Z.B.; Zhang, X.; Li, X.J. Abnormal mitochondrial transport and morphology as early pathological changes in human models of spinal muscular atrophy. Dis. Model. Mech. 2016, 9, 39–49. [Google Scholar] [CrossRef]
- Borgia, D.; Malena, A.; Spinazzi, M.; Desbats, M.A.; Salviati, L.; Russell, A.P.; Miotto, G.; Tosatto, L.; Pegoraro, E.; Sorarù, G.; et al. Increased mitophagy in the skeletal muscle of spinal and bulbar muscular atrophy patients. Hum. Mol. Genet. 2017, 26, 1087–1103. [Google Scholar] [CrossRef]
- Acsadi, G.; Lee, I.; Li, X.; Khaidakov, M.; Pecinova, A.; Parker, G.C.; Hüttemann, M. Mitochondrial dysfunction in a neural cell model of spinal muscular atrophy. J. Neurosci. Res. 2009, 87, 2748–2756. [Google Scholar] [CrossRef]
- Stifani, N. Motor neurons and the generation of spinal motor neuron diversity. Front. Cell Neurosci. 2014, 8, 293. [Google Scholar] [CrossRef]
- Wijngaarde, C.A.; Blank, A.C.; Stam, M.; Wadman, R.I.; van den Berg, L.H.; van der Pol, W.L. Cardiac pathology in spinal muscular atrophy: A systematic review. Orphanet. J. Rare Dis. 2017, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Fortune, M.D.; Guo, H.; Burren, O.; Schofield, E.; Walker, N.M.; Ban, M.; Sawcer, S.J.; Bowes, J.; Worthington, J.; Barton, A.; et al. Statistical colocalization of genetic risk variants for related autoimmune diseases in the context of common controls. Nat. Genet. 2015, 47, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Brown, B.C.; Ye, C.J.; Price, A.L.; Zaitlen, N.; Asian Genetic Epidemiology Network Type 2 Diabetes Consortium. Transethnic genetic-correlation estimates from summary statistics. Am. J. Hum. Genet. 2016, 99, 76–88. [Google Scholar] [CrossRef]
- Pickrell, J.K.; Berisa, T.; Liu, J.Z.; Ségurel, L.; Tung, J.Y.; Hinds, D.A. Detection and interpretation of shared genetic influences on 42 human traits. Nat. Genet. 2016, 48, 709–717. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Chan, K.H.K.; Zhang, G.; Huan, T.; Kurt, Z.; Zhao, Y.; Codoni, V.; Trégouët, D.A.; Yang, J.; Wilson, J.G.; et al. Shared genetic regulatory networks for cardiovascular disease and type 2 diabetes in multiple populations of diverse ethnicities in the United States. PLoS Genet. 2017, 13, e1007040. [Google Scholar] [CrossRef] [PubMed]
- Arneson, D.; Zhang, Y.; Yang, X.; Narayanan, M. Shared mechanisms among neurodegenerative diseases: From genetic factors to gene networks. J. Genet. 2018, 97, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.C.; Lockwood, A.H.; Sonawane, B.R. Neurodegenerative diseases: An overview of environmental risk factors. Environ. Health Perspect. 2005, 113, 1250–1256. [Google Scholar] [CrossRef]
- Myhre, O.; Utkilen, H.; Duale, N.; Brunborg, G.; Hofer, T. Metal dyshomeostasis and inflammation in Alzheimer’s and Parkinson’s diseases: Possible impact of environmental exposures. Oxid. Med. Cell Longev. 2013, 2013, 726954. [Google Scholar] [CrossRef]
- Cutler, N.R.; Sramek, J.J. Review of the next generation of Alzheimer’s disease therapeutics: Challenges for drug development. Prog. Neuropsychopharmacol. Biol. Psychiatry 2001, 25, 27–57. [Google Scholar] [CrossRef]
- Pogačnik, L.; Ota, A.; Ulrih, N.P. An overview of crucial dietary substances and their modes of action for prevention of neurodegenerative diseases. Cells 2020, 9, 576. [Google Scholar] [CrossRef]
- Wilkins, H.M.; Morris, J.K. New therapeutics to modulate mitochondrial function in neurodegenerative disorders. Curr. Pharm. Des. 2017, 23, 731–752. [Google Scholar] [CrossRef] [PubMed]
