
Chemical Profiling and Cholinesterase Inhibitory Activity of Five Phaedranassa Herb. (Amaryllidaceae) Species from Ecuador

 ,
, 
 ,
,  and
and
Abstract

1. Introduction
2. Results and Discussion
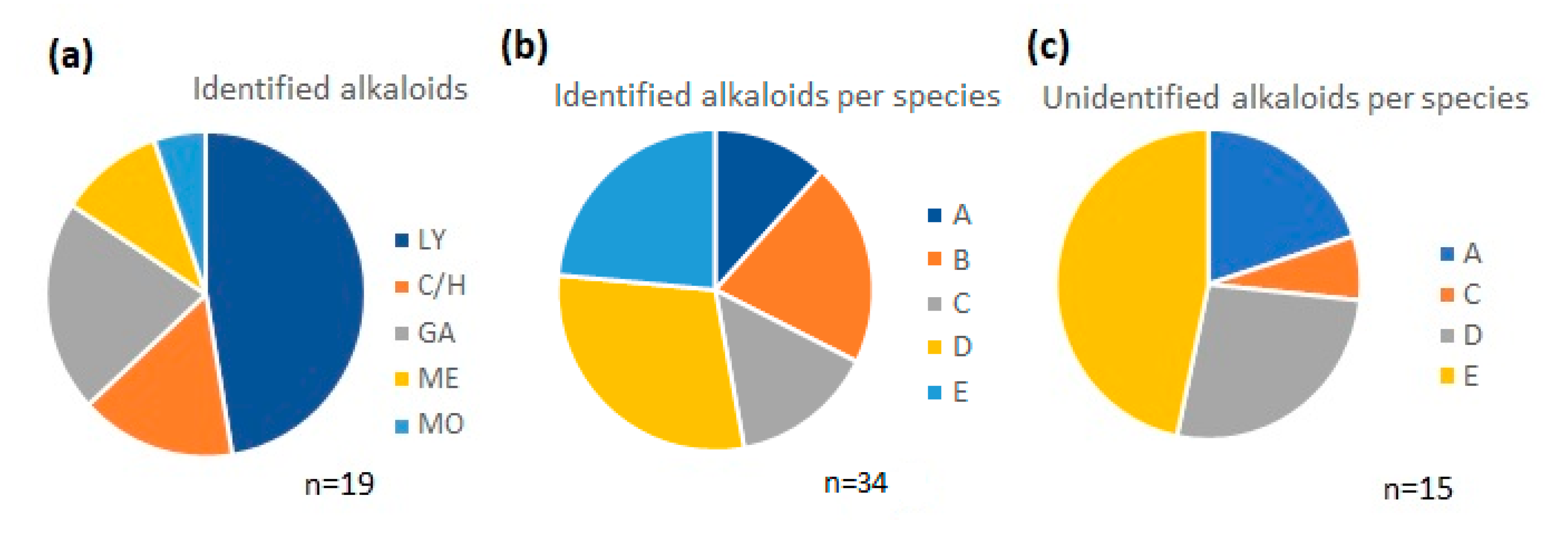
2.1. Alkaloids Identified by GC-MS
2.2. AChE and BuChE Inhibitory Activities
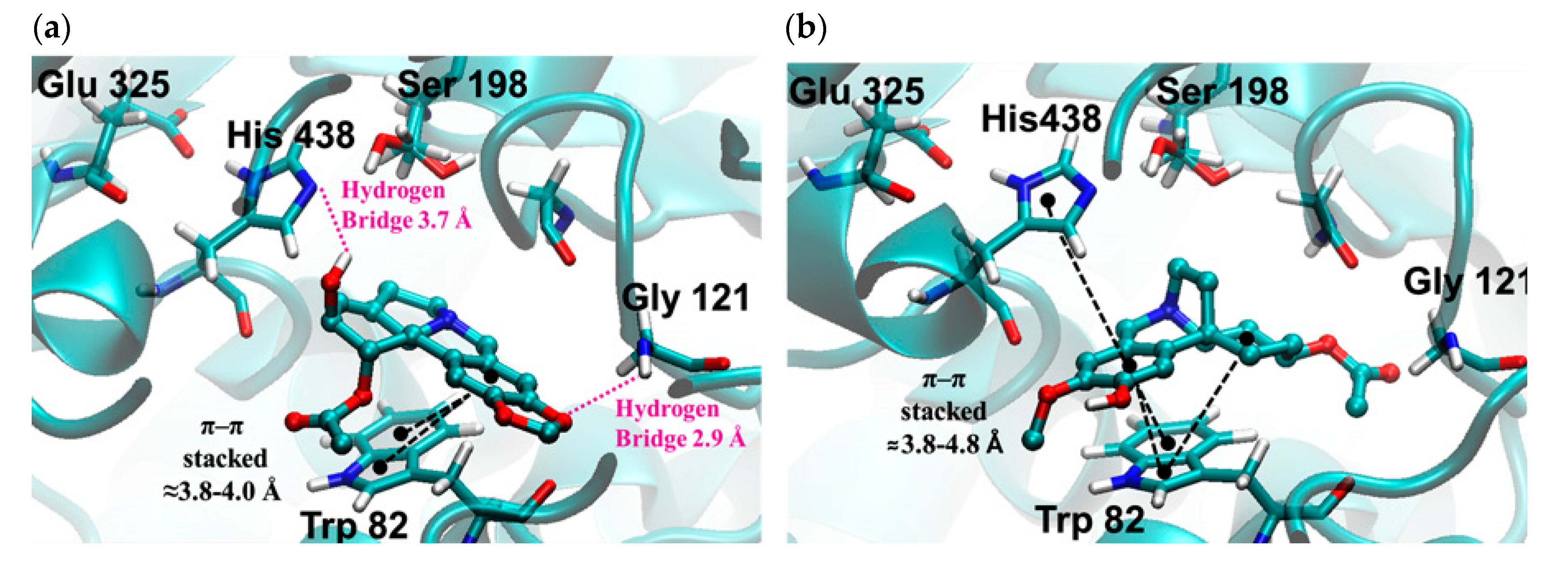
2.3. Molecular Docking
3. Materials and Methods
3.1. Plant Material
3.2. Extractions
3.3. Acetylcholinesterase (AChE) and Butyrylcholinesterase (BuChE) Inhibitory Activity
3.4. Alkaloid Identification and Quantification
3.5. Statistical Analysis
3.6. Molecular Docking
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A. Strategies for discovering drugs from previously unexplored natural products. Drug Discov. Today 2000, 5, 294–300. [Google Scholar] [CrossRef]
- Rodrigues, T.; Reker, D.; Schneider, P.; Schneider, G. Counting on natural products for drug design. Nat. Chem. 2016, 8, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Stratton, C.F.; Newman, D.J.; Tan, D.S. Cheminformatic comparison of approved drugs from natural products versus synthetic origins. Bioorg. Med. Chem. Lett. 2015, 25, 4802–4807. [Google Scholar] [CrossRef]
