Variation in MicroRNA Expression Profile of Uterine Leiomyoma with Endometrial Cavity Distortion and Endometrial Cavity Non-Distortion



 and
and
Abstract
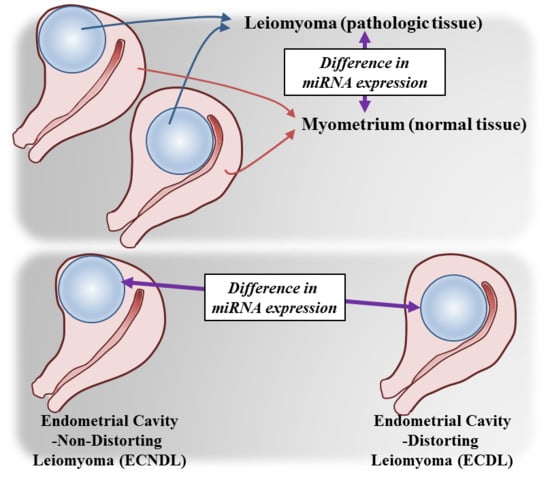
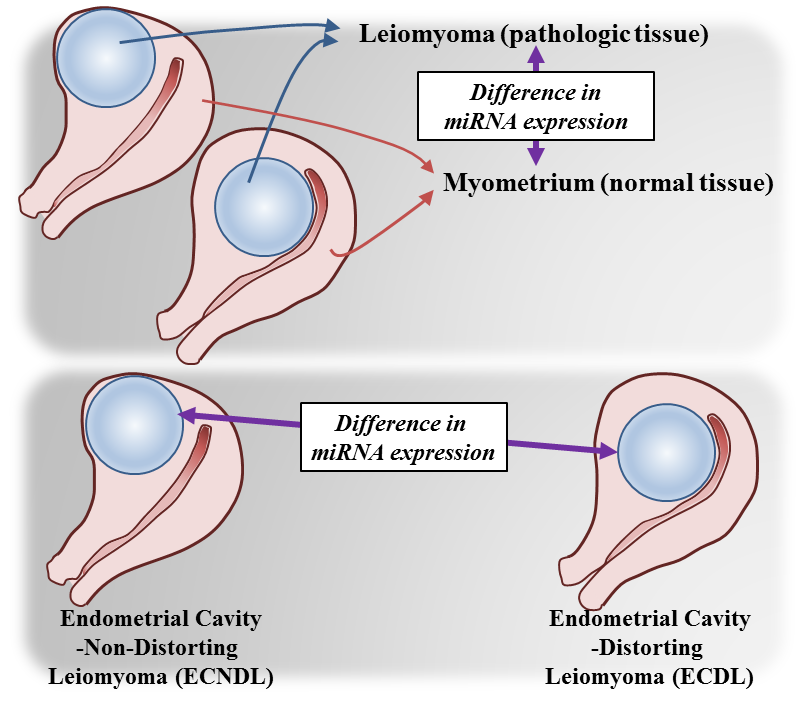
1. Introduction
2. Results
2.1. Characteristics of the Patients and Uterine Leiomyoma (UL) Tissues
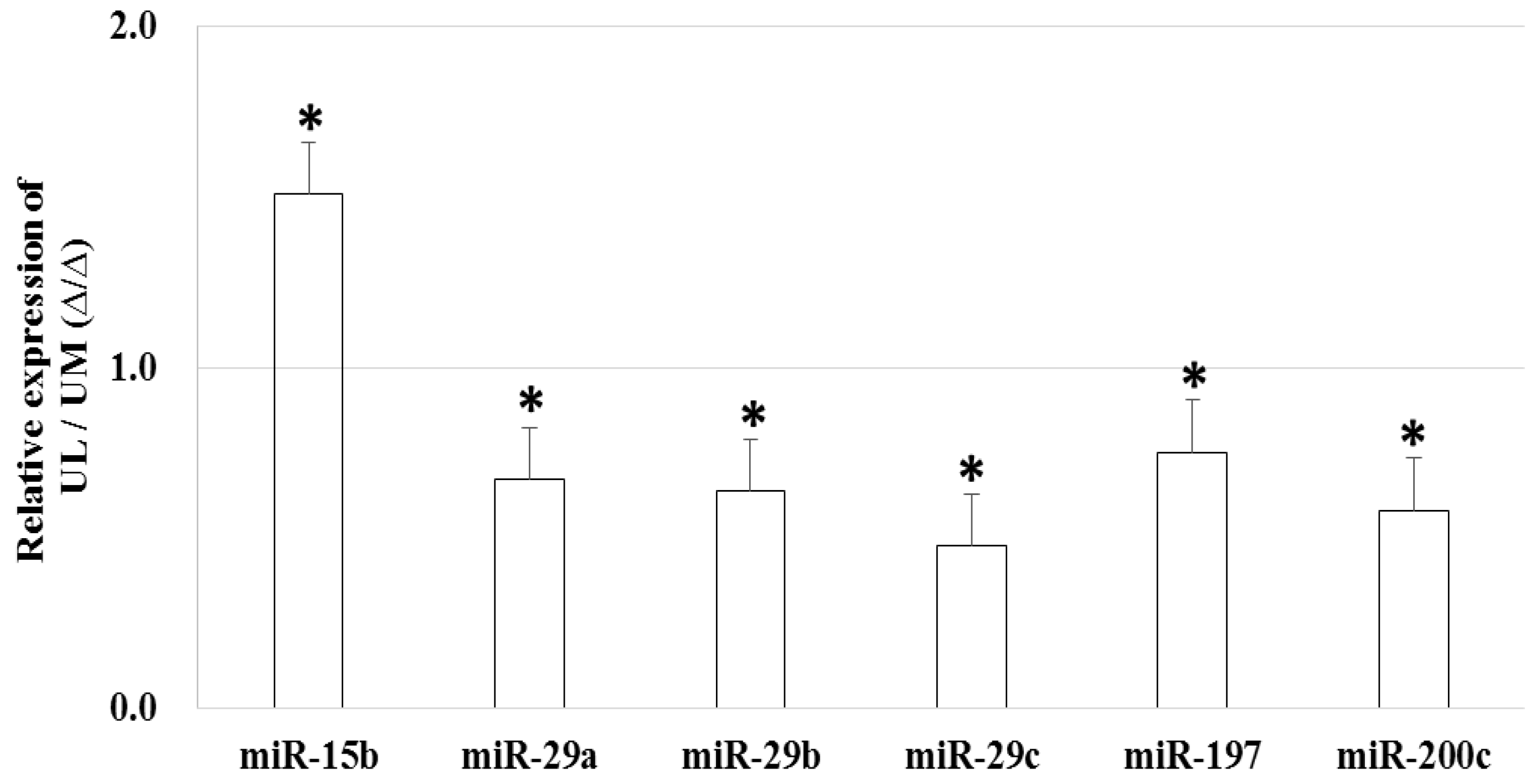
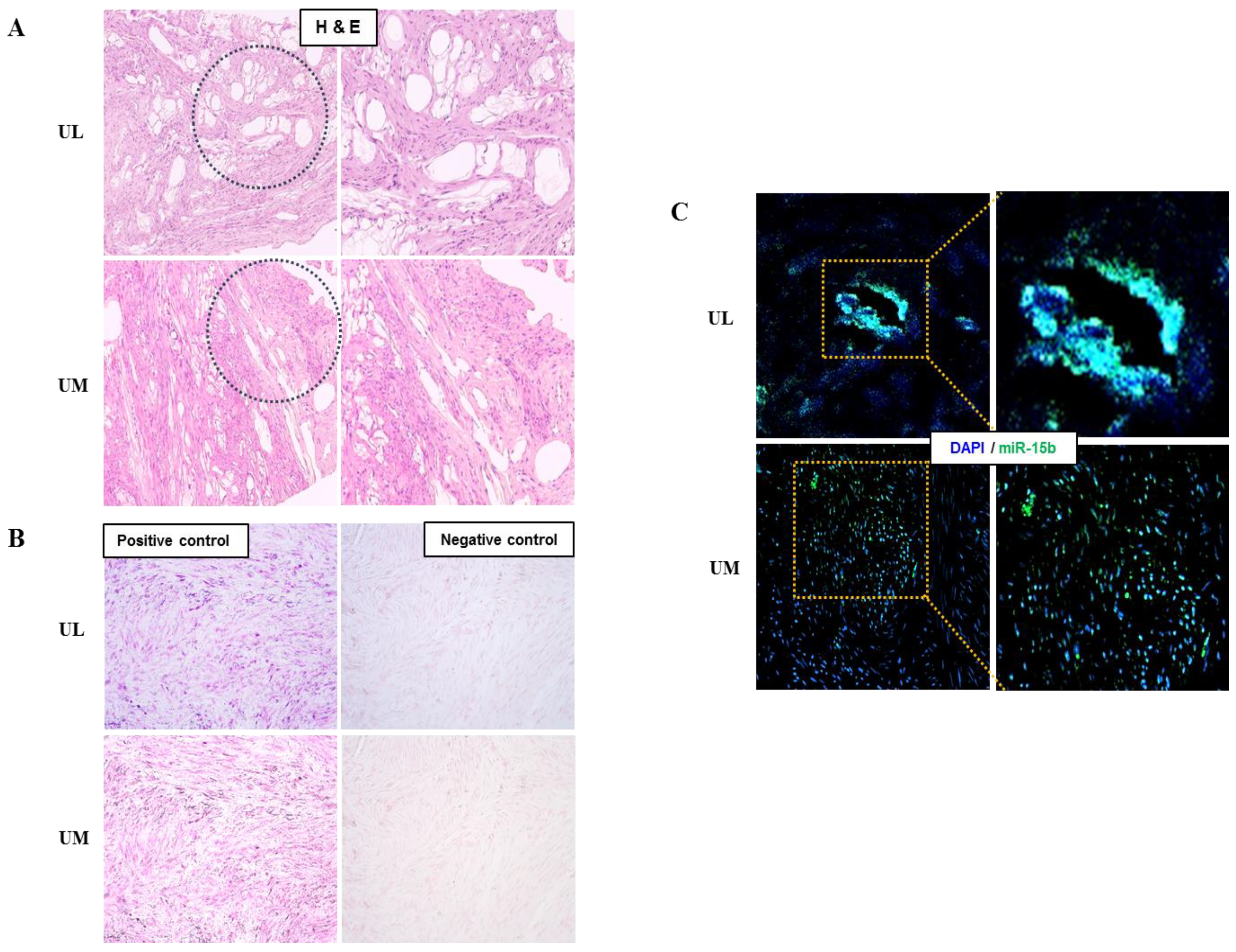
2.2. The Expression Profiles of miRNAs in UL Cells
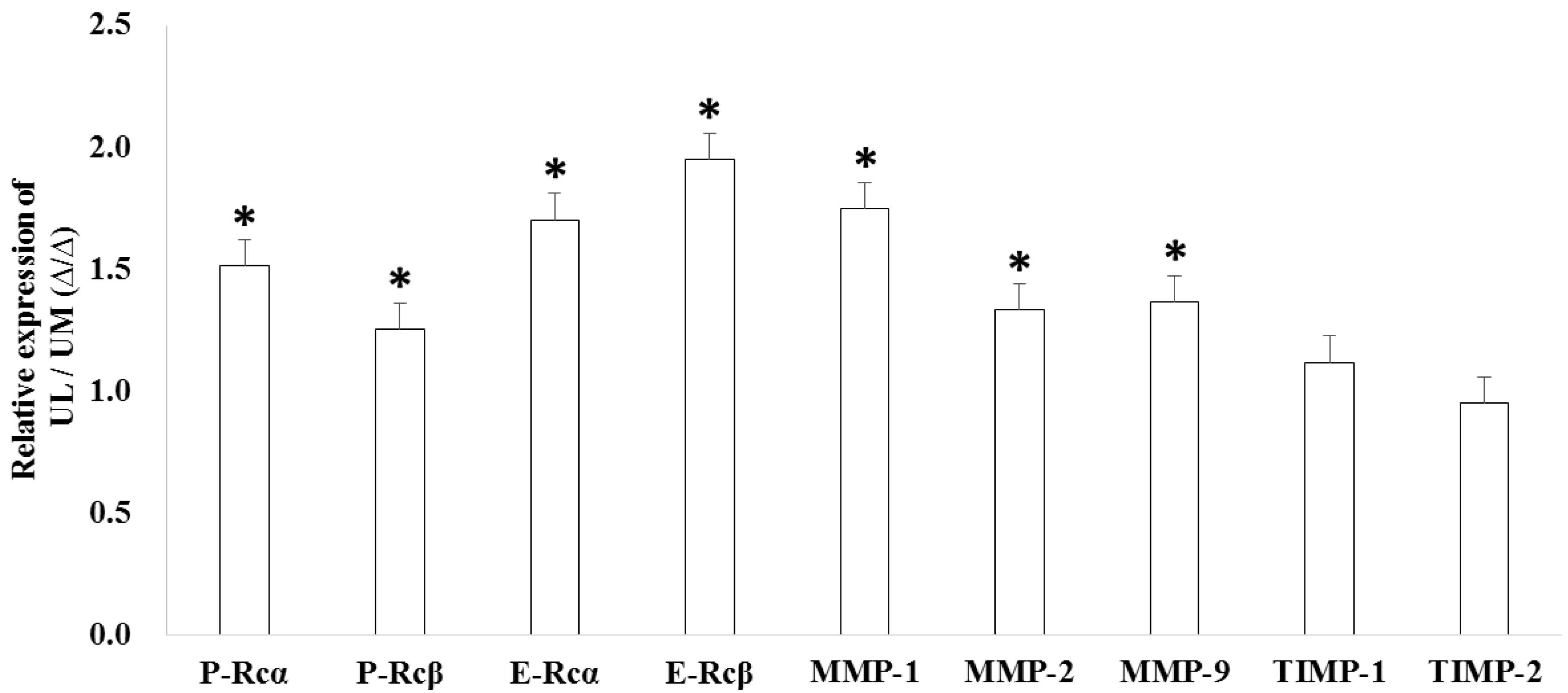
2.3. The Expression Levels of Candidate Target Genes in UL Cells
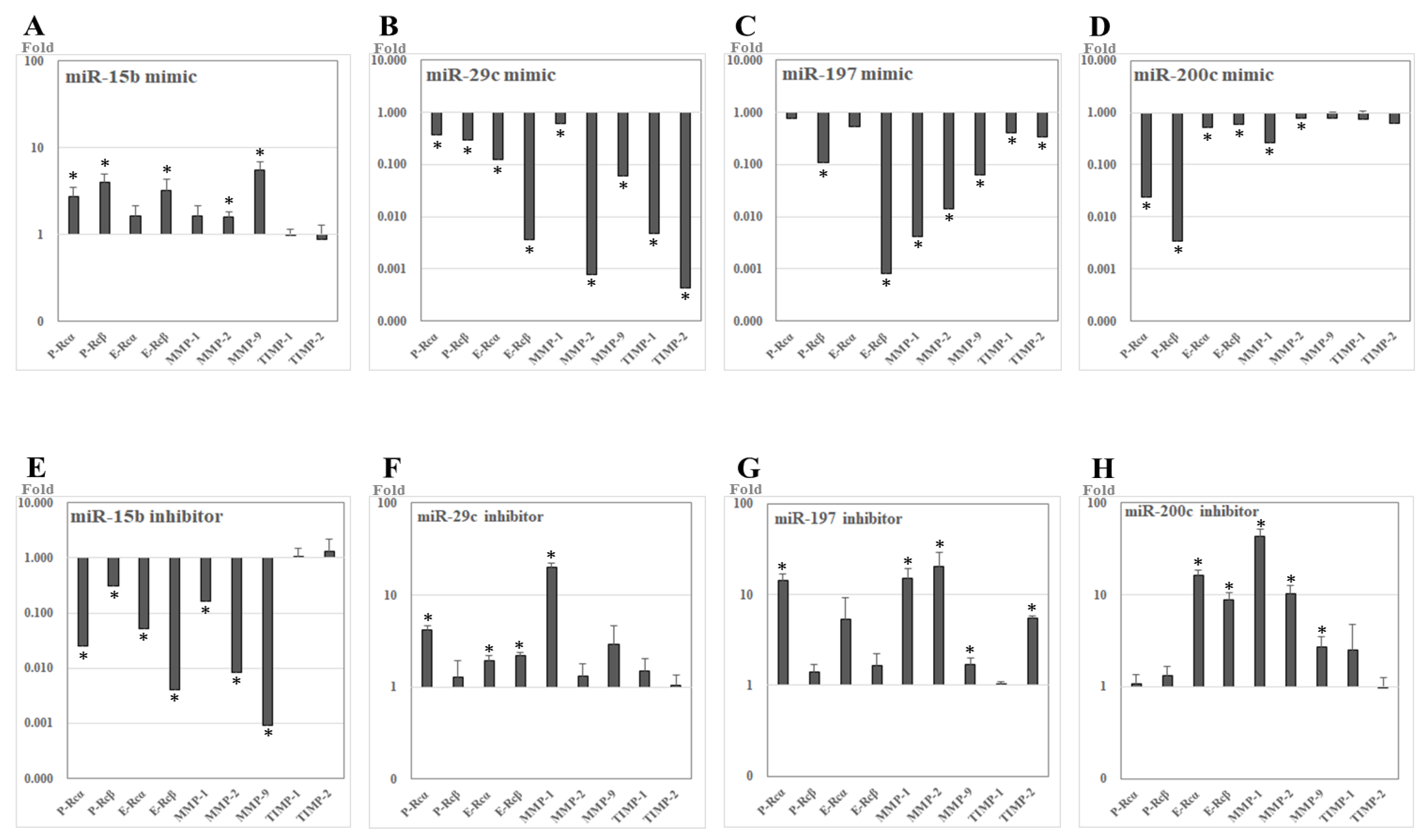
2.4. The Expression Levels of Candidate Target Genes after miRNA Transfection into UL Cells
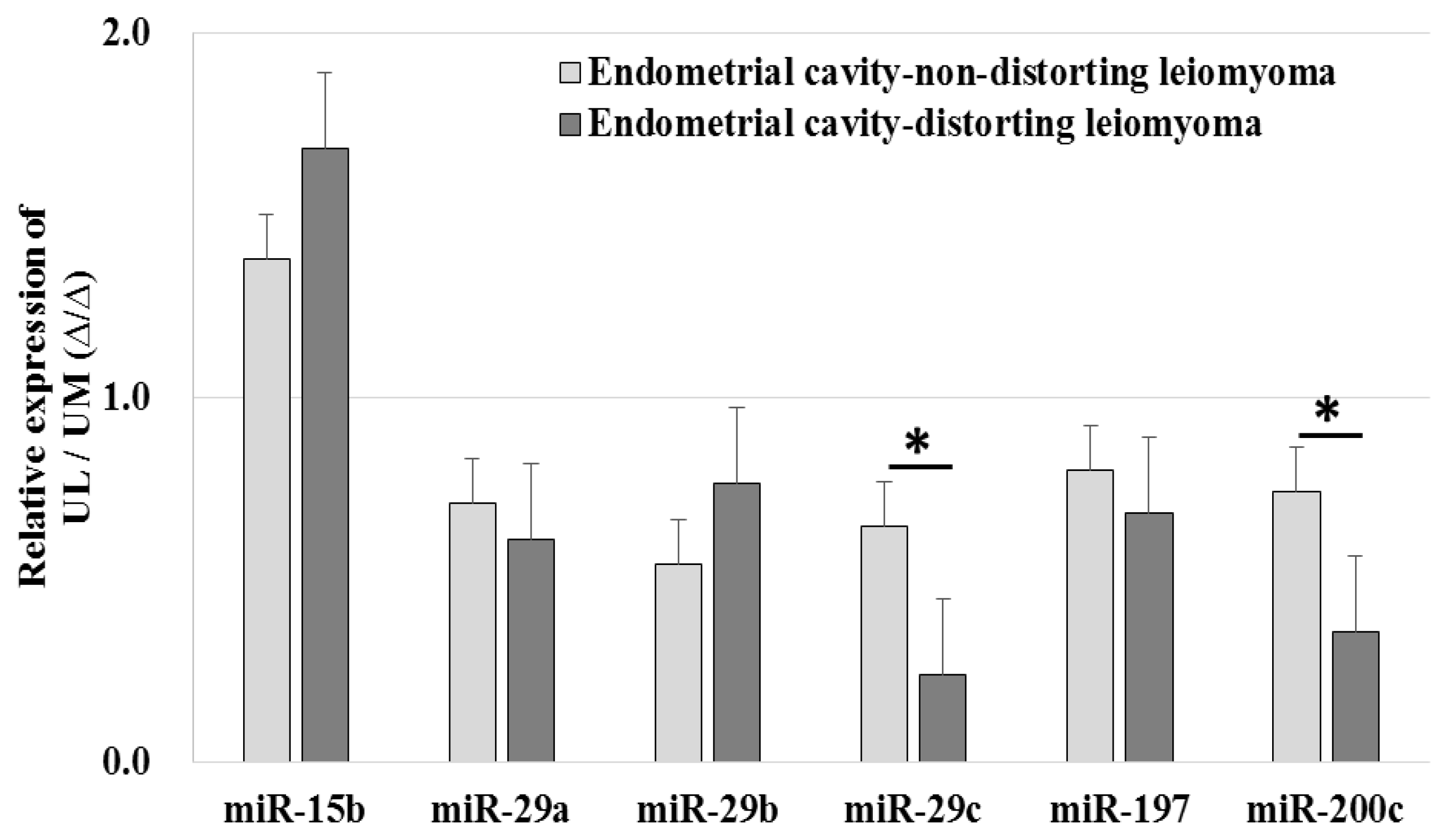
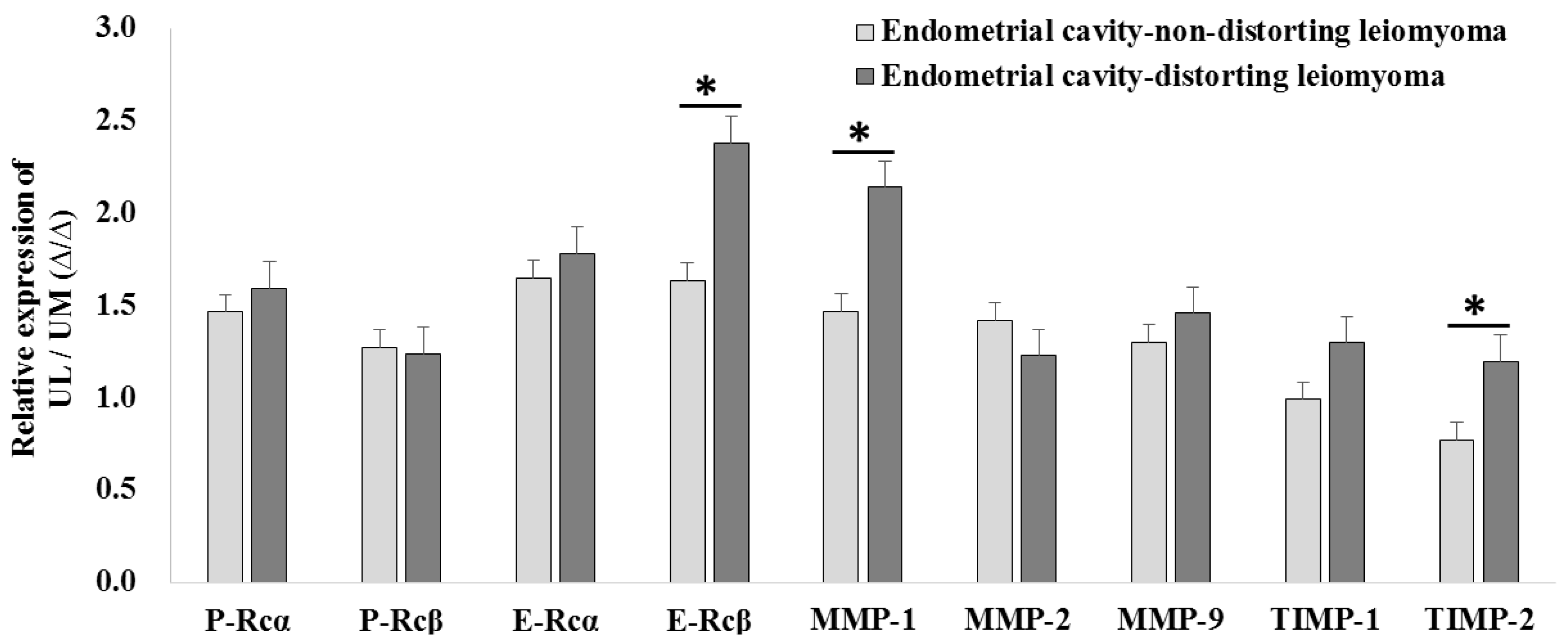
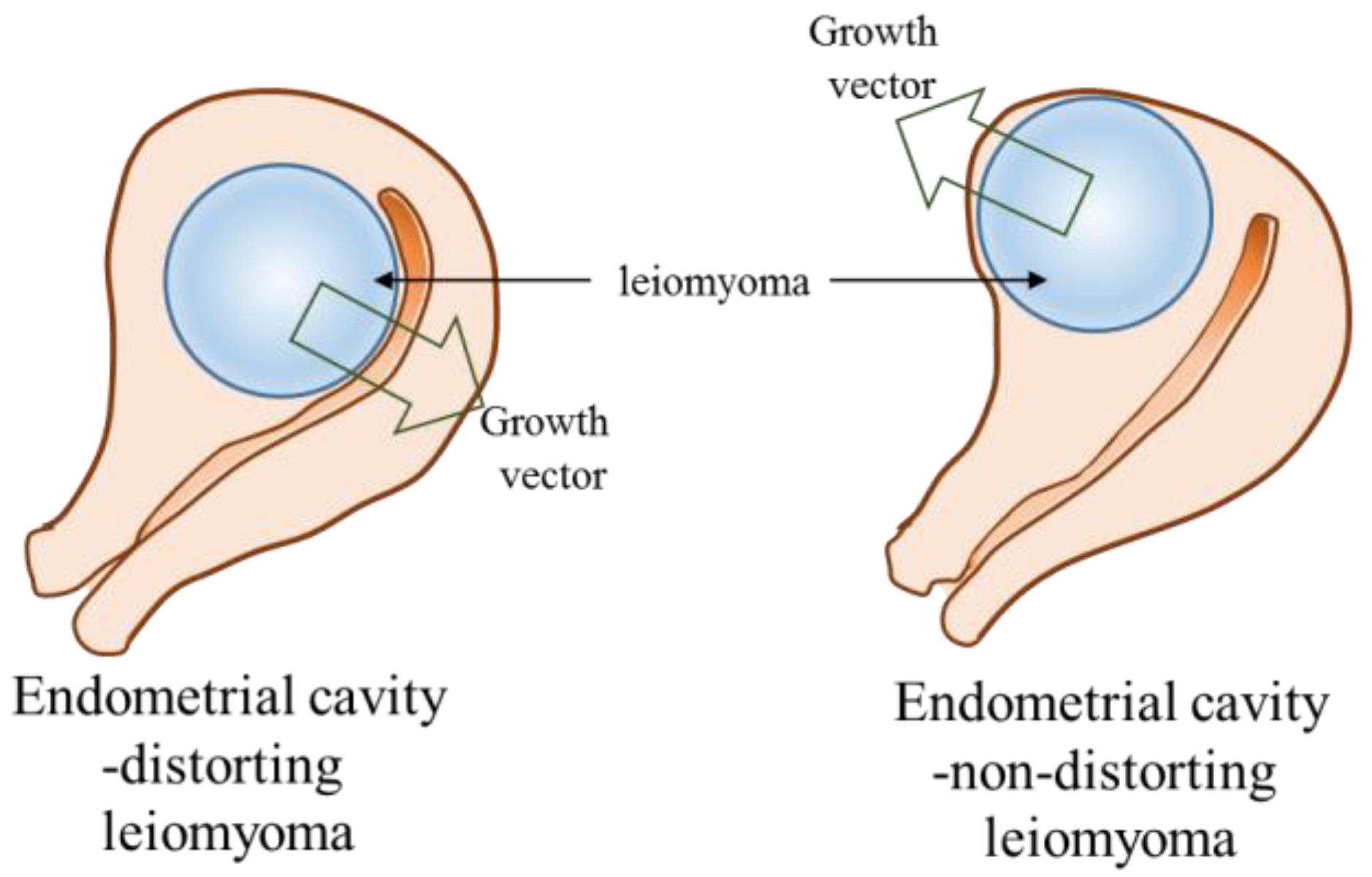
2.5. The Expression Levels of miRNAs and Candidate Target Genes Associated to Endometrial Cavity Distortion (ECD)
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Measurement of Hardness of Uterine Leiomyoma (UL) Tissues
4.3. RNA Isolation from Uterine Leiomyoma (UL) and Uterine Myometrium (UM) Tissues
4.4. qRT-PCR to Assess the Levels of miRNAs
4.5. qPCR to Assess the Levels of Candidate Target Genes
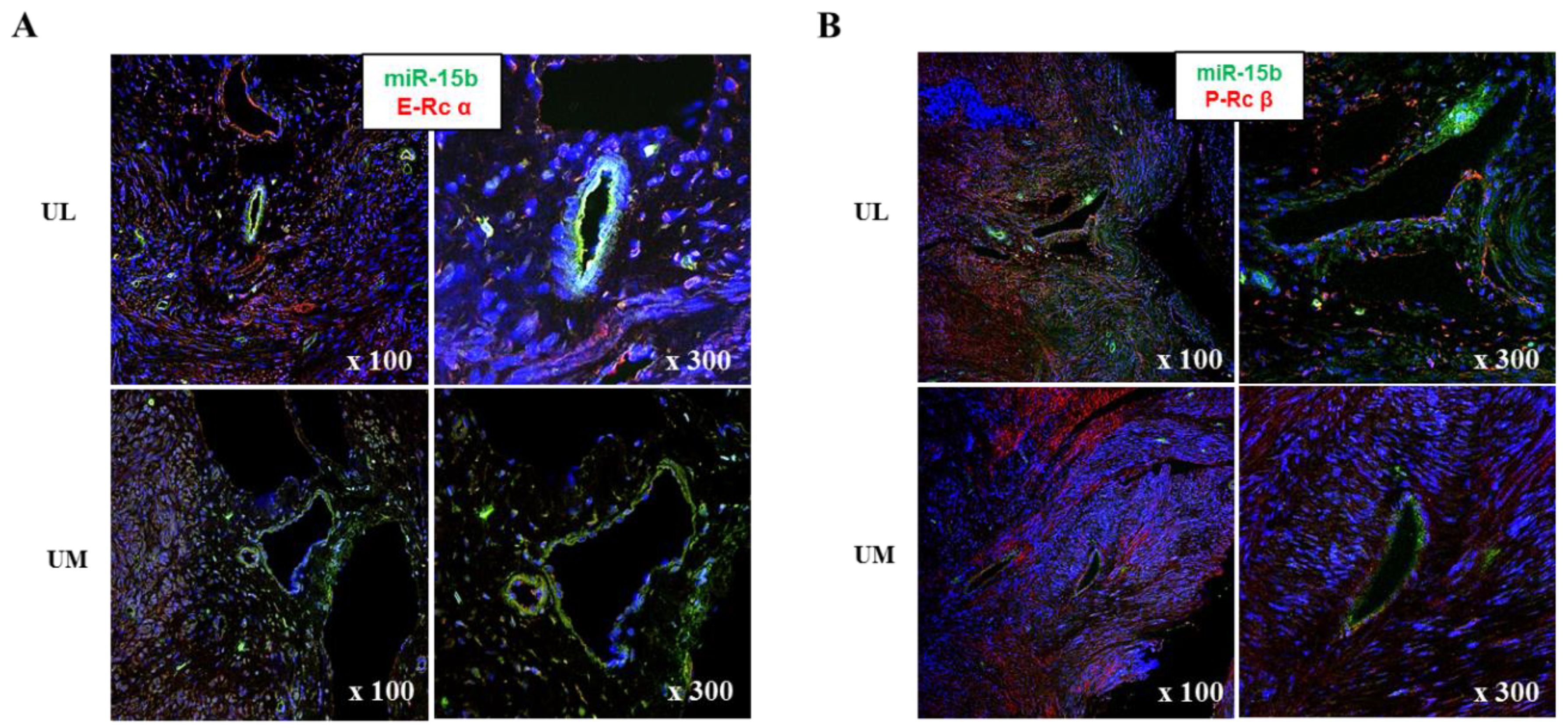
4.6. In Situ Hybridization of miRNAs
4.7. Culture of UL Cells
4.8. Transfection of Mimics or Inhibitors of miRNAs into In Vitro Cultured UL Cells
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Al-Hendy, A.; Salama, S. Gene Therapy and uterine leiomyoma: A review. Hum. Reprod. Update 2006, 12, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Drayer, S.M.; Catherino, W.H. Prevalence, morbidity, and current medical management of uterine leiomyomas. Int. J. Gynaecol. Obstet. 2015, 131, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Benson, C.B.; Chow, J.S.; Chang-Lee, W.; Hill, J.A., 3rd; Doubilet, P.M. Outcome of pregnancies in women with uterine leiomyomas identified by sonography in the first trimester. J. Clin. Ultrasound 2001, 29, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Hart, R.; Khalaf, Y.; Yeong, C.T.; Seed, P.; Taylor, A.; Braude, P. A Prospective controlled study of the effect of intramural uterine fibroids on the outcome of assisted conception. Hum. Reprod. 2001, 16, 2411–2417. [Google Scholar] [CrossRef] [PubMed]
