

A Branched and Double Alpha-Gal-Bearing Synthetic Neoglycoprotein as a Biomarker for Chagas Disease
 , , , , ,
, , , , ,  , , ,
, , ,  add
Show full author list
add
Show full author list
Abstract
1. Introduction
2. Results and Discussion
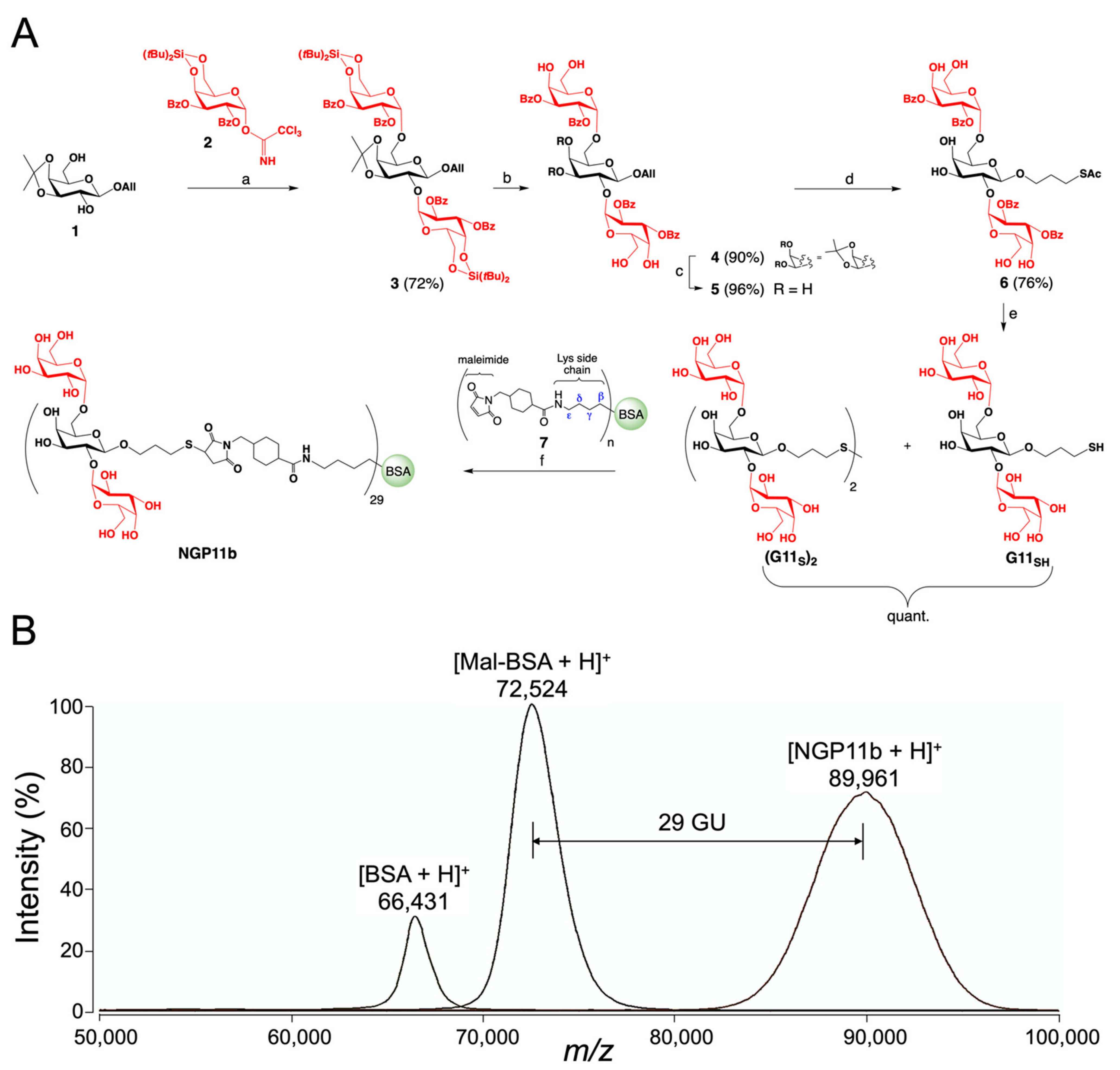
2.1. Chemical Synthesis of NGP11b
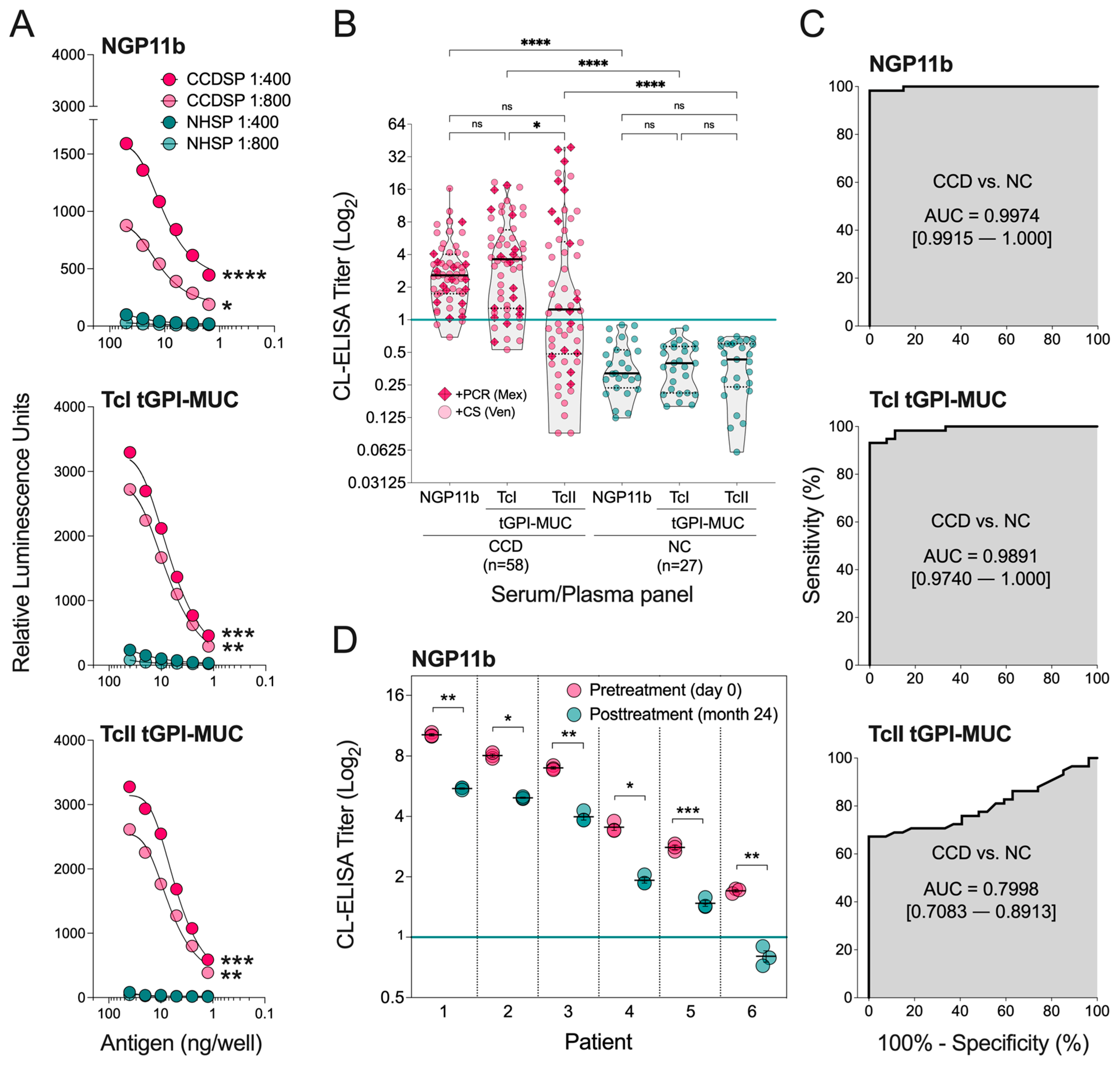
2.2. Evaluation of NGP11b as a Biomarker for Chagas Disease by CL-ELISA
3. Materials and Methods
3.1. General Information
3.2. Carbohydrate Synthesis and Characterization
3.3. Conjugation of the Glycan with Maleimide-Derivatized Bovine Serum Albumin (BSA)
3.4. MALDI-TOF-MS
3.5. Purification of tGPI-MUC from T. cruzi Colombiana (DTU TcI) and (DTU TcII) Y Strains
3.6. Human Serum and Plasma Samples
3.7. CL-ELISA
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Committee, W.H.O.E. Control of Chagas disease. World Health Organ. Tech. Rep. Ser. 2002, 905, 1–109. [Google Scholar]
- Carod-Artal, F.J.; Gascon, J. Chagas disease and stroke. Lancet Neurol. 2010, 9, 533–542. [Google Scholar] [CrossRef]
- Perez-Molina, J.A.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Perez-Molina, J.A.; Norman, F.; Lopez-Velez, R. Chagas disease in non-endemic countries: Epidemiology, clinical presentation and treatment. Curr. Infect. Dis. Rep. 2012, 14, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Schmunis, G.A. Epidemiology of Chagas disease in non-endemic countries: The role of international migration. Mem. Inst. Oswaldo Cruz. 2007, 102 (Suppl. 1), 75–86. [Google Scholar] [CrossRef]
- Manne-Goehler, J.; Umeh, C.A.; Montgomery, S.P.; Wirtz, V.J. Estimating the Burden of Chagas Disease in the United States. PLoS Negl. Trop. Dis. 2016, 10, e0005033. [Google Scholar] [CrossRef] [PubMed]
- Bern, C.; Montgomery, S.P. An estimate of the burden of Chagas disease in the United States. Clin. Infect. Dis. 2009, 49, e52–e54. [Google Scholar] [CrossRef]
- Rassi, A., Jr.; Rassi, A.; Marin-Neto, J.A. Chagas disease. Lancet 2010, 375, 1388–1402. [Google Scholar] [CrossRef]
- Bern, C.; Kjos, S.; Yabsley, M.J.; Montgomery, S.P. Trypanosoma cruzi and Chagas’ Disease in the United States. Clin. Microbiol. Rev. 2011, 24, 655–681. [Google Scholar] [CrossRef]
