


Influence of the Casein Genotype on Goat Milk Bioactivity: An In Silico Analysis of the Casein Peptidome
 ,
,  ,
,
Abstract
1. Introduction
2. Results
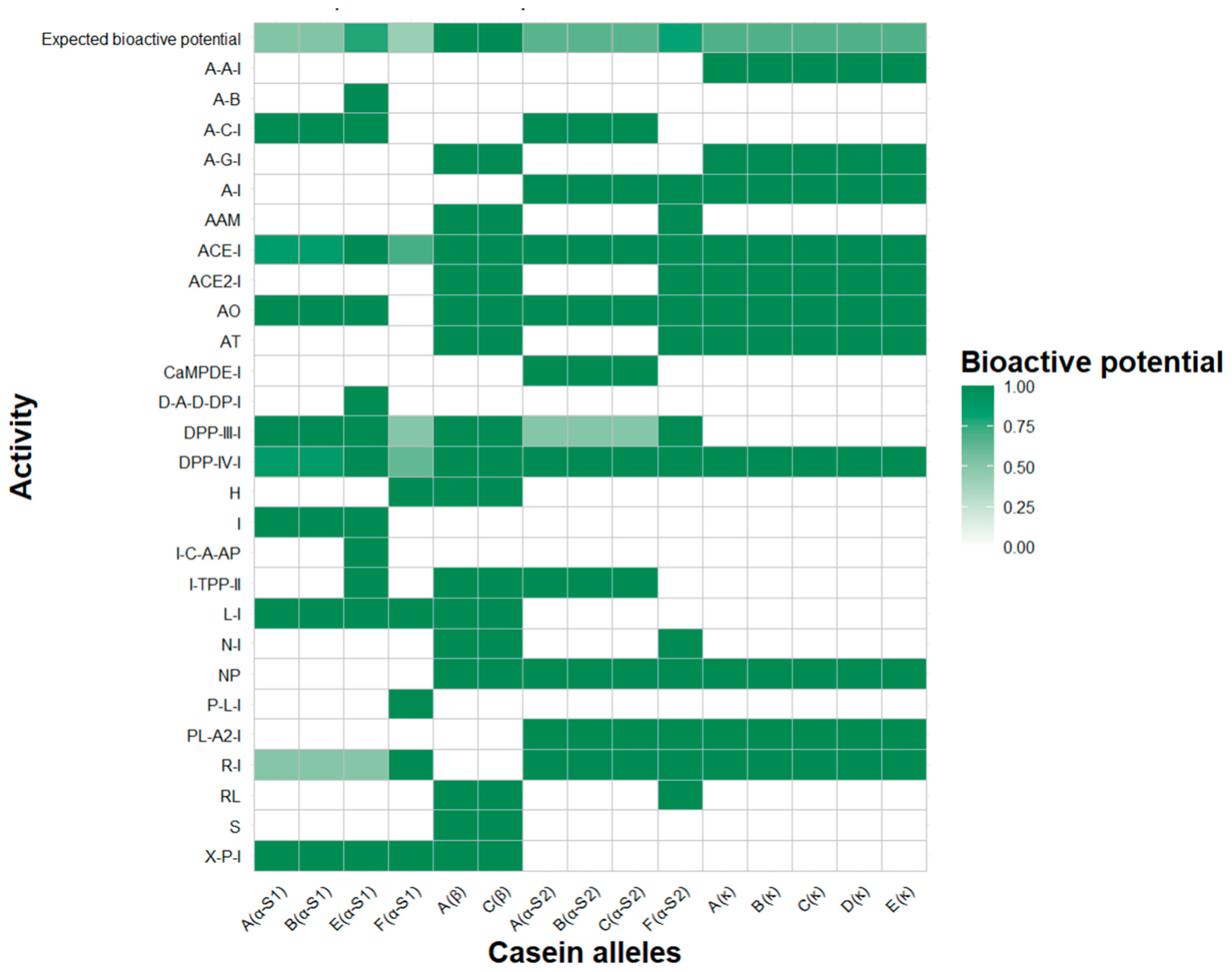
2.1. Analysis of Caseins as a Source of Bioactive Peptides on the Basis of Specific Genotypes
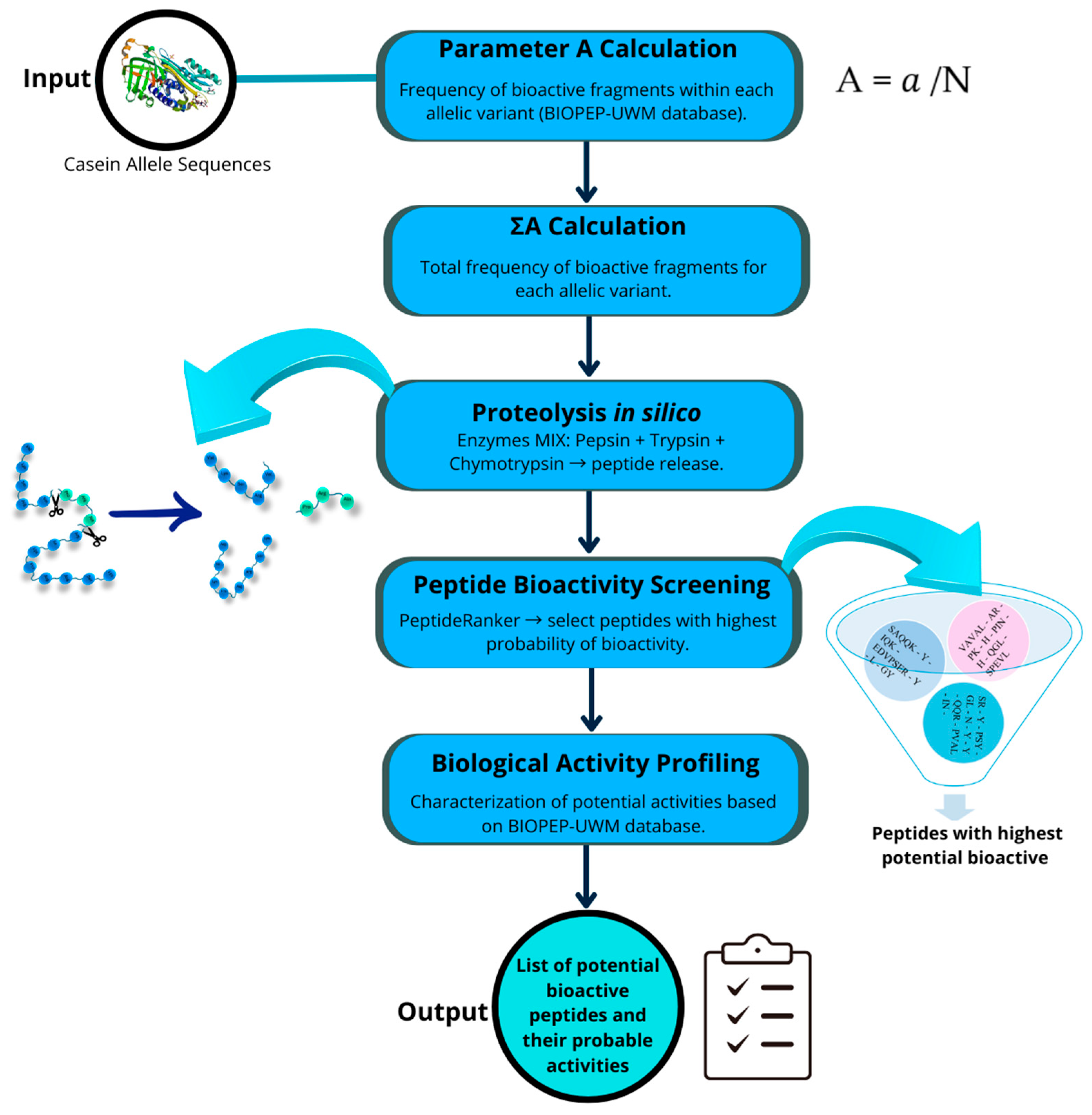
2.2. In Silico Proteolytic Release and Prediction of Bioactive Functions
3. Discussion
4. Materials and Methods
4.1. Analysis of Capra Hircus Caseins as a Source of Bioactive Peptides
- a = Number of fragments with a specific activity in a protein sequence;
- N = Total number of amino acid residues in the protein.
4.2. In Silico Proteolytic Release and Prediction of Bioactive Functions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ∑ Functional Detected Activities | α-S1-Casein | Β-Casein | α-S2-Casein | κ-Casein | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Allele A | Allele B | Allele E | Allele F | Allele A | Allele C | Allele A | Allele B | Allele C | Allele F | Allele A | Allele B | Allele C | Allele D | Allele E | |
| Alpha-amylase inhibitor | 0.0093 | 0.0093 | 0.0093 | 0 | 0 | 0 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| ACE 2 activator | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| ACE inhibitor | 0.5794 | 0.5794 | 0.5841 | 0.4609 | 0.6441 | 0.638 | 0.4888 | 0.4843 | 0.4798 | 0.4888 | 0.5802 | 0.5864 | 0.5864 | 0.5926 | 0.6049 |
| ACE2 inhibitor | 0.0047 | 0.0047 | 0.0047 | 0.0087 | 0.0315 | 0.0317 | 0 | 0 | 0 | 0.0045 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Activating ubiquitin-mediated proteolysis | 0.0093 | 0.0093 | 0.0093 | 0.0087 | 0.0135 | 0.0136 | 0.009 | 0.009 | 0.009 | 0.009 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Alanine carboxypeptidase inhibitor | 0.0047 | 0.0047 | 0.0047 | 0 | 0.0045 | 0.0045 | 0.009 | 0.009 | 0.009 | 0.0045 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Alpha-glucosidase inhibitor | 0.0374 | 0.0374 | 0.0327 | 0.0261 | 0.0405 | 0.0407 | 0.0179 | 0.0179 | 0.0179 | 0.0179 | 0.0494 | 0.0494 | 0.0556 | 0.0556 | 0.0494 |
| Anti inflammatory | 0 | 0 | 0 | 0 | 0.0045 | 0.0045 | 0.0135 | 0.0135 | 0.0135 | 0.0135 | 0.0185 | 0.0185 | 0.0185 | 0.0185 | 0.0185 |
| Anti-apoptotic | 0 | 0 | 0 | 0 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Antiamnestic | 0.0047 | 0.0047 | 0.0047 | 0 | 0.018 | 0.0181 | 0.0045 | 0.0045 | 0.0045 | 0.009 | 0.0123 | 0.0123 | 0.0123 | 0.0123 | 0.0123 |
| Antibacterial | 0.0093 | 0.0093 | 0.0047 | 0 | 0.009 | 0.009 | 0.0179 | 0.0179 | 0.009 | 0.0179 | 0.0185 | 0.0185 | 0.0185 | 0.0185 | 0.0185 |
| Anticancer | 0.0187 | 0.0187 | 0.0187 | 0.0087 | 0.009 | 0.009 | 0.009 | 0.009 | 0.0045 | 0.009 | 0 | 0 | 0 | 0 | 0 |
| Antidiabetic | 0.0047 | 0.0047 | 0.0047 | 0 | 0.009 | 0.009 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 |
| Antifungal | 0.0047 | 0.0047 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Antioxidative | 0.243 | 0.243 | 0.2336 | 0.0783 | 0.1622 | 0.1629 | 0.139 | 0.139 | 0.139 | 0.139 | 0.1914 | 0.1914 | 0.1914 | 0.1914 | 0.1914 |