- Hoy, S.M. Nusinersen: First global approval. Drugs 2017, 77, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Potkin, K.T.; Potkin, S.G. New directions in therapeutics for Huntington disease. Future Neurol. 2018, 13, 101–121. [Google Scholar] [CrossRef] [PubMed]
- Atri, A. Current and future treatments in Alzheimer’s disease. Semin. Neurol. 2019, 39, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Hoy, S.M. Onasemnogene abeparvovec: First global approval. Drugs 2019, 79, 1255–1262. [Google Scholar] [CrossRef]
- Mejzini, R.; Flynn, L.L.; Pitout, I.L.; Fletcher, S.; Wilton, S.D.; Akkari, P.A. ALS genetics, mechanisms, and therapeutics: Where are we now? Front. Neurosci. 2019, 13, 1310. [Google Scholar] [CrossRef]
- Reich, S.G.; Savitt, J.M. Parkinson’s disease. Med. Clin. North. Am. 2019, 103, 337–350. [Google Scholar] [CrossRef]
- Cenini, G.; Voos, W. Mitochondria as potential targets in Alzheimer disease therapy: An update. Front. Pharmacol. 2019, 10, 902. [Google Scholar] [CrossRef]
- Iturria-Medina, Y.; Carbonell, F.M.; Sotero, R.C.; Chouinard-Decorte, F.; Evans, A.C.; Alzheimer’s Disease Neuroimaging Initiative. Multifactorial causal model of brain (dis)organization and therapeutic intervention: Application to Alzheimer’s disease. Neuroimage 2017, 152, 60–77. [Google Scholar] [CrossRef]
- Veitch, D.P.; Weiner, M.W.; Aisen, P.S.; Beckett, L.A.; Cairns, N.J.; Green, R.C.; Harvey, D.; Jack, C.R.; Jagust, W.; Morris, J.C.; et al. Understanding disease progression and improving Alzheimer’s disease clinical trials: Recent highlights from the Alzheimer’s disease neuroimaging initiative. Alzheimers Dement. 2019, 15, 106–152. [Google Scholar] [CrossRef]
- Moss, D.E.; Perez, R.G.; Kobayashi, H. Cholinesterase inhibitor therapy in Alzheimer’s disease: The limits and tolerability of irreversible CNS-selective acetylcholinesterase inhibition in primates. J. Alzheimers Dis. 2017, 55, 1285–1294. [Google Scholar] [CrossRef]
- Folch, J.; Busquets, O.; Ettcheto, M.; Sánchez-López, E.; Castro-Torres, R.D.; Verdaguer, E.; Garcia, M.L.; Olloquequi, J.; Casadesús, G.; Beas-Zarate, C.; et al. Memantine for the treatment of dementia: A review on its current and future applications. J. Alzheimers Dis. 2018, 62, 1223–1240. [Google Scholar] [CrossRef]
- Sharma, K. Cholinesterase inhibitors as Alzheimer’s therapeutics (review). Mol. Med. Rep. 2019, 20, 1479–1487. [Google Scholar] [CrossRef]
- Rascol, O.; Payoux, P.; Ory, F.; Ferreira, J.J.; Brefel-Courbon, C.; Montastruc, J.L. Limitations of current Parkinson’s disease therapy. Ann. Neurol 2003, 53, S3–S12. [Google Scholar] [CrossRef]
- Kong, M.; Ba, M.; Ren, C.; Yu, L.; Dong, S.; Yu, G.; Liang, H. An updated meta-analysis of amantadine for treating dyskinesia in Parkinson’s disease. Oncotarget 2017, 8, 57316–57326. [Google Scholar] [CrossRef] [PubMed]
- Khairoalsindi, O.A.; Abuzinadah, A.R. Maximizing the survival of amyotrophic lateral sclerosis patients: Current perspectives. Neurol. Res. Int. 2018, 2018, 6534150. [Google Scholar] [CrossRef] [PubMed]
- Dangouloff, T.; Servais, L. Clinical evidence supporting early treatment of patients with spinal muscular atrophy: Current perspectives. Ther. Clin. Risk Manag. 2019, 15, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative stress: Harms and benefits for human health. Oxid. Med. Cell Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef] [PubMed]
- Morató, L.; Bertini, E.; Verrigni, D.; Ardissone, A.; Ruiz, M.; Ferrer, I.; Uziel, G.; Pujol, A. Mitochondrial dysfunction in central nervous system white matter disorders. Glia 2014, 62, 1878–1894. [Google Scholar] [CrossRef]
- Carrera-Juliá, S.; Moreno, M.L.; Barrios, C.; de la Rubia Ortí, J.E.; Drehmer, E. Antioxidant alternatives in the treatment of amyotrophic lateral sclerosis: A comprehensive review. Front. Physiol. 2020, 11, 63. [Google Scholar] [CrossRef]