- Casini, M.L.; Rossi, F.; Agostini, R.; Unfer, V. Effects of the position of fibroids on fertility. Gynecol. Endocrinol. 2006, 22, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Pritts, E.A.; Parker, W.H.; Olive, D.L. Fibroids and infertility: An updated systematic review of the evidence. Fertil. Steril. 2009, 91, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Sokol, N.S.; Ambros, V. Mesodermally expressed drosophila microRNA-1 is regulated by twist and is required in muscles during larval growth. Genes Dev. 2005, 19, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Poy, M.N.; Eliasson, L.; Krutzfeldt, J.; Kuwajima, S.; Ma, X.; MacDonald, P.E.; Pfeffer, S.; Tuschl, T.; Rajewsky, N.; Rorsman, P.; et al. A pancreatic islet-specific microRNA regulates insulin secretion. Nature 2004, 432, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Lai, E.C. MiRNAs: Whys and wherefores of miRNA-mediated regulation. Curr. Biol. 2005, 15, R458–R460. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Boutet, S.; Vazquez, F.; Liu, J.; Béclin, C.; Fagard, M.; Gratias, A.; Morel, J.B.; Crété, P.; Chen, X.; Vaucheret, H. Arabidopsis HEN1: A genetic link between endogenous miRNA controlling development and siRNA controlling transgene silencing and virus resistance. Curr. Biol. 2003, 13, 843–848. [Google Scholar] [CrossRef]
- Kim, Y.J.; Ku, S.Y.; Kim, Y.Y.; Liu, H.C.; Chi, S.W.; Kim, S.H.; Choi, Y.M.; Kim, J.G.; Moon, S.Y. MicroRNAs transfected into granulosa cells may regulate oocyte meiotic competence during in vitro maturation of mouse follicles. Hum. Reprod. 2013, 28, 3050–3061. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Ku, S.Y.; Rosenwaks, Z.; Liu, H.C.; Chi, S.W.; Kang, J.S.; Lee, W.J.; Jung, K.C.; Kim, S.H.; Choi, Y.M.; et al. MicroRNA expression profiles are altered by gonadotropins and vitamin c status during in vitro follicular growth. Reprod. Sci. 2010, 17, 1081–1089. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Ku, S.Y.; Kim, Y.Y.; Suh, C.S.; Kim, S.H.; Choi, Y.M. MicroRNA profile of granulosa cells after ovarian stimulation differs according to maturity of retrieved oocytes. Geburtshilfe Frauenheilkd. 2016, 76, 704–708. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.Y.; Min, H.; Kim, H.; Choi, Y.M.; Liu, H.C.; Ku, S.Y. Differential microRNA expression profile of human embryonic stem cell-derived cardiac lineage cells. Tissue Eng. Regen. Med. 2017, 14, 163–169. [Google Scholar] [CrossRef]
- Chuang, T.D.; Khorram, O. Expression profiling of lncRNAs, miRNAs, and mRNAs and their differential expression in leiomyoma using next-generation RNA sequencing. Reprod. Sci. 2018, 25, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Marsh, E.E.; Steinberg, M.L.; Parker, J.B.; Wu, J.; Chakravarti, D.; Bulun, S.E. Decreased expression of microRNA-29 family in leiomyoma contributes to increased major fibrillar collagen production. Fertil. Steril. 2016, 106, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Luo, X.; Chegini, N. MicroRNA 21: Response to hormonal therapies and regulatory function in leiomyoma, transformed leiomyoma and leiomyosarcoma cells. Mol. Hum. Reprod. 2010, 16, 215–227. [Google Scholar] [PubMed]
- Parker, W.H. Etiology, symptomatology, and diagnosis of uterine myomas. Fertil. Steril. 2007, 87, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Brady, P.C.; Stanic, A.K.; Styer, A.K. Uterine fibroids and subfertility: An update on the role of myomectomy. Curr. Opin. Obstet. Gynecol. 2013, 25, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Lagana, A.S.; Vergara, D.; Favilli, A.; La Rosa, V.L.; Tinelli, A.; Gerli, S.; Noventa, M.; Vitagliano, A.; Triolo, O.; Rapisarda, A.M.C.; et al. Epigenetic and genetic landscape of uterine leiomyomas: A current view over a common gynecological disease. Arch. Gynecol. Obstet. 