- Bastida, J.; Lavilla, R.; Viladomat, F. Chemical and biological aspects of Narcissus Alkaloids. In The Alkaloids: Chemistry and Physiology; Cordell, G.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 63, pp. 87–179. [Google Scholar] [CrossRef]
- Minga, D.; Ulloa, C.U.; Oleas, N.; Verdugo, A. A new species of Phaedranassa (Amaryllidaceae) from Ecuador. Phytotaxa 2015, 192, 50–53. [Google Scholar] [CrossRef]
- Oleas, N. Landscape Genetics of Phaedranassa Herb. (Amaryllidaceae) in Ecuador. Ph.D. Thesis, Florida International University, Miami, FL, USA, 2011. [Google Scholar]
- Oleas, N. Amaryllidaceae. In Libro Rojo de Plantas Endémicas del Ecuador; León-Yánez, S., Valencia, R., Pitman, N., Endara, L., Ulloa Ulloa, C., Navarrete, H., Eds.; Publicaciones del Herbario QCA, Pontificia Universidad Católica del Ecuador: Quito, Ecuador, 2011. [Google Scholar]
- Oleas, N.H.; Meerow, A.W.; Francisco-Ortega, J. Population dynamics of the endangered plant, Phaedranassa tunguraguae, from the Tropical Andean Hotspot. J. Hered 2012, 103, 557–569. [Google Scholar] [CrossRef][Green Version]
- Oleas, N.H.; Meerow, A.W.; Francisco-Ortega, J. Genetic structure of the threatened Phaedranassa schizantha (Amaryllidaceae). Bot. J. Linn. Soc. 2016, 182, 169–179. [Google Scholar] [CrossRef]
- Alzheimer’s Disease International. 2018. Available online: https://www.alz.co.uk/research/WorldAlzheimerReport2018.pdf (accessed on 30 January 2020).