- Innamorato, N.G.; Lastres-Becker, I.; Cuadrado, A. Role of microglial redox balance in modulation of neuroinflammation. Curr. Opin. Neurol. 2009, 22, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Dheen, S.T.; Kaur, C.; Ling, E.A. Microglial activation and its implications in the brain diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef] [PubMed]
- Kelso, G.F.; Porteous, C.M.; Coulter, C.V.; Hughes, G.; Porteous, W.K.; Ledgerwood, E.C.; Smith, R.A.; Murphy, M.P. Selective targeting of a redox-active ubiquinone to mitochondria within cells: Antioxidant and antiapoptotic properties. J. Biol. Chem. 2001, 276, 4588–4596. [Google Scholar] [CrossRef] [PubMed]
- Young, M.L.; Franklin, J.L. The mitochondria-targeted antioxidant MitoQ inhibits memory loss, neuropathology, and extends lifespan in aged 3xTg-AD mice. Mol. Cell Neurosci. 2019, 101, 103409. [Google Scholar] [CrossRef]
- Xi, Y.; Feng, D.; Tao, K.; Wang, R.; Shi, Y.; Qin, H.; Murphy, M.P.; Yang, Q.; Zhao, G. MitoQ protects dopaminergic neurons in a 6-OHDA induced PD model by enhancing Mfn2-dependent mitochondrial fusion via activation of PGC-1α. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 2859–2870. [Google Scholar] [CrossRef]
- Grünewald, A.; Kumar, K.R.; Sue, C.M. New insights into the complex role of mitochondria in Parkinson’s disease. Prog. Neurobiol. 2019, 177, 73–93. [Google Scholar] [CrossRef]
- Miquel, E.; Cassina, A.; Martínez-Palma, L.; Souza, J.M.; Bolatto, C.; Rodríguez-Bottero, S.; Logan, A.; Smith, R.A.; Murphy, M.P.; Barbeito, L.; et al. Neuroprotective effects of the mitochondria-targeted antioxidant MitoQ in a model of inherited amyotrophic lateral sclerosis. Free Radic. Biol. Med. 2014, 70, 204–213. [Google Scholar] [CrossRef]
- Pinho, B.R.; Duarte, A.I.; Canas, P.M.; Moreira, P.I.; Murphy, M.P.; Oliveira, J.M.A. The interplay between redox signalling and proteostasis in neurodegeneration: In vivo effects of a mitochondria-targeted antioxidant in Huntington’s disease mice. Free Radic. Biol. Med. 2020, 146, 372–382. [Google Scholar] [CrossRef]
- Weissig, V. Drug development for the therapy of mitochondrial diseases. Trends Mol. Med. 2020, 26, 40–57. [Google Scholar] [CrossRef]
- Stefanova, N.A.; Muraleva, N.A.; Maksimova, K.Y.; Rudnitskaya, E.A.; Kiseleva, E.; Telegina, D.V.; Kolosova, N.G. An antioxidant specifically targeting mitochondria delays progression of Alzheimer’s disease-like pathology. Aging 2016, 8, 2713–2733. [Google Scholar] [CrossRef]
- Brenza, T.M.; Ghaisas, S.; Ramirez, J.E.V.; Harischandra, D.; Anantharam, V.; Kalyanaraman, B.; Kanthasamy, A.G.; Narasimhan, B. Neuronal protection against oxidative insult by polyanhydride nanoparticle-based mitochondria-targeted antioxidant therapy. Nanomedicine 2017, 13, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Langley, M.R.; Harischandra, D.S.; Neal, M.L.; Jin, H.; Anantharam, V.; Joseph, J.; Brenza, T.; Narasimhan, B.; Kanthasamy, A.; et al. Mitoapocynin treatment protects against neuroinflammation and dopaminergic neurodegeneration in a preclinical animal model of Parkinson’s disease. J. Neuroimmune Pharmacol. 2016, 11, 259–278. [Google Scholar] [CrossRef] [PubMed]
- Langley, M.; Ghosh, A.; Charli, A.; Sarkar, S.; Ay, M.; Luo, J.; Zielonka, J.; Brenza, T.; Bennett, B.; Jin, H.; et al. Mito-apocynin prevents mitochondrial dysfunction, microglial activation, oxidative damage, and progressive neurodegeneration in MitoPark transgenic mice. Antioxid. Redox Signal. 2017, 27, 1048–1066. [Google Scholar] [CrossRef] [PubMed]