| Antithrombotic | 0 | 0 | 0 | 0 | 0.0135 | 0.0136 | 0.009 | 0.009 | 0.009 | 0.0135 | 0.037 | 0.037 | 0.037 | 0.037 | 0.037 |
| Antiviral | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Bacterial permease ligand | 0.0047 | 0.0047 | 0.0047 | 0 | 0.0045 | 0.0045 | 0.0135 | 0.0135 | 0.009 | 0.0135 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Binding | 0.0047 | 0.0047 | 0.0047 | 0 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Calpain 1 inhibitor | 0.0047 | 0.0047 | 0.0047 | 0.0087 | 0 | 0 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 |
| CaMPDE inhibitor | 0 | 0 | 0 | 0 | 0.0045 | 0.0045 | 0.0224 | 0.0224 | 0.0224 | 0.0179 | 0 | 0 | 0 | 0 | 0 |
| Cathepsin B inhibitor | 0 | 0 | 0 | 0 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Citrate lyase deacetylase inhibitor | 0.0093 | 0.0093 | 0.0093 | 0 | 0.0135 | 0.0136 | 0.0404 | 0.0404 | 0.0404 | 0.0404 | 0 | 0 | 0 | 0 | 0 |
| Contracting | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0247 | 0.0247 | 0.0247 | 0.0247 | 0.0247 |
| D-Ala-D-Ala dipeptidase inhibitor | 0 | 0 | 0.0047 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Dipeptidyl peptidase III inhibitor | 0.1075 | 0.1075 | 0.1075 | 0.0957 | 0.0946 | 0.0905 | 0.0717 | 0.0717 | 0.0717 | 0.0762 | 0.0741 | 0.0741 | 0.0802 | 0.0864 | 0.0741 |
| Dipeptidyl peptidase IV inhibitor | 0.6168 | 0.6168 | 0.6075 | 0.5826 | 0.8514 | 0.8462 | 0.6771 | 0.6682 | 0.6771 | 0.6771 | 0.7778 | 0.784 | 0.7778 | 0.7778 | 0.7778 |
| Glutamate carboxypeptidase II inhibitor | 0.028 | 0.0327 | 0.0327 | 0.0087 | 0.0225 | 0.0226 | 0.0583 | 0.0538 | 0.0583 | 0.0583 | 0.0185 | 0.0185 | 0.0185 | 0.0185 | 0.0185 |
| Glutamate carboxypeptidase inhibitor | 0.0093 | 0.0093 | 0.0093 | 0 | 0.0225 | 0.0226 | 0.0404 | 0.0404 | 0.0404 | 0.0404 | 0 | 0 | 0 | 0 | 0 |
| HMG-CoA reductase inhibitor | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Hypotensive | 0.0187 | 0.0187 | 0.0234 | 0.0087 | 0.0225 | 0.0226 | 0.009 | 0.009 | 0.009 | 0.0135 | 0.0185 | 0.0185 | 0.0185 | 0.0185 | 0.0185 |
| Hypouricemic | 0.0327 | 0.0327 | 0.0234 | 0.0348 | 0.036 | 0.0362 | 0.0179 | 0.0179 | 0.0179 | 0.0179 | 0.0494 | 0.0494 | 0.0494 | 0.0494 | 0.0494 |
| Immunomodulating | 0.0047 | 0.0047 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Immunostimulating | 0.0047 | 0.0047 | 0.0047 | 0 | 0.009 | 0.009 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Inhibitor | 0.0047 | 0.0047 | 0.0047 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Inhibitor of cytosol alanyl aminopeptidase | 0.014 | 0.014 | 0.0187 | 0.0174 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 |
| Inhibitor of tripeptidyl peptidase II | 0.0374 | 0.0374 | 0.0421 | 0.0522 | 0.0315 | 0.0362 | 0.0269 | 0.0269 | 0.0269 | 0.0224 | 0.0185 | 0.0185 | 0.0185 | 0.0185 | 0.0185 |
| Lactocepin inhibitor | 0.028 | 0.028 | 0.028 | 0.0261 | 0.045 | 0.0452 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0123 | 0.0123 | 0.0123 | 0.0123 | 0.0123 |