- de Noya, B.A.; Gonzalez, O.N. An ecological overview on the factors that drives to Trypanosoma cruzi oral transmission. Acta Trop. 2015, 151, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Coura, J.R.; Borges-Pereira, J. Chagas disease: 100 years after its discovery. A systemic review. Acta Trop. 2010, 115, 5–13. [Google Scholar] [CrossRef]
- Custer, B.; Agapova, M.; Bruhn, R.; Cusick, R.; Kamel, H.; Tomasulo, P.; Biswas, H.; Tobler, L.; Lee, T.H.; Caglioti, S.; et al. Epidemiologic and laboratory findings from 3 years of testing United States blood donors for Trypanosoma cruzi. Transfusion. 2012, 52, 1901–1911. [Google Scholar] [CrossRef] [PubMed]
- Zingales, B. Trypanosoma cruzi genetic diversity: Something new for something known about Chagas disease manifestations, serodiagnosis and drug sensitivity. Acta Trop. 2018, 184, 38–52. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, L.; Gollob, K.; Zingales, B.; Dutra, W. Pathogen diversity, immunity, and the fate of infections: Lessons learned from Trypanosoma cruzi human-host interactions. Lancet Microbe 2022, 3, e711–e722. [Google Scholar] [CrossRef]
- Urbina, J.A. Recent clinical trials for the etiological treatment of chronic chagas disease: Advances, challenges and perspectives. J. Eukaryot. Microbiol. 2015, 62, 149–156. [Google Scholar] [CrossRef]
- Viotti, R.; Vigliano, C.; Lococo, B.; Alvarez, M.G.; Petti, M.; Bertocchi, G.; Armenti, A. Side effects of benznidazole as treatment in chronic Chagas disease: Fears and realities. Expert Rev. Anti Infect. Ther. 2009, 7, 157–163. [Google Scholar] [CrossRef]
- Pinazo, M.J.; Thomas, M.C.; Bua, J.; Perrone, A.; Schijman, A.G.; Viotti, R.J.; Ramsey, J.M.; Ribeiro, I.; Sosa-Estani, S.; Lopez, M.C.; et al. Biological markers for evaluating therapeutic efficacy in Chagas disease, a systematic review. Expert Rev. Anti Infect. Ther. 2014, 12, 479–496. [Google Scholar] [CrossRef]
- Ribeiro, I.; Sevcsik, A.M.; Alves, F.; Diap, G.; Don, R.; Harhay, M.O.; Chang, S.; Pecoul, B. New, improved treatments for Chagas disease: From the R&D pipeline to the patients. PLoS Negl. Trop. Dis. 2009, 3, e484. [Google Scholar]
