Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells

 and
and
Abstract

1. Introduction
2. Results
2.1. Effects of nCGA and LPS on Cell Viability in A549 Cells
2.2. Effects of nCGA on Inflammatory Cytokines Production in LPS-Stimulated A549 Cells
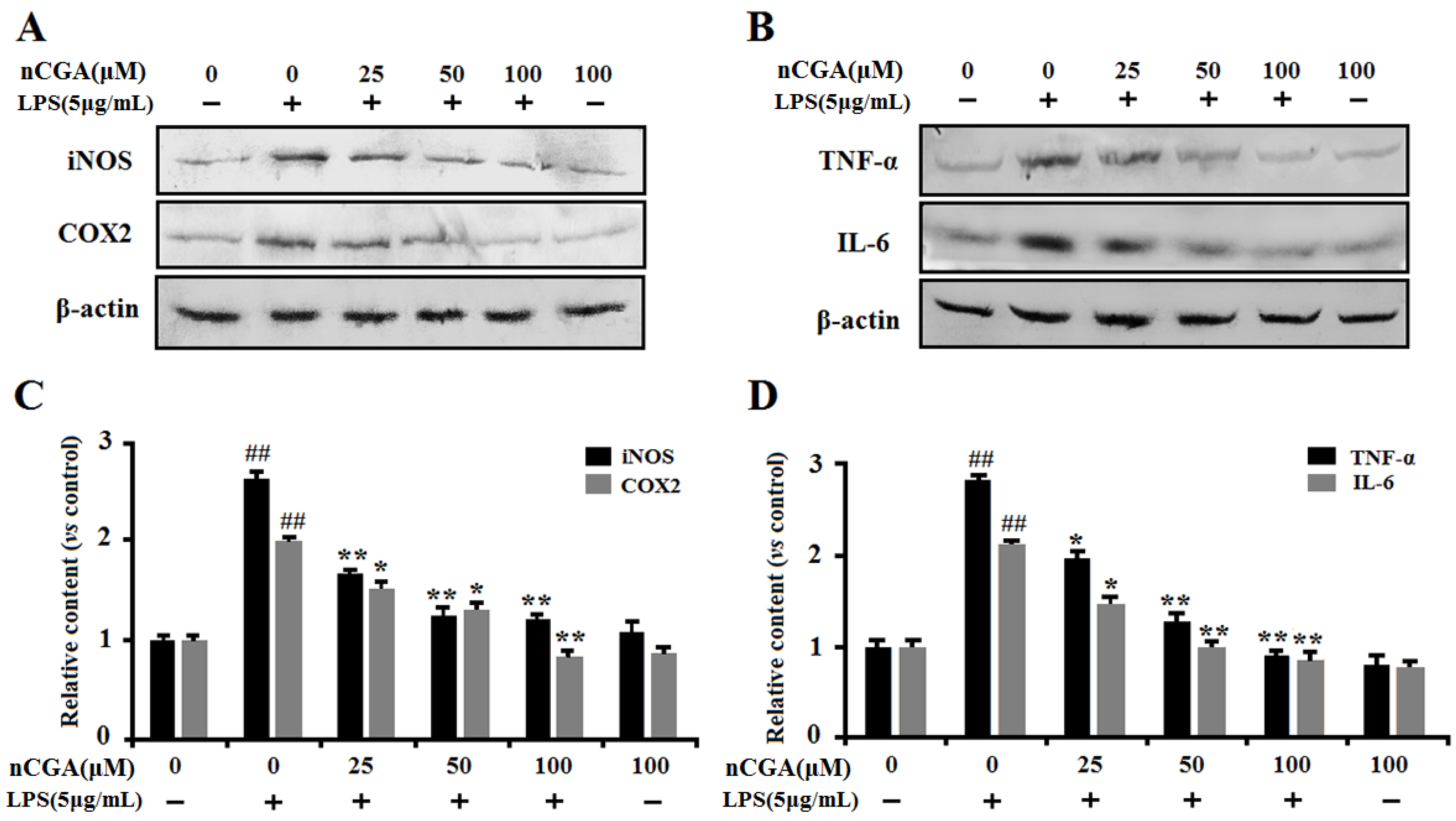
2.3. Effects of nCGA on the Expression of Inflammatory-Related Proteins in LPS-Stimulated A549 Cells
2.4. nCGA Attenuated LPS-Stimulated Inflammatory Responses via Inhibiting MAPK Phosphorylation and NF-κB Activation in A549 Cells
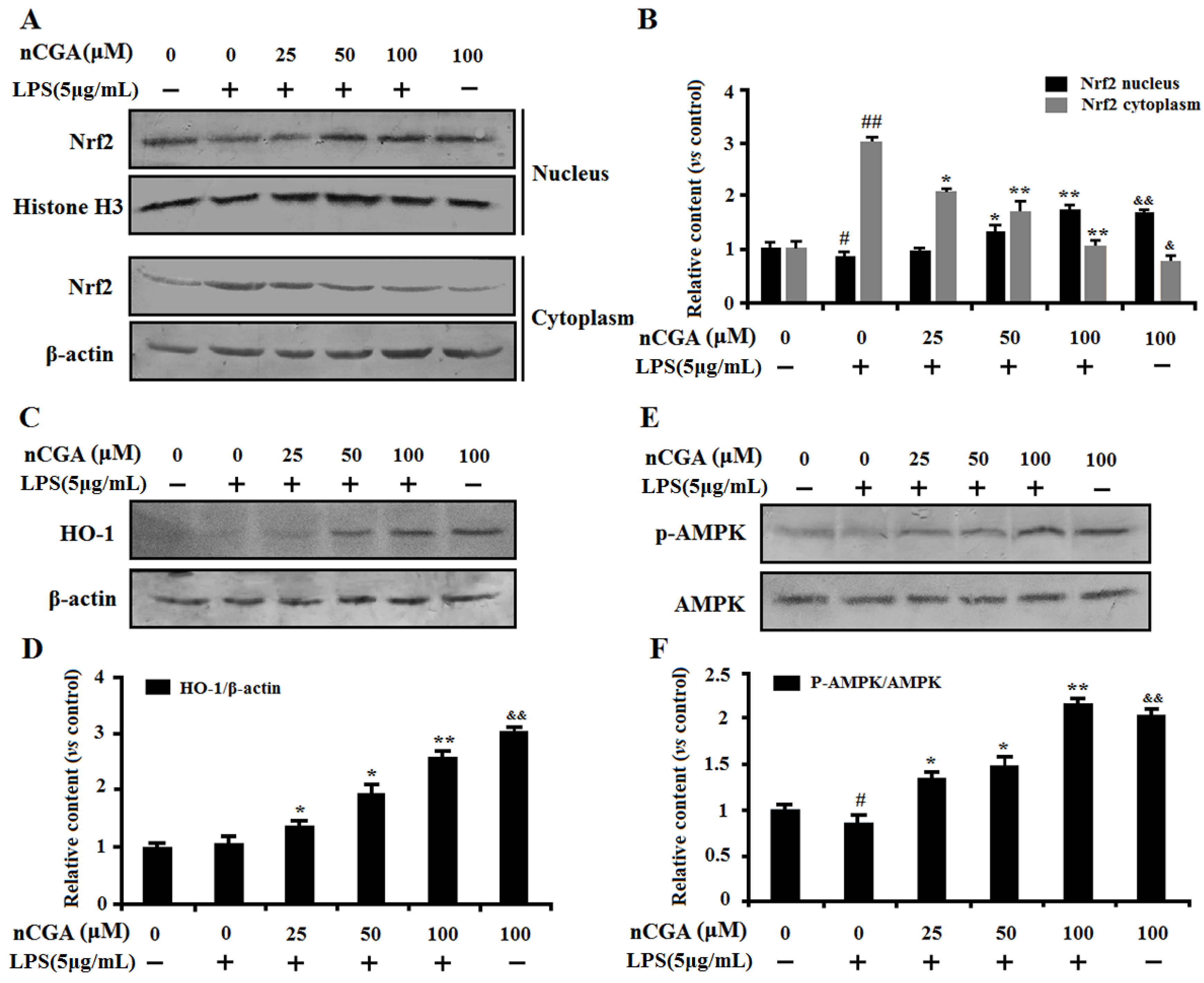
2.5. Effects of nCGA on Regulation of Nrf2 and AMPK Against LPS-Induced Inflammatory Responses in A549 Cells
2.6. The Importance of AMPK Activation in the Process of nCGA Against LPS-Induced Inflammatory Responses
3. Discussion
4. Materials and Method
4.1. Reagents and Chemicals
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. Nitrite Assay
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Quantitative Real-Time PCR
4.7. Immunofluorescence Assay
4.8. Western Blot Analysis
4.9. Preparation of Cytoplasmic and Nuclear Protein Extracts
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MTT | 3-[4–dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide |
| LPS | lipopolysaccharide |
| nCGA | neochlorogenic acid |
| AMPK | AMP-activated protein kinase |
| MAPK | mitogen-activated protein kinase |
| Nrf2 | nuclear factor-crythroid 2-related factor |
| HO-1 | heme oxygenase-1 |
| TNF-α | tumor necrosis factor α |
| IL-6 | interleukin-6 |
| iNOS | nitric oxide synthase |
| COX2 | Cyclooxygenase 2 |
| NO | nitric oxide |
| C.C | Compound C |
References
- Oliwa, J.N.; Marais, B.J. Vaccines to prevent pneumonia in children–a developing country perspective. Paediatr. Respir. Rev. 2017, 22, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-W.; Chien, M.-H.; Su, C.-S.; Yang, S.-F. New markers in pneumonia. Clin. Chim. Acta 2013, 419, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Singh, V. The burden of pneumonia in children: An asian perspective. Paediatr. Respir. Rev. 2005, 6, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Lu, C.; Deng, Q. Prenatal exposure to diurnal temperature variation and early childhood pneumonia. J. Therm. Biol. 2017, 65, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Zar, H.J. Bacterial and viral pneumonia: New insights from the drakenstein child health study. Paediatr. Respir. Rev. 2017, 24, 8–10. [Google Scholar] [CrossRef]
