

Curcumin-Induced Molecular Mechanisms in U-87 MG Glioblastoma Cells: Insights from Global Gene Expression Profiling

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Curcumin Treatment Decreases U87 MG Cell Viability and Proliferation
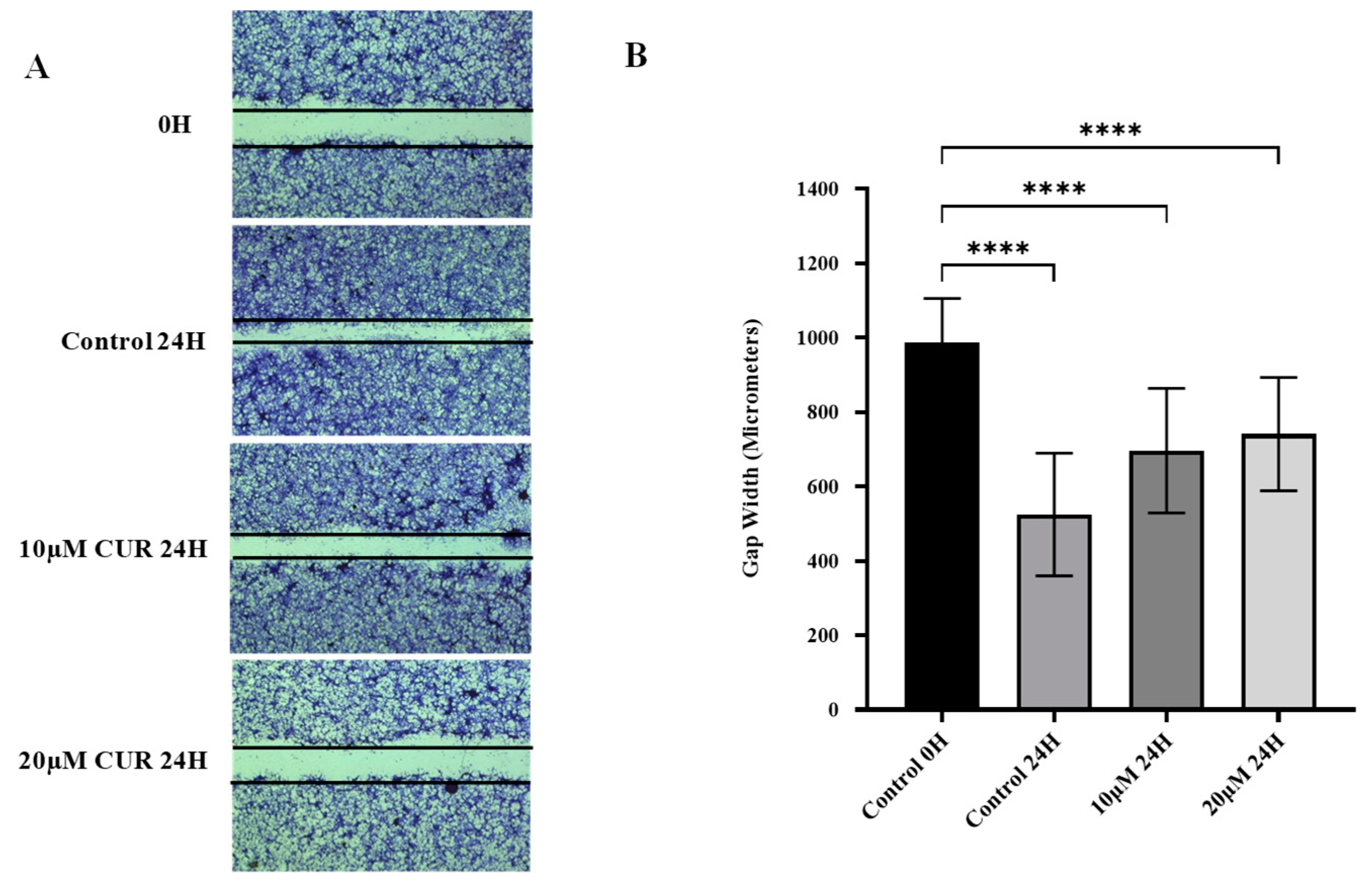
2.2. Curcumin Reduced Migration of U87 MG Cells
2.3. Curcumin Induced Apoptosis but Not Senescence in U87 MG Cells
2.4. Curcumin Induced Differential Gene Expression in U87 MG Cells
3. Discussion
3.1. Curcumin Suppresses Viability and Migration of U87 MG Cells
3.2. Curcumin Triggers Apoptosis in U87 MG Glioblastoma Cells
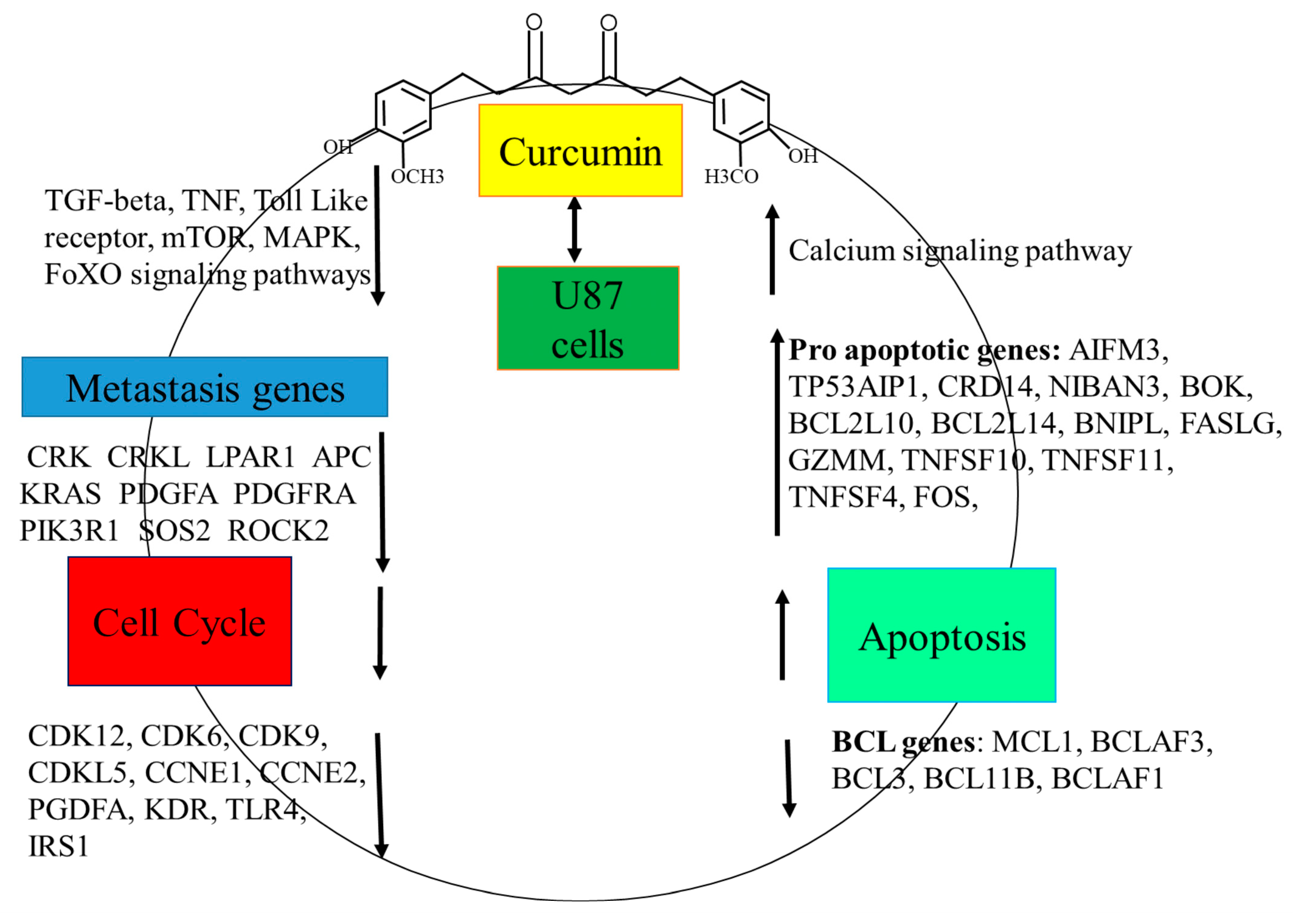
3.3. Curcumin Treatment Altered Gene Expression in U87 MG Cells
4. Materials and Methods
4.1. Curcumin
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Poly-Caspase Assay
4.5. SA-Beta Galactosidase Assay
4.6. RNA Isolation and Library Preparation
4.7. RNA-Seq Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agosti, E.; Zeppieri, M.; De Maria, L.; Tedeschi, C.; Fontanella, M.M.; Panciani, P.P.; Ius, T. Glioblastoma immunotherapy: A systematic review of the present strategies and prospects for advancements. Int. J. Mol. Sci. 2023, 24, 15037. [Google Scholar] [CrossRef]
- Ren, X.; Qin, L. Glioblastoma Management in the Post-COVID-19 Era: Challenges, Strategies, and Adaptations; Intechopen: London, UK, 2025. [Google Scholar]
- Jackson, G.A.; Adamson, D.C. Similarities in Mechanisms of Ovarian Cancer Metastasis and Brain Glioblastoma Multiforme Invasion Suggest Common Therapeutic Targets. Cells 2025, 14, 171. [Google Scholar] [CrossRef]
- Aleksakhina, S.N.; Kashyap, A.; Imyanitov, E.N. Mechanisms of acquired tumor drug resistance. Biochim. Et Biophys. Acta (BBA)-Rev. Cancer 2019, 1872, 188310. [Google Scholar] [CrossRef] [PubMed]
- Srirangan, P.; Sabina, E.P. Protective effects of herbal compounds against cyclophosphamide-induced organ toxicity: A pathway-centered approach. Drug Chem. Toxicol. 2025, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Reddy, C.S.; Natarajan, P.; Nimmakayala, P.; Hankins, G.R.; Reddy, U.K. From Fruit Waste to Medical Insight: The Comprehensive Role of Watermelon Rind Extract on Renal Adenocarcinoma Cellular and Transcriptomic Dynamics. Int. J. Mol. Sci. 2023, 24, 15615. [Google Scholar] [CrossRef]
- Chinreddy, S.R.; Mashozhera, N.T.; Rashrash, B.; Flores-Iga, G.; Nimmakayala, P.; Hankins, G.R.; Harris, R.T.; Reddy, U.K. Unraveling TRPV1’s Role in Cancer: Expression, Modulation, and Therapeutic Opportunities with Capsaicin. Molecules 2024, 29, 4729. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Saha, P.; Dey, S.; Sinha, D. Natural Products as Chemosensitizers for Adjunct Therapy in Cancer Management. Pharmacother. Bot. Cancer Chemoprev. 2020, 154, 67–119. [Google Scholar]