- Buscaglia, C.A.; Campo, V.A.; Frasch, A.C.; Di Noia, J.M. Trypanosoma cruzi surface mucins: Host-dependent coat diversity. Nat. Rev. Microbiol. 2006, 4, 229–236. [Google Scholar] [CrossRef]
- Frasch, A.C. Functional diversity in the trans-sialidase and mucin families in Trypanosoma cruzi. Parasitol. Today. 2000, 16, 282–286. [Google Scholar] [CrossRef]
- De Pablos, L.M.; Osuna, A. Multigene families in Trypanosoma cruzi and their role in infectivity. Infect. Immun. 2012, 80, 2258–2264. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Serrano, A.; Almeida, I.C.; Freitas-Junior, L.H.; Yoshida, N.; Schenkman, S. The mucin-like glycoprotein super-family of Trypanosoma cruzi: Structure and biological roles. Mol. Biochem. Parasitol. 2001, 114, 143–150. [Google Scholar] [CrossRef]
- Teixeira, S.M.; El-Sayed, N.M.; Araujo, P.R. The genome and its implications. Adv. Parasitol. 2011, 75, 209–230. [Google Scholar] [PubMed]
- Almeida, I.C.; Ferguson, M.A.; Schenkman, S.; Travassos, L.R. Lytic anti-alpha-galactosyl antibodies from patients with chronic Chagas’ disease recognize novel O-linked oligosaccharides on mucin-like glycosyl-phosphatidylinositol-anchored glycoproteins of Trypanosoma cruzi. Biochem. J. 1994, 304, 793–802. [Google Scholar] [CrossRef]
- Galili, U. Anti-Gal in Humans and Its Antigen the α-Gal Epitope. In The Natural Anti-Gal Antibody As Foe Turned Friend In Medicine, 1st ed.; Galili, U., Ed.; Academic Press: Cambridge, MA, USA, 2017; pp. 3–22. [Google Scholar]
- Galili, U.; Swanson, K. Gene sequences suggest inactivation of alpha-1,3-galactosyltransferase in catarrhines after the divergence of apes from monkeys. Proc. Natl. Acad. Sci. USA 1991, 88, 7401–7404. [Google Scholar] [CrossRef]
- Almeida, I.C.; Milani, S.R.; Gorin, P.A.; Travassos, L.R. Complement-mediated lysis of Trypanosoma cruzi trypomastigotes by human anti-alpha-galactosyl antibodies. J. Immunol. 1991, 146, 2394–2400. [Google Scholar] [PubMed]
- Gazzinelli, R.T.; Pereira, M.E.; Romanha, A.; Gazzinelli, G.; Brener, Z. Direct lysis of Trypanosoma cruzi: A novel effector mechanism of protection mediated by human anti-gal antibodies. Parasite Immunol. 1991, 13, 345–356. [Google Scholar] [CrossRef]
