
In the Swim of Cannabis: Developmental Toxicity and Metabolomic Pathway Alterations of Zebrafish Larvae Exposed to THC for the Assessment of Its Potential Environmental and Human Health Impact

 ,
,  ,
,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Zebrafish Housing and Husbandry
2.3. Zebrafish Toxicity Testing
2.4. Lethal Concentration (LC50) Determination
2.5. Heart Rate
2.6. Behavior Screening
2.6.1. Touching Motor Response (TMR)
2.6.2. Locomotor Activity
2.6.3. Vibrational Startle Response (VSR)
2.7. Metabolomic Study and Data Processing
2.7.1. Metabolite Extraction
2.7.2. Metabolome Study and Data Processing
2.8. Statistical Analyses
2.9. Ethics Statement
3. Results
3.1. Lethal Concentration (LC50) Determination
3.2. Morphological and Function Alterations
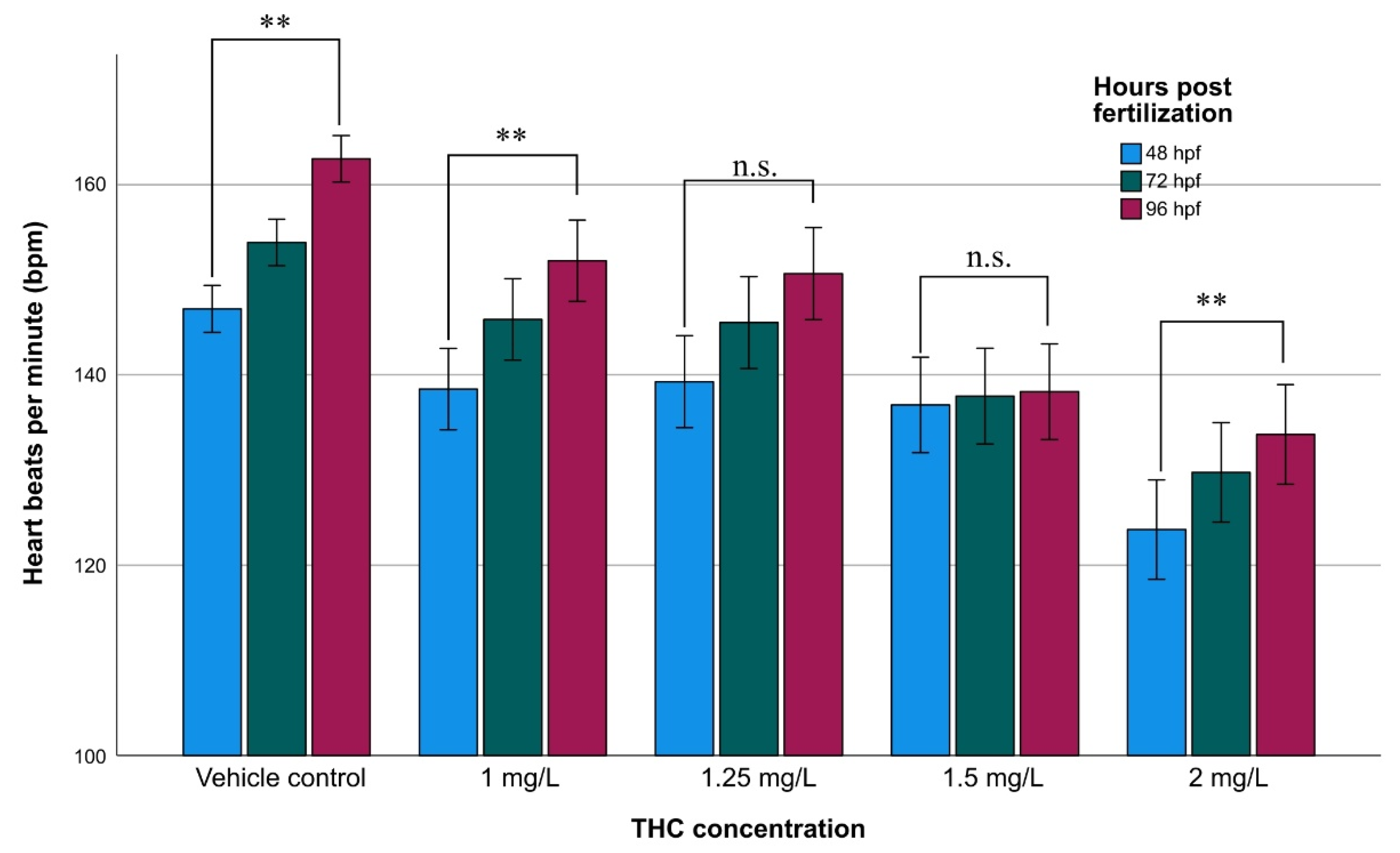
3.3. Heart Rate
3.4. Behavioral Analysis
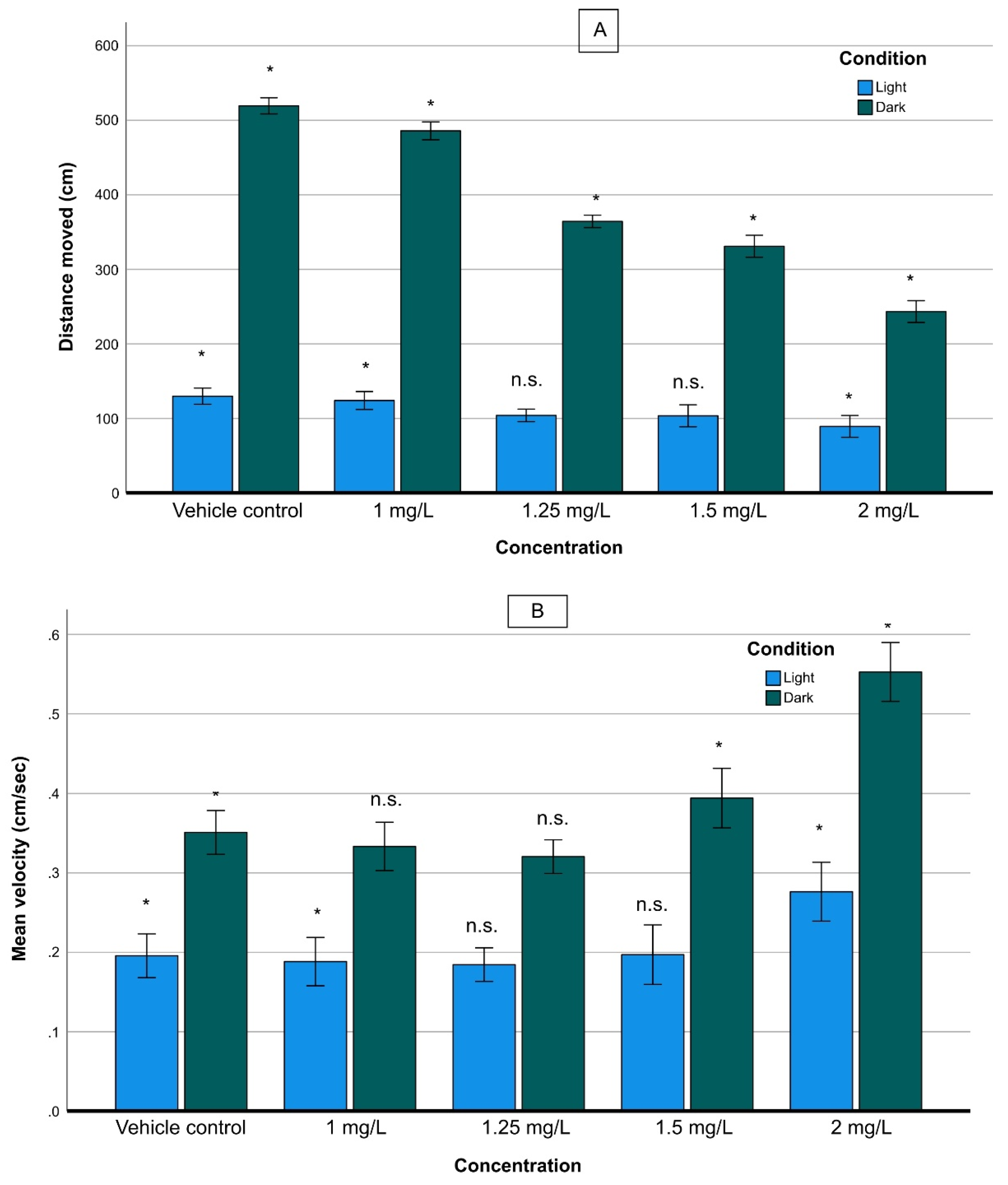
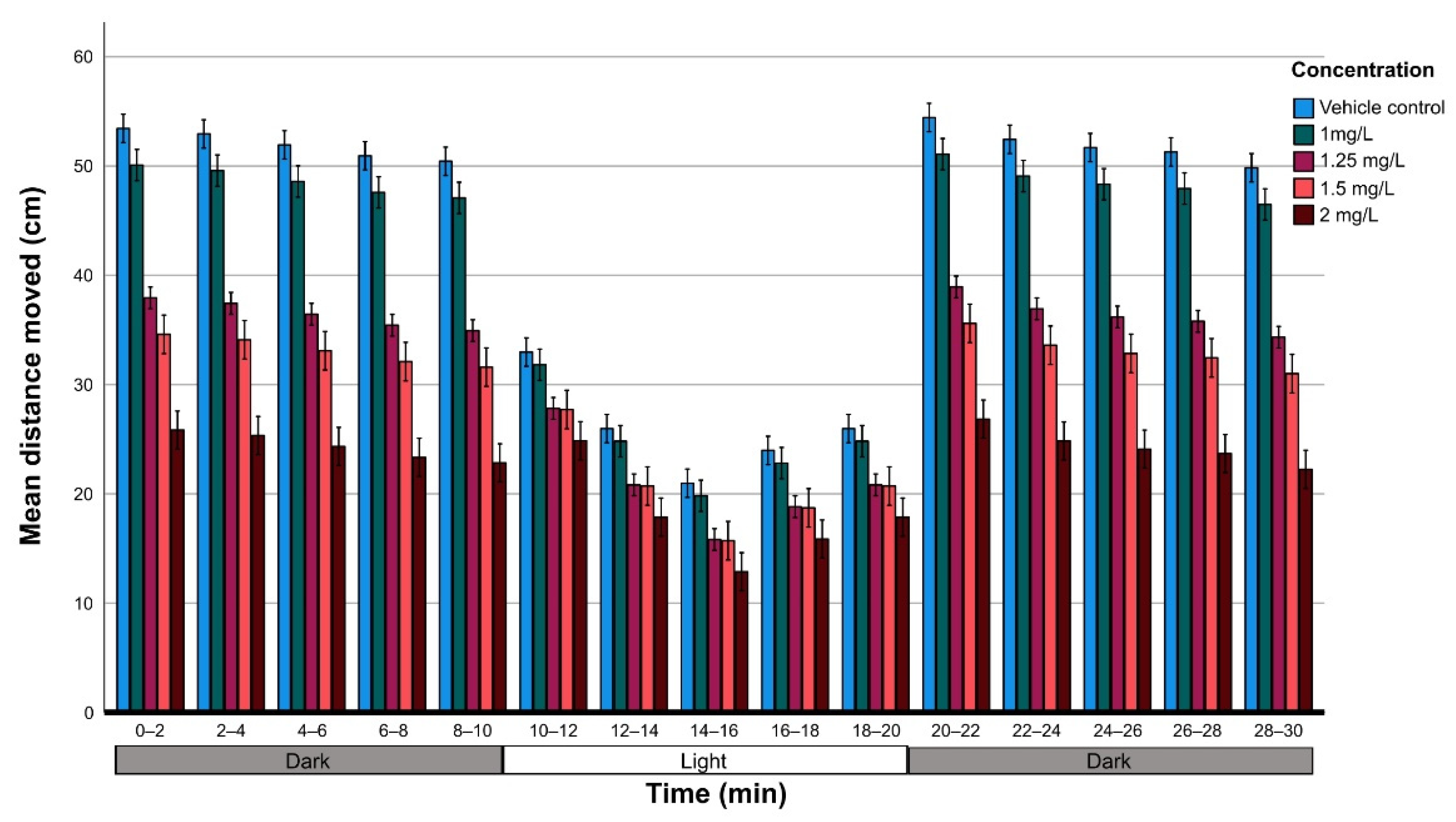
3.4.1. Larval Activity
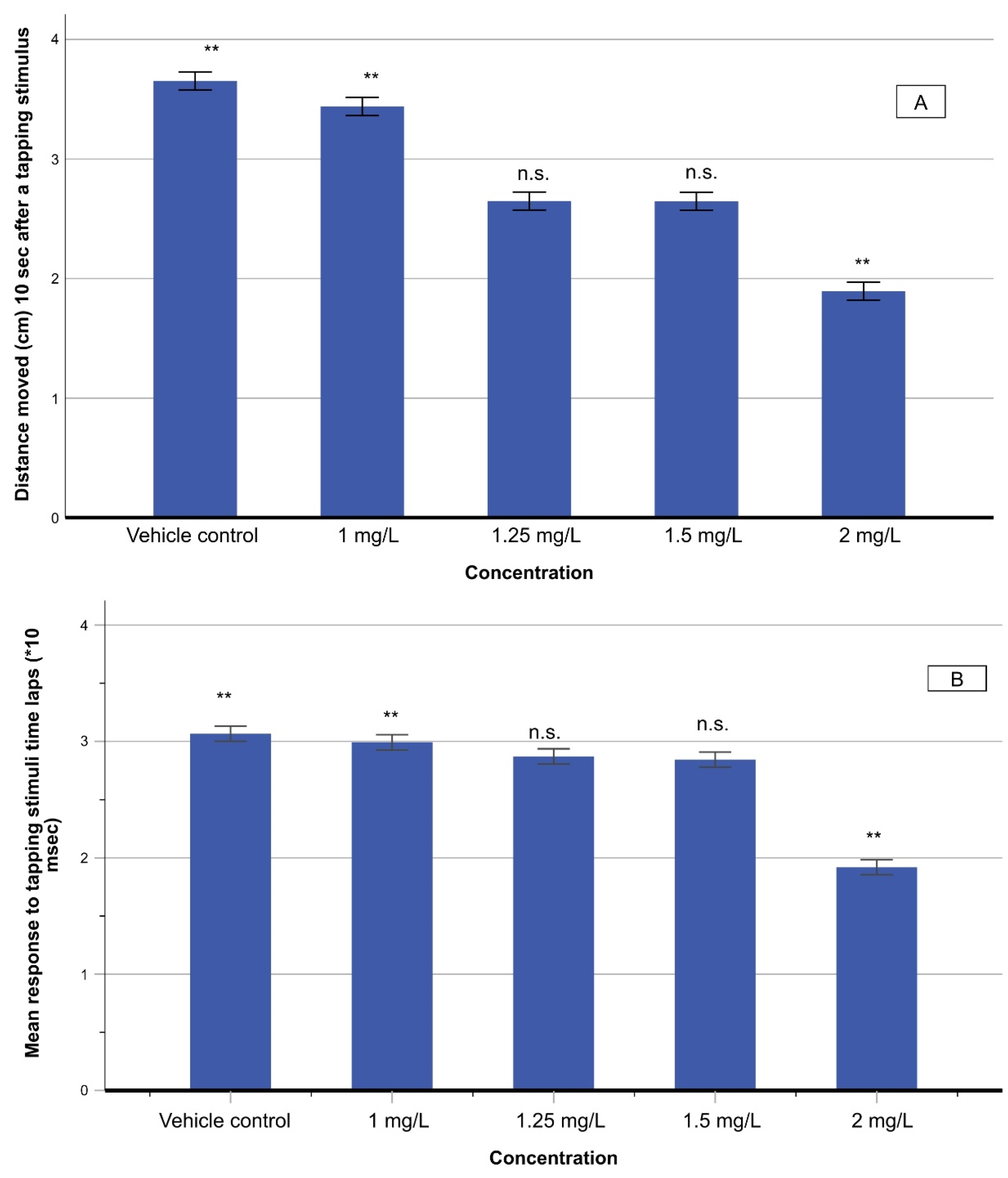
3.4.2. Touch Motor Response (TMR) and Vibrational Startle Response (VSR)
3.5. Metabolomic Study
4. Discussion
4.1. Behavior
4.2. Metabolic Alterations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis Sativa: A Comprehensive Ethnopharmacological Review of a Medicinal Plant with a Long History. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Handbook of Cannabis; Oxford University Press: New York, NY, USA, 2015; Volume 53. [Google Scholar]
- Fathordoobady, F.; Singh, A.; Kitts, D.D.; Pratap Singh, A. Hemp (Cannabis sativa L.) Extract: Anti-Microbial Properties, Methods of Extraction, and Potential Oral Delivery. Food Rev. Int. 2019, 35, 664–684. [Google Scholar] [CrossRef]