| Leucyltransferase inhibitor | 0.0047 | 0.0047 | 0.0047 | 0.0087 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Lipoxygenase inhibitor | 0 | 0 | 0 | 0 | 0.009 | 0.009 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Neprilysin 2 inhibitor | 0.0047 | 0.0047 | 0.0047 | 0.0087 | 0 | 0 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 |
| Neprilysin inhibitor | 0.0327 | 0.0374 | 0.0374 | 0.0348 | 0.0586 | 0.0633 | 0.0314 | 0.0269 | 0.0269 | 0.0359 | 0.0617 | 0.0617 | 0.0617 | 0.0617 | 0.0617 |
| Neurolysin inhibitor | 0.0047 | 0.0047 | 0.0047 | 0.0087 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Neuropeptide | 0.0374 | 0.0374 | 0.0374 | 0.0174 | 0.018 | 0.0181 | 0.0224 | 0.0224 | 0.0224 | 0.0224 | 0.0185 | 0.0185 | 0.0185 | 0.0247 | 0.0247 |
| Opioid | 0.014 | 0.014 | 0.014 | 0 | 0.0135 | 0.0136 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 | 0 |
| Opioid agonist | 0.0047 | 0.0047 | 0.0047 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Opioid antagonist | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0.0247 | 0.0247 | 0.0247 | 0.0247 | 0.0247 |
| Osteoanabolic | 0.0093 | 0.0093 | 0.0093 | 0 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0.0123 | 0.0123 | 0.0123 | 0.0123 | 0.0123 |
| Pancreatic lipase inhibitor | 0 | 0 | 0 | 0.0087 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0 | 0 | 0 |
| Peptidylprolyl isomerase inhibitor | 0.0093 | 0.0093 | 0.0093 | 0.0087 | 0 | 0 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0 | 0 | 0.0062 | 0.0062 | 0 |
| Phospholipase A2 inhibitor | 0 | 0 | 0 | 0 | 0 | 0 | 0.009 | 0.009 | 0.009 | 0.009 | 0.0123 | 0.0123 | 0.0123 | 0.0123 | 0.0123 |
| Pseudolysin inhibitor | 0 | 0 | 0 | 0 | 0 | 0 | 0.0045 | 0.0045 | 0.0045 | 0.0045 | 0.0062 | 0.0062 | 0.0062 | 0.0062 | 0.0062 |
| Regulating | 0.0047 | 0.0047 | 0.0047 | 0 | 0.036 | 0.0362 | 0.0045 | 0.0045 | 0.0045 | 0.009 | 0 | 0 | 0 | 0 | 0 |
| Renin inhibitor | 0.028 | 0.028 | 0.0234 | 0.0261 | 0.0315 | 0.0317 | 0.0448 | 0.0448 | 0.0448 | 0.0448 | 0.0185 | 0.0185 | 0.0185 | 0.0185 | 0.0185 |
| Stimulating | 0.0888 | 0.0888 | 0.0841 | 0.0696 | 0.1036 | 0.0995 | 0.0852 | 0.0852 | 0.0897 | 0.0852 | 0.037 | 0.037 | 0.037 | 0.0309 | 0.037 |
| Thymidylate synthase inhibitor | 0.0093 | 0.0093 | 0.0093 | 0 | 0.0135 | 0.0136 | 0.0404 | 0.0404 | 0.0404 | 0.0404 | 0 | 0 | 0 | 0 | 0 |
| Toxic | 0.0093 | 0.0093 | 0.0093 | 0.0087 | 0 | 0 | 0.009 | 0.009 | 0.0045 | 0.009 | 0 | 0 | 0 | 0 | 0 |
| Tubulin-tyrosine ligase inhibitor | 0.014 | 0.014 | 0.014 | 0 | 0 | 0 | 0.009 | 0.009 | 0.009 | 0.009 | 0.0247 | 0.0247 | 0.0247 | 0.0247 | 0.0247 |
| Xaa-pro inhibitor | 0.0093 | 0.0093 | 0.0093 | 0.0087 | 0.018 | 0.0181 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ∑A | 2.1447 | 2.1541 | 2.1216 | 1.6351 | 2.4500 | 2.4429 | 1.9959 | 1.9735 | 1.9645 | 2.0094 | 2.1914 | 2.2038 | 2.2161 | 2.2286 | 2.2223 |
Appendix B

Appendix C
| No | Peptide | Activity Prediction | No | Peptide | Activity Prediction | No | Peptide | Activity Prediction |
|---|---|---|---|---|---|---|---|---|
| 1 | IL | 0.392515 | 59 | TTM | 0.137765 | 117 | AN | 0.156915 |
| 2 | TCL | 0.47708 | 60 | AAM | 0.55858 | 118 | EEEY | 0.0347581 |