- Avila, J.L.; Rojas, M.; Galili, U. Immunogenic Gal alpha 1—3Gal carbohydrate epitopes are present on pathogenic American. Trypanos. Leishmania. J. Immunol. 1989, 142, 2828–2834. [Google Scholar]
- Milani, S.R.; Travassos, L.R. Anti-alpha-galactosyl antibodies in chagasic patients. Possible biological significance. Braz. J. Med. Biol. Res. 1988, 21, 1275–1286. [Google Scholar]
- Almeida, I.C.; Covas, D.T.; Soussumi, L.M.; Travassos, L.R. A highly sensitive and specific chemiluminescent enzyme-linked immunosorbent assay for diagnosis of active Trypanosoma cruzi infection. Transfusion 1997, 37, 850–857. [Google Scholar] [CrossRef]
- Almeida, I.C.; Krautz, G.M.; Krettli, A.U.; Travassos, L.R. Glycoconjugates of Trypanosoma cruzi: A 74 kD antigen of trypomastigotes specifically reacts with lytic anti-alpha-galactosyl antibodies from patients with chronic Chagas disease. J. Clin. Lab. Anal. 1993, 7, 307–316. [Google Scholar] [CrossRef]
- Torrico, F.; Gascon, J.; Ortiz, L.; Alonso-Vega, C.; Pinazo, M.J.; Schijman, A.; Almeida, I.C.; Alves, F.; Strub-Wourgaft, N.; Ribeiro, I.; et al. Treatment of adult chronic indeterminate Chagas disease with benznidazole and three E1224 dosing regimens: A proof-of-concept, randomised, placebo-controlled trial. Lancet Infect. Dis. 2018, 18, 419–430. [Google Scholar] [CrossRef]
- Izquierdo, L.; Marques, A.F.; Gallego, M.; Sanz, S.; Tebar, S.; Riera, C.; Quinto, L.; Aldasoro, E.; Almeida, I.C.; Gascon, J. Evaluation of a chemiluminescent enzyme-linked immunosorbent assay for the diagnosis of Trypanosoma cruzi infection in a nonendemic setting. Mem. Inst. Oswaldo Cruz 2013, 108, 928–931. [Google Scholar] [CrossRef][Green Version]
- Torrico, F.; Gascon, J.; Barreira, F.; Blum, B.; Almeida, I.C.; Alonso-Vega, C.; Barboza, T.; Bilbe, G.; Correia, E.; Garcia, W.; et al. Bendita study group, New regimens of benznidazole monotherapy and in combination with fosravuconazole for treatment of Chagas disease (BENDITA): A phase 2, double-blind, randomised trial. Lancet Infect. Dis. 2021, 21, 1129–1140. [Google Scholar] [CrossRef]