- Moosmann, B.; Skutella, T.; Beyer, K.; Behl, C. Protective activity of aromatic amines and imines against oxidative nerve cell death. Biol. Chem. 2001, 382, 1601–1612. [Google Scholar] [CrossRef]
- Bachurin, S.O.; Makhaeva, G.F.; Shevtsova, E.F.; Boltneva, N.P.; Kovaleva, N.V.; Lushchekina, S.V.; Rudakova, E.V.; Dubova, L.G.; Vinogradova, D.V.; Sokolov, V.B.; et al. Conjugates of methylene blue with γ-carboline derivatives as new multifunctional agents for the treatment of neurodegenerative diseases. Sci. Rep. 2019, 9, 4873. [Google Scholar] [CrossRef]
- Smith, E.S.; Clark, M.E.; Hardy, G.A.; Kraan, D.J.; Biondo, E.; Gonzalez-Lima, F.; Cormack, L.K.; Monfils, M.; Lee, H.J. Daily consumption of methylene blue reduces attentional deficits and dopamine reduction in a 6-OHDA model of Parkinson’s disease. Neuroscience 2017, 359, 8–16. [Google Scholar] [CrossRef]
- Rocha, M.; Hernandez-Mijares, A.; Garcia-Malpartida, K.; Bañuls, C.; Bellod, L.; Victor, V.M. Mitochondria-targeted antioxidant peptides. Curr. Pharm. Des. 2010, 16, 3124–3131. [Google Scholar] [CrossRef]
- Szeto, H.H.; Birk, A.V. Serendipity and the discovery of novel compounds that restore mitochondrial plasticity. Clin. Pharmacol. Ther. 2014, 96, 672–683. [Google Scholar] [CrossRef]
- Searls, R.L.; Sanadi, D.R. α-Ketoglutaric dehydrogenase. VIII. Isolation and some properties of a flavoprotein component. J. Biol. Chem. 1960, 235, 2485–2491. [Google Scholar]
- Dos Santos, S.M.; Romeiro, C.F.R.; Rodrigues, C.A.; Cerqueira, A.R.L.; Monteiro, M.C. Mitochondrial dysfunction and alpha-lipoic acid: Beneficial or harmful in Alzheimer’s disease? Oxid. Med. Cell Longev. 2019, 2019, 8409329. [Google Scholar] [CrossRef]
- Zhang, N.; Shu, H.Y.; Huang, T.; Zhang, Q.L.; Li, D.; Zhang, G.Q.; Peng, X.Y.; Liu, C.F.; Luo, W.F.; Hu, L.F. Nrf2 signaling contributes to the neuroprotective effects of urate against 6-OHDA toxicity. PLoS ONE 2014, 9, e100286. [Google Scholar] [CrossRef] [PubMed]
- Paganoni, S.; Schwarzschild, M.A. Urate as a marker of risk and progression of neurodegenerative disease. Neurotherapeutics 2017, 14, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Schwarzschild, M.A.; Ascherio, A.; Beal, M.F.; Cudkowicz, M.E.; Curhan, G.C.; Hare, J.M.; Hooper, D.C.; Kieburtz, K.D.; Macklin, E.A.; Oakes, D.; et al. Inosine to increase serum and cerebrospinal fluid urate in Parkinson disease: A randomized clinical trial. JAMA Neurol. 2014, 71, 141–150. [Google Scholar] [CrossRef]
- Nicholson, K.; Chan, J.; Macklin, E.A.; Levine-Weinberg, M.; Breen, C.; Bakshi, R.; Grasso, D.L.; Wills, A.M.; Jahandideh, S.; Taylor, A.A.; et al. Pilot trial of inosine to elevate urate levels in amyotrophic lateral sclerosis. Ann. Clin. Transl. Neurol. 2018, 5, 1522–1533. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal. Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Plummer, B.F.; Hardies, L.J.; Weintraub, S.T.; Vijayalaxmi; Shepherd, A.M. A novel melatonin metabolite, cyclic 3-hydroxymelatonin: A biomarker of in vivo hydroxyl radical generation. Biochem. Biophys. Res. Commun. 1998, 253, 614–620. [Google Scholar] [CrossRef]
- Cheng, Y.; Feng, Z.; Zhang, Q.Z.; Zhang, J.T. Beneficial effects of melatonin in experimental models of Alzheimer disease. Acta Pharmacol. Sin. 2006, 27, 129–139. [Google Scholar] [CrossRef]
- Weishaupt, J.H.; Bartels, C.; Pölking, E.; Dietrich, J.; Rohde, G.; Poeggeler, B.; Mertens, N.; Sperling, S.; Bohn, M.; Hüther, G.; et al. Reduced oxidative damage in ALS by high-dose enteral melatonin treatment. J. Pineal. Res. 2006, 41, 313–323. [Google Scholar] [CrossRef]