- Hill, M.N.; Gorzalka, B.B. Is There a Role for the Endocannabinoid System in the Etiology and Treatment of Melancholic Depression? Behav. Pharmacol. 2005, 16, 333–352. [Google Scholar] [CrossRef]
- Kerr, A.; Walston, V.; Wong, V.S.S.; Kellogg, M.; Ernst, L. Marijuana Use among Patients with Epilepsy at a Tertiary Care Center. Epilepsy Behav. 2019, 97, 144–148. [Google Scholar] [CrossRef]
- Devinsky, O.; Cross, J.H.; Laux, L.; Marsh, E.; Miller, I.; Nabbout, R.; Scheffer, I.E.; Thiele, E.A.; Wright, S. Trial of Cannabidiol for Drug-Resistant Seizures in the Dravet Syndrome. N. Engl. J. Med. 2017, 376, 2011–2020. [Google Scholar] [CrossRef] [PubMed]
- Kosiba, J.D.; Maisto, S.A.; Ditre, J.W. Patient-Reported Use of Medical Cannabis for Pain, Anxiety, and Depression Symptoms: Systematic Review and Meta-Analysis. Soc. Sci. Med. 2019, 233, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, I.; Monaghan, M. Cannabis and Psychosis: Are We Any Closer to Understanding the Relationship? Curr. Psychiatry Rep. 2019, 21, 48. [Google Scholar] [CrossRef]
- Boehnke, K.F.; Scott, J.R.; Litinas, E.; Sisley, S.; Williams, D.A.; Clauw, D.J. Pills to Pot: Observational Analyses of Cannabis Substitution among Medical Cannabis Users with Chronic Pain. J. Pain 2019, 20, 830–841. [Google Scholar] [CrossRef]
- Guzmán, M. Cannabis for the Management of Cancer Symptoms: THC Version 2.0? Cannabis Cannabinoid Res. 2018, 3, 117–119. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Jenabian, M.A. Acute Inflammation and Pathogenesis of SARS-CoV-2 Infection: Cannabidiol as a Potential Anti-Inflammatory Treatment? Cytokine Growth Factor Rev. 2020, 53, 63–65. [Google Scholar] [CrossRef]
