
In Vitro Calli Production Resulted in Different Profiles of Plant-Derived Medicinal Compounds in Phyllanthus amarus


Abstract
1. Introduction
2. Results
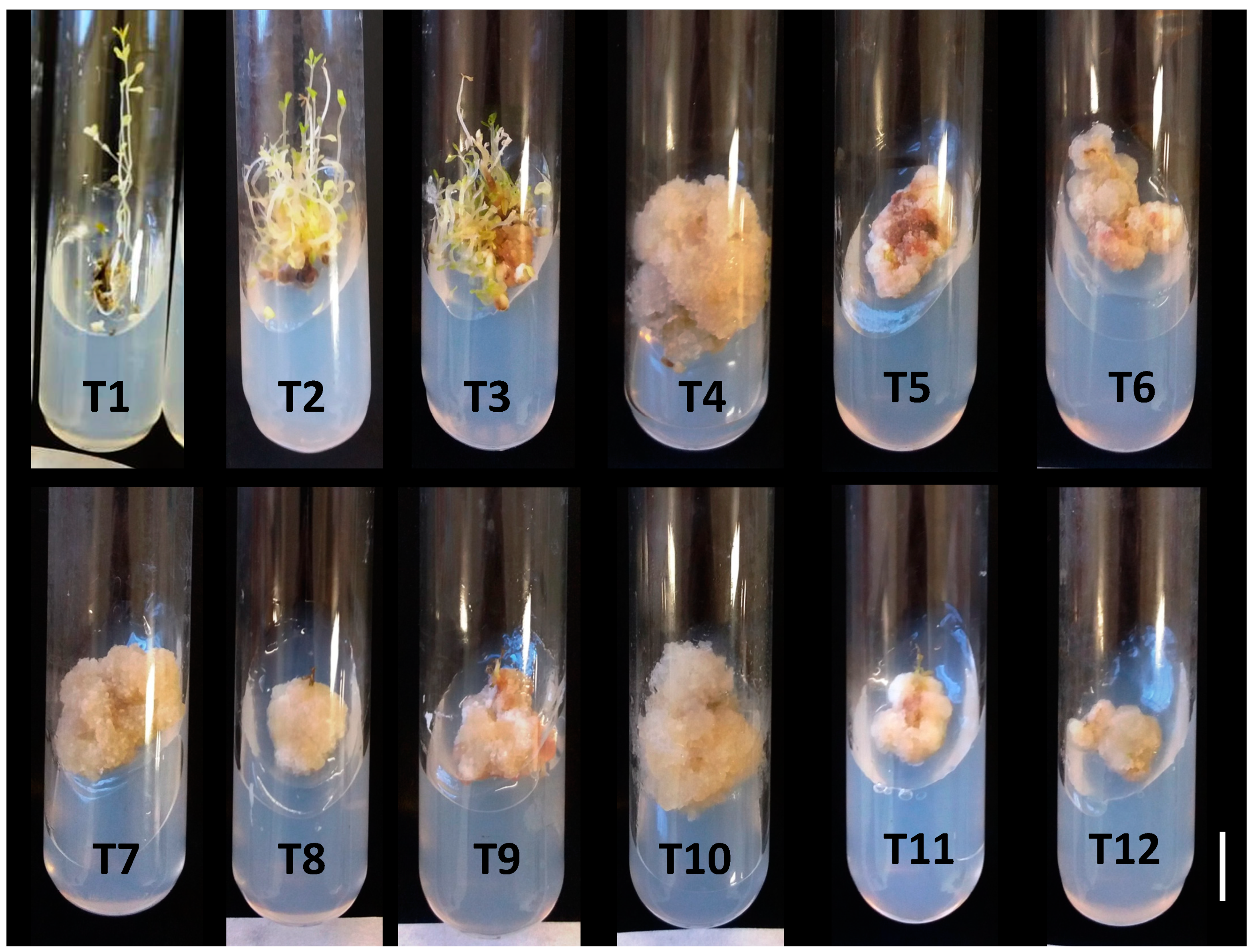
2.1. Experiments for In Vitro Callogenesis in Phyllantus amarus
2.2. Chemical Elicitation of Calli Obtained in Medium with 2,4-D in the Dark and with BA in the Light
3. Discussion
3.1. 2,4-D Was the Major Elicitor Responsible for Induction and Proliferation of Calli in Dark Conditions
3.2. The Use of Cytokinin BA, in the Presence of Light, Changed the Pattern of Callogenesis in P. amarus
3.3. The Use of Calli In Vitro Did Not Result in the Production of Phyllanthin, Even When Combined with Elicitors
3.4. In Vitro Calli Are a Source of Other Metabolites of Great Medicinal Importance
4. Material and Methods
4.1. Phyllantus amarus Genetic Material and In Vitro Seeding
4.2. Experiments for Induction and Proliferation of Calli in Nodal Segments of Phyllantus amarus
4.3. Chemical Elicitation of Calli Obtained in Medium with 2,4-D in the Dark and with BA in the Light
4.4. Cultivation of Micropropagated Plants in a Greenhouse
4.5. GC-MS Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Patel, J.R.; Tripathi, P.; Sharma, V.; Chauhan, N.S.; Dixit, V.K. Phyllanthus amarus: Ethnomedicinal uses, phytochemistry and pharmacology: A review. J. Ethnopharmacol. 2011, 138, 286–313. [Google Scholar] [CrossRef] [PubMed]
- Nisar, M.; He, J.; Ahmed, A.; Yang, Y.; Li, M.; Wan, C. Chemical Components and Biological Activities of the Genus Phyllanthus: A Review of the Recent Literature. Molecules 2018, 23, 2567. [Google Scholar] [CrossRef] [PubMed]
- Conceição, L.O.; Aoyama, E.M. Anatomia e histoquímica da lâmina foliar de espécies conhecidas por quebra-pedra (Euphorbia prostrata Aiton, Euphorbia hyssopifolia L., Phyllanthus amarus Schumach. & Thonn e Phyllanthus tenellus Roxb.). Rev. Bras. Plantas Med. 2016, 18, 571–581. [Google Scholar] [CrossRef]
- Mao, X.; Wu, L.-F.; Guo, H.-L.; Chen, W.-J.; Cui, Y.-P.; Qi, Q.; Li, S.; Liang, W.-Y.; Yang, G.-H.; Shao, Y.-Y.; et al. The Genus Phyllanthus: An Ethnopharmacological, Phytochemical, and Pharmacological Review. Evid. Based Complement. Altern. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Alamri, S.A.; Al-Whaibi, M.H.; Hussain, Z.; Ali, H.M.; El-Zaidy, M.E. A mini-review of anti-hepatitis B virus activity of medicinal plants. Biotechnol. Biotechnol. Equip. 2017, 31, 9–15. [Google Scholar] [CrossRef]