2017, 296, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Vitale, S.G.; Sapia, F.; Rapisarda, A.M.C.; Valenti, G.; Santangelo, F.; Rossetti, D.; Chiofalo, B.; Sarpietro, G.; La Rosa, V.L.; Triolo, O.; et al. Hysteroscopic morcellation of submucous myomas: A systematic review. Biomed. Res. Int. 2017, 6848250. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.S.; Ku, S.Y.; Jee, B.C.; Suh, C.S.; Choi, Y.M.; Kim, J.G.; Moon, S.Y.; Kim, S.H. Comparison of follicular fluid IGF-i, IGF-ii, IGFBP-3, IGFBP-4 and PAPP-a concentrations and their ratios between gnrh agonist and GNRH antagonist protocols for controlled ovarian stimulation in ivf-embryo transfer patients. Hum. Reprod. 2006, 21, 2015–2021. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Ku, S.Y.; Jee, B.C.; Suh, C.S.; Kim, S.H.; Choi, Y.M.; Kim, J.G.; Moon, S.Y. A comparative study on the outcomes of in vitro fertilization between women with polycystic ovary syndrome and those with sonographic polycystic ovary-only in gnrh antagonist cycles. Arch. Gynecol. Obstet. 2010, 282, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.Y.; Suh, C.S.; Kim, S.H.; Choi, Y.M.; Kim, J.G.; Moon, S.Y. A pilot study of the use of low dose human menopausal gonadotropin in ovulation induction. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 109, 55–59. [Google Scholar] [CrossRef]
- Karmon, A.E.; Cardozo, E.R.; Rueda, B.R.; Styer, A.K. MicroRNAs in the development and pathobiology of uterine leiomyomata: Does evidence support future strategies for clinical intervention? Hum. Reprod. Update 2014, 20, 670–687. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, B.; Milev, I.; Minkov, I.; Dimitrova, I.; Bradford, A.P.; Baev, V. Characterization of the uterine leiomyoma microRNAome by deep sequencing. Genomics 2012, 99, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Chiofalo, B.; Lagana, A.S.; Vaiarelli, A.; La Rosa, V.L.; Rossetti, D.; Palmara, V.; Valenti, G.; Rapisarda, A.M.C.; Granese, R.; Sapia, F.; et al. Do miRNAs play a role in fetal growth restriction? a fresh look to a busy corner. Biomed. Res. Int. 2017, 2017, 6073167. [Google Scholar] [CrossRef] [PubMed]