- Pinazo, M.J.; Posada Ede, J.; Izquierdo, L.; Tassies, D.; Marques, A.F.; de Lazzari, E.; Aldasoro, E.; Munoz, J.; Abras, A.; Tebar, S.; et al. Altered Hypercoagulability Factors in Patients with Chronic Chagas Disease: Potential Biomarkers of Therapeutic Response. PLoS Negl. Trop. Dis. 2016, 10, e0004269. [Google Scholar] [CrossRef]
- de Andrade, A.L.; Zicker, F.; de Oliveira, R.M.; Almeida Silva, S.; Luquetti, A.; Travassos, L.R.; Almeida, I.C.; de Andrade, S.S.; de Andrade, J.G.; Martelli, C.M. Randomised trial of efficacy of benznidazole in treatment of early Trypanosoma cruzi infection. Lancet 1996, 348, 1407–1413. [Google Scholar] [CrossRef]
- Andrade, A.L.; Martelli, C.M.; Oliveira, R.M.; Silva, S.A.; Aires, A.I.; Soussumi, L.M.; Covas, D.T.; Silva, L.S.; Andrade, J.G.; Travassos, L.R.; et al. Short report: Benznidazole efficacy among Trypanosoma cruzi-infected adolescents after a six-year follow-up. Am. J. Trop. Med. Hyg. 2004, 71, 594–597. [Google Scholar] [CrossRef]
- Ortega-Rodriguez, U.; Portillo, S.; Ashmus, R.A.; Duran, J.A.; Schocker, N.S.; Iniguez, E.; Montoya, A.L.; Zepeda, B.G.; Olivas, J.J.; Karimi, N.H.; et al. Purification of Glycosylphosphatidylinositol-Anchored Mucins from Trypanosoma cruzi Trypomastigotes and Synthesis of alpha-Gal-Containing Neoglycoproteins: Application as Biomarkers for Reliable Diagnosis and Early Assessment of Chemotherapeutic Outcomes of Chagas Disease. Methods Mol. Biol. 2019, 1955, 287–308. [Google Scholar]
- Ashmus, R.A.; Schocker, N.S.; Cordero-Mendoza, Y.; Marques, A.F.; Monroy, E.Y.; Pardo, A.; Izquierdo, L.; Gallego, M.; Gascon, J.; Almeida, I.C.; et al. Potential use of synthetic alpha-galactosyl-containing glycotopes of the parasite Trypanosoma cruzi as diagnostic antigens for Chagas disease. Org. Biomol. Chem. 2013, 11, 5579–5583. [Google Scholar] [CrossRef] [PubMed]
- Schocker, N.S.; Portillo, S.; Brito, C.R.; Marques, A.F.; Almeida, I.C.; Michael, K. Synthesis of Galalpha(1,3)Galbeta(1,4)GlcNAcalpha-, Galbeta(1,4)GlcNAcalpha- and GlcNAc-containing neoglycoproteins and their immunological evaluation in the context of Chagas disease. Glycobiology 2016, 26, 39–50. [Google Scholar]
- Montoya, A.L.; Gil, E.R.; Heydemann, E.L.; Estevao, I.L.; Luna, B.E.; Ellis, C.C.; Jankuru, S.R.; Alarcón de Noya, B.; Noya, O.; Zago, M.P.; et al. Specific Recognition of beta-Galactofuranose-Containing Glycans of Synthetic Neoglycoproteins by Sera of Chronic Chagas Disease Patients. Molecules 2022, 27, 411. [Google Scholar] [CrossRef] [PubMed]