- Rudnitskaya, E.A.; Muraleva, N.A.; Maksimova, K.Y.; Kiseleva, E.; Kolosova, N.G.; Stefanova, N.A. Melatonin attenuates memory impairment, amyloid-β accumulation, and neurodegeneration in a rat model of sporadic Alzheimer’s disease. J. Alzheimers Dis. 2015, 47, 103–116. [Google Scholar] [CrossRef]
- Lee, J.G.; Woo, Y.S.; Park, S.W.; Seog, D.H.; Seo, M.K.; Bahk, W.M. The neuroprotective effects of melatonin: Possible role in the pathophysiology of neuropsychiatric disease. Brain Sci. 2019, 9, 285. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin: Clinical perspectives in neurodegeneration. Front. Endocrinol. 2019, 10, 480. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Brown, G.M.; Spence, D.W.; Bharti, V.K.; Kaur, C.; Hardeland, R.; Cardinali, D.P. Melatonin antioxidative defense: Therapeutical implications for aging and neurodegenerative processes. Neurotox. Res. 2013, 23, 267–300. [Google Scholar] [CrossRef] [PubMed]
- Samuni, Y.; Goldstein, S.; Dean, O.M.; Berk, M. The chemistry and biological activities of N-acetylcysteine. Biochim. Biophys. Acta 2013, 1830, 4117–4129. [Google Scholar] [CrossRef]
- Aldini, G.; Altomare, A.; Baron, G.; Vistoli, G.; Carini, M.; Borsani, L.; Sergio, F. N-Acetylcysteine as an antioxidant and disulphide breaking agent: The reasons why. Free Radic. Res. 2018, 52, 751–762. [Google Scholar] [CrossRef] [PubMed]
- Tardiolo, G.; Bramanti, P.; Mazzon, E. Overview on the effects of N-acetylcysteine in neurodegenerative diseases. Molecules 2018, 23, 3305. [Google Scholar] [CrossRef]
- Bavarsad Shahripour, R.; Harrigan, M.R.; Alexandrov, A.V. N-acetylcysteine (NAC) in neurological disorders: Mechanisms of action and therapeutic opportunities. Brain Behav. 2014, 4, 108–122. [Google Scholar] [CrossRef]
- Remington, R.; Bechtel, C.; Larsen, D.; Samar, A.; Doshanjh, L.; Fishman, P.; Luo, Y.; Smyers, K.; Page, R.; Morrell, C.; et al. A phase II randomized clinical trial of a nutritional formulation for cognition and mood in Alzheimer’s disease. J. Alzheimers Dis. 2015, 45, 395–405. [Google Scholar] [CrossRef]
- Stahl, W.; Sies, H. Antioxidant defense: Vitamins E and C and carotenoids. Diabetes 1997, 46, S14–S18. [Google Scholar] [CrossRef]
- Medina, S.; Martínez, M.; Hernanz, A. Antioxidants inhibit the human cortical neuron apoptosis induced by hydrogen peroxide, tumor necrosis factor alpha, dopamine and beta-amyloid peptide 1-42. Free Radic. Res. 2002, 36, 1179–1184. [Google Scholar] [CrossRef]
- Kook, S.Y.; Lee, K.M.; Kim, Y.; Cha, M.Y.; Kang, S.; Baik, S.H.; Lee, H.; Park, R.; Mook-Jung, I. High-dose of vitamin C supplementation reduces amyloid plaque burden and ameliorates pathological changes in the brain of 5XFAD mice. Cell Death Dis. 2014, 5, e1083. [Google Scholar] [CrossRef]
- Dixit, S.; Fessel, J.P.; Harrison, F.E. Mitochondrial dysfunction in the APP/PSEN1 mouse model of Alzheimer’s disease and a novel protective role for ascorbate. Free Radic. Biol. Med. 2017, 112, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Selvaraju, T.R.; Khaza’ai, H.; Vidyadaran, S.; Abd Mutalib, M.S.; Vasudevan, R. The neuroprotective effects of tocotrienol rich fraction and alpha tocopherol against glutamate injury in astrocytes. Bosn. J. Basic Med. Sci. 2014, 14, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Schloesser, A.; Esatbeyoglu, T.; Piegholdt, S.; Dose, J.; Ikuta, N.; Okamoto, H.; Ishida, Y.; Terao, K.; Matsugo, S.; Rimbach, G. Dietary tocotrienol/γ-cyclodextrin complex increases mitochondrial membrane potential and ATP concentrations in the brains of aged mice. Oxid. Med. Cell Longev. 2015, 2015, 789710. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Huang, X.; Zhen, J.; Van Halm-Lutterodt, N.; Wang, J.; Zhou, C.; Yuan, L. Dietary vitamin E status dictates oxidative stress outcomes by modulating effects of fish oil supplementation in Alzheimer disease model APP. Mol. Neurobiol. 2018, 55, 9204–9219. [Google Scholar] [CrossRef]