- Lycoudi, A.; Mavreli, D.; Mavrou, A.; Papantoniou, N.; Kolialexi, A. MiRNAs in pregnancy-related complications. Expert. Rev. Mol. Diagn. 2015, 15, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Guo, L.; Zukerberg, L.; Rueda, B.R.; Styer, A.K. MicroRNA-15b regulates reversion-inducing cysteine-rich protein with kazal motifs (reck) expression in human uterine leiomyoma. Reprod. Biol. Endocrinol. 2016, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ling, J.; Fu, Z.; Ji, C.; Wu, J.; Xu, Q. Effects of miRNA-197 overexpression on proliferation, apoptosis and migration in levonorgestrel treated uterine leiomyoma cells. Biomed. Pharmacother. 2015, 71, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Chuang, T.D.; Khorram, O. Mir-200c regulates Il8 expression by targeting IKBKB: A potential mediator of inflammation in leiomyoma pathogenesis. PLoS ONE 2014, 9, e95370. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Zhang, D.; Du, R.; Pan, Y.; Zhao, L.; Sun, S.; Hong, L.; Liu, J.; Fan, D. MiR-15b and miR-16 modulate multidrug resistance by targeting bcl2 in human gastric cancer cells. Int. J. Cancer 2008, 123, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Takahashi, R.; Kondo, S.; Mizoguchi, A.; Adachi, E.; Sasahara, R.M.; Nishimura, S.; Imamura, Y.; Kitayama, H.; Alexander, D.B.; et al. The membrane-anchored mmp inhibitor reck is a key regulator of extracellular matrix integrity and angiogenesis. Cell 2001, 107, 789–800. [Google Scholar] [CrossRef]
- Wang, X.; Tang, S.; Le, S.Y.; Lu, R.; Rader, J.S.; Meyers, C.; Zheng, Z.M. Aberrant expression of oncogenic and tumor-suppressive microRNAs in cervical cancer is required for cancer cell growth. PLoS ONE 2008, 3, e2557. [Google Scholar] [CrossRef] [PubMed]
