

Isolation of Allosteric Tryptase Inhibitor from Methanol Extract of Rhubarb and Enhancement of Its Tryptase Inhibitory Activity by Compounds That Were Screened by In Silico Screening
 , , , and
, , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. The Identification of a Tryptase Inhibitor in the Extract of Rhubarb (RHEI RHIZOMA) and an Analysis of Specificity of the Tryptase Inhibitor
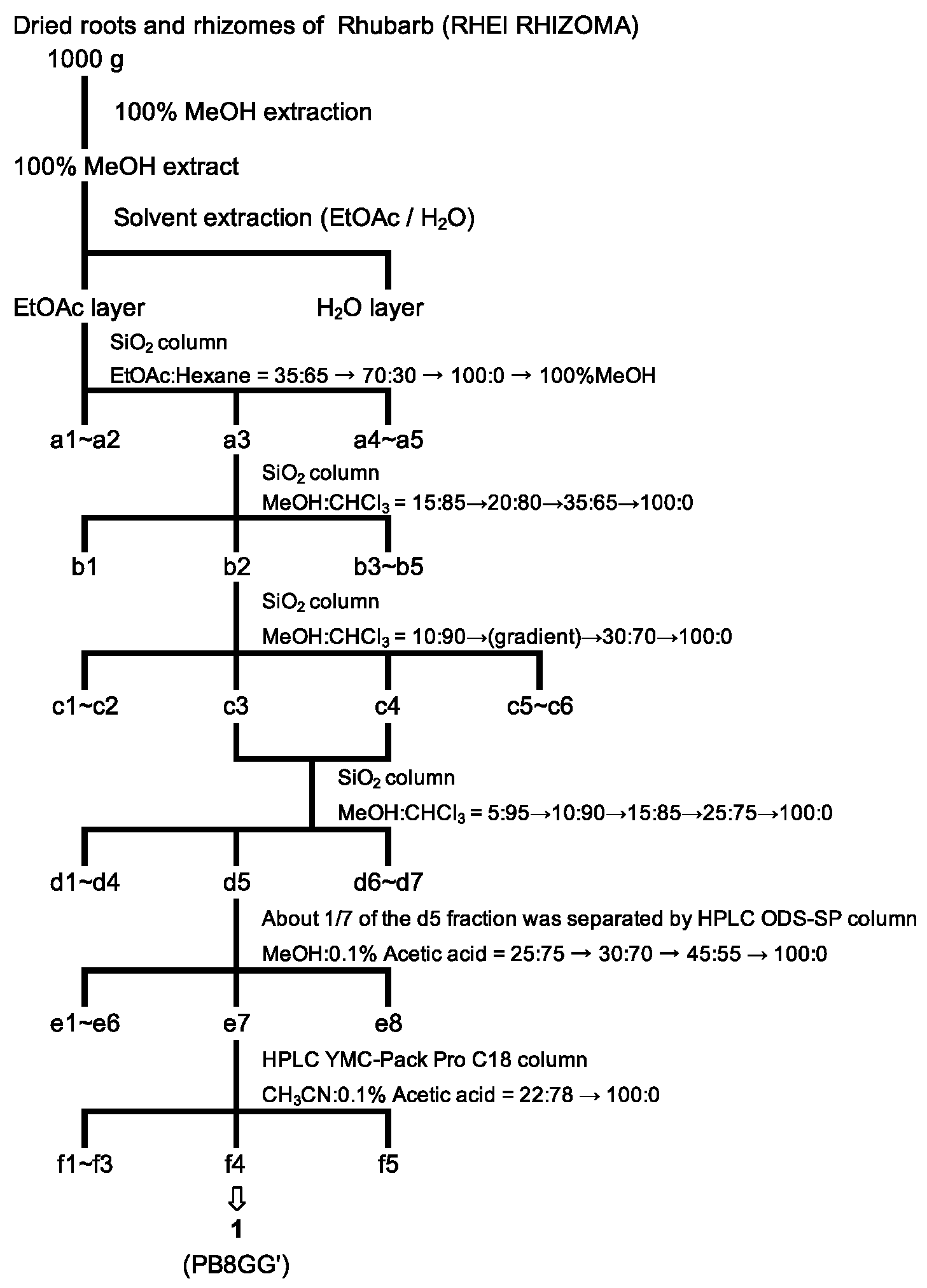
2.1.1. The Purification of a Tryptase Inhibitor from the Methanol Extract of Rhubarb
2.1.2. The Identification of the Tryptase Inhibitor in the Methanol Extract of Rhubarb
2.2. Mode of Tryptase Inhibition by PB8GG’ and Determination of Kinetic Parameters
2.2.1. Specific Inhibition of Tryptase by PB8GG’
2.2.2. Allosteric Inhibition of Tryptase by PB8GG’
2.3. In Silico Screening of the Other Tryptase Inhibitors Using AutoDock Vina
2.3.1. Carpinins B and E Are Candidate Compounds That Bind to Tryptase
2.3.2. Enhancement of the Tryptase Inhibitory Activity of PB8GG’ by Carpinins B and E
3. Discussion
4. Materials and Methods
4.1. Chemical Reagents and Instruments
4.2. Preparation of Extract of Rhubarb and Purification of Tryptase Inhibitors
4.3. Procyanidin B8 3,3′-Di-O-Gallate (1)
4.4. Measurement of Inhibitory Activity of the Inhibitor Against Tryptase and Trypsin
4.5. Kinetic Analysis of Tryptase Inhibition
4.6. Docking Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hogan, A.D.; Schwartz, L.B. Markers of mast cell degranulation. Methods 1997, 13, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, L.B.; Bradford, T.R. Regulation of tryptase from human lung mast cells by heparin. Stabilization of the active tetramer. J. Biol. Chem. 1986, 261, 7372–7379. [Google Scholar] [CrossRef] [PubMed]
- Vitte, J. Human mast cell tryptase in biology and medicine. Mol. Immunol. 2015, 63, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, H.; Abiko, Y.; Akimoto, T. A 50-year history of new drugs in Japan-the development and trends of hemostatics and antithrombotic drugs. Yakushigaku Zasshi 2003, 38, 93–105. [Google Scholar]
- Iwaki, M.; Ino, Y.; Motoyoshi, A.; Ozeki, M.; Sato, T.; Kurumi, M.; Aoyama, T. Pharmacological studies of FUT-175, nafamostat mesilate. V. Effects on the pancreatic enzymes and experimental acute pancreatitis in rats. Jpn. J. Pharmacol. 1986, 41, 155–162. [Google Scholar] [CrossRef]