- Browne, D.; McGuinness, B.; Woodside, J.V.; McKay, G.J. Vitamin E and Alzheimer’s disease: What do we know so far? Clin. Interv. Aging 2019, 14, 1303–1317. [Google Scholar] [CrossRef]
- Gurney, M.E.; Cutting, F.B.; Zhai, P.; Doble, A.; Taylor, C.P.; Andrus, P.K.; Hall, E.D. Benefit of vitamin E, riluzole, and gabapentin in a transgenic model of familial amyotrophic lateral sclerosis. Ann. Neurol. 1996, 39, 147–157. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Al-Amin, M.M.; Akhter, S.; Hasan, A.T.; Alam, T.; Nageeb Hasan, S.M.; Saifullah, A.R.; Shohel, M. The antioxidant effect of astaxanthin is higher in young mice than aged: A region specific study on brain. Metab. Brain Dis. 2015, 30, 1237–1246. [Google Scholar] [CrossRef]
- Lobos, P.; Bruna, B.; Cordova, A.; Barattini, P.; Galáz, J.L.; Adasme, T.; Hidalgo, C.; Muñoz, P.; Paula-Lima, A. Astaxanthin protects primary hippocampal neurons against noxious effects of Aβ-oligomers. Neural Plast. 2016, 2016, 3456783. [Google Scholar] [CrossRef]
- Uittenbogaard, M.; Chiaramello, A. Mitochondrial biogenesis: A therapeutic target for neurodevelopmental disorders and neurodegenerative diseases. Curr. Pharm. Des. 2014, 20, 5574–5593. [Google Scholar] [CrossRef]
- Eschbach, J.; von Einem, B.; Müller, K.; Bayer, H.; Scheffold, A.; Morrison, B.E.; Rudolph, K.L.; Thal, D.R.; Witting, A.; Weydt, P.; et al. Mutual exacerbation of peroxisome proliferator-activated receptor γ coactivator 1α deregulation and α-synuclein oligomerization. Ann. Neurol. 2015, 77, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Baines, C.P.; Kaiser, R.A.; Purcell, N.H.; Blair, N.S.; Osinska, H.; Hambleton, M.A.; Brunskill, E.W.; Sayen, M.R.; Gottlieb, R.A.; Dorn, G.W.; et al. Loss of cyclophilin D reveals a critical role for mitochondrial permeability transition in cell death. Nature 2005, 434, 658–662. [Google Scholar] [CrossRef] [PubMed]
- Bordet, T.; Berna, P.; Abitbol, J.L.; Pruss, R.M. Olesoxime (TRO19622): A novel mitochondrial-targeted neuroprotective compound. Pharmaceuticals 2010, 3, 345–368. [Google Scholar] [CrossRef] [PubMed]
- Petrov, D.; Mansfield, C.; Moussy, A.; Hermine, O. ALS clinical trials review: 20 years of failure. Are we any closer to registering a new treatment? Front. Aging Neurosci. 2017, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.J.; Clemensson, L.E.; Schiöth, H.B.; Nguyen, H.P. Olesoxime in neurodegenerative diseases: Scrutinising a promising drug candidate. Biochem. Pharmacol. 2019, 168, 305–318. [Google Scholar] [CrossRef]
- Demarin, V.; Podobnik, S.S.; Storga-Tomic, D.; Kay, G. Treatment of Alzheimer’s disease with stabilized oral nicotinamide adenine dinucleotide: A randomized, double-blind study. Drugs Exp. Clin. Res. 2004, 30, 27–33. [Google Scholar]
- Harlan, B.A.; Pehar, M.; Sharma, D.R.; Beeson, G.; Beeson, C.C.; Vargas, M.R. Enhancing NAD+ salvage pathway reverts the toxicity of primary astrocytes expressing amyotrophic lateral sclerosis-linked mutant superoxide dismutase 1 (SOD1). J. Biol. Chem. 2016, 291, 10836–10846. [Google Scholar] [CrossRef]
- Roe, C.R.; Mochel, F. Anaplerotic diet therapy in inherited metabolic disease: Therapeutic potential. J. Inherit. Metab. Dis. 2006, 29, 332–340. [Google Scholar] [CrossRef]
- Adanyeguh, I.M.; Rinaldi, D.; Henry, P.G.; Caillet, S.; Valabregue, R.; Durr, A.; Mochel, F. Triheptanoin improves brain energy metabolism in patients with Huntington disease. Neurology 2015, 84, 490–495. [Google Scholar] [CrossRef]