| 3 | VAVAL | 0.114473 | 61 | VL | 0.126025 | 119 | SIR | 0.292181 |
| 4 | AR | 0.394584 | 62 | ACL | 0.746733 | 120 | SSSEESAEVAPEEIK | 0.128476 |
| 5 | PK | 0.325772 | 63 | VAL | 0.127235 | 121 | ITVDDK | 0.0507604 |
| 6 | PIN | 0.392694 | 64 | AL | 0.43789 | 122 | EIN | 0.0574424 |
| 7 | QGL | 0.533495 | 65 | EQEEL | 0.0476195 | 123 | PQY | 0.465473 |
| 8 | SPEVL | 0.240941 | 66 | VVGETVESL | 0.044042 | 124 | QY | 0.233017 |
| 9 | SPEVPN | 0.288857 | 67 | SSSEESITH | 0.123839 | 125 | PY | 0.736696 |
| 10 | EN | 0.0363788 | 68 | IEK | 0.0431926 | 126 | QGPIVL | 0.474397 |
| 11 | VVAPF | 0.482403 | 69 | QSEEQQQTEDEL | 0.0562204 | 127 | PW | 0.992911 |
| 12 | PEVF | 0.416246 | 70 | QDK | 0.0742183 | 128 | DQVK | 0.0569847 |
| 13 | IN | 0.152106 | 71 | IH | 0.213873 | 129 | AGAF | 0.873661 |
| 14 | EL | 0.0728272 | 72 | PF | 0.99343 | 130 | AGPF | 0.969222 |
| 15 | SK | 0.0732708 | 73 | AQAQSL | 0.187449 | 131 | TPTVN | 0.0691266 |
| 16 | DIGSESTEDQAM | 0.118262 | 74 | VY | 0.0989681 | 132 | STSEEN | 0.0452869 |
| 17 | EDAK | 0.0508357 | 75 | TGPIPN | 0.416883 | 133 | TIDM | 0.226127 |
| 18 | QM | 0.607122 | 76 | SL | 0.330018 | 134 | ESTEVF | 0.118496 |
| 19 | AGSSSSSEEIVPN | 0.150856 | 77 | PQN | 0.285353 | 135 | TEEEK | 0.0196166 |
| 20 | AGSSCQVR | 0.361516 | 78 | TQTPVVVPPF | 0.213655 | 136 | ISQY | 0.183534 |
| 21 | PIVL | 0.3331 | 79 | QPEIM | 0.368217 | 137 | AW | 0.9669 |
| 22 | SR | 0.308189 | 80 | GVPK | 0.24186 | 138 | TVDQH | 0.048737 |
| 23 | STF | 0.58977 | 81 | VK | 0.03329 | 139 | AM | 0.74549 |
| 24 | CPL | 0.916551 | 82 | ETM | 0.0952962 | 140 | TQPK | 0.117696 |
| 25 | SVTW | 0.38436 | 83 | VPK | 0.135867 | 141 | TN | 0.0512668 |
| 26 | VIW | 0.610494 | 84 | EM | 0.194256 | 142 | AIPY | 0.613395 |
| 27 | SF | 0.948796 | 85 | PVEPF | 0.604782 | 143 | VR | 0.114691 |
| 28 | SD | 0.102577 | 86 | TESQSL | 0.0615565 | 144 | CCEK | 0.328494 |
| 29 | SAQQK | 0.115388 | 87 | TL | 0.144542 | 145 | DER | 0.0799595 |
| 30 | SAEQK | 0.0721709 | 88 | TDVEK | 0.0283043 | 146 | DDK | 0.0807603 |
| 31 | IQK | 0.0864674 | 89 | VQSW | 0.328419 | 147 | IAK | 0.112126 |
| 32 | EDVPSER | 0.0960623 | 90 | QPPQPL | 0.823819 | 148 | IPIQY | 0.360781 |
| 33 | GY | 0.741592 | 91 | SPTVM | 0.441571 | 149 | PSY | 0.46941 |
| 34 | EQL | 0.0721403 | 92 | PPQSVL | 0.447296 | 150 | GL | 0.808777 |
| 35 | VPQL | 0.306426 | 93 | SQPK | 0.227354 | 151 | QQR | 0.201093 |
| 36 | EIVPK | 0.0893465 | 94 | PVPQK | 0.223551 | 152 | PVAL | 0.277634 |
| 37 | SAEEQL | 0.104281 | 95 | AVPQR | 0.253856 | 153 | PVAVR | 0.134071 |
| 38 | SM | 0.628268 | 96 | VVPQR | 0.140161 | 154 | PIAVR | 0.239292 |
| 39 | EGN | 0.094901 | 97 | DM | 0.607466 | 155 | SPAQTL | 0.459196 |
| 40 | PAH | 0.432769 | 98 | PIQAF | 0.749093 | 156 | QW | 0.928524 |
| 41 | QK | 0.0619833 | 99 | QEPVL | 0.19215 | 157 | QVL | 0.120783 |
| 42 | QPM | 0.810167 | 100 | GPVR | 0.488393 | 158 | PN | 0.480465 |
| 43 | IAVN | 0.0822104 | 101 | GPF | 0.989324 | 159 | TVPAK | 0.0926354 |
| 44 | QEL | 0.0839476 | 102 | PIL | 0.641797 | 160 | SCQDQPTTL | 0.428259 |