- Qiang, W.; Liu, Z.; Serna, V.A.; Druschitz, S.A.; Liu, Y.; Espona-Fiedler, M.; Wei, J.J.; Kurita, T. Down-regulation of mir-29b is essential for pathogenesis of uterine leiomyoma. Endocrinology 2014, 155, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Roslan, S.; Johnstone, C.N.; Wright, J.A.; Bracken, C.P.; Anderson, M.; Bert, A.G.; Selth, L.A.; Anderson, R.L.; Goodall, G.J.; et al. MiR-200 can repress breast cancer metastasis through zeb1-independent but moesin-dependent pathways. Oncogene 2014, 33, 4077–4088. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kurie, J.M.; Ahn, Y.H. Bmp4 Depletion by MiR-200 inhibits tumorigenesis and metastasis of lung adenocarcinoma cells. Mol. Cancer 2015, 14, 173. [Google Scholar] [CrossRef] [PubMed]
- Bogusiewicz, M.; Stryjecka-Zimmer, M.; Postawski, K.; Jakimiuk, A.J.; Rechberger, T. Activity of matrix metalloproteinase-2 and -9 and contents of their tissue inhibitors in uterine leiomyoma and corresponding myometrium. Gynecol. Endocrinol. 2007, 23, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Ishi, K.; Serna, V.A.; Kakazu, R.; Bulun, S.E.; Kurita, T. Progesterone is essential for maintenance and growth of uterine leiomyoma. Endocrinology 2010, 151, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Reierstad, S.; Demura, M.; Rademaker, A.W.; Kasai, T.; Inoue, M.; Usui, H.; Shozu, M.; Bulun, S.E. High aromatase expression in uterine leiomyoma tissues of African-American women. J. Clin. Endocrinol. Metab. 2009, 94, 1752–1756. [Google Scholar] [CrossRef] [PubMed]
- Cittelly, D.M.; Finlay-Schultz, J.; Howe, E.N.; Spoelstra, N.S.; Axlund, S.D.; Hendricks, P.; Jacobsen, B.M.; Sartorius, C.A.; Richer, J.K. Progestin suppression of mir-29 potentiates dedifferentiation of breast cancer cells via klf4. Oncogene 2013, 32, 2555–2564. [Google Scholar] [CrossRef] [PubMed]
- Vares, G.; Sai, S.; Wang, B.; Fujimori, A.; Nenoi, M.; Nakajima, T. Progesterone generates cancer stem cells through membrane progesterone receptor-triggered signaling in basal-like human mammary cells. Cancer Lett. 2015, 362, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Muluhngwi, P.; Richardson, K.; Napier, J.; Rouchka, E.C.; Mott, J.L.; Klinge, C.M. Regulation of miR-29b-1/a transcription and identification of target mRNAs in cho-k1 cells. Mol. Cell. Endocrinol. 2017, 444, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.Y.; Kim, Y.J.; Cho, K.M.; Kim, S.H.; Park, K.E.; Kang, B.C.; Jung, K.C.; Kim, M.S.; Ku, S.Y. The expression profile of angiotensin system on thawed murine ovaries. Tissue Eng. Regen. Med. 2016, 13, 724–731. [Google Scholar] [CrossRef]
