
Melatonin: A Neurotrophic Factor?

 , , and
, , and
Abstract
1. Introduction
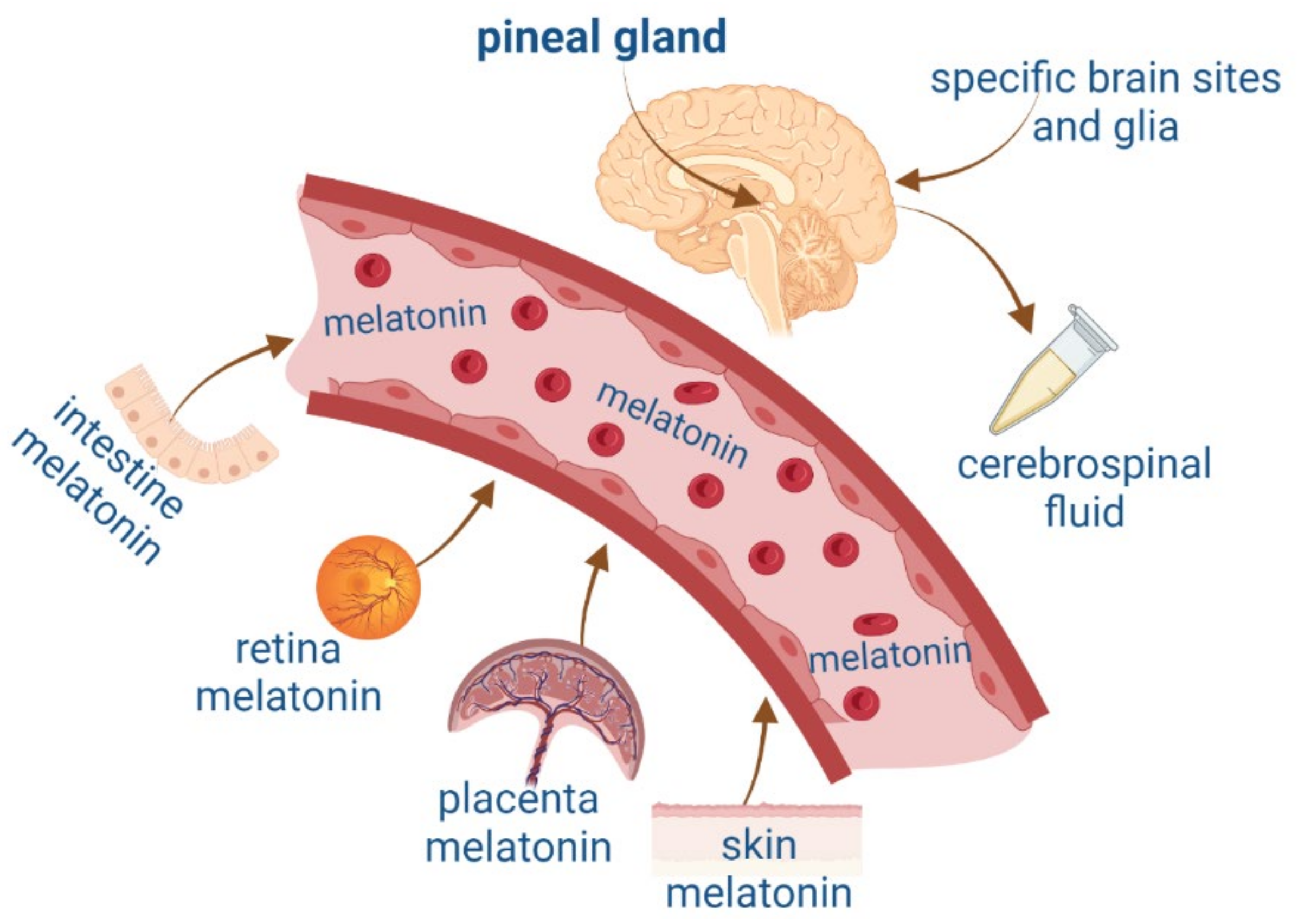
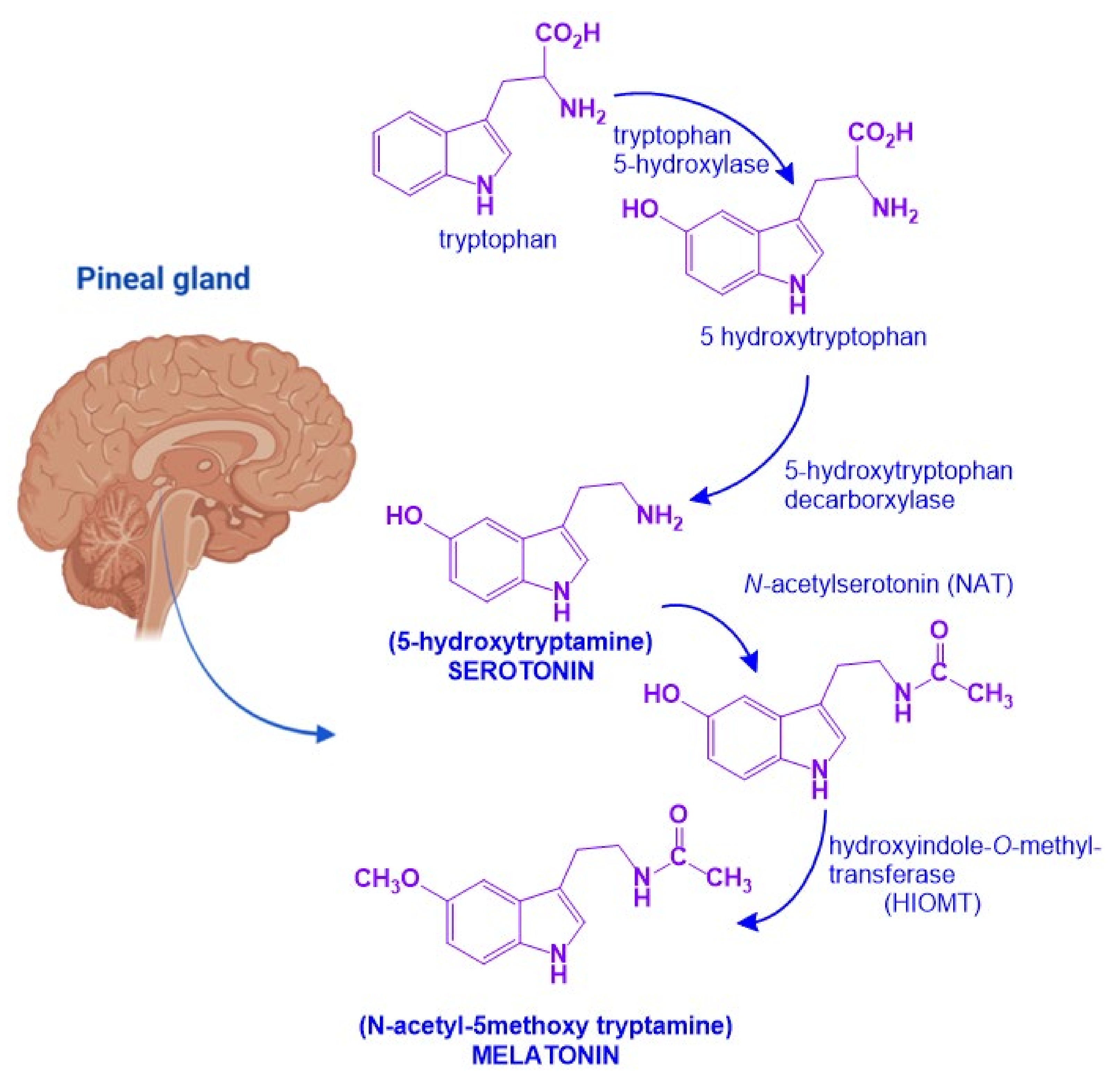
2. Melatonin and Neurotrophic Factors Synthesis
3. The Neurodevelopment in the Adult Brain
4. Evidence That Supports Melatonin Acts as a Neurotrophic Factor
Mechanism of Action Involved in Neurogenesis and Neural Differentiation: Neurotrophic Factors and Melatonin
5. Therapeutic Implications
6. Final Considerations: Is Melatonin a Neurotrophic Factor?
7. Method
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of Melatonin, Pineal Factor That Lightens Melanocytes. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Zhou, Z.; Cruz, M.H.C.; Fuentes-Broto, L.; Galano, A. Phytomelatonin: Assisting Plants to Survive and Thrive. Molecules 2015, 20, 7396–7437. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Atigadda, V.R.; Brzeminski, P.; Fabisiak, A.; Tang, E.K.Y.; Tuckey, R.C.; Reiter, R.J.; Slominski, A.T. Detection of Serotonin, Melatonin, and Their Metabolites in Honey. ACS Food Sci. Technol. 2021, 1, 1228–1235. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Fabisiak, A.; Brzeminski, P.; Reiter, R.J.; Slominski, A.T. Serotonin, Melatonin and Their Precursors and Metabolites and Vitamin D3 Derivatives in Honey. Melatonin Res. 2022, 5, 374–380. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef]
- Tricoire, H.; Møller, M.; Chemineau, P.; Malpaux, B. Origin of Cerebrospinal Fluid Melatonin and Possible Function in the Integration of Photoperiod. Reprod. Suppl. 2003, 61, 311–321. [Google Scholar] [CrossRef]
- Hardeland, R.; Pandi-Perumal, S.R.; Cardinali, D.P. Melatonin. Int. J. Biochem. Cell Biol. 2006, 38, 313–316. [Google Scholar] [CrossRef]
- Reiter, R.J. Melatonin: The Chemical Expression of Darkness. Mol. Cell. Endocrinol. 1991, 79, C153–C158. [Google Scholar] [CrossRef]
- Prakash, Y.; Thompson, M.A.; Meuchel, L.; Pabelick, C.M.; Mantilla, C.B.; Zaidi, S.; Martin, R.J. Neurotrophins in Lung Health and Disease. Expert Rev. Respir. Med. 2010, 4, 395–411. [Google Scholar] [CrossRef]
- Thoenen, H. Neurotrophins and Neuronal Plasticity. Science 1995, 270, 593–598. [Google Scholar] [CrossRef]
- Duman, R.S.; Nakagawa, S.; Malberg, J. Regulation of Adult Neurogenesis by Antidepressant Treatment. Neuropsychopharmacology 2001, 25, 836–844. [Google Scholar] [CrossRef]
- Landreth, G.E. Growth Factors. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects; Siegel, G.J., Albers, R.W., Brady, S.T., Price, D.L., Eds.; Elsevier: Oxford, UK, 2006; pp. 471–484. ISBN 0-12-088397-X. [Google Scholar]
- Duman, R.S.; Monteggia, L.M. A Neurotrophic Model for Stress-Related Mood Disorders. Biol. Psychiatry 2006, 59, 1116–1127. [Google Scholar] [CrossRef] [PubMed]
- Sariola, H. The Neurotrophic Factors in Non-Neuronal Tissues. Cell. Mol. Life Sci. 2001, 58, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.M.; Wu, H.C.; Sun, Z.G.; Lian, F.; Leung, P.C.K. Neurotrophins and Glial Cell Linederived Neurotrophic Factor in the Ovary: Physiological and Pathophysiological Implications. Hum. Reprod. Update 2019, 25, 224–242. [Google Scholar] [CrossRef]