- Montoya, A.L.; Austin, V.M.; Portillo, S.; Vinales, I.; Ashmus, R.A.; Estevao, I.; Jankuru, S.R.; Alraey, Y.; Al-Salem, W.S.; Acosta-Serrano, A.; et al. Reversed Immunoglycomics Identifies alpha-Galactosyl-Bearing Glycotopes Specific for Leishmania major Infection. JACS Au 2021, 1, 1275–1287. [Google Scholar] [CrossRef] [PubMed]
- Viana, S.M.; Montoya, A.L.; Carvalho, A.M.; de Mendonça, B.; Portillo, S.; Olivas, J.J.; Karimi, N.H.; Estevao, I.L.; Ortega-Rodriguez, U.; Carvalho, E.M.; et al. Serodiagnosis and therapeutic monitoring of New-World tegumentary leishmaniasis using synthetic type-2 glycoinositolphospholipid-based neoglycoproteins. Emerg. Microbes Infect. 2022. [Google Scholar] [CrossRef] [PubMed]
- Zingales, B.; Andrade, S.G.; Briones, M.R.; Campbell, D.A.; Chiari, E.; Fernandes, O.; Guhl, F.; Lages-Silva, E.; Macedo, A.M.; Machado, C.R.; et al. A new consensus for Trypanosoma cruzi intraspecific nomenclature: Second revision meeting recommends TcI to TcVI. Mem. Inst. Oswaldo Cruz 2009, 104, 1051–1054. [Google Scholar] [CrossRef] [PubMed]
- Schocker, N.S.; Portillo, S.; Ashmus, R.A.; Brito, C.R.N.; Silva, I.E.; Cordero-Mendoza, Y.; Marques, A.F.; Monroy, E.Y.; Pardo, A.; Izquierdo, L.; et al. Probing for Trypanosoma cruzi cell surface glycobiomarkers for the diagnosis and follow-up of chemotherapy of Chagas disease. In Coupling and Decoupling of Diverse Molecular Units in Glycosciences; Witzczak, Z.J., Bielski, R., Eds.; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 195–211. [Google Scholar]
- Imamura, A.; Matsuzawa, N.; Sakai, S.; Udagawa, T.; Nakashima, S.; Ando, H.; Ishida, H.; Kiso, M. The Origin of High Stereoselectivity in Di-tert-butylsilylene-Directed α-Galactosylation. J. Org. Chem. 2006, 81, 8086–9104. [Google Scholar] [CrossRef]
- Soares, R.P.; Torrecilhas, A.C.; Assis, R.R.; Rocha, M.N.; Moura e Castro, F.A.; Freitas, G.F.; Murta, S.M.; Santos, S.L.; Marques, A.F.; Almeida, I.C.; et al. Intraspecies variation in Trypanosoma cruzi GPI-mucins: Biological activities and differential expression of alpha-galactosyl residues. Am. J. Trop. Med. Hyg. 2012, 87, 87–96. [Google Scholar] [CrossRef]