- Islam, M.R.; Rauf, A.; Akash, S.; Trisha, S.I.; Nasim, A.H.; Akter, M.; Dhar, P.S.; Ogaly, H.A.; Hemeg, H.A.; Wilairatana, P. Targeted therapies of curcumin focus on its therapeutic benefits in cancers and human health: Molecular signaling pathway-based approaches and future perspectives. Biomed. Pharmacother. 2024, 170, 116034. [Google Scholar] [CrossRef]
- Banerjee, S.; Singh, S.K.; Chowdhury, I.; Lillard, J.W., Jr.; Singh, R. Combinatorial effect of curcumin with docetaxel modulates apoptotic and cell survival molecules in prostate cancer. Front. Biosci. 2017, 9, 235. [Google Scholar]
- Tiryaki, S.; Macit, M.; Zemheri, I.E.; Süt, P.A.; Duman, G.; Telci, D. Anticancer Activity of Soy Lecithin-Based Curcumin in Prostate Cancer. J. Appl. Polym. Sci. 2025, 142, e56816. [Google Scholar] [CrossRef]
- Du, B.; Jiang, L.; Xia, Q.; Zhong, L. Synergistic inhibitory effects of curcumin and 5-fluorouracil on the growth of the human colon cancer cell line HT-29. Chemotherapy 2005, 52, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Cespedes-Acuña, C.L.; Yang, Z.; Abu Bakar, M.Z.; Chan, K.W.; Deng, X. Plant foods and their bioactives as dietary enhancers for colon cancer treatment with 5-fluorouracil. Food Rev. Int. 2025, 1–50. [Google Scholar] [CrossRef]
- Casarcia, N.; Rogers, P.; Guld, E.; Iyer, S.; Li, Y.; Burcher, J.T.; Deliberto, L.K.; Banerjee, S.; Bishayee, A. Phytochemicals for the prevention and treatment of pancreatic cancer: Current progress and future prospects. Br. J. Pharmacol. 2023, 182, 2181–2234. [Google Scholar] [CrossRef] [PubMed]
- Allegra, A.; Mirabile, G.; Ettari, R.; Pioggia, G.; Gangemi, S. The impact of curcumin on immune response: An immunomodulatory strategy to treat sepsis. Int. J. Mol. Sci. 2022, 23, 14710. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Rane, G.; Kanchi, M.M.; Arfuso, F.; Chinnathambi, A.; Zayed, M.; Alharbi, S.A.; Tan, B.K.; Kumar, A.P.; Sethi, G. The multifaceted role of curcumin in cancer prevention and treatment. Molecules 2015, 20, 2728–2769. [Google Scholar] [CrossRef]
- Dhandapani, K.M.; Mahesh, V.B.; Brann, D.W. Curcumin suppresses growth and chemoresistance of human glioblastoma cells via AP-1 and NFκB transcription factors. J. Neurochem. 2007, 102, 522–538. [Google Scholar] [CrossRef]