- Skaper, S.D. Nerve Growth Factor: A Neuroimmune Crosstalk Mediator for All Seasons. Immunology 2017, 151, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Castroviejo, D.; Escames, G.; Venegas, C.; Díaz-Casado, M.E.; Lima-Cabello, E.; López, L.C.; Rosales-Corral, S.; Tan, D.-X.X.; Reiter, R.J. Extrapineal Melatonin: Sources, Regulation, and Potential Functions. Cell. Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Kim, T.K.; Janjetovic, Z.; Slominski, R.M.; Zmijewski, J.W. Melatonin, Mitochondria, and the Skin. Cell. Mol. Life Sci. 2017, 74, 3913–3925. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Hardeland, R.; Lopez-Burillo, S.; Mayo, J.C.; Sainz, R.M.; Reiter, R.J. Melatonin: A Hormone, a Tissue Factor, an Autocoid, a Paracoid, and an Antioxidant Vitamin. J. Pineal Res. 2003, 34, 75–78. [Google Scholar] [CrossRef]
- Mannino, G.; Pernici, C.; Serio, G.; Gentile, C.; Bertea, C.M. Melatonin and Phytomelatonin: Chemistry, Biosynthesis, Metabolism, Distribution and Bioactivity in Plants and Animals-An Overview. Int. J. Mol. Sci. 2021, 22, 9996. [Google Scholar] [CrossRef]
- Schomerus, C.; Korf, H.W. Mechanisms Regulating Melatonin Synthesis in the Mammalian Pineal Organ. Ann. N. Y. Acad. Sci. 2005, 1057, 372–383. [Google Scholar] [CrossRef]
- Leone, A.M.; Francis, P.L.; McKenzie-Gray, B. Rapid and Simple Synthesis for the Sulphate Esters of 6-Hydroxy-Melatonin and N-Acetyl-Serotonin. J. Pineal Res. 1988, 5, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Skene, D.J.; Papagiannidou, E.; Hashemi, E.; Snelling, J.; Lewis, D.F.V.; Fernandez, M.; Ioannides, C. Contribution of CYP1A2 in the Hepatic Metabolism of Melatonin: Studies with Isolated Microsomal Preparations and Liver Slices. J. Pineal Res. 2001, 31, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Melatonin Metabolism in the Central Nervous System. Curr. Neuropharmacol. 2010, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Francis, P.L.; Leone, A.M.; Young, I.M.; Stovell, P.; Silman, R.E. Gas Chromatographic-Mass Spectrometric Assay for 6-Hydroxymelatonin Sulfate and 6-Hydroxymelatonin Glucuronide in Urine. Clin. Chem. 1987, 33, 453–457. [Google Scholar] [CrossRef]
- Keefe, K.; Sheikh, I.; Smith, G. Targeting Neurotrophins to Specific Populations of Neurons: NGF, BDNF, and NT-3 and Their Relevance for Treatment of Spinal Cord Injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef]
- Skaper, S.D. Neurotrophic Factors: An Overview. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2018; Volume 1727, pp. 1–17. [Google Scholar]
- Jackman, S.L.; Regehr, W.G. The Mechanisms and Functions of Synaptic Facilitation. Neuron 2017, 94, 447–464. [Google Scholar] [CrossRef]
- Song, M.; Martinowich, K.; Lee, F.S. BDNF at the Synapse: Why Location Matters. Mol. Psychiatry 2017, 22, 1370–1375. [Google Scholar] [CrossRef]
- Huang, E.J.; Reichardt, L.F. Neurotrophins: Roles in Neuronal Development and Function. Annu. Rev. Neurosci. 2001, 24, 677–736. [Google Scholar] [CrossRef]
- Levi-Montalcini, R. The Nerve Growth Factor 35 Years Later. Science 1987, 237, 1154–1162. [Google Scholar] [CrossRef]
- Alvarez-Buylla, A.; García-Verdugo, J.M.; Tramontin, A.D. A Unified Hypothesis on the Lineage of Neural Stem Cells. Nat. Rev. Neurosci. 2001, 2, 287–293. [Google Scholar] [CrossRef]
- Nualart, F. Unconventional Neurogenic Niches and Neurogenesis Modulation by Vitamins. J. Stem Cell Res. Ther. 2014, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Bray, D.; Lewis, J.; Raff, M.; Roberts, K.; Watson, D.J. Biología Molecular de La Célula, 3rd ed.; Durfort, M., Llobera, M., Eds.; Ediciones Omega: Barcelona, Spain, 2002; ISBN 84-282-1011-X. [Google Scholar]