- Yan, S.Q.; Xiong, X.L.; Wan, Q.; Liu, F.; Tang, J.Q.; Jiang, Z.X.; Zhou, L.S.; Yuan, K.; Xie, D. Preventative effect of massage on gastric volvulus in infants with gastroesophageal reflux-induced pneumonia. J. Tradit. Chin. Med. 2015, 35, 520–527. [Google Scholar]
- Wang, P.; Qiao, Q.; Li, J.; Wang, W.; Yao, L.P.; Fu, Y.J. Inhibitory effects of geraniin on LPS-induced inflammation via regulating NF-κB and Nrf2 pathways in RAW 264.7 cells. Chem.-Biol. Interact. 2016, 253, 134–142. [Google Scholar] [CrossRef]
- Knapp, S. LPS and bacterial lung inflammation models. Drug Discov. Today: Dis. Model. 2009, 6, 113–118. [Google Scholar] [CrossRef]
- Liu, Y.; Song, M.; Zhu, G.F.; Xi., X.; Li, K.; Wu, C.T.; Huang, L.X. Corynoline attenuates LPS-induced acute lung injury in mice by activating Nrf2. Int. Immunopharmacol. 2017, 48, 96–101. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Greiffo, F.R.; Rigonato-Oliveira, N.C.; Custódio, R.W.A.; Silva, V.R.; Damaceno-Rodrigues, N.R.; Almeida, F.M.; Albertini, R.; Lopes-Martins, R.A.; Oliveira, L.V.F.; et al. Low level laser therapy reduces acute lung inflammation in a model of pulmonary and extrapulmonary LPS-induced ARDS. J. Photochem. Photobiol. B: Biol. 2014, 134, 57–63. [Google Scholar] [CrossRef]
- Silverman, N.; Maniatis, T. NF-kappa B signaling pathways in mammalian and insect innate immunity. Genes Dev. 2001, 15, 2321–2342. [Google Scholar] [CrossRef] [PubMed]
- Jane, T.J.; Xi, Q.; Jos, L.J.; Shi, B.C.; David, H.M.; Stevenson, F.; Stevenson, M.H.; Karolyn, G.L.; Robert, W.S.; James, D.N.; et al. Glutathione S-transferase pi modulates NF-κB activation and pro-inflammatory responses in lung epithelial cells. Redox Biol. 2016, 8, 375–382. [Google Scholar]
- Hoffmann, A.; Levchenko, A.; Scott, M.L.; Baltimore, D. The IκB-NF-κB signaling module: Temporal control and selective gene activation. Science 2002, 298, 1241–1245. [Google Scholar] [CrossRef]
- He, G.D.; Zhang, X.; Chen, Y.H.; Chen, J.; Li, J.; Xie, Y.B. Isoalantolactone inhibits LPS-induced inflammation via NF-κB inactivation in peritoneal macrophages and improves survival in sepsis. Biomed. Pharmacother. 2017, 90, 598–607. [Google Scholar] [CrossRef]