- Lin, S.-S.; Lai, K.-C.; Hsu, S.-C.; Yang, J.-S.; Kuo, C.-L.; Lin, J.-P.; Ma, Y.-S.; Wu, C.-C.; Chung, J.-G. Curcumin inhibits the migration and invasion of human A549 lung cancer cells through the inhibition of matrix metalloproteinase-2 and-9 and Vascular Endothelial Growth Factor (VEGF). Cancer Lett. 2009, 285, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.X.; Zhao, L.; Deng, C.; Yang, L.; Wang, Y.; Guo, T.; Li, L.; Lin, J.; Zhang, L. Curcumin suppresses proliferation and induces apoptosis of human hepatocellular carcinoma cells via the wnt signaling pathway. Int. J. Oncol. 2013, 43, 1951–1959. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Feng, T.; Liu, X.; Liu, Q. Curcumin inhibits migration and invasion of non-small cell lung cancer cells through up-regulation of miR-206 and suppression of PI3K/AKT/mTOR signaling pathway. Acta Pharm. 2020, 70, 399–409. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, K.; Liu, J.; Yang, J.; Tian, Y.; Yang, C.; Li, Y.; Shao, M.; Su, W.; Song, N. Curcumin regulates cancer progression: Focus on ncRNAs and molecular signaling pathways. Front. Oncol. 2021, 11, 660712. [Google Scholar] [CrossRef]
- Khazei, K.; Jamali, M.; Sarhadi, S.; Dadashpour, M.; Shokrollahzade, S.; Zarghami, N. Transcriptome profiling of curcumin-treated T47D human breast cancer cells by a system-based approach. Gene Rep. 2022, 27, 101556. [Google Scholar] [CrossRef]
- Wang, R.; Yu, H.; Chen, P.; Yuan, T.; Zhang, J. Integrated transcriptome and molecular docking to identify the hub superimposed attenuation targets of curcumin in breast cancer cells. Int. J. Mol. Sci. 2023, 24, 12479. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liang, C.; Yang, H.; Li, X.; Deng, X.; Liang, X.; Li, L.; Huang, Z.; Lu, D.; Ma, Y. Curcumin induces apoptosis and inhibits the growth of adrenocortical carcinoma: Identification of potential candidate genes and pathways by transcriptome analysis. Oncol. Lett. 2021, 21, 476. [Google Scholar] [CrossRef]
- Li, R.; Zhang, J.; Zhou, Y.; Gao, Q.; Wang, R.; Fu, Y.; Zheng, L.; Yu, H. Transcriptome investigation and in vitro verification of curcumin-induced HO-1 as a feature of ferroptosis in breast cancer cells. Oxidative Med. Cell. Longev. 2020, 2020, 3469840. [Google Scholar] [CrossRef] [PubMed]
- Mo, F.; Xiao, Y.; Zeng, H.; Fan, D.; Song, J.; Liu, X.; Luo, M.; Ma, X. Curcumin-induced global profiling of transcriptomes in small cell lung cancer cells. Front. Cell Dev. Biol. 2021, 8, 588299. [Google Scholar] [CrossRef]
- Salam, R.; Saliou, A.; Bielle, F.; Bertrand, M.; Antoniewski, C.; Carpentier, C.; Alentorn, A.; Capelle, L.; Sanson, M.; Huillard, E. Cellular senescence in malignant cells promotes tumor progression in mouse and patient Glioblastoma. Nat. Commun. 2023, 14, 441. [Google Scholar] [CrossRef]
- Giordano, A.; Tommonaro, G. Curcumin and cancer. Nutrients 2019, 11, 2376. [Google Scholar] [CrossRef]
- Wong, S.C.; Kamarudin, M.N.A.; Naidu, R. Anticancer mechanism of curcumin on human glioblastoma. Nutrients 2021, 13, 950. [Google Scholar] [CrossRef]