- Tefera, T.W.; Wong, Y.; Barkl-Luke, M.E.; Ngo, S.T.; Thomas, N.K.; McDonald, T.S.; Borges, K. Triheptanoin protects motor neurons and delays the onset of motor symptoms in a mouse model of amyotrophic lateral sclerosis. PLoS ONE 2016, 11, e0161816. [Google Scholar] [CrossRef]
- Kolaj, I.; Imindu Liyanage, S.; Weaver, D.F. Phenylpropanoids and Alzheimer’s disease: A potential therapeutic platform. Neurochem. Int. 2018, 120, 99–111. [Google Scholar] [CrossRef]
- Bors, W.; Heller, W.; Michel, C.; Saran, M. Flavonoids as antioxidants: Determination of radical-scavenging efficiencies. Methods Enzymol. 1990, 186, 343–355. [Google Scholar] [PubMed]
- Moskaug, J.; Carlsen, H.; Myhrstad, M.C.; Blomhoff, R. Polyphenols and glutathione synthesis regulation. Am. J. Clin. Nutr. 2005, 81, 277S–283S. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.L.; Wilhelmus, M.M.; de Vries, H.E.; Drukarch, B.; Hoozemans, J.J.; van Horssen, J. Antioxidative defense mechanisms controlled by Nrf2: State-of-the-art and clinical perspectives in neurodegenerative diseases. Arch. Toxicol. 2014, 88, 1773–1786. [Google Scholar] [CrossRef] [PubMed]
- Fiorani, M.; Guidarelli, A.; Blasa, M.; Azzolini, C.; Candiracci, M.; Piatti, E.; Cantoni, O. Mitochondria accumulate large amounts of quercetin: Prevention of mitochondrial damage and release upon oxidation of the extramitochondrial fraction of the flavonoid. J. Nutr. Biochem. 2010, 21, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.H.; Kwon, H.; Kim, K.S.; Kim, J.; Kim, M.H.; Yu, H.J.; Kim, M.; Lee, K.W.; Do, B.R.; Jung, H.K.; et al. Epigallocatechin gallate prevents oxidative-stress-induced death of mutant Cu/Zn-superoxide dismutase (G93A) motoneuron cells by alteration of cell survival and death signals. Toxicology 2004, 202, 213–225. [Google Scholar] [CrossRef]
- Jiang, W.; Luo, T.; Li, S.; Zhou, Y.; Shen, X.Y.; He, F.; Xu, J.; Wang, H.Q. Quercetin protects against okadaic acid-induced injury via MAPK and PI3K/Akt/GSK3β signaling pathways in HT22 hippocampal neurons. PLoS ONE 2016, 11, e0152371. [Google Scholar] [CrossRef]
- Huang, D.S.; Yu, Y.C.; Wu, C.H.; Lin, J.Y. Protective effects of wogonin against Alzheimer’s disease by inhibition of amyloidogenic pathway. Evid. Based Complement. Alternat. Med. 2017, 2017, 3545169. [Google Scholar] [CrossRef]
- Mancuso, R.; del Valle, J.; Modol, L.; Martinez, A.; Granado-Serrano, A.B.; Ramirez-Núñez, O.; Pallás, M.; Portero-Otin, M.; Osta, R.; Navarro, X. Resveratrol improves motoneuron function and extends survival in SOD1(G93A) ALS mice. Neurotherapeutics 2014, 11, 419–432. [Google Scholar]
- Moussa, C.; Hebron, M.; Huang, X.; Ahn, J.; Rissman, R.A.; Aisen, P.S.; Turner, R.S. Resveratrol regulates neuro-inflammation and induces adaptive immunity in Alzheimer’s disease. J. Neuroinflammation 2017, 14, 1. [Google Scholar] [CrossRef]
- Zhu, C.W.; Grossman, H.; Neugroschl, J.; Parker, S.; Burden, A.; Luo, X.; Sano, M. A randomized, double-blind, placebo-controlled trial of resveratrol with glucose and malate (RGM) to slow the progression of Alzheimer’s disease: A pilot study. Alzheimers Dement. 2018, 4, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Ferrari-Toninelli, G.; Maccarinelli, G.; Uberti, D.; Buerger, E.; Memo, M. Mitochondria-targeted antioxidant effects of S(-) and R(+) pramipexole. BMC Pharmacol. 2010, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Cassarino, D.S.; Fall, C.P.; Smith, T.S.; Bennett, J.P. Pramipexole reduces reactive oxygen species production in vivo and in vitro and inhibits the mitochondrial permeability transition produced by the parkinsonian neurotoxin methylpyridinium ion. J. Neurochem. 1998, 71, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, I.; Parvez, S.; Winkler-Stuck, K.; Seitz, G.; Trieu, I.; Wallesch, C.W.; Schönfeld, P.; Siemen, D. Patch clamp reveals powerful blockade of the mitochondrial permeability transition pore by the D2-receptor agonist pramipexole. FASEB J. 2006, 20, 556–558. [Google Scholar] [CrossRef] [PubMed]