- Maia-Almeida, C.I.; Ming, L.C.; Marques, M.O.M.; Magalhães, P.M.; Queiroz, S.C.N.; Scramim, S.; Mischan, M.M.; Montanari, I., Jr.; Pereira, B.; Ferreira, M.I. Densidade de plantio e idade de colheita de quebra-pedra [Phyllanthus amarus (Schumach. & Thonning) genótipo UNICAMP-CPQBA 14] na produtividade de filantina. Rev. Bras. Plantas Med. 2011, 13, 633–641. [Google Scholar] [CrossRef]
- Gandhi, S.G.; Mahajan, V.; Bedi, Y.S. Changing trends in biotechnology of secondary metabolism in medicinal and aromatic plants. Planta 2015, 241, 303–317. [Google Scholar] [CrossRef]
- Cardoso, J.C.; de Oliveira, M.E.B.S.; de Cardoso, F.C.I. Advances and challenges on the in vitro production of secondary metabolites from medicinal plants. Hortic. Bras. 2019, 37, 124–132. [Google Scholar] [CrossRef]
- Keshvari, T.; Najaphy, A.; Kahrizi, D.; Zebarjadi, A. Callus induction and somatic embryogenesis in Stevia rebaudiana Bertoni as a medicinal plant. Cell. Mol. Biol. 2018, 64, 46–49. [Google Scholar] [CrossRef]
- Miroshnichenko, D.; Chaban, I.; Chernobrovkina, M.; Dolgov, S. Protocol for efficient regulation of in vitro morphogenesis in einkorn (Triticum monococcum L.), a recalcitrant diploid wheat species. PLoS ONE 2017, 12, e0173533. [Google Scholar] [CrossRef]
- Santos, M.R.A.; Ferreira, M.G.R.; Guimarães, M.C.M.; Lima, R.A.; Oliveira, C.L.L.G. Callogenesis in leaves of Kalanchoe pinnata Lam. by 2, 4-D and BA action. Rev. Bras. Plantas Med. 2014, 16, 760–764. [Google Scholar] [CrossRef][Green Version]
- Cooper, W.R.; Horton, D.R. Elicitors of Host Plant Defenses Partially Suppress Cacopsylla pyricola (Hemiptera: Psyllidae) Populations Under Field Conditions. J. Insect Sci. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Coste, A.; Vlase, L.; Halmagyi, A.; Deliu, C.; Coldea, G. Effects of plant growth regulators and elicitors on production of secondary metabolites in shoot cultures of Hypericum hirsutum and Hypericum maculatum. Plant Cell. Tissue Organ Cult. 2011, 106, 279–288. [Google Scholar] [CrossRef]
- Zhao, J.L.; Zhou, L.G.; Wu, J.Y. Effects of biotic and