- Da Silva, J.S.; Dotti, C.G. Breaking the Neuronal Sphere: Regulation of the Actin Cytoskeleton in Neuritogenesis. Nat. Rev. Neurosci. 2002, 3, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Shu, T.; Wu, T.; Pang, M.; Liu, C.; Wang, X.; Wang, J.; Liu, B.; Rong, L. Effects and Mechanisms of Melatonin on Neural Differentiation of Induced Pluripotent Stem Cells. Biochem. Biophys. Res. Commun. 2016, 474, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Galván-Arrieta, T.; Trueta, C.; Cercós, M.G.; Valdés-Tovar, M.; Alarcón, S.; Oikawa, J.; Zamudio-Meza, H.; Benítez-King, G. The Role of Melatonin in the Neurodevelopmental Etiology of Schizophrenia: A Study in Human Olfactory Neuronal Precursors. J. Pineal Res. 2017, 63, e12421. [Google Scholar] [CrossRef]
- Valdés-Tovar, M.; Estrada-Reyes, R.; Solís-Chagoyán, H.; Argueta, J.; Dorantes-Barrón, A.M.; Quero-Chávez, D.; Cruz-Garduño, R.; Cercós, M.G.; Trueta, C.; Oikawa-Sala, J.; et al. Circadian Modulation of Neuroplasticity by Melatonin: A Target in the Treatment of Depression. Br. J. Pharmacol. 2018, 175, 3200–3208. [Google Scholar] [CrossRef] [PubMed]
- Moriya, T.; Horie, N.; Mitome, M.; Shinohara, K. Melatonin Influences the Proliferative and Differentiative Activity of Neural Stem Cells. J. Pineal Res. 2007, 42, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rodríguez, G.; Klempin, F.; Babu, H.; Benítez-King, G.; Kempermann, G. Melatonin Modulates Cell Survival of New Neurons in the Hippocampus of Adult Mice. Neuropsychopharmacology 2009, 34, 2180–2191. [Google Scholar] [CrossRef]
- Sotthibundhu, A.; Phansuwan-Pujito, P.; Govitrapong, P. Melatonin Increases Proliferation of Cultured Neural Stem Cells Obtained from Adult Mouse Subventricular Zone. J. Pineal Res. 2010, 49, 291–300. [Google Scholar] [CrossRef]
- Liu, J.; Somera-Molina, K.C.; Hudson, R.L.; Dubocovich, M.L. Melatonin Potentiates Running Wheel-Induced Neurogenesis in the Dentate Gyrus of Adult C3H/HeN Mice Hippocampus. J. Pineal Res. 2013, 54, 222–231. [Google Scholar] [CrossRef]
- Estrada-Reyes, R.; Quero-Chávez, D.B.; Trueta, C.; Miranda, A.; Valdés-Tovar, M.; Alarcón-Elizalde, S.; Oikawa-Sala, J.; Argueta, J.; Constantino-Jonapa, L.A.; Muñoz-Estrada, J.; et al. Low Doses of Ketamine and Melatonin in Combination Produce Additive Antidepressant-like Effects in Mice. Int. J. Mol. Sci. 2021, 22, 9225. [Google Scholar] [CrossRef]
- Estrada-Reyes, R.; Quero-Chávez, D.B.; Alarcón-Elizalde, S.; Cercós, M.G.; Trueta, C.; Constantino-Jonapa, L.A.; Oikawa-Sala, J.; Argueta, J.; Cruz-Garduño, R.; Dubocovich, M.L.; et al. Antidepressant Low Doses of Ketamine and Melatonin in Combination Produce Additive Neurogenesis in Human Olfactory Neuronal Precursors. Molecules 2022, 27, 5650. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Alonso, A.; Ramírez-Rodríguez, G.; Benítez-King, G. Melatonin Increases Dendritogenesis in the Hilus of Hippocampal Organotypic Cultures. J. Pineal Res. 2012, 52, 427–436. [Google Scholar] [CrossRef]
- Benítez-King, G. Melatonin as a Cytoskeletal Modulator: Implications for Cell Physiology and Disease. J. Pineal Res. 2006, 40, 1–9. [Google Scholar] [CrossRef]
- Bellon, A.; Ortíz-López, L.; Ramírez-Rodríguez, G.; Antón-Tay, F.; Benítez-King, G. Melatonin Induces Neuritogenesis at Early Stages in N1E-115 Cells through Actin Rearrangements via Activation of Protein Kinase C and Rho-Associated Kinase. J. Pineal Res. 2007, 42, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Rodriguez, G.; Ortíz-López, L.; Domínguez-Alonso, A.; Benítez-King, G.A.; Kempermann, G. Chronic Treatment with Melatonin Stimulates Dendrite Maturation and Complexity in Adult Hippocampal Neurogenesis of Mice. J. Pineal Res. 2011, 50, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Alonso, A.; Valdés-Tovar, M.; Solís-Chagoyán, H.; Benítez-King, G. Melatonin Stimulates Dendrite Formation and Complexity in the Hilar Zone of the Rat Hippocampus: Participation of the Ca++/Calmodulin Complex. Int. J. Mol. Sci. 2015, 16, 1907–1927. [Google Scholar] [CrossRef]