- Matsuzawa, A.; Saegusa, K.; Noguchi, T.; Sadamitsu, C.; Nishitoh, H.; Nagai, S.; Koyasu, S.; Matsumoto, K.; Takeda, K.; Ichijo, H. ROS-dependent activation of the TRAF6-ASK1-p38 pathway is selectively required for TLR4-mediated innate immunity. Nat. Immunol. 2005, 6, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, S.M.; El-Shal, A.S.; Abd-Allah, S.H.; Selim, A.O.; Selim, S.A.; Gouda, Z.A.; Abd El Motteleb, D.M.; Zanfaly, H.E.; EL-Assar, H.M.; Abdelazim, S. Mesenchymal stromal cell injection protects against oxidative stress in Escherichia coli–induced acute lung injury in mice. Cytotherapy 2014, 16, 764–775. [Google Scholar] [CrossRef]
- Min, L.H.; He, B.K.; Hui, L. Mitogen-activated protein kinases in hepatocellular carcinoma development. Semin. Cancer Biol. 2011, 21, 10–20. [Google Scholar] [CrossRef]
- Chepelev, N.L.; Zhang, H.Q.; Liu, H.L.; McBride, S.; Seal, A.J.; Morgan, T.E.; Finch, C.E.; Willmore, W.G.; Davies, K.J.A.; Forman, H.J. Competition of nuclear factor-erythroid 2 factors related transcription factor isoforms, Nrf1 and Nrf2, in antioxidant enzyme induction. Redox Biol. 2013, 1, 183–189. [Google Scholar] [CrossRef]
- Zhu, H.; Zhang, L.; Itoh, K.; Yamamoto, M.; Ross, D.; Trush, M.A.; Zweier, J.L.; Li, Y.B. Nrf2 controls bone marrow stromal cell susceptibility to oxidative and electrophilic stress. Free Radic. Biol. Med. 2006, 41, 132–143. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Guo, Z.K.; Zhan, Y.T.; Li, H.Y.; Wu, S.Y. Role of histone acetylation in activation of nuclear factor erythroid 2-related factor 2/heme oxygenase 1 pathway by manganese chloride. Toxicol. Appl. Pharmacol. 2017, 336, 94–100. [Google Scholar] [CrossRef]
- Park, E.J.; Kim, Y.M.; Park, S.W.; Kim, H.J.; Lee, J.H.; Lee, D.U.; Chang, K.C. Induction of HO-1 through p38 MAPK/Nrf2 signaling pathway by ethanol extract of Inula helenium L. reduces inflammation in LPS-activated RAW 264.7 cells and CLP-induced septic mice. Food Chem. Toxicol. 2013, 55, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Mo, C.; Wang, L.; Zhang, J.; Numazawa, S.; Tang, H.; Tang, X.; Han, X.; Li, J.; Yang, M.; Wang, Z.; et al. The crosstalk between Nrf2 and AMPK signal pathways is important for the anti-inflammatory effect of Berberine in LPS-stimulated macrophages and endotoxin-shocked mice. Antioxid. Redox Signal. 2014, 20, 574–588. [Google Scholar] [CrossRef] [PubMed]