| 45 | AY | 0.346246 | 103 | IF | 0.949173 | 161 | PH | 0.541688 |
| 46 | PQL | 0.527882 | 104 | AVAL | 0.158386 | 162 | AIPPK | 0.575607 |
| 47 | QF | 0.946135 | 105 | AK | 0.0979062 | 163 | DQDK | 0.0824949 |
| 48 | QL | 0.2924 | 106 | EH | 0.0533694 | 164 | TEVPAIN | 0.0754348 |
| 49 | DAY | 0.234477 | 107 | VSSSEEPIN | 0.148998 | 165 | TEIPAIN | 0.108978 |
| 50 | PSGAW | 0.870583 | 108 | ISSSEEPIN | 0.19248 | 166 | TIASAEPTVH | 0.0919084 |
| 51 | PL | 0.811148 | 109 | QEIY | 0.12052 | 167 | STPTTEAIVN | 0.0575411 |
| 52 | GTQY | 0.184477 | 110 | QEK | 0.0353735 | 168 | TVDN | 0.0382016 |
| 53 | TDAPSF | 0.417727 | 111 | AIH | 0.212314 | 169 | PEASSESIASASETN | 0.131678 |
| 54 | SDIPN | 0.394725 | 112 | PR | 0.787626 | 170 | PEASSESIVSAPETN | 0.126334 |
| 55 | PIGSEN | 0.146949 | 113 | EK | 0.0247356 | 171 | TAQVTSTEV | 0.0300416 |
| 56 | SGK | 0.220066 | 114 | CTTSCEEVVR | 0.0950641 | 172 | SCQGQPTTL | 0.424559 |
| 57 | SSSEESAKVAPEEIK | 0.167177 | 115 | IISQY | 0.162259 | |||
| 58 | TK | 0.0329947 | 116 | QR | 0.274416 |
References
- Nayik, G.A.; Jagdale, Y.D.; Gaikwad, S.A.; Devkatte, A.N.; Dar, A.H.; Ansari, M.J. Nutritional profile, processing and potential products: A comparative review of goat milk. Dairy 2022, 3, 622–647. [Google Scholar] [CrossRef]
- Rahmatalla, S.A.; Arends, D.; Brockmann, G.A. Genetic and protein variants of milk caseins in goats. Front. Genet. 2022, 13, 995349. [Google Scholar] [CrossRef] [PubMed]
- Marletta, D.; Criscione, A.; Bordonaro, S.; Guastella, A.M.; d’Urso, G. Casein polymorphism in goat’s milk. Dairy Sci. Technol. 2007, 87, 491–504. [Google Scholar] [CrossRef]
- Pal, S.; Woodford, K.; Kukuljan, S.; Ho, S. Milk intolerance, beta-casein and lactose. Nutrients 2015, 7, 7285–7297. [Google Scholar] [CrossRef]
- Dantas, A.; Kumar, H.; Prudencio, E.S.; de Avila Junior, L.B.; Orellana-Palma, P.; Dosoky, N.S.; Kumar, D. An approach on detection, quantification, technological properties, and trends market of A2 cow milk. Food Res. Int. 2023, 167, 112690. [Google Scholar] [CrossRef]
- Giribaldi, M.; Lamberti, C.; Cirrincione, S.; Giuffrida, M.G.; Cavallarin, L. A2 milk and BCM-7 peptide as emerging parameters of milk quality. Front. Nutr. 2022, 9, 842375. [Google Scholar] [CrossRef]
- Kashung, P.; Karuthapandian, D. Milk-derived bioactive peptides. Food Prod. Process Nutr. 2025, 7, 6. [Google Scholar] [CrossRef]
- Ouafi, A.T.; Babilliot, J.M.; Leroux, C.; Martin, P. Genetic diversity of the two main Moroccan goat breeds: Phylogenetic relationships with four breeds reared in France. Small Rumin. Res. 2002, 45, 225–233. [Google Scholar] [CrossRef]
- Maga, E.A.; Daftari, P.; Kültz, D.; Penedo, M.C.T. Prevalence of αs1-casein genotypes in American dairy goats. J. Anim. Sci. 2009, 87, 3464–3469. [Google Scholar] [CrossRef]