- Ramírez-Rodríguez, G.B.; Palacios-Cabriales, D.M.; Ortiz-López, L.; Estrada-Camarena, E.M.; Vega-Rivera, N.M. Melatonin Modulates Dendrite Maturation and Complexity in the Dorsal-and Ventral-Dentate Gyrus Concomitantly with Its Antidepressant-like Effect in Male Balb/c Mice. Int. J. Mol. Sci. 2020, 21, 1724. [Google Scholar] [CrossRef]
- González-Burgos, I.; Letechipía-Vallejo, G.; López-Loeza, E.; Moralí, G.; Cervantes, M. Long-Term Study of Dendritic Spines from Hippocampal CA1 Pyramidal Cells, after Neuroprotective Melatonin Treatment Following Global Cerebral Ischemia in Rats. Neurosci. Lett. 2007, 423, 162–166. [Google Scholar] [CrossRef]
- Solís-Chagoyán, H.; Domínguez-Alonso, A.; Valdés-Tovar, M.; Argueta, J.; Sánchez-Florentino, Z.A.; Calixto, E.; Benítez-King, G. Melatonin Rescues the Dendrite Collapse Induced by the Pro-Oxidant Toxin Okadaic Acid in Organotypic Cultures of Rat Hilar Hippocampus. Molecules 2020, 25, 5508. [Google Scholar] [CrossRef]
- Ramírez-Rodríguez, G.B.; Olvera-Hernández, S.; Vega-Rivera, N.M.; Ortiz-López, L. Melatonin Influences Structural Plasticity in the Axons of Granule Cells in the Dentate Gyrus of Balb/C Mice. Int. J. Mol. Sci. 2018, 20, 73. [Google Scholar] [CrossRef]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on Melatonin Receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef] [PubMed]
- Delcourt, N.; Bockaert, J.; Marin, P. GPCR-Jacking: From a New Route in RTK Signalling to a New Concept in GPCR Activation. Trends Pharmacol. Sci. 2007, 28, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Lemmon, M.A.; Schlessinger, J. Cell Signaling by Receptor Tyrosine Kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [PubMed]
- Deinhardt, K.; Kim, T.; Spellman, D.S.; Mains, R.E.; Eipper, B.A.; Neubert, T.A.; Chao, M.V.; Hempstead, B.L. Neuronal Growth Cone Retraction Relies on Proneurotrophin Receptor Signaling through Rac. Sci. Signal. 2011, 4, ra82. [Google Scholar] [CrossRef]
- Barford, K.; Deppmann, C.; Winckler, B. The Neurotrophin Receptor Signaling Endosome: Where Trafficking Meets Signaling. Dev. Neurobiol. 2017, 77, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, M.V.; Melo, C.V.; Pereira, D.B.; Carvalho, R.F.; Carvalho, A.L.; Duarte, C.B. BDNF Regulates the Expression and Traffic of NMDA Receptors in Cultured Hippocampal Neurons. Mol. Cell. Neurosci. 2007, 35, 208–219. [Google Scholar] [CrossRef]
- Schinder, A.F.; Berninger, B.; Poo, M. ming Postsynaptic Target Specificity of Neurotrophin-Induced Presynaptic Potentiation. Neuron 2000, 25, 151–163. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Delagrange, P.; Krause, D.N.; Sugden, D.; Cardinali, D.P.; Olcese, J. International Union of Basic and Clinical Pharmacology. LXXV. Nomenclature, Classification, and Pharmacology of G Protein-Coupled Melatonin Receptors. Pharmacol. Rev. 2010, 63, 343–380. [Google Scholar] [CrossRef]
- Hardeland, R. Melatonin: Signaling Mechanisms of a Pleiotropic Agent. Biofactors 2009, 35, 183–192. [Google Scholar] [CrossRef]
- Liu, D.; Wei, N.; Man, H.Y.; Lu, Y.; Zhu, L.Q.; Wang, J.Z. The MT2 Receptor Stimulates Axonogenesis and Enhances Synaptic Transmission by Activating Akt Signaling. Cell Death Differ. 2015, 22, 583–596. [Google Scholar] [CrossRef]