- Kim, Y.J.; Kim, Y.Y.; Kim, D.W.; Joo, J.K.; Kim, H.; Ku, S.Y. Profile of microRNA expression in endometrial cell during in vitro culture according to progesterone concentration. Tissue Eng. Regen. Med. 2017, 14, 617–629. [Google Scholar] [CrossRef]
- Tian, R.; Wang, Z.; Shi, Z.; Li, D.; Wang, Y.; Zhu, Y.; Lin, W.; Gui, Y.; Zheng, X.L. Differential expression of g-protein-coupled estrogen receptor-30 in human myometrial and uterine leiomyoma smooth muscle. Fertil. Steril. 2013, 99, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, A.; Ohara, N.; Xu, Q.; Nakabayashi, K.; DeManno, D.A.; Chwalisz, K.; Yoshida, S.; Maruo, T. Selective progesterone receptor modulator asoprisnil down-regulates collagen synthesis in cultured human uterine leiomyoma cells through up-regulating extracellular matrix metalloproteinase inducer. Hum. Reprod. 2008, 23, 944–951. [Google Scholar] [CrossRef] [PubMed]
- Wolanska, M.; Sobolewski, K.; Bankowski, E.; Jaworski, S. Matrix metalloproteinases of human leiomyoma in various stages of tumor growth. Gynecol. Obstet. Investig. 2004, 58, 14–18. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Endometrial Cavity-Non-Distorting Leiomyoma (n = 15) | Endometrial Cavity-Distorting Leiomyoma (n = 11) |
|---|---|---|
| Age (years) | 32.5 ± 3.2 | 31.7 ± 4.3 |
| Size of leiomyoma (maximal diameter, cm) | 4.2 ± 1.3 | 4.5 ± 0.9 |
| Chief complaints | ||
| Dysmenorrhea | 7/15 (46.7%) | 8/11 (72.7%) |
| Menorrhagia | 9/15 (60.0%) | 9/11 (81.8%) |
| Infertility | 15/15 (100%) | 11/11 (100%) |
| Tissue Hardness * | 1+ | 2+ | 3+ | Total |
|---|---|---|---|---|
| Endometrial cavity-non-distorting leiomyoma (n) | 2 | 6 | 7 | 15 |
| Endometrial cavity-distorting leiomyoma (n) | 3 | 5 | 3 | 11 |
| miRNA | Sequence |
|---|---|
| U6 | gtgctcgcttcggcagcacatatac |