- Chiodo, F.; Marradi, M.; Tefsen, B.; Snippe, H.; van Die, I.; Penades, S. High sensitive detection of carbohydrate binding proteins in an ELISA-solid phase assay based on multivalent glyconanoparticles. PLoS ONE 2013, 8, e73027. [Google Scholar] [CrossRef]
- Oyelaran, O.; Li, Q.; Farnsworth, D.; Gildersleeve, J.C. Microarrays with varying carbohydrate density reveal distinct subpopulations of serum antibodies. J. Proteome Res. 2009, 8, 3529–3538. [Google Scholar] [CrossRef]
- Geissner, A.; Seeberger, P.H. Glycan Arrays: From Basic Biochemical Research to Bioanalytical and Biomedical Applications. Annu. Rev. Anal. Chem. (Palo Alto Calif) 2016, 9, 223–247. [Google Scholar] [CrossRef]
- Alonso-Vega, C.; Urbina, J.A.; Sanz, S.; Pinazo, M.J.; Pinto, J.J.; Gonzalez, V.R.; Rojas, G.; Ortiz, L.; Garcia, W.; Lozano, D.; et al. New chemotherapy regimens and biomarkers for Chagas disease: The rationale and design of the TESEO study, an open-label, randomised, prospective, phase-2 clinical trial in the Plurinational State of Bolivia. BMJ Open 2021, 11, e052897. [Google Scholar] [CrossRef]
- Balcerzak, A.K.; Ferreira, S.S.; Trant, J.F.; Ben, R.N. Structurally diverse disaccharide analogs of antifreeze glycoproteins and their ability to inhibit ice recrystallization. Bioorg. Med. Chem. Lett. 2012, 22, 1719–1721. [Google Scholar] [CrossRef] [PubMed]
- Kimura, A.; Imamura, A.; Ando, H.; Ishida, H.; Kiso, M. A novel synthetic route to a-galactosyl ceramides and iGb3 using DTBS-directed a-selective galactosylation. Synlett 2006, 2379–2382. [Google Scholar] [CrossRef]
- Imamura, A.; Kimura, A.; Ando, H.; Ishida, H.; Kiso, M. Extended application of di-tert-butylsilylene-directed a-predominant galactosylation compatible with C2-participating groups toward the assembly of various glycosides. Chem. Eur. J. 2006, 12, 8862–8870. [Google Scholar] [CrossRef] [PubMed]
- Frey, A.; Di Canzio, J.; Zurakowski, D. A statistically defined endpoint titer determination method for immunoassays. J. Immunol. Methods 1998, 221, 35–41. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Disease/Control | n | NGP11b | TcI tGPI-MUC | TcII tGPI-MUC | |||