- Colombo, S.L.; Moncada, S. AMPKα1 regulates the antioxidant status of vascular endothelial cells. Biochem. J. 2009, 421, 163–169. [Google Scholar] [CrossRef]
- Lv, H.; Liu, Q.; Wen, Z.; Feng, H.; Deng, X.; Ci, X. Xanthohumol ameliorates lipopolysaccharide (LPS)-induced acute lung injury via induction of AMPK/GSK3β-Nrf2 signal axis. Redox Biology. 2017, 12, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salcedo, E.M.; Mena, P.; García-Viguera, C.; Hernández, F.; Martínez, J.J. (Poly)phenolic compounds and antioxidant activity of white (Morus alba) and black (Morus nigra) mulberry leaves: Their potential for new products rich in phytochemicals. J. Funct. Foods 2015, 18, 1039–1046. [Google Scholar] [CrossRef]
- Gryn-Rynko, A.; Bazylak, G.; Olszewska-Slonina, D. New potential phytotherapeutics obtained from white mulberry (Morus alba L.) leaves. Biomed. Pharmacother. 2016, 84, 628–636. [Google Scholar] [CrossRef]
- CHAN, E.W.C.; LYE, P.Y.; Wong, S.K. Phytochemistry, pharmacology, and clinical trials of Morus alba. Chin. J. Nat. Med. 2016, 14, 17–30. [Google Scholar]
- He, X.; Fang, J.; Ruan, Y.; Wang, X.; Sun, Y.; Wu, N.; Zhao, Z.; Chang, Y.; Ning, N.; Guo, H.; et al. Structures, bioactivities and future prospective of polysaccharides from Morus alba (white mulberry): A review. Food Chem. 2018, 245, 899–910. [Google Scholar] [CrossRef]
- Jeszka-Skowron, M.; Flaczyk, E.; Flaczyk, E.; Jeszka, J.; Krejpcio, Z.; Król, E.; Buchowski, M.S. Mulberry leaf extract intake reduces hyperglycaemia in streptozotocin (STZ)-induced diabetic rats fed high-fat diet. J. Funct. Foods 2014, 8, 9–17. [Google Scholar] [CrossRef]
- Cheng, B.C.Y.; Yu, H.; Su, T.; Fu, X.Q.; Guo, H.; Li, T.; Cao, H.H.; Tse, A.K.W.; Kwan, H.Y.; Yu, Z.L. A herbal formula comprising Rosae Multiflorae Fructus and Lonicerae Japonicae Flos inhibits the production of inflammatory mediators and the IRAK-1/TAK1 and TBK1/IRF3 pathways in RAW 264.7 and THP-1 cells. J. Ethnopharmacol. 