- Chessa, S.; Budelli, E.; Chiatti, F.; Cito, A.M.; Bolla, P.; Caroli, A. Predominance of β-casein (CSN2) C allele in goat breeds reared in Italy. Journal of dairy science. J. Dairy Sci. 2005, 88, 1878–1881. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Ahmed, A.S.; Miyata, T. Novel angiotensin-converting enzyme inhibitory peptides from caseins and whey proteins of goat milk. J. Adv. Res. 2017, 8, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Rout, P.K.; Kaushik, R.; Singh, G. Identification of bioactive peptides in goat milk and their health application. J. Adv. Dairy Res. 2017, 5, 1000191. [Google Scholar] [CrossRef]
- Guha, S.; Sharma, H.; Deshwal, G.K.; Rao, P.S. A comprehensive review on bioactive peptides derived from milk and milk products of minor dairy species. Food Prod. Process. Nutr. 2021, 3, 2. [Google Scholar] [CrossRef]
- Dos Santos, W.M.; Gomes, A.C.G.; de Caldas Nobre, M.S.; de Souza Pereira, Á.M.; dos Santos Pereira, E.V.; dos Santos, K.M.O.; Buriti, F.C.A. Goat milk as a natural source of bioactive compounds and strategies to enhance the amount of these beneficial components. Int. Dairy J. 2023, 137, 105515. [Google Scholar] [CrossRef]
- Dave, L.A.; Montoya, C.A.; Rutherfurd, S.M.; Moughan, P.J. Gastrointestinal endogenous proteins as a source of bioactive peptides-an in silico study. PLoS ONE 2014, 9, e98922. [Google Scholar] [CrossRef]
- Lin, K.; Zhang, L.W.; Han, X.; Xin, L.; Meng, Z.X.; Gong, P.M.; Cheng, D.Y. Yak milk casein as potential precursor of angiotensin I-converting enzyme inhibitory peptides based on in silico proteolysis. Food Chem. 2018, 254, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Li, X.; Qi, X.; Ma, Y.; Chan, E.C.Y. In silico identification of novel ACE and DPP-IV inhibitory peptides derived from buffalo milk proteins and evaluation of their inhibitory mechanisms. Amino Acids 2023, 55, 161–171. [Google Scholar] [CrossRef]
- Sundarraj, A.A.; Rajathi, A.A.; Vishaal, S.C.; Rohit, D.; Prakash, M.S.; Sam, A.A.; Seihenbalg, S.S. Food biotechnology applications in dairy and dairy products. J. Pharm. Res. 2018, 12, 520–525. [Google Scholar]
- Balthazar, C.F.; Pimentel, T.C.; Ferrão, L.L.; Almada, C.N.; Santillo, A.; Albenzio, M.; Cruz, A.G. Sheep milk: Physicochemical characteristics and relevance for functional food development. Compr. Rev. Food Sci. Food Saf. 2017, 16, 247–262. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. Strategies for the discovery and identification of food protein-derived biologically active peptides. Trends Food Sci. Technol. 2017, 69, 289–305. [Google Scholar] [CrossRef]
- Cui, Q.; Duan, Y.; Zhang, M.; Liang, S.; Sun, Y.; Cheng, J.; Guo, M. Peptide profiles and antioxidant capacity of extensive hydrolysates of milk protein concentrate. J. Dairy Sci. 2022, 105, 7972–7985. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, J.; Buretić-Tomljanović, A. Peptidomics as a tool for characterizing bioactive milk peptides. Food Chem. 2017, 230, 91–98. [Google Scholar] [CrossRef]
- Yang, M.; Yang, Z.; Everett, D.W.; Gilbert, E.P.; Singh, H.; Ye, A. Digestion of food proteins: The role of pepsin. Crit. Rev. Food Sci. Nutr. 2025, 1–22. [Google Scholar] [CrossRef]