- Liu, J.; Clough, S.J.; Dubocovich, M.L. Role of the MT1 and MT2 Melatonin Receptors in Mediating Depressive- and Anxiety-like Behaviors in C3H/HeN Mice. Genes Brain Behav. 2017, 16, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Tocharus, C.; Puriboriboon, Y.; Junmanee, T.; Tocharus, J.; Ekthuwapranee, K.; Govitrapong, P. Melatonin Enhances Adult Rat Hippocampal Progenitor Cell Proliferation via ERK Signaling Pathway through Melatonin Receptor. Neuroscience 2014, 275, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Mendivil-Perez, M.; Soto-Mercado, V.; Guerra-Librero, A.; Fernandez-Gil, B.I.; Florido, J.; Shen, Y.Q.; Tejada, M.A.; Capilla-Gonzalez, V.; Rusanova, I.; Garcia-Verdugo, J.M.; et al. Melatonin Enhances Neural Stem Cell Differentiation and Engraftment by Increasing Mitochondrial Function. J. Pineal Res. 2017, 63, e12415. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef]
- De Faria Poloni, J.; Feltes, B.C.; Bonatto, D. Melatonin as a Central Molecule Connecting Neural Development and Calcium Signaling. Funct. Integr. Genom. 2011, 11, 383–388. [Google Scholar] [CrossRef]
- Phonchai, R.; Phermthai, T.; Kitiyanant, N.; Suwanjang, W.; Kotchabhakdi, N.; Chetsawang, B. Potential Effects and Molecular Mechanisms of Melatonin on the Dopaminergic Neuronal Differentiation of Human Amniotic Fluid Mesenchymal Stem Cells. Neurochem. Int. 2019, 124, 82–93. [Google Scholar] [CrossRef]
- Ortiz-López, L.; González-Olvera, J.J.; Vega-Rivera, N.M.; García-Anaya, M.; Carapia-Hernández, A.K.; Velázquez-Escobar, J.C.; Ramírez-Rodríguez, G.B. Human Neural Stem/Progenitor Cells Derived from the Olfactory Epithelium Express the TrkB Receptor and Migrate in Response to BDNF. Neuroscience 2017, 355, 84–100. [Google Scholar] [CrossRef]
- Ghareghani, M.; Sadeghi, H.; Zibara, K.; Danaei, N.; Azari, H.; Ghanbari, A. Melatonin Increases Oligodendrocyte Differentiation in Cultured Neural Stem Cells. Cell. Mol. Neurobiol. 2017, 37, 1319–1324. [Google Scholar] [CrossRef]
- Argueta, J.; Solís-Chagoyán, H.; Estrada-Reyes, R.; Constantino-Jonapa, L.A.; Oikawa-Sala, J.; Velázquez-Moctezuma, J.; Benítez-King, G. Further Evidence of the Melatonin Calmodulin Interaction: Effect on CaMKII Activity. Int. J. Mol. Sci. 2022, 23, 2479. [Google Scholar] [CrossRef]
- Assimakopoulou, M.; Kondyli, M.; Gatzounis, G.; Maraziotis, T.; Varakis, J. Neurotrophin Receptors Expression and JNK Pathway Activation in Human Astrocytomas. BMC Cancer 2007, 7, 202. [Google Scholar] [CrossRef]
- Wang, J.; Xiao, X.; Zhang, Y.; Shi, D.; Chen, W.; Fu, L.; Liu, L.; Xie, F.; Kang, T.; Huang, W.; et al. Simultaneous Modulation of COX-2, P300, Akt, and Apaf-1 Signaling by Melatonin to Inhibit Proliferation and Induce Apoptosis in Breast Cancer Cells. J. Pineal Res. 2012, 53, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Santarelli, L.; Saxe, M.; Gross, C.; Surget, A.; Battaglia, F.; Dulawa, S.; Weisstaub, N.; Lee, J.; Duman, R.; Arancio, O.; et al. Requirement of Hippocampal Neurogenesis for the Behavioral Effects of Antidepressants. Science 2003, 301, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Hare, B.D.; Duman, R.S. Prefrontal Cortex Circuits in Depression and Anxiety: Contribution of Discrete Neuronal Populations and Target Regions. Mol. Psychiatry 2020, 25, 2742–2758. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.; Baktir, M.A.; Srivatsan, M.; Salehi, A. Neuroprotective Effects of Physical Activity on the Brain: A Closer Look at Trophic Factor Signaling. Front. Cell. Neurosci. 2014, 8, 170. [Google Scholar] [CrossRef]