- Zoi, V.; Kyritsis, A.P.; Galani, V.; Lazari, D.; Sioka, C.; Voulgaris, S.; Alexiou, G.A. The role of curcumin in cancer: A focus on the PI3K/Akt pathway. Cancers 2024, 16, 1554. [Google Scholar] [CrossRef]
- Hesari, A.; Rezaei, M.; Rezaei, M.; Dashtiahangar, M.; Fathi, M.; Rad, J.G.; Momeni, F.; Avan, A.; Ghasemi, F. Effect of curcumin on glioblastoma cells. J. Cell. Physiol. 2019, 234, 10281–10288. [Google Scholar] [CrossRef]
- Yang, C.-L.; Liu, Y.-Y.; Ma, Y.-G.; Xue, Y.-X.; Liu, D.-G.; Ren, Y.; Liu, X.-B.; Li, Y.; Li, Z. Curcumin blocks small cell lung cancer cells migration, invasion, angiogenesis, cell cycle and neoplasia through Janus kinase-STAT3 signalling pathway. PLoS ONE 2012, 7, e37960. [Google Scholar] [CrossRef]
- Chen, Q.-Y.; Zheng, Y.; Jiao, D.-M.; Chen, F.-Y.; Hu, H.-Z.; Wu, Y.-Q.; Song, J.; Yan, J.; Wu, L.-J.; Lv, G.-Y. Curcumin inhibits lung cancer cell migration and invasion through Rac1-dependent signaling pathway. J. Nutr. Biochem. 2014, 25, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sun, L.; Lei, J.; Wu, Z.; Ma, Q.; Wang, Z. Curcumin inhibits pancreatic cancer cell invasion and EMT by interfering with tumor-stromal crosstalk under hypoxic conditions via the IL-6/ERK/NF-κB axis. Oncol. Rep. 2020, 44, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Weissenberger, J.; Priester, M.; Bernreuther, C.; Rakel, S.; Glatzel, M.; Seifert, V.; Kögel, D. Dietary curcumin attenuates glioma growth in a syngeneic mouse model by inhibition of the JAK1, 2/STAT3 signaling pathway. Clin. Cancer Res. 2010, 16, 5781–5795. [Google Scholar] [CrossRef]
- Li, J.; Ye, L.; Parr, C.; Douglas-Jones, A.; Kynaston, H.G.; Mansel, R.E.; Jiang, W.G. The aberrant expression of bone morphogenetic protein 12 (BMP-12) in human breast cancer and its potential prognostic value. Gene Ther. Mol. Biol. 2009, 13, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Abdullah Thani, N.A.; Sallis, B.; Nuttall, R.; Schubert, F.R.; Ahsan, M.; Davies, D.; Purewal, S.; Cooper, A.; Rooprai, H.K. Induction of apoptosis and reduction of MMP gene expression in the U373 cell line by polyphenolics in Aronia melanocarpa and by curcumin. Oncol. Rep. 2012, 28, 1435–1442. [Google Scholar] [CrossRef]
- Karmakar, S.; Banik, N.L.; Ray, S.K. Curcumin suppressed anti-apoptotic signals and activated cysteine proteases for apoptosis in human malignant glioblastoma U87MG cells. Neurochem. Res. 2007, 32, 2103–2113. [Google Scholar] [CrossRef]
- Cheng, C.; Jiao, J.T.; Qian, Y.; Guo, X.Y.; Huang, J.; Dai, M.C.; Zhang, L.; Ding, X.P.; Zong, D.; Shao, J.F. Curcumin induces G2/M arrest and triggers apoptosis via FoxO1 signaling in U87 human glioma cells. Mol. Med. Rep. 2016, 13, 3763–3770. [Google Scholar] [CrossRef]