- Martin, P.; Ollivier-Bousquet, M.; Grosclaude, F. Genetic polymorphism of caseins: A tool to investigate casein micelle organization. Int. Dairy J. 1999, 9, 163–171. [Google Scholar] [CrossRef]
- Ramunno, L.; Cosenza, G.; Rando, A.; Pauciullo, A.; Illario, R.; Gallo, D.; Masina, P. Comparative analysis of gene sequence of goat CSN1S1 F and N alleles and characterization of CSN1S1 transcript variants in mammary gland. Gene 2005, 345, 289–299. [Google Scholar] [CrossRef]
- Lunagariya, N.A.; Patel, N.K.; Jagtap, S.C.; Bhutani, K.K. Inhibitors of pancreatic lipase: State of the art and clinical perspectives. EXCLI J. 2014, 13, 897–921. [Google Scholar] [PubMed]
- Udenigwe, C.C.; Aluko, R.E. Multifunctional cationic peptide fractions from flaxseed protein hydrolysates. Plant Foods Hum. Nutr. 2012, 67, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Zhang, Y.J.; Zhou, F.; Ge, Y.; Zhao, M.H.; Liu, Y.; Yang, K.W. Real-time monitoring of D-Ala-D-Ala dipeptidase activity of VanX in living bacteria by isothermal titration calorimetry. Anal. Biochem. 2019, 578, 29–35. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Cadamuro, C.; Le Gouic, A.; Mudgil, P.; Maqsood, S.; FitzGerald, R.J. Dipeptidyl peptidase IV (DPP-IV) inhibitory properties of a camel whey protein enriched hydrolysate preparation. Food Chem. 2019, 279, 70–79. [Google Scholar] [CrossRef]
- Daliri, E.B.M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2018, 5813, 2273–2284. [Google Scholar] [CrossRef]
- Singh, B.P.; Aluko, R.E.; Hati, S.; Solanki, D. Bioactive peptides in the management of lifestyle-related diseases: Current trends and future perspectives. Crit. Rev. Food Sci. Nutr. 2022, 62, 4593–4606. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, J.; Mu, T.; Zhang, H.; Cao, J.; Li, H.; Zhao, K. Selection of goat β-casein derived ACE-inhibitory peptide SQPK and insights into its effect and regulatory mechanism on the function of endothelial cells. Int. J. Biol. Macromol. 2023, 253, 127312. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C. Bioinformatics approaches, prospects and challenges of food bioactive peptide research. Trends Food Sci Technol. 2014, 36, 137–143. [Google Scholar] [CrossRef]
- Srinivas, S.; Prakash, V. Bioactive peptides from bovine milk α-casein: Isolation, characterization and multifunctional properties. Int. J. Pept. Res. Ther. 2010, 16, 7–15. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM database of bioactive peptides: Current opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [CrossRef]
- Minkiewicz, P.; Darewicz, M.; Iwaniak, A.; Sokołowska, J.; Starowicz, P.; Bucholska, J.; Hrynkiewicz, M. Common amino acid subsequences in a universal proteome—Relevance for food science. Int. J. Mol. Sci. 2015, 16, 20748–20773. [Google Scholar] [CrossRef]
- Dziuba, J.; Minkiewicz, P.; Nałecz, D.; Iwaniak, A. Database of biologically active peptide sequences. Food/Nahrung 1999, 43, 190–195. [Google Scholar] [CrossRef]
- Posit team. RStudio: Integrated Development Environment for R; Posit Software, PBC: Boston, MA, USA, 2024; Available online: http://www.posit.co/ (accessed on 25 April 2025).