2015, 174, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Jin, M.L.; Hye, Y.E.; Yoon, K.; Geuntae, P. Neochlorogenic acid inhibits against LPS-activated inflammatory responses through up-regulation of Nrf2/HO-1 and involving AMPK pathway. Environ. Toxicol. Pharmacol. 2018, 62, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Zhu, D.Y.; Wang, C.H.; Zhang, Y.S.; Liu, F.R.; Ye, T.Q.; Chen, X.; Wei, Z.J. Simultaneous and fast separation of three chlorogenic acids and two flavonoids from bamboo leaves extracts using zirconia. Food Chem. Toxicol. 2018, 119, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Talhaoui, N.; Vezza, T.; Gómez-Caravaca, A.M.; Fernández-Gutiérrez, A.; Gálvez, J.; Segura-Carretero, A. Phenolic compounds and in vitro immunomodulatory properties of three Andalusian olive leaf extracts. J. Funct. Foods 2016, 22, 270–277. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Rosadini, C.V.; Kagan, J.C. Early innate immune responses to bacterial LPS. Curr. Opin. Immunol.. 2017, 44, 14–19. [Google Scholar] [CrossRef]
- Goodman, R.B.; Pugin, J.; Lee, J.S.; Matthay, M.A. Cytokine-mediated inflammation in acute lung injury. Cytokine Growth Factor Rev. 2003, 14, 523–535. [Google Scholar] [CrossRef]
- Song, J.; Pan, W.; Sun, Y.; Han, J.; Shi, W.; Liao, W. Aspergillus fumigatus-induced early inflammatory response in pulmonary microvascular endothelial cells: Role of p38 MAPK and inhibition by silibinin. Int. Immunopharmacol. 2017, 49, 195–202. [Google Scholar] [CrossRef]
- Yeom, M.; Kim, J.H.; Min, J.H.; Hwang, M.K.; Jung, H.S.; Sohn, Y. Xanthii fructus inhibits inflammatory responses in LPS-stimulated RAW 264.7 macrophages through suppressing NF-κB and JNK/p38 MAPK. J. Ethnopharmacol. 2015, 176, 394–401. [Google Scholar] [CrossRef]
- Jaiswal, A.K. Nrf2 signaling in coordinated activation of antioxidant gene expression. Free Radic. Biol. Med. 2004, 36, 1199–1207. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef]
- Lee, M.Y.; Lee, J.A.; Seo, C.S.; Ha, H.; Lee, H.; Son, J.K.; Shin, H.K. Anti-inflammatory activity of Angelica dahurica ethanolic extract on RAW264.7 cells via upregulation of heme oxygenase-1. Food Chem. Toxicol. 2011, 49, 1047–1055. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Qiu, X.C.; Jiang, H.; Zhi, Y.; Fu, J.F.; Liu, J. Ulinastatin inhibits the inflammation of LPS-induced acute lung injury in mice via regulation of AMPK/NF-κB pathway. Int. Immunopharmacol. 