- Mori, S.; Itoh, Y.; Shinohata, R.; Sendo, T.; Oishi, R.; Nishibori, M. Nafamostat mesilate is an extremely potent inhibitor of human tryptase. J. Pharmacol. Sci. 2003, 92, 420–423. [Google Scholar] [CrossRef]
- Pejler, G. The emerging role of mast cell proteases in asthma. Eur. Respir. J. 2019, 54, 1900685. [Google Scholar] [CrossRef]
- Maun, H.R.; Jackman, J.K.; Choy, D.F.; Loyet, K.M.; Staton, T.L.; Jia, G.; Dressen, A.; Hackney, J.A.; Bremer, M.; Walters, B.T.; et al. An Allosteric Anti-tryptase Antibody for the Treatment of Mast Cell-Mediated Severe Asthma. Cell 2019, 179, 417–431.e419. [Google Scholar] [CrossRef]
- Rhee, H.; Henderson, L.M.; Bauer, R.N.; Wong, K.; Staton, T.L.; Choy, D.F.; Banerjee, P.; Poon, V.; Yoshida, K.; Chen, C.; et al. Airway tryptase levels inform the lack of clinical efficacy of the tryptase inhibitor MTPS9579A in asthma. Allergy 2024, 79, 2993–3004. [Google Scholar] [CrossRef]
- Li, R.R.; Liu, X.F.; Feng, S.X.; Shu, S.N.; Wang, P.Y.; Zhang, N.; Li, J.S.; Qu, L.B. Pharmacodynamics of Five Anthraquinones (Aloe-emodin, Emodin, Rhein, Chysophanol, and Physcion) and Reciprocal Pharmacokinetic Interaction in Rats with Cerebral Ischemia. Molecules 2019, 24, 1898. [Google Scholar] [CrossRef]
- Hiraoka, A.; Koike, S.; Sakaguchi, S.; Masuda, Y.; Terai, F.; Miura, I.; Kawasaki, T.; Kawai, H.; Sakakibara, M. The sennoside constituents of rhei rhizoma and sennae folium as inhibitors of serum monoamine oxidase. Chem. Pharm. Bull. 1989, 37, 2744–2746. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein-ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef]
- Kojima, D.; Shimizu, K.; Aritake, K.; Era, M.; Matsuo, Y.; Saito, Y.; Tanaka, T.; Nonaka, G.I. Highly Oxidized Ellagitannins of Carpinus japonica and Their Oxidation-Reduction Disproportionation. J. Nat. Prod. 2020, 83, 3424–3434. [Google Scholar] [CrossRef]
- Sato, S.; Yamamoto, K.; Ito, M.; Nishino, K.; Otsuka, T.; Irie, K.; Nagao, M. Enhancement of Inhibitory Activity by Combining Allosteric Inhibitors Putatively Binding to Different Allosteric Sites on Cathepsin K. Molecules 2023, 28, 4197. [Google Scholar] [CrossRef]
- Esatbeyoglu, T.; Wray, V.; Winterhalter, P. Dimeric procyanidins: Screening for B1 to B8 and semisynthetic preparation of B3, B4, B6, And B8 from a polymeric procyanidin fraction of white willow bark (Salix alba). J. Agric. Food Chem. 2010, 58, 7820–7830. [Google Scholar] [CrossRef]