- Selkoe, D.J. Alzheimer’s Disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef]
- Konrath, E.L.; dos Santos Passos, C.; Klein-Júnior, L.C.; Henriques, A.T. Alkaloids as a source of potential anticholinesterase inhibitors for the treatment of Alzheimer’s disease. J. Pharm. Pharm. 2013, 65, 1701–1725. [Google Scholar] [CrossRef]
- Maelicke, A.; Samachocki, M.; Jostock, R.; Fehrenbacher, A.; Ludwig, J.; Albuquerque, E.X.; Zerlin, M. Allosteric sensitization of nicotine receptors by galanthamine, a new treatment strategy for Alzheimer’s disease. Biol. Psychiatry 2001, 49, 279–288. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Tallini, L.; Viladomat, F.; Bastida, J. Research in natural products: Amaryllidaceae ornamental plants as sources of bioactive compounds. In Recent Advances in Pharmaceutical Sciences VII; Muñoz-Torrero, D., Riu, M., Feliu, C., Eds.; Research Signpost Trivandrum: Kerala, India, 2017; pp. 69–82. ISBN 978-81-308-0573-3. [Google Scholar]
- Guo, Y.; Pigni, N.B.; Zheng, Y.; de Andrade, J.P.; Torras-Claveria, L.; Borges, W.S.; Viladomat, F.; Codina, C.; Bastida, J. Analysis of bioactive Amaryllidaceae alkaloid profiles in Lycoris species by GC-MS. Nat. Prod. Commun. 2014, 9, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Tallini, L.R.; Bastida, J.; Cortes, N.; Osorio, E.H.; Theoduloz, C.; Schemeda-Hirschmann, G. Cholinesterase inhibition activity, alkaloid profiling and molecular docking of Chilean Rhodophiala (Amaryllidaceae). Molecules 2018, 23, 1532. [Google Scholar] [CrossRef] [PubMed]
- Osorio, E.J.; Berkov, S.; Brun, R.; Codina, C.; Viladomat, F.; Cabezas, F.; Bastida, J. In vitro antiprotozoal activity of alkaloids from Phaedranassa dubia (Amaryllidaceae). Phytochem. Lett. 2010, 3, 161–163. [Google Scholar] [CrossRef]
- Acosta, K.; Pigni, N.; Oleas, N.; Bastida, J. Identification of the alkaloids of Stenomesson aurantiacum (Kunth) Herb., an Amaryllidaceae species from the Ecuadorian Andes. Pharmacol. Online 2014, 3, 178–183. [Google Scholar]
- Berkov, S.; Georgieva, L.; Kondakova, V.; Atanassov, A.; Viladomat, F.; Bastida, J.; Codina, C. Plant sources of galanthamine: Phytochemical and biotechnological aspects. Biotechnol. Biotechnol. Equip. 2009, 23, 1170–1176. [Google Scholar] [CrossRef]
- Bores, G.M.; Huger, F.P.; Petko, W.; Mutlib, A.E.; Camacho, F.; Rush, D.K.; Selk, D.E.; Wolf, V.; Kosley, R.W., Jr.; Davis, L.; et al. Pharmaceutical evaluation of novel Alzheimer’s disease therapeutics: Acetylcholinesterase inhibitors related to galanthamine. J. Pharm. Exp. 1996, 277, 728–738. [Google Scholar]
- Iannello, C.; Pigni, N.B.; Antognoni, F.; Poli, F.; Maxia, A.; de Andrade, J.P.; Bastida, J. A potent acetylcholinesterase inhibitor from Pancratium illyricum L. Fitoterapia 2014, 92, 163–167. [Google Scholar] [CrossRef]