- Rizzo, M.D.; Henriquez, J.E.; Blevins, L.K.; Bach, A.; Crawford, R.B.; Kaminski, N.E. Targeting Cannabinoid Receptor 2 on Peripheral Leukocytes to Attenuate Inflammatory Mechanisms Implicated in HIV-Associated Neurocognitive Disorder. J. Neuroimmune Pharmacol. 2020, 15, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Tortora, C.; Argenziano, M.; Di Paola, A.; Punzo, F. Cannabinoid Receptor Type 2: A Possible Target in SARS-CoV-2 (CoV-19) Infection? Int. J. Mol. Sci. 2020, 21, 3809. [Google Scholar] [CrossRef] [PubMed]
- van Breemen, R.B.; Muchiri, R.N.; Bates, T.A.; Weinstein, J.B.; Leier, H.C.; Farley, S.; Tafesse, F.G. Cannabinoids Block Cellular Entry of SARS-CoV-2 and the Emerging Variants. J. Nat. Prod. 2022, 85, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Krug, R.G.; Clark, K.J. Elucidating Cannabinoid Biology in Zebrafish (Danio rerio). Gene 2015, 570, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Carty, D.R.; Miller, Z.S.; Thornton, C.; Pandelides, Z.; Kutchma, M.L.; Willett, K.L. Multigenerational Consequences of Early-Life Cannabinoid Exposure in Zebrafish. Toxicol. Appl. Pharmacol. 2019, 364, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Chousidis, I.; Chatzimitakos, T.; Leonardos, D.; Filiou, M.D.; Stalikas, C.D.; Leonardos, I.D. Cannabinol in the Spotlight: Toxicometabolomic Study and Behavioral Analysis of Zebrafish Embryos Exposed to the Unknown Cannabinoid. Chemosphere 2020, 252, 126417. [Google Scholar] [CrossRef] [PubMed]
- Pandelides, Z.; Thornton, C.; Lovitt, K.G.; Faruque, A.S.; Whitehead, A.P.; Willett, K.L.; Ashpole, N.M. Developmental Exposure to Δ9-Tetrahydrocannabinol (THC) Causes Biphasic Effects on Longevity, Inflammation, and Reproduction in Aged Zebrafish (Danio rerio). GeroScience 2020, 42, 923–936. [Google Scholar] [CrossRef]
- Samarut, É.; Nixon, J.; Kundap, U.P.; Drapeau, P.; Ellis, L.D. Single and Synergistic Effects of Cannabidiol and δ-9-Tetrahydrocannabinol on Zebrafish Models of Neuro-Hyperactivity. Front. Pharmacol. 2019, 10, 226. [Google Scholar] [CrossRef]