- Kowiański, P.; Lietzau, G.; Czuba, E.; Waśkow, M.; Steliga, A.; Moryś, J. BDNF: A Key Factor with Multipotent Impact on Brain Signaling and Synaptic Plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Numakawa, T.; Odaka, H.; Adachi, N. Actions of Brain-Derived Neurotrophin Factor in the Neurogenesis and Neuronal Function, and Its Involvement in the Pathophysiology of Brain Diseases. Int. J. Mol. Sci. 2018, 19, 3650. [Google Scholar] [CrossRef]
- Labban, S.; Alshehri, F.S.; Kurdi, M.; Alatawi, Y.; Alghamdi, B.S. Melatonin Improves Short-Term Spatial Memory in a Mouse Model of Alzheimer’s Disease. Degener. Neurol. Neuromuscul. Dis. 2021, 11, 15–27. [Google Scholar] [CrossRef]
- Labban, S.; Alghamdi, B.S.; Alshehri, F.S.; Kurdi, M. Effects of Melatonin and Resveratrol on Recognition Memory and Passive Avoidance Performance in a Mouse Model of Alzheimer’s Disease. Behav. Brain Res. 2021, 402, 113100. [Google Scholar] [CrossRef]
- Guaiana, G.; Gupta, S.; Chiodo, D.; Davies, S.J.; Haederle, K.; Koesters, M. Agomelatine versus Other Antidepressive Agents for Major Depression. Cochrane Database Syst. Rev. 2013, 2013, CD008851. [Google Scholar] [CrossRef]
- Taniguti, E.H.; Ferreira, Y.S.; Stupp, I.J.V.; Fraga-Junior, E.B.; Mendonça, C.B.; Rossi, F.L.; Ynoue, H.N.; Doneda, D.L.; Lopes, L.; Lima, E.; et al. Neuroprotective Effect of Melatonin against Lipopolysaccharide-Induced Depressive-like Behavior in Mice. Physiol. Behav. 2018, 188, 270–275. [Google Scholar] [CrossRef]
- Estrada-Reyes, R.; Valdés-Tovar, M.; Arrieta-Baez, D.; Dorantes-Barrón, A.M.; Quero-Chávez, D.; Solís-Chagoyán, H.; Argueta, J.; Dubocovich, M.L.; Benítez-King, G. The Timing of Melatonin Administration Is Crucial for Its Antidepressant-like Effect in Mice. Int. J. Mol. Sci. 2018, 19, 2278. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Rodríguez, G.; Vega-Rivera, N.M.; Oikawa-Sala, J.; Gómez-Sánchez, A.; Ortiz-López, L.; Estrada-Camarena, E. Melatonin Synergizes with Citalopram to Induce Antidepressant-like Behavior and to Promote Hippocampal Neurogenesis in Adult Mice. J. Pineal Res. 2014, 56, 450–461. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.A.R.; Al-Khalifa, I.I.; Jasim, N.A.; Gorial, F.I. Adjuvant Use of Melatonin for Treatment of Fibromyalgia. J. Pineal Res. 2011, 50, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Cuzzocrea, S.; Zingarelli, B.; Gilad, E.; Hake, P.; Salzman, A.L.; Szabó, C. Protective Effect of Melatonin in Carrageenan-Induced Models of Local Inflammation: Relationship to Its Inhibitory Effect on Nitric Oxide Production and Its Peroxynitrite Scavenging Activity. J. Pineal Res. 1997, 23, 106–116. [Google Scholar] [CrossRef]
- Bilici, D.; Akpinar, E.; Kiziltunç, A. Protective effect of melatonin in carrageenan-induced acute local inflammation. Pharmacol. Res. 2002, 46, 133–139. [Google Scholar] [CrossRef]
- Posa, L.; De Gregorio, D.; Gobbi, G.; Comai, S. Targeting Melatonin MT2 Receptors: A Novel Pharmacological Avenue for Inflammatory and Neuropathic Pain. Curr. Med. Chem. 2018, 25, 3866–3882. [Google Scholar] [CrossRef]
- Posa, L.; Lopez-Canul, M.; Rullo, L.; De Gregorio, D.; Dominguez-Lopez, S.; Kaba Aboud, M.; Caputi, F.F.; Candeletti, S.; Romualdi, P.; Gobbi, G. Nociceptive Responses in Melatonin MT2 Receptor Knockout Mice Compared to MT1 and Double MT1/MT2 Receptor Knockout Mice. J. Pineal Res. 2020, 69, e12671. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, T.; Lee, T.H. Cellular Mechanisms of Melatonin: Insight from Neurodegenerative Diseases. Biomolecules 2020, 10, 1158. [Google Scholar] [CrossRef]