- Cahill, K.N.; Katz, H.R.; Cui, J.; Lai, J.; Kazani, S.; Crosby-Thompson, A.; Garofalo, D.; Castro, M.; Jarjour, N.; DiMango, E.; et al. KIT Inhibition by Imatinib in Patients with Severe Refractory Asthma. N. Engl. J. Med. 2017, 376, 1911–1920. [Google Scholar] [CrossRef]
- Hallgren, J.; Estrada, S.; Karlson, U.; Alving, K.; Pejler, G. Heparin antagonists are potent inhibitors of mast cell tryptase. Biochemistry 2001, 40, 7342–7349. [Google Scholar] [CrossRef]
- Ishizaki, M.; Tanaka, H.; Kajiwara, D.; Toyohara, T.; Wakahara, K.; Inagaki, N.; Nagai, H. Nafamostat mesilate, a potent serine protease inhibitor, inhibits airway eosinophilic inflammation and airway epithelial remodeling in a murine model of allergic asthma. J. Pharmacol. Sci. 2008, 108, 355–363. [Google Scholar] [CrossRef]
- Sendo, T.; Itoh, Y.; Goromaru, T.; Sumimura, T.; Saito, M.; Aki, K.; Yano, T.; Oishi, R. A potent tryptase inhibitor nafamostat mesilate dramatically suppressed pulmonary dysfunction induced in rats by a radiographic contrast medium. Br. J. Pharmacol. 2003, 138, 959–967. [Google Scholar] [CrossRef]
- Rice, K.D.; Tanaka, R.D.; Katz, B.A.; Numerof, R.P.; Moore, W.R. Inhibitors of tryptase for the treatment of mast cell-mediated diseases. Curr. Pharm. Des. 1998, 4, 381–396. [Google Scholar] [CrossRef] [PubMed]
- Kaneda, H.; Matsui, N.; Asano, Y.; Tabei, K.; Naganuma, S.; Koide, K.; Junko Toyama, T.; Nakamura, K.; Sekiguchi, T.; Suzuki, Y. Clinical evaluation of FUT-175 as a regional anti-coagulant in hemodialysis—Multi-center cooperative study. J. Jpn. Soc. Dial. Ther. 1987, 20, 951–963. [Google Scholar]
- Luo, W.; Su, X.; Gong, S.; Qin, Y.; Liu, W.; Li, J.; Yu, H.; Xu, Q. Anti-SARS coronavirus 3C-like protease effects of Rheum palmatum L. extracts. Biosci. Trends 2009, 3, 124–126. [Google Scholar] [PubMed]
- Addington, A.K.; Johnson, D.A. Inactivation of human lung tryptase: Evidence for a re-activatable tetrameric intermediate and active monomers. Biochemistry 1996, 35, 13511–13518. [Google Scholar] [CrossRef]
- Schechter, N.M.; Eng, G.Y.; Selwood, T.; McCaslin, D.R. Structural changes associated with the spontaneous inactivation of the serine proteinase human tryptase. Biochemistry 1995, 34, 10628–10638. [Google Scholar] [CrossRef]