- Kalueff, A.V.; Stewart, A.M.; Gerlai, R. Zebrafish as an Emerging Model for Studying Complex Brain Disorders. Trends Pharmacol. Sci. 2014, 35, 63–75. [Google Scholar] [CrossRef]
- Ellis, L. Zebrafish as a High-Throughput in vivo Model for Testing the Bioactivity of Cannabinoids. In Recent Advances in Cannabinoid Research; IntechOpen: London, UK, 2019. [Google Scholar]
- Henn, K.; Braunbeck, T. Dechorionation as a tool to improve the fish embryo toxicity test (FET) with the zebrafish (Danio rerio). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 91–98. [Google Scholar] [CrossRef]
- OECD OECD GUIDELINES FOR THE TESTING OF CHEMICALS nr 236: Fish Embryo Acute Toxicity (FET) Test. OECD Guidel. Test. Chem. Sect. 2, OECD Publ. 2013. Available online: https://doi.org/10.1787/9789264203709-en (accessed on 4 July 2022).
- Akhtar, M.T.; Ali, S.; Rashidi, H.; Van Der Kooy, F.; Verpoorte, R.; Richardson, M.K. Developmental Effects of Cannabinoids on Zebrafish Larvae. Zebrafish 2013, 10, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.J. The Toxicologic and Teratologic Effects of Δ9-Tetrahydrocannabinol in the Zebrafish Embryo. Toxicol. Appl. Pharmacol. 1975, 32, 184–190. [Google Scholar] [CrossRef]
- Chen, J.N.; Haffter, P.; Odenthal, J.; Vogelsang, E.; Brand, M.; Van Eeden, F.J.M.; Furutani-Seiki, M.; Granato, M.; Hammerschmidt, M.; Heisenberg, C.P.; et al. Mutations Affecting the Cardiovascular System and Other Internal Organs in Zebrafish. Development 1996, 123, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Fumagalli, M.; Lecca, D.; Abbracchio, M.P.; Ceruti, S. Pathophysiological Role of Purines and Pyrimidines in Neurodevelopment: Unveiling New Pharmacological Approaches to Congenital Brain Diseases. Front. Pharmacol. 2017, 8, 941. [Google Scholar] [CrossRef]