- Wander, C.M.; Song, J. The Neurogenic Niche in Alzheimer’s Disease. Neurosci. Lett. 2021, 762, 136109. [Google Scholar] [CrossRef]
- Rosales-Corral, S.; Acuna-Castroviejo, D.; Tan, D.X.; López-Armas, G.; Cruz-Ramos, J.; Munoz, R.; Melnikov, V.G.; Manchester, L.C.; Reiter, R.J. Accumulation of Exogenous Amyloid- Beta Peptide in Hippocampal Mitochondria Causes Their Dysfunction: A Protective Role for Melatonin. Oxidative Med. Cell. Longev. 2012, 2012, 843649. [Google Scholar] [CrossRef]
- Permpoonputtana, K.; Tangweerasing, P.; Mukda, S.; Boontem, P.; Nopparat, C.; Govitrapong, P. Long-Term Administration of Melatonin Attenuates Neuroinflammation in the Aged Mouse Brain. EXCLI J. 2018, 17, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, S.; Salari, A.-A.; Abedi, A.; Mohammadi, P.; Amani, M. Melatonin Treatment Improves Cognitive Deficits by Altering Inflammatory and Neurotrophic Factors in the Hippocampus of Obese Mice. Physiol. Behav. 2022, 254, 113919. [Google Scholar] [CrossRef] [PubMed]
- Madhu, L.N.; Kodali, M.; Attaluri, S.; Shuai, B.; Melissari, L.; Rao, X.; Shetty, A.K. Melatonin Improves Brain Function in a Model of Chronic Gulf War Illness with Modulation of Oxidative Stress, NLRP3 Inflammasomes, and BDNF-ERK-CREB Pathway in the Hippocampus. Redox Biol. 2021, 43, 101973. [Google Scholar] [CrossRef] [PubMed]
- Suwannakot, K.; Sritawan, N.; Naewla, S.; Aranarochana, A.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Melatonin Attenuates Methotrexate-Induced Reduction of Antioxidant Activity Related to Decreases of Neurogenesis in Adult Rat Hippocampus and Prefrontal Cortex. Oxidative Med. Cell. Longev. 2022, 2022, 1596362. [Google Scholar] [CrossRef] [PubMed]
- Aranarochana, A.; Sirichoat, A.; Pannangrong, W.; Wigmore, P.; Welbat, J.U. Melatonin Ameliorates Valproic Acid-Induced Neurogenesis Impairment: The Role of Oxidative Stress in Adult Rats. Oxidative Med. Cell. Longev. 2021, 2021, 9997582. [Google Scholar] [CrossRef]
- Sugiyama, A.; Kato, H.; Takakura, H.; Osawa, S.; Maeda, Y.; Izawa, T. Effects of Physical Activity and Melatonin on Brain-derived Neurotrophic Factor and Cytokine Expression in the Cerebellum of High-fat Diet-fed Rats. Neuropsychopharmacol. Rep. 2020, 40, 291–296. [Google Scholar] [CrossRef]
- Mateș, L.; Popa, D.-S.; Rusu, M.E.; Fizeșan, I.; Leucuța, D. Walnut Intake Interventions Targeting Biomarkers of Metabolic Syndrome and Inflammation in Middle-Aged and Older Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Antioxidants 2022, 11, 1412. [Google Scholar] [CrossRef]
- López-Armas, G.; Flores-Soto, M.E.; Chaparro-Huerta, V.; Jave-Suarez, L.F.; Soto-Rodríguez, S.; Rusanova, I.; Acuña-Castroviejo, D.; González-Perez, O.; González-Castañeda, R.E. Prophylactic Role of Oral Melatonin Administration on Neurogenesis in Adult Balb/C Mice during REM Sleep Deprivation. Oxidative Med. Cell. Longev. 2016, 2016, 2136902. [Google Scholar] [CrossRef]
- Kim, W.; Hahn, K.R.; Jung, H.Y.; Kwon, H.J.; Nam, S.M.; Kim, J.W.; Park, J.H.; Yoo, D.Y.; Kim, D.W.; Won, M.H.; et al. Melatonin Ameliorates Cuprizone-Induced Reduction of Hippocampal Neurogenesis, Brain-Derived Neurotrophic Factor, and Phosphorylation of Cyclic AMP Response Element-Binding Protein in the Mouse Dentate Gyrus. Brain Behav. 2019, 9, e01388. [Google Scholar] [CrossRef]