- Niles, A.L.; Maffitt, M.; Haak-Frendscho, M.; Wheeless, C.J.; Johnson, D.A. Recombinant human mast cell tryptase beta: Stable expression in Pichia pastoris and purification of fully active enzyme. Biotechnol. Appl. Biochem. 1998, 28 Pt 2, 125–131. [Google Scholar] [CrossRef]
- Deeks, E.D. Asciminib: First Approval. Drugs 2022, 82, 219–226. [Google Scholar] [CrossRef]
- Hoy, S.M. Deucravacitinib: First Approval. Drugs 2022, 82, 1671–1679. [Google Scholar] [CrossRef]
- Nussinov, R.; Tsai, C.J. The different ways through which specificity works in orthosteric and allosteric drugs. Curr. Pharm. Des. 2012, 18, 1311–1316. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef]
- Ohtera, A.; Miyamae, Y.; Yoshida, K.; Maejima, K.; Akita, T.; Kakizuka, A.; Irie, K.; Masuda, S.; Kambe, T.; Nagao, M. Identification of a New Type of Covalent PPARγ Agonist using a Ligand-Linking Strategy. ACS Chem. Biol. 2015, 10, 2794–2804. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fujii, H.; Ito, M.; Nishioka, K.; Nishino, K.; Otsuka, T.; Irie, K.; Tanaka, T.; Nagao, M. Isolation of Allosteric Tryptase Inhibitor from Methanol Extract of Rhubarb and Enhancement of Its Tryptase Inhibitory Activity by Compounds That Were Screened by In Silico Screening. Molecules 2025, 30, 1341. https://doi.org/10.3390/molecules30061341
Fujii H, Ito M, Nishioka K, Nishino K, Otsuka T, Irie K, Tanaka T, Nagao M. Isolation of Allosteric Tryptase Inhibitor from Methanol Extract of Rhubarb and Enhancement of Its Tryptase Inhibitory Activity by Compounds That Were Screened by In Silico Screening. Molecules. 2025; 30(6):1341. https://doi.org/10.3390/molecules30061341
Chicago/Turabian StyleFujii, Hidetoshi, Moeno Ito, Kentaro Nishioka, Katsutoshi Nishino, Takanao Otsuka, Kazuhiro Irie, Takashi Tanaka, and Masaya Nagao. 2025. "Isolation of Allosteric Tryptase Inhibitor from Methanol Extract of Rhubarb and Enhancement of Its Tryptase Inhibitory Activity by Compounds That Were Screened by In Silico Screening" Molecules 30, no. 6: 1341. https://doi.org/10.3390/molecules30061341
APA StyleFujii, H., Ito, M., Nishioka, K., Nishino, K., Otsuka, T., Irie, K., Tanaka, T., & Nagao, M. (2025). Isolation of Allosteric Tryptase Inhibitor from Methanol Extract of Rhubarb and Enhancement of Its Tryptase Inhibitory Activity by Compounds That Were Screened by In Silico Screening. Molecules, 30(6), 1341. https://doi.org/10.3390/molecules30061341