| miR-15b | tagcagcacatcatggtttaca |
| miR-29a | tagcaccatctgaaatcggtt |
| miR-29b | tagcaccatttgaaatcagtgtt |
| miR-29c | tagcaccatttgaaatcggt |
| miR-197 | ttcaccaccttctccacccagc |
| miR-200c | taatactgccgggtaatgatgga |
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| P-Rcα | GAGCACTGGATGCTGTTGCT | GGCTTAGGGCTTGGCTTTC |
| P-Rcβ | TGGGATCTGAGATCTTCGGAG | GAAGGGTCGGACTTCTGCTG |
| E-Rcα | TACTGACCAACCTGGCAGACAG | TGGACCTGATCATGGAGGGT |
| E-Rcβ | AGTTGGCCGACAAGGAGTTG | CGCACTTGGTCGAACAGG |
| MMP-1 | ACGGATACCCCAAGGACATCT | TCAGAAAGAGCATCGATATG |
| MMP-2 | GGACACACTAAAGAAGATGCAGAAGT | CGCATGGTCTCGATGGTATTC |
| MMP-9 | CCCGGAGTGAGTTGAACCA | GGATTTACATGGCACTGCC |
| TIMP-1 | CTGCGGATACTTCCACAGGTC | GCAAGAGTCCATCCTGCAGTT |
| TIMP-2 | ATAAGCAGGCCTCCAACGC | GAGCTGGACCAGTCGAAACC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.J.; Kim, Y.Y.; Shin, J.H.; Kim, H.; Ku, S.-Y.; Suh, C.S. Variation in MicroRNA Expression Profile of Uterine Leiomyoma with Endometrial Cavity Distortion and Endometrial Cavity Non-Distortion. Int. J. Mol. Sci. 2018, 19, 2524. https://doi.org/10.3390/ijms19092524
Kim YJ, Kim YY, Shin JH, Kim H, Ku S-Y, Suh CS. Variation in MicroRNA Expression Profile of Uterine Leiomyoma with Endometrial Cavity Distortion and Endometrial Cavity Non-Distortion. International Journal of Molecular Sciences. 2018; 19(9):2524. https://doi.org/10.3390/ijms19092524
Chicago/Turabian StyleKim, Yong Jin, Yoon Young Kim, Jung Ho Shin, Hoon Kim, Seung-Yup Ku, and Chang Suk Suh. 2018. "Variation in MicroRNA Expression Profile of Uterine Leiomyoma with Endometrial Cavity Distortion and Endometrial Cavity Non-Distortion" International Journal of Molecular Sciences 19, no. 9: 2524. https://doi.org/10.3390/ijms19092524
APA StyleKim, Y. J., Kim, Y. Y., Shin, J. H., Kim, H., Ku, S.-Y., & Suh, C. S. (2018). Variation in MicroRNA Expression Profile of Uterine Leiomyoma with Endometrial Cavity Distortion and Endometrial Cavity Non-Distortion. International Journal of Molecular Sciences, 19(9), 2524. https://doi.org/10.3390/ijms19092524