- Wang, P.; Hao, X.; Li, X.; Yan, Y.; Tian, W.; Xiao, L.; Wang, Z.; Dong, J. Curcumin inhibits adverse psychological stress-induced proliferation and invasion of glioma cells via down-regulating the ERK/MAPK pathway. J. Cell. Mol. Med. 2021, 25, 7190–7203. [Google Scholar] [CrossRef]
- Wang, L.; Hu, R.; Dai, A. Curcumin Increased the Sensitivity of Non-Small-Cell Lung Cancer to Cisplatin through the Endoplasmic Reticulum Stress Pathway. Evid.-Based Complement. Altern. Med. 2022, 2022, 6886366. [Google Scholar] [CrossRef]
- Huang, Y.-F.; Zhu, D.-J.; Chen, X.-W.; Chen, Q.-K.; Luo, Z.-T.; Liu, C.-C.; Wang, G.-X.; Zhang, W.-J.; Liao, N.-Z. Curcumin enhances the effects of irinotecan on colorectal cancer cells through the generation of reactive oxygen species and activation of the endoplasmic reticulum stress pathway. Oncotarget 2017, 8, 40264. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Chen, S.; Lu, H.; Li, H.; Qin, C. Study on the inhibitory effect of curcumin on GBM and its potential mechanism. Drug Des. Dev. Ther. 2021, 15, 2769–2781. [Google Scholar] [CrossRef]
- Huang, T.-Y.; Tsai, T.-H.; Hsu, C.-W.; Hsu, Y.-C. Curcuminoids suppress the growth and induce apoptosis through caspase-3-dependent pathways in glioblastoma multiforme (GBM) 8401 cells. J. Agric. Food Chem. 2010, 58, 10639–10645. [Google Scholar] [CrossRef]
- Mosieniak, G.; Adamowicz, M.; Alster, O.; Jaskowiak, H.; Szczepankiewicz, A.A.; Wilczynski, G.M.; Ciechomska, I.A.; Sikora, E. Curcumin induces permanent growth arrest of human colon cancer cells: Link between senescence and autophagy. Mech. Ageing Dev. 2012, 133, 444–455. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, R.; Yu, Y.; Liu, J.; Luo, T.; Fan, F. Glioblastoma treatment modalities besides surgery. J. Cancer 2019, 10, 4793. [Google Scholar] [CrossRef]
- Cheng, J.; Zeng, M.; Peng, B.; Li, P.; Zhao, S. Transient receptor potential vanilloid-1 (TRPV1) channels act as suppressors of the growth of glioma. Brain Res. Bull. 2024, 211, 110950. [Google Scholar] [CrossRef]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative approaches for cancer treatment: Current perspectives and new challenges. ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.B.; Sundaram, C.; Malani, N.; Ichikawa, H. Curcumin: The Indian solid gold. Mol. Targets Ther. Uses Curcumin Health Dis. 2007, 595, 1–75. [Google Scholar]
- Avci, C.; Dodurga, Y.; Susluer, S.; Sıgva, Z.; Yucebas, M.; Caglar, H.; Akalin, T.; Dalbasti, T.; Oktar, N.; Gunduz, C. Promoter hypermethylation-mediated down-regulation of RUNX3 gene in human brain tumors. Ir. J. Med. Sci. 2014, 183, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.; Ito, Y. RUNX3 is multifunctional in carcinogenesis of multiple solid tumors. Oncogene 2010, 29, 2605–2615. [Google Scholar] [CrossRef]