|---|---|---|---|---|---|---|---|
| Positive | Negative | Positive | Negative | Positive | Negative | ||
| Chronic Chagas disease | 58 | 55 (95.8%) | 3 (5.2%) | 51 (87.9%) | 7 (12.1%) | 32 (55.2%) | 26 (44.8%) |
| Healthy control a | 27 | 0 | 27 (100%) | 0 | 27 (100%) | 0 | 27 (100%) |
| Parameter a,b | NGP11b | TcI tGPI-MUC | TcII tGPI-MUC |
|---|---|---|---|
| % | |||
| Sensitivity | 94.8 | 87.9 | 55.2 |
| Specificity | 100.0 | 100.0 | 100.0 |
| FPR | 0.0 | 0.0 | 0.0 |
| PPV | 100.0 | 100.0 | 100.0 |
| NPV | 90.0 | 79.4 | 50.9 |
| Accuracy | 96.6 | 92.4 | 76.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montoya, A.L.; Carvajal, E.G.; Ortega-Rodriguez, U.; Estevao, I.L.; Ashmus, R.A.; Jankuru, S.R.; Portillo, S.; Ellis, C.C.; Knight, C.D.; Alonso-Padilla, J.; et al. A Branched and Double Alpha-Gal-Bearing Synthetic Neoglycoprotein as a Biomarker for Chagas Disease. Molecules 2022, 27, 5714. https://doi.org/10.3390/molecules27175714
Montoya AL, Carvajal EG, Ortega-Rodriguez U, Estevao IL, Ashmus RA, Jankuru SR, Portillo S, Ellis CC, Knight CD, Alonso-Padilla J, et al. A Branched and Double Alpha-Gal-Bearing Synthetic Neoglycoprotein as a Biomarker for Chagas Disease. Molecules. 2022; 27(17):5714. https://doi.org/10.3390/molecules27175714
Chicago/Turabian StyleMontoya, Alba L., Elisa G. Carvajal, Uriel Ortega-Rodriguez, Igor L. Estevao, Roger A. Ashmus, Sohan R. Jankuru, Susana Portillo, Cameron C. Ellis, Colin D. Knight, Julio Alonso-Padilla, and et al. 2022. "A Branched and Double Alpha-Gal-Bearing Synthetic Neoglycoprotein as a Biomarker for Chagas Disease" Molecules 27, no. 17: 5714. https://doi.org/10.3390/molecules27175714
APA StyleMontoya, A. L., Carvajal, E. G., Ortega-Rodriguez, U., Estevao, I. L., Ashmus, R. A., Jankuru, S. R., Portillo, S., Ellis, C. C., Knight, C. D., Alonso-Padilla, J., Izquierdo, L., Pinazo, M.-J., Gascon, J., Suarez, V., Watts, D. M., Malo, I. R., Ramsey, J. M., Alarcón De Noya, B., Noya, O., ... Michael, K. (2022). A Branched and Double Alpha-Gal-Bearing Synthetic Neoglycoprotein as a Biomarker for Chagas Disease. Molecules, 27(17), 5714. https://doi.org/10.3390/molecules27175714