- Sussman, J.L.; Harel, M.; Frolow, F.; Oefner, C.; Goldman, A.; Toker, L.; Silman, I. Atomic structure of acetylcholinesterase from Torpedo californica: A prototypic acetylcholine-binding protein. Science 1991, 253, 872–879. [Google Scholar] [CrossRef]
- Nicolet, Y. Crystal structure of human butyrylcholinesterase and of its complexes with substrate and products. J. Biol. Chem. 2003, 278, 41141–41147. [Google Scholar] [CrossRef]
- Cortes, N.; Sierra, K.; Alzate, F.; Osorio, E.H.; Osorio, E. Alkaloids of Amaryllidaceae as inhibitors of cholinesterases (AChEs and BuChEs): An integrated bioguided study. Phytochem. Anal. 2018, 29, 217–227. [Google Scholar] [CrossRef]
- Tallini, L.R.; Osorio, E.H.; dos Santos, V.D.; De Souza Borges, W.; Kaiser, M.; Viladomat, F.; Zuanazzi, J.A.S.; Bastida, J. Hippeastrum reticulatum (Amaryllidaceae): Alkaloid, profiling, biological activities and molecular docking. Molecules 2017, 22, 2191. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres Jr., V.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- López, S.; Bastida, J.; Viladomat, F.; Codina, C. Acetylcholinesterase inhibitory activity of some Amaryllidaceae alkaloids and Narcissus extracts. Life Sci. 2002, 71, 2521–2529. [Google Scholar] [CrossRef]
- Torras-Claveria, L.; Berkov, S.; Codina, C.; Viladomat, F.; Bastida, J. Metabolomic analysis of bioactive Amaryllidaceae alkaloids of ornamental varieties of Narcissus by GC-MS combined with k-means cluster analysis. Ind. Crop. Prod. 2014, 56, 211–222. [Google Scholar] [CrossRef]
- Greenblatt, H.M.; Kryger, G.; Lewis, T.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with (-)-galanthamine at 2.3 Å resolution. Febs Lett. 1999, 463, 321–326. [Google Scholar] [CrossRef]
- Nachon, F.; Carletti, E.; Ronco, C.; Trovaslet, M.; Nicolet, Y.; Jean, L.; Renard, P.-Y. Crystal structures of human cholinesterases in complex with huprine W and tacrine: Elements of specificity for anti-Alzheimer’s drugs targeting acetyl- and butyryl-cholinesterase. Biochem. J. 2013, 453, 393–399. [Google Scholar] [CrossRef]
- Adamo, C.; Barone, V. Toward reliable density functional methods without adjustable parameters: The PBE0 model. J. Chem. Phys. 1999, 110, 6158–6170. [Google Scholar] [CrossRef]
- Ernzerhof, M.; Scuseria, G.E. Assessment of the Perdew–Burke– Ernzerhof exchange-correlation functional. J. Chem. Phys. 1999, 110, 5029–5036. [Google Scholar] [CrossRef]
- Petersson, G.A.; Bennett, A.; Tensfeldt, T.G.; Al-Laham, M.A.; Shirley, W.A.; Mantzaris, J.; Mantzaris, J. A complete basis set model chemistry. I. The total energies of closed-shell atoms and hydrides of the first-row elements. J. Chem. Phys. 1988, 89, 2193–2218. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09, Revis. E.01; Gaussian, Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Moris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef]