- Shin, E.J.; Nam, Y.; Lee, J.W.; Nguyen, P.T.; Yoo, J.E.; Tran, T.V.; Jeong, J.H.; Jang, C.G.; Oh, Y.J.; Youdim, M.B.H.; et al. N-Methyl, N-propynyl-2-phenylethylamine (MPPE), a selegiline analog, attenuates MPTP-induced dopaminergic toxicity with guaranteed behavioral safety: Involvement of inhibitions of mitochondrial oxidative burdens and p53 gene-elicited pro-apoptotic change. Mol. Neurobiol. 2016, 53, 6251–6269. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; Qiu, K.; Liu, H.; He, Q.; Bai, J.H.; Lu, W. Application and prospects of butylphthalide for the treatment of neurologic diseases. Chin. Med. J. 2019, 132, 1467–1477. [Google Scholar] [CrossRef]
- Kapoor, S. Dl-3-n-butylphthalide and its emerging beneficial effects in neurology. Chin. Med. J. 2012, 125, 3360. [Google Scholar]
- Xiong, N.; Huang, J.; Chen, C.; Zhao, Y.; Zhang, Z.; Jia, M.; Hou, L.; Yang, H.; Cao, X.; Liang, Z.; et al. Dl-3-n-butylphthalide, a natural antioxidant, protects dopamine neurons in rotenone models for Parkinson’s disease. Neurobiol. Aging 2012, 33, 1777–1791. [Google Scholar] [CrossRef]
- Zhou, H.; Ye, M.; Xu, W.; Yu, M.; Liu, X.; Chen, Y. DL-3-n-butylphthalide therapy for Parkinson’s disease: A randomized controlled trial. Exp. Ther. Med. 2019, 17, 3800–3806. [Google Scholar] [CrossRef]
- Paumgartner, G.; Beuers, U. Ursodeoxycholic acid in cholestatic liver disease: Mechanisms of action and therapeutic use revisited. Hepatology 2002, 36, 525–531. [Google Scholar] [CrossRef]
- Ackerman, H.D.; Gerhard, G.S. Bile acids in neurodegenerative disorders. Front. Aging Neurosci. 2016, 8, 263. [Google Scholar] [CrossRef] [PubMed]
- Keene, C.D.; Rodrigues, C.M.; Eich, T.; Chhabra, M.S.; Steer, C.J.; Low, W.C. Tauroursodeoxycholic acid, a bile acid, is neuroprotective in a transgenic animal model of Huntington’s disease. Proc. Natl. Acad. Sci. USA 2002, 99, 10671–10676. [Google Scholar] [CrossRef] [PubMed]
- Vaz, A.R.; Cunha, C.; Gomes, C.; Schmucki, N.; Barbosa, M.; Brites, D. Glycoursodeoxycholic acid reduces matrix metalloproteinase-9 and caspase-9 activation in a cellular model of superoxide dismutase-1 neurodegeneration. Mol. Neurobiol. 2015, 51, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.M.; Barnes, K.; Clemmens, H.; Al-Rafiah, A.R.; Al-Ofi, E.A.; Leech, V.; Bandmann, O.; Shaw, P.J.; Blackburn, D.J.; Ferraiuolo, L.; et al. Ursodeoxycholic acid improves mitochondrial function and redistributes Drp1 in fibroblasts from patients with either sporadic or familial Alzheimer’s disease. J. Mol. Biol. 2018, 430, 3942–3953. [Google Scholar] [CrossRef]
- Elia, A.E.; Lalli, S.; Monsurrò, M.R.; Sagnelli, A.; Taiello, A.C.; Reggiori, B.; La Bella, V.; Tedeschi, G.; Albanese, A. Tauroursodeoxycholic acid in the treatment of patients with amyotrophic lateral sclerosis. Eur. J. Neurol. 2016, 23, 45–52. [Google Scholar] [CrossRef]
- Daruich, A.; Picard, E.; Boatright, J.H.; Behar-Cohen, F. Review: The bile acids urso- and tauroursodeoxycholic acid as neuroprotective therapies in retinal disease. Mol. Vis. 2019, 25, 610–624. [Google Scholar]
- Wirth, B.; Karakaya, M.; Kye, M.J.; Mendoza-Ferreira, N. Twenty-five years of spinal muscular atrophy research: From phenotype to genotype to therapy, and what comes next. Annu. Rev. Genomics Hum. Genet. 2020, 21. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).