- Lee, H.K.; Kim, K.; Lee, J.; Lee, J.; Lee, J.; Kim, S.; Lee, S.E.; Kim, J.H. Targeted Toxicometabolomics of Endosulfan Sulfate in Adult Zebrafish (Danio rerio) Using GC-MS/MS in Multiple Reaction Monitoring Mode; Elsevier B.V.: Amsterdam, The Netherlands, 2020; Volume 389, ISBN 8228804644. [Google Scholar]
- Wang, D.; Zhang, Y.; Li, J.; Dahlgren, R.A.; Wang, X.; Huang, H.; Wang, H. Risk Assessment of Cardiotoxicity to Zebrafish (Danio rerio) by Environmental Exposure to Triclosan and Its Derivatives. Environ. Pollut. 2020, 265, 114995. [Google Scholar] [CrossRef]
- Nyhan, W.L. Disorders of Purine and Pyrimidine Metabolism. Mol. Genet. Metab. 2005, 86, 25–33. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, L.; Han, C.; Lv, H.; Chen, D.; Shen, G.; Wu, K.; Pan, S.; Ye, F. Design, Synthesis, and Biological Activity of Tetrahydrobenzo[4,5]Thieno[2,3-d]Pyrimidine Derivatives as Anti-Inflammatory Agents. Molecules 2017, 22, 1960. [Google Scholar] [CrossRef]
- Hartsel, J.A.; Eades, J.; Hickory, B.; Makriyannis, A. Cannabis Sativa and Hemp. In Nutraceuticals: Efficacy, Safety and Toxicity; Academic Press: New York, NY, USA, 2016; pp. 735–754. ISBN 9780128021477. [Google Scholar]
- Maximino, C.; de Brito, T.M.; da Silva Batista, A.W.; Herculano, A.M.; Morato, S.; Gouveia, A. Measuring Anxiety in Zebrafish: A Critical Review. Behav. Brain Res. 2010, 214, 157–171. [Google Scholar] [CrossRef]
- Campos, A.C.; Moreira, F.A.; Gomes, F.V.; del Bel, E.A.; Guimarães, F.S. Multiple Mechanisms Involved in the Large-Spectrum Therapeutic Potential of Cannabidiol in Psychiatric Disorders. Philos. Trans. R. Soc. Biol. Sci. 2012, 367, 3364–3378. [Google Scholar] [CrossRef]