- Ambriz-Tututi, M.; Granados-Soto, V. Oral and Spinal Melatonin Reduces Tactile Allodynia in Rats via Activation of MT2 and Opioid Receptors. Pain 2007, 132, 273–280. [Google Scholar] [CrossRef]
- Ortiz-López, L.; Pérez-Beltran, C.; Ramírez-Rodríguez, G. Chronic Administration of a Melatonin Membrane Receptor Antagonist, Luzindole, Affects Hippocampal Neurogenesis without Changes in Hopelessness-like Behavior in Adult Mice. Neuropharmacology 2016, 103, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Sotthibundhu, A.; Ekthuwapranee, K.; Govitrapong, P. Comparison of Melatonin with Growth Factors in Promoting Precursor Cells Proliferation in Adult Mouse Subventricular Zone. EXCLI J. 2016, 15, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Bae, J.H.; Lee, J.H.; Kim, Y.N.; Kim, D.K. The Melatonin Signaling Pathway in a Long-Term Memory In Vitro Study. Molecules 2018, 23, 737. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MEL Neurotrophic-like Characteristics | References |
|---|---|
| MEL has membrane receptors coupled to G protein | [54,61] |
| MEL promotes neuronal differentiation, proliferation, and neuronal survival in newly formed neurons | [39,40,41,63,64,65,66,69,71,104,105] |
| MEL participates in mature brain neuroplasticity and neurodevelopment by stimulating second messenger cascades (Ca2+, CaMKII, Trk receptors) | [63,65,69,80,81,106] |
| Exerts antidepressant and anxiolytic-like effects associated to increased neurogenesis | [44,45,50,64,80,81,85] |
| Adjuvant in antidepressant treatments due to its synergistic effect with other molecules administered as antidepressant such as SSRIs (fluoxetine, citalopram) or ketamine | [43,44,85,86] |
| MEL can be used as neuroprotector in diseases whose treatments cause toxicity | [97,98] |
| MEL has anti-inflammatory properties and restorative effects in toxin-induced rodent models of persistent/chronic and neuropathic pain and spinal cord injury | [66,87,88,89,90,94,95,96,103] |
| MEL protects and maintains immune cells in non-neuronal places, like NTFs does | [14,19] |
| MEL can change the reorganization of the cytoskeleton | [46,47] |
| Promotes neuron’s maturation, neuritogenesis, dendritic growth, and axogenesis. MEL also increases the complexity of the dendrite trees in crucial brain regions affected in neurodegenerative diseases. | [37,45,46,47,48,49,50,51] |
| Free radical scavenging properties of MEL equip it with antioxidant and neuroprotective effects. MEL modulates the production of other antioxidant molecules | [17,19,66,87,88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miranda-Riestra, A.; Estrada-Reyes, R.; Torres-Sanchez, E.D.; Carreño-García, S.; Ortiz, G.G.; Benítez-King, G. Melatonin: A Neurotrophic Factor? Molecules 2022, 27, 7742. https://doi.org/10.3390/molecules27227742
Miranda-Riestra A, Estrada-Reyes R, Torres-Sanchez ED, Carreño-García S, Ortiz GG, Benítez-King G. Melatonin: A Neurotrophic Factor? Molecules. 2022; 27(22):7742. https://doi.org/10.3390/molecules27227742
Chicago/Turabian StyleMiranda-Riestra, Armida, Rosa Estrada-Reyes, Erandis D. Torres-Sanchez, Silvia Carreño-García, Genaro Gabriel Ortiz, and Gloria Benítez-King. 2022. "Melatonin: A Neurotrophic Factor?" Molecules 27, no. 22: 7742. https://doi.org/10.3390/molecules27227742
APA StyleMiranda-Riestra, A., Estrada-Reyes, R., Torres-Sanchez, E. D., Carreño-García, S., Ortiz, G. G., & Benítez-King, G. (2022). Melatonin: A Neurotrophic Factor? Molecules, 27(22), 7742. https://doi.org/10.3390/molecules27227742