- Li, X.; Zheng, J.; Diao, H.; Liu, Y. RUNX 3 is down-regulated in glioma by Myc-regulated miR-4295. J. Cell. Mol. Med. 2016, 20, 518–525. [Google Scholar] [CrossRef]
- Steponaitis, G.; Kazlauskas, A.; Vaitkienė, P.; Deltuva, V.P.; Mikuciunas, M.; Skiriutė, D. Oncosuppressive role of RUNX3 in human astrocytomas. J. Oncol. 2019, 2019, 1232434. [Google Scholar] [CrossRef]
- Mei, P.-J.; Bai, J.; Liu, H.; Li, C.; Wu, Y.-P.; Yu, Z.-Q.; Zheng, J.-N. RUNX3 expression is lost in glioma and its restoration causes drastic suppression of tumor invasion and migration. J. Cancer Res. Clin. Oncol. 2011, 137, 1823–1830. [Google Scholar] [CrossRef]
- Sun, J.; Jia, Z.; Li, B.; Zhang, A.; Wang, G.; Pu, P.; Chen, Z.; Wang, Z.; Yang, W. MiR-19 regulates the proliferation and invasion of glioma by RUNX3 via β-catenin/Tcf-4 signaling. Oncotarget 2017, 8, 110785. [Google Scholar] [CrossRef]
- Sun, J.; Li, B.; Jia, Z.; Zhang, A.; Wang, G.; Chen, Z.; Shang, Z.; Zhang, C.; Cui, J.; Yang, W. RUNX3 inhibits glioma survival and invasion via suppression of the β-catenin/TCF-4 signaling pathway. J. Neuro-Oncol. 2018, 140, 15–26. [Google Scholar] [CrossRef]
- Newton, A.C.; Brognard, J. Reversing the paradigm: Protein kinase C as a tumor suppressor. Trends Pharmacol. Sci. 2017, 38, 438–447. [Google Scholar] [CrossRef]
- Pilo, C.A.; Newton, A.C. Two sides of the same coin: Protein kinase C γ in cancer and neurodegeneration. Front. Cell Dev. Biol. 2022, 10, 929510. [Google Scholar] [CrossRef]
- Satow, R.; Suzuki, Y.; Asada, S.; Ota, S.; Idogawa, M.; Kubota, S.; Ikeo, N.; Yoneda, A.; Fukami, K. Downregulation of protein kinase C gamma reduces epithelial property and enhances malignant phenotypes in colorectal cancer cells. Iscience 2022, 25, 105501. [Google Scholar] [CrossRef]
- Liu, L.; Wang, G.; Wang, L.; Yu, C.; Li, M.; Song, S.; Hao, L.; Ma, L.; Zhang, Z. Computational identification and characterization of glioma candidate biomarkers through multi-omics integrative profiling. Biol. Direct 2020, 15, 10. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Wang, X.; Wang, X.-J.; Zheng, B.-Z.; Wang, Y.; Liang, B. Curcumin inhibits the growth via Wnt/β-catenin pathway in non-small-cell lung cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7492–7499. [Google Scholar]
- Zhang, X.; Dong, S.; Xu, F. Structural and druggability landscape of frizzled G protein-coupled receptors. Trends Biochem. Sci. 2018, 43, 1033–1046. [Google Scholar] [CrossRef]
- Huang, K.; Xu, H.; Han, L.; Xu, R.; Xu, Z.; Xie, Y. Identification of therapeutic targets and prognostic biomarkers among frizzled family genes in glioma. Front. Mol. Biosci. 2023, 9, 1054614. [Google Scholar] [CrossRef] [PubMed]