- Casarotto, P.C.; Gomes, F.V.; Resstel, L.B.M.; Guimarães, F.S. Cannabidiol Inhibitory Effect on Marble-Burying Behaviour: Involvement of CB1 Receptors. Behav. Pharmacol. 2010, 21, 353–358. [Google Scholar] [CrossRef]
- Moreira, F.A.; Guimarães, F.S. Cannabidiol Inhibits the Hyperlocomotion Induced by Psychotomimetic Drugs in Mice. Eur. J. Pharmacol. 2005, 512, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Viveros, M.P.; Marco, E.M.; File, S.E. Endocannabinoid System and Stress and Anxiety Responses. Pharmacol. Biochem. Behav. 2005, 81, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.J.; Byun, D.; Nam, T.S.; Choi, S.Y.; Lee, B.G.; Kim, M.K.; Kim, S. Zebrafish as an Animal Model in Epilepsy Studies with Multichannel EEG Recordings. Sci. Rep. 2017, 7, 3099. [Google Scholar] [CrossRef] [PubMed]
- Donvito, G.; Nass, S.R.; Wilkerson, J.L.; Curry, Z.A.; Schurman, L.D.; Kinsey, S.G.; Lichtman, A.H. The Endogenous Cannabinoid System: A Budding Source of Targets for Treating Inflammatory and Neuropathic Pain. Neuropsychopharmacology 2018, 43, 52–79. [Google Scholar] [CrossRef]
- Khatri, D.; Zizioli, D.; Tiso, N.; Facchinello, N.; Vezzoli, S.; Gianoncelli, A.; Memo, M.; Monti, E.; Borsani, G.; Finazzi, D. Down-Regulation of Coasy, the Gene Associated with NBIA-VI, Reduces Bmp Signaling, Perturbs Dorso-Ventral Patterning and Alters Neuronal Development in Zebrafish. Sci. Rep. 2016, 6, 37660. [Google Scholar] [CrossRef]
- Pacheco-Alvarez, D.; Solórzano-Vargas, R.S.; Del Río, A.L. Biotin in Metabolism and Its Relationship to Human Disease. Arch. Med. Res. 2002, 33, 439–447. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, D.Q.; Zhang, P.W.; Chen, X.T.; Liu, B.J.; Cheng, D.M.; Zhang, Z.X. Integrated LC–MS and GC–MS-Based Untargeted Metabolomics Studies of the Effect of Azadirachtin on Bactrocera Dorsalis Larvae. Sci. Rep. 2020, 10, 2306. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Patterson, J.; Kimmel, R.O. The Development and Behavioral Characteristics of the Startle Response in the Zebra Fish. Dev. Psychobiol. 1974, 7, 47–60. [Google Scholar] [CrossRef]
- Flores-Valverde, A.M.; Horwood, J.; Hill, E.M. Disruption of the Steroid Metabolome in Fish Caused by Exposure to the Environmental Estrogen 17α-Ethinylestradiol. Environ. Sci. Technol. 2010, 44, 3552–3558. [Google Scholar] [CrossRef]
- Gu, J.; Hu, X.; Shao, W.; Ji, T.; Yang, W.; Zhuo, H.; Jin, Z.; Huang, H.; Chen, J.; Huang, C.; et al. Metabolomic Analysis Reveals Altered Metabolic Pathways in a Rat Model of Gastric Carcinogenesis. Oncotarget 2016, 7, 60053–60073. [Google Scholar] [CrossRef] [Green Version]
- Elie, M.R.; Choi, J.; Nkrumah-Elie, Y.M.; Gonnerman, G.D.; Stevens, J.F.; Tanguay, R.L. Metabolomic Analysis to Define and Compare the Effects of PAHs and Oxygenated PAHs in Developing Zebrafish. Environ. Res. 2015, 140, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of Embryonic Development of the Zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolic Pathway | Vehicle Control | THC 1.00 mg/L | THC 1.125 mg/L | THC 1.25 mg/L |
|---|---|---|---|---|
| Amino sugar and nucleotide sugar metabolism | ✓ | ✓ | ✓ | ✓ |
| Cysteine and methionine metabolism | ✓ | ✓ | ✓ | ✓ |
| Riboflavin metabolism | ✓ | ✓ | ✓ | ✓ |
| Starch and sucrose metabolism | ✓ | ✓ | ✓ | ✓ |
| Galactose metabolism | ✓ | ✓ | ✓ | ✓ |
| Neomycin, kanamycin, and gentamicin biosynthesis | ✓ | ✓ | ✓ | ✓ |
| Pentose and glucuronate interconversions | ✓ | ✓ | ✓ | |
| Pyrimidine metabolism | ✓ | ✓ | ✓ | |
| Aminoacyl-tRNA biosynthesis | ✓ | ✓ | ||