2015, 29, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Bai, A.P.; Ma, A.G.; Yong, M.; Weiss, C.R.; Ma, Y.; Guan, Q.; Bernstein, C.N.; Peng, Z. AMPK agonist downregulates innate and adaptive immune responses in TNBS-induced murine acute and relapsing colitis. Biochem. Pharmacol. 2010, 80, 1708–1717. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.T.; Gao, M.Z.; Yang, Q.; Cui, Q.; Jian, Y.; Fan, X.H.; Yao, L.P.; Fu, Y.J. An Efficient Strategy Based on Liquid–Liquid Extraction With Acid Condition and HSCCC for Rapid Enrichment and Preparative Separation of Three Caffeoylquinic Acid Isomers From Mulberry Leaves. J. Chromatogr. Sci. 2019, 57, 738–744. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer | |
|---|---|---|
| TNF-α | 5′-CCAACGGCATGGATCTCAA-3′ | 5′-TTGACGGCAGAGAGGAGGT-3′ |
| IL-6 | 5′-TCCATCCAGTTGCCTTCTTG-3′ | 5′-AAGCCTCCGACTTGTGAAGT-3′ |
| β-actin | 5′-CTCCATCCTGGCCTCGCTGT-3′ | 5′-GCTGTCACCTTCACCGTTCC-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.-h.; Zhang, S.-d.; Wang, L.-t.; Yu, L.; Zhao, X.-l.; Ni, H.-y.; Wang, Y.-q.; Wang, J.-d.; Shan, C.-h.; Fu, Y.-j. Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells. Molecules 2020, 25, 1385. https://doi.org/10.3390/molecules25061385
Gao X-h, Zhang S-d, Wang L-t, Yu L, Zhao X-l, Ni H-y, Wang Y-q, Wang J-d, Shan C-h, Fu Y-j. Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells. Molecules. 2020; 25(6):1385. https://doi.org/10.3390/molecules25061385
Chicago/Turabian StyleGao, Xiao-han, Sun-dong Zhang, Li-tao Wang, Liang Yu, Xue-lian Zhao, Hai-yan Ni, Yan-qiu Wang, Jian-dong Wang, Chun-hua Shan, and Yu-jie Fu. 2020. "Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells" Molecules 25, no. 6: 1385. https://doi.org/10.3390/molecules25061385
APA StyleGao, X.-h., Zhang, S.-d., Wang, L.-t., Yu, L., Zhao, X.-l., Ni, H.-y., Wang, Y.-q., Wang, J.-d., Shan, C.-h., & Fu, Y.-j. (2020). Anti-Inflammatory Effects of Neochlorogenic Acid Extract from Mulberry Leaf (Morus alba L.) Against LPS-Stimulated Inflammatory Response through Mediating the AMPK/Nrf2 Signaling Pathway in A549 Cells. Molecules, 25(6), 1385. https://doi.org/10.3390/molecules25061385