- Thomsen, R.; Christensen, M.H. MolDock: A new technique for high-accuracy molecular docking. J. Med. Chem. 2006, 49, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alkaloid | [M+] | B.P. | R.I. | A *a | B *b | C *c | D *d | E *e |
|---|---|---|---|---|---|---|---|---|
| Lycorine-type | - | 121.09 | 85.21 | 114.18 | 85.34 | |||
| Anhydrolycorine (1) | 251 | 250 | 2534.4 | - | 2.39 | 3.52 | 2.69 | - |
| 1-O-Acetylcaranine (2) | 313 | 252 | 2554.2 | - | - | - | 2.32 | - |
| 11,12-Dehydroanhydrolycorine (3) | 249 | 248 | 2638.6 | - | 2.39 | 3.43 | 2.80 | - |
| Galanthine (4) | 317 | 242 | 2730.1 | - | - | - | - | 5.36 |
| 1-O-Acetyllycorine (5) | 329 | 226 | 2747.2 | - | 2.59 | - | 2.35 | - |
| Dihydrolycorine (6) | 289 | 288 | 2781.5 | - | - | - | - | 11.80 |
| Lycorine (7) | 287 | 226 | 2795.8 | - | 113.72 | 78.26 | 93.92 | 57.04 |
| Sternbergine (8) | 331 | 228 | 2831.4 | - | - | - | 10.10 | 8.64 |
| 2-Methoxypratosine (9) | 309 | 309 | 3084.0 | - | - | - | - | 2.50 |
| Crinine/haemanthamine-type | - | - | - | 6.34 | 7.70 | |||
| 8-O-Demethylmaritidine (10) | 273 | 273 | 2532.2 | - | - | - | - | 3.81 |
| Cantabricine (11) | 317 | 317 | 2639.9 | - | - | - | - | 3.89 |
| Haemanthamine (12) | 301 | 272 | 2668.8 | - | - | - | 6.34 | - |
| Galanthamine-type | 27.87 | 70.10 | 15.80 | 2.30 | 2.33 | |||
| Galanthamine (13) | 287 | 286 | 2385.2 | 15.99 | 48.60 | 7.16 | 2.30 | 2.33 |
| Sanguinine (14) | 273 | 273 | 2403.1 | - | 18.85 | - | - | - |
| N-Demethylgalanthamine (15) | 273 | 272 | 2428.3 | 5.51 | 2.65 | 8.64 | - | - |
| N-Formylnorgalanthamine (16) | 301 | 301 | 2834.4 | 6.37 | - | - | - | - |
| Mesembranone-type | - | - | - | 5.15 | - | |||
| 6-Epimesembranol (17) | 291 | 290 | 2379.7 | - | - | - | 2.90 | - |
| Mesembrine (18) | 289 | 218 | 2396.5 | - | - | - | 2.25 | - |
| Montanine-type | 6.17 | - | - | - | - | |||
| Pancratinine C (19) | 287 | 176 | 2622.2 | 6.17 | - | - | - | - |
| Unidentified | 17.22 | - | 2.40 | 11.76 | 27.54 | |||
| Unknown (20) (homolycorine type) *g | 301 *f | 109 | 2500.8 | - | - | - | - | 2.62 |
| Unknown (21) | 371 *f | 130 | 2513.4 | 5.76 | - | - | - | - |
| Unknown (22) (lycorine type) *g | 315 *f | 228 | 2544.5 | - | - | - | 3.23 | - |
| Unknown (23) | 239 *f | 239 | 2548.8 | 5.86 | - | - | - | - |
| Unknown (24) (crinine/haem. type) *g | 275 *f | 275 | 2574.4 | - | - | - | - | 8.87 |
| Unknown (25) (lycorine type) *g | 265 *f | 264 | 2707.4 | 5.60 | - | - | - | 3.00 |
| Unknown (26) (lycorine type) *g | 331 *f | 228 | 2738.0 | - | - | - | 3.37 | - |
| Unknown (27) (lycorine type) *g | 303 *f | 228 | 2771.7 | - | - | - | - | 3.32 |
| Unknown (28) (lycorine type) *g | 315 *f | 240 | 2805.7 | - | - | - | - | 2.26 |
| Unknown (29) (lycorine type) *g | 329 *f | 268 | 2873.2 | - | - | 2.40 | - | - |
| Unknown (30) (lycorine type) *g | 289 *f | 228 | 2873.9 | - | - | - | 2.81 | - |
| Unknown (31) (homolycorine type) *g | 331 *f | 125 | 2928.3 | - | - | - | 2.35 | - |