| Arginine and proline metabolism | ✓ | ✓ | ||
| D-Glutamine and D-glutamate metabolism | ✓ | ✓ | ||
| Nitrogen metabolism | ✓ | |||
| Steroid biosynthesis | ✓ | |||
| Biotin metabolism | ✓ | |||
| Pantothenate and CoA biosynthesis | ✓ | ✓ | ||
| Fatty acid degradation | ✓ | ✓ | ||
| Glutathione metabolism | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatzimitakos, T.; Chousidis, I.; Leonardos, D.; Stalikas, C.; Leonardos, I. In the Swim of Cannabis: Developmental Toxicity and Metabolomic Pathway Alterations of Zebrafish Larvae Exposed to THC for the Assessment of Its Potential Environmental and Human Health Impact. Molecules 2022, 27, 5506. https://doi.org/10.3390/molecules27175506
Chatzimitakos T, Chousidis I, Leonardos D, Stalikas C, Leonardos I. In the Swim of Cannabis: Developmental Toxicity and Metabolomic Pathway Alterations of Zebrafish Larvae Exposed to THC for the Assessment of Its Potential Environmental and Human Health Impact. Molecules. 2022; 27(17):5506. https://doi.org/10.3390/molecules27175506
Chicago/Turabian StyleChatzimitakos, Theodoros, Ieremias Chousidis, Dimitrios Leonardos, Constantine Stalikas, and Ioannis Leonardos. 2022. "In the Swim of Cannabis: Developmental Toxicity and Metabolomic Pathway Alterations of Zebrafish Larvae Exposed to THC for the Assessment of Its Potential Environmental and Human Health Impact" Molecules 27, no. 17: 5506. https://doi.org/10.3390/molecules27175506
APA StyleChatzimitakos, T., Chousidis, I., Leonardos, D., Stalikas, C., & Leonardos, I. (2022). In the Swim of Cannabis: Developmental Toxicity and Metabolomic Pathway Alterations of Zebrafish Larvae Exposed to THC for the Assessment of Its Potential Environmental and Human Health Impact. Molecules, 27(17), 5506. https://doi.org/10.3390/molecules27175506