- Hirano, H.; Yonezawa, H.; Yunoue, S.; Habu, M.; Uchida, H.; Yoshioka, T.; Kishida, S.; Kishida, M.; Oyoshi, T.; Fujio, S. Immunoreactivity of Wnt5a, Fzd2, Fzd6, and Ryk in glioblastoma: Evaluative methodology for DAB chromogenic immunostaining. Brain Tumor Pathol. 2014, 31, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Ran, Y.; Han, S.; Gao, D.; Chen, X.; Liu, C. Interference of FZD2 suppresses proliferation, vasculogenic mimicry and stemness in glioma cells via blocking the Notch/NF-κB signaling pathway. Exp. Ther. Med. 2024, 28, 373. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Zhang, L.; Jiang, Z.; Tan, G.; Wang, Z. FZD1/KLF10-hsa-miR-4762–5p/miR-224–3p-circular RNAs axis as prognostic biomarkers and therapeutic targets for glioblastoma: A comprehensive report. BMC Med. Genom. 2023, 16, 21. [Google Scholar] [CrossRef]
- Ma, S.; Guo, Z.; Wang, B.; Yang, M.; Yuan, X.; Ji, B.; Wu, Y.; Chen, S. A Computational Framework to Identify Biomarkers for Glioma Recurrence and Potential Drugs Targeting Them. Front. Genet. 2022, 12, 832627. [Google Scholar] [CrossRef]
- Ohka, F.; Shinjo, K.; Deguchi, S.; Matsui, Y.; Okuno, Y.; Katsushima, K.; Suzuki, M.; Kato, A.; Ogiso, N.; Yamamichi, A. Pathogenic epigenetic consequences of genetic alterations in IDH-wild-type diffuse astrocytic gliomas. Cancer Res. 2019, 79, 4814–4827. [Google Scholar] [CrossRef]
- Li, Y.; Liu, Z.; Zhang, Y. Expression and prognostic impact of FZDs in pancreatic adenocarcinoma. BMC Gastroenterol. 2021, 21, 1–14. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, W.; Zhao, C. Frizzled receptors in tumors, focusing on signaling, roles, modulation mechanisms, and targeted therapies. Oncol. Res. 2021, 28, 661. [Google Scholar] [CrossRef]
- Yin, S.; Xu, L.; Bonfil, R.D.; Banerjee, S.; Sarkar, F.H.; Sethi, S.; Reddy, K.B. Tumor-initiating cells and FZD8 play a major role in drug resistance in triple-negative breast cancer. Mol. Cancer Ther. 2013, 12, 491–498. [Google Scholar] [CrossRef]
- Caja, L.; Bellomo, C.; Moustakas, A. Transforming growth factor β and bone morphogenetic protein actions in brain tumors. FEBS Lett. 2015, 589, 1588–1597. [Google Scholar] [CrossRef] [PubMed]
- Tenen, D.G. Disruption of differentiation in human cancer: AML shows the way. Nat. Rev. Cancer 2003, 3, 89–101. [Google Scholar] [CrossRef]
- Hanavadi, S.; Martin, T.; Watkins, G.; Mansel, R.; Jiang, W. The role of growth differentiation factor-9 (GDF-9) and its analog, GDF-9b/BMP-15, in human breast cancer. Ann. Surg. Oncol. 2007, 14, 2159–2166. [Google Scholar] [CrossRef]