| Unknown (32) (lycorine type) *g | 374 *f | 284 | 2953.7 | - | - | - | - | 3.04 |
| Unknown (33) (lycorine type) *g | 295 *f | 294 | 2963.1 | - | - | - | - | 4.43 |
| Total | 51.26 | 191.19 | 103.41 | 139.73 | 122.91 |
| Alkaloid | AChE | BuChE | Reference |
|---|---|---|---|
| Lycorine-Type | |||
| Anhydrolycorine (1) | −8.38 a; −8.35 c | −8.14 b | [25] |
| 1-O-Acetylcaranine (2) | −9.55 a | −8.78 b | Calculated values |
| 11,12-Dehydroanhydrolycorine (3) | −8.41 a | −7.44 b | [17] |
| Galanthine (4) | −8.43 a | −8.37 b | Calculated values |
| 1-O-Acetyllycorine (5) | −8.82 a | −9.02 b | Calculated values |
| Dihydrolycorine (6) | −8.76 a; −9.07 c | −8.80 b | [25] |
| Lycorine (7) | −8.82 a | −8.94 b | [17] |
| Sternbergine (8) | −8.61 a | −8.71 b | Calculated values |
| 2-Methoxypratosine (9) | −8.14 a | −7.81 b | Calculated values |
| Homolycorine-type | |||
| Crinine/haemanthamine-type | |||
| 8-O-Demethylmaritidine (10) | −8.74 a | −8.93 b | [17] |
| Cantabricine (11) | −9.15 a | −9.07 b | Calculated values |
| Haemanthamine (12) | −8.80 a | −8.34 b | [17] |
| Galanthamine-type | |||
| Galanthamine (13) | −10.10 a | −8.23 b | Calculated values |
| Sanguinine (14) | −9.40 a | −7.92 b | [25] |
| N-Demethylgalanthamine (15) | −9.09 a | −8.81 b | Calculated values |
| N-Formylnorgalanthamine (16) | −8.72 a | −8.12 b | Calculated values |
| Mesembranone-type | |||
| 6-Epimesembranol (17) | −8.15 a | −7.59 b | Calculated values |
| Mesembrine (18) | −8.77 a | −8.14 b | Calculated values |
| Montanine-type | |||
| Pancratinine C (19) | −8.53 a | −8.12 b | Calculated values |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno, R.; Tallini, L.R.; Salazar, C.; Osorio, E.H.; Montero, E.; Bastida, J.; Oleas, N.H.; Acosta León, K. Chemical Profiling and Cholinesterase Inhibitory Activity of Five Phaedranassa Herb. (Amaryllidaceae) Species from Ecuador. Molecules 2020, 25, 2092. https://doi.org/10.3390/molecules25092092
Moreno R, Tallini LR, Salazar C, Osorio EH, Montero E, Bastida J, Oleas NH, Acosta León K. Chemical Profiling and Cholinesterase Inhibitory Activity of Five Phaedranassa Herb. (Amaryllidaceae) Species from Ecuador. Molecules. 2020; 25(9):2092. https://doi.org/10.3390/molecules25092092
Chicago/Turabian StyleMoreno, Raúl, Luciana R. Tallini, Cristina Salazar, Edison H. Osorio, Evelin Montero, Jaume Bastida, Nora H. Oleas, and Karen Acosta León. 2020. "Chemical Profiling and Cholinesterase Inhibitory Activity of Five Phaedranassa Herb. (Amaryllidaceae) Species from Ecuador" Molecules 25, no. 9: 2092. https://doi.org/10.3390/molecules25092092
APA StyleMoreno, R., Tallini, L. R., Salazar, C., Osorio, E. H., Montero, E., Bastida, J., Oleas, N. H., & Acosta León, K. (2020). Chemical Profiling and Cholinesterase Inhibitory Activity of Five Phaedranassa Herb. (Amaryllidaceae) Species from Ecuador. Molecules, 25(9), 2092. https://doi.org/10.3390/molecules25092092