| Casein Variant | ΣA * |
|---|---|
| α-S1-casein | |
| A | 2.1447 |
| B | 2.1541 |
| E | 2.1216 |
| F | 1.6351 |
| β-casein | |
| A | 2.4500 |
| C | 2.4429 |
| α-S2-casein | |
| A | 1.9959 |
| B | 1.9735 |
| C | 1.9645 |
| F | 2.0094 |
| κ-casein | |
| A | 2.1914 |
| B | 2.2038 |
| C | 2.2161 |
| D | 2.2286 |
| E | 2.2223 |
| Peptide | Peptide Ranker * | Bioactive Activity | α-S1-Casein | β-Casein | α-S2-Casein | κ-Casein | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | B | E | F | A | C | A | B | C | F | A | B | C | D | E | |||
| QGL | 0.533495 | ACE-I; DPP-IV-I | ✓ | ✓ | ✓ | ✓ | |||||||||||
| QM | 0.607122 | - | ✓ | ✓ | ✓ | ✓ | |||||||||||
| STF | 0.58977 | ACE-I; H; DPP-IV-I; DPP-III-I; R-I; P-L-I | + | ||||||||||||||
| CPL | 0.916551 | ACE-I; DPP-IV-I; X-P-I; L-I | + | ||||||||||||||
| VIW | 0.610494 | ACE-I; DPP-IV-I | + | ||||||||||||||
| SF | 0.948796 | ACE-I; DPP-IV-I; R-I | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||||||||
| GY | 0.741592 | ACE-I; DPP-IV-I; DPP-III-I | ✓ | ✓ | ✓ | ||||||||||||
| SM | 0.628268 | DPP-III-I | ✓ | ✓ | ✓ | ||||||||||||
| QPM | 0.810167 | ACE-I; DPP-IV-I | ✓ | ✓ | ✓ | ||||||||||||
| PQL | 0.527882 | ACE-I; DPP-IV-I | ✓ | ✓ | ✓ | ||||||||||||
| QF | 0.946135 | DPP-IV-I; R-I | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | |||
| PSGAW | 0.870583 | ACE-I; AO; I; DPP-IV-I; A-C-I | ✓ | ✓ | ✓ | ||||||||||||
| PL | 0.811148 | ACE-I; DPP-IV-I; X-P-I; L-I | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| AAM | 0.55858 | ACE-I; A-B; H; DPP-IV-I; I-TPP-II; I-C-A-AP; D-A-D-DP-I | + | ||||||||||||||
| ACL | 0.746733 | - | ✓ | ✓ | |||||||||||||
| PF | 0.99343 | DPP-IV-I; DPP-III-I; ACE2-I | ✓ | ✓ | |||||||||||||
| PVEPF | 0.604782 | ACE-I; AO; DPP-IV-I; A-G-I; DPP-III-I; ACE2-I; N-I | ✓ | ✓ | |||||||||||||
| QPPQPL | 0.823819 | ACE-I; DPP-IV-I; A-G-I; X-P-I; L-I | ✓ | ✓ | |||||||||||||
| DM | 0.607466 | ACE-I | ✓ | ✓ | |||||||||||||
| PIQAF | 0.749093 | ACE-I; DPP-IV-I; I-TPP-II | ✓ | ✓ | |||||||||||||
| GPF | 0.989324 | AAM; ACE-I; AT; RL; AO; H; DPP-IV-I; DPP-III-I; ACE2-I; N-I | ✓ | ✓ | |||||||||||||
| PIL | 0.641797 | ACE-I; S; NP; DPP-IV-I | ✓ | ✓ | |||||||||||||
| IF | 0.949173 | ACE-I | ✓ | ✓ | ✓ | ✓ | |||||||||||
| PR | 0.787626 | ACE-I; DPP-III-I | ✓ | ✓ | ✓ | ✓ | |||||||||||
| PY | 0.736696 | NP; A-I; DPP-IV-I; PL-A2-I | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||
| PW | 0.992911 | AO; DPP-IV-I | ✓ | ✓ | ✓ | ✓ | |||||||||||
| AGAF | 0.873661 | ACE-I; AO; DPP-IV-I; CaMPDE-I; I-TPP-II; A-C-I | ✓ | ✓ | ✓ | ||||||||||||
| AGPF | 0.969222 | AAM; ACE-I; AT; RL; AO; DPP-IV-I; DPP-III-I; ACE2-I; N-I | + | ||||||||||||||
| AW | 0.9669 | ACE-I; AO; DPP-IV-I | ✓ | ✓ | ✓ | ✓ | |||||||||||
| AM | 0.74549 | - | ✓ | ✓ | ✓ | ✓ | |||||||||||
| AIPY | 0.613395 | ACE-I; NP; A-I; DPP-IV-I; PL-A2-I | ✓ | ✓ | ✓ | ✓ | |||||||||||
| GL | 0.808777 | ACE-I; DPP-IV-I | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| QW | 0.928524 | DPP-IV-I | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| PH | 0.541688 | ACE-I; DPP-IV-I; AO | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
| AIPPK | 0.575607 | A-A-I; ACE-I; AT; AO; A-I; DPP-IV-I; A-G-I; ACE2-I | ✓ | ✓ | ✓ | ✓ | ✓ | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio-Reyes, A.Y.; Delgado-Enciso, I.; Casas, E.; Garza-Brenner, E.; Sifuentes-Rincón, A.M. Influence of the Casein Genotype on Goat Milk Bioactivity: An In Silico Analysis of the Casein Peptidome. Molecules 2025, 30, 2601. https://doi.org/10.3390/molecules30122601
Rubio-Reyes AY, Delgado-Enciso I, Casas E, Garza-Brenner E, Sifuentes-Rincón AM. Influence of the Casein Genotype on Goat Milk Bioactivity: An In Silico Analysis of the Casein Peptidome. Molecules. 2025; 30(12):2601. https://doi.org/10.3390/molecules30122601
Chicago/Turabian StyleRubio-Reyes, Aram Y., Iván Delgado-Enciso, Eduardo Casas, Estela Garza-Brenner, and Ana M. Sifuentes-Rincón. 2025. "Influence of the Casein Genotype on Goat Milk Bioactivity: An In Silico Analysis of the Casein Peptidome" Molecules 30, no. 12: 2601. https://doi.org/10.3390/molecules30122601
APA StyleRubio-Reyes, A. Y., Delgado-Enciso, I., Casas, E., Garza-Brenner, E., & Sifuentes-Rincón, A. M. (2025). Influence of the Casein Genotype on Goat Milk Bioactivity: An In Silico Analysis of the Casein Peptidome. Molecules, 30(12), 2601. https://doi.org/10.3390/molecules30122601