- Varadaraj, A.; Patel, P.; Serrao, A.; Bandyopadhay, T.; Lee, N.Y.; Jazaeri, A.A.; Huang, Z.; Murphy, S.K.; Mythreye, K. Epigenetic regulation of GDF2 suppresses anoikis in ovarian and breast epithelia. Neoplasia 2015, 17, 826–838. [Google Scholar] [CrossRef]
- Du, P.; Ye, L.; Li, H.; Ruge, F.; Yang, Y.; Jiang, W.G. Growth differentiation factor-9 expression is inversely correlated with an aggressive behaviour in human bladder cancer cells. Int. J. Mol. Med. 2012, 29, 428–434. [Google Scholar] [PubMed]
- Codó, P.; Weller, M.; Kaulich, K.; Schraivogel, D.; Silginer, M.; Reifenberger, G.; Meister, G.; Roth, P. Control of glioma cell migration and invasiveness by GDF-15. Oncotarget 2016, 7, 7732. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Li, Z.; Fu, J.; Zhou, R. Growth and differentiation factor 15 regulates PD-L1 expression in glioblastoma. Cancer Manag. Res. 2019, 11, 2653–2661. [Google Scholar] [CrossRef]
- Kadowaki, M.; Yoshioka, H.; Kamitani, H.; Watanabe, T.; Wade, P.A.; Eling, T.E. DNA methylation-mediated silencing of nonsteroidal anti-inflammatory drug-activated gene (NAG-1/GDF15) in glioma cell lines. Int. J. Cancer 2012, 130, 267–277. [Google Scholar] [CrossRef]
- Bolger, A.; Giorgi, F. Trimmomatic: A flexible read trimming tool for Illumina NGS data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Love, M.; Anders, S.; Huber, W. Differential analysis of count data–the DESeq2 package. Genome Biol 2014, 15, 10–1186. [Google Scholar]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39 (Suppl. S2), W316–W322. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mashozhera, N.T.; Reddy, C.S.; Ranasinghe, Y.N.; Natarajan, P.; Reddy, U.K.; Hankins, G. Curcumin-Induced Molecular Mechanisms in U-87 MG Glioblastoma Cells: Insights from Global Gene Expression Profiling. Molecules 2025, 30, 2108. https://doi.org/10.3390/molecules30102108
Mashozhera NT, Reddy CS, Ranasinghe YN, Natarajan P, Reddy UK, Hankins G. Curcumin-Induced Molecular Mechanisms in U-87 MG Glioblastoma Cells: Insights from Global Gene Expression Profiling. Molecules. 2025; 30(10):2108. https://doi.org/10.3390/molecules30102108
Chicago/Turabian StyleMashozhera, Nicole Tendayi, Chinreddy Subramanyam Reddy, Yevin Nenuka Ranasinghe, Purushothaman Natarajan, Umesh K. Reddy, and Gerald Hankins. 2025. "Curcumin-Induced Molecular Mechanisms in U-87 MG Glioblastoma Cells: Insights from Global Gene Expression Profiling" Molecules 30, no. 10: 2108. https://doi.org/10.3390/molecules30102108
APA StyleMashozhera, N. T., Reddy, C. S., Ranasinghe, Y. N., Natarajan, P., Reddy, U. K., & Hankins, G. (2025). Curcumin-Induced Molecular Mechanisms in U-87 MG Glioblastoma Cells: Insights from Global Gene Expression Profiling. Molecules, 30(10), 2108. https://doi.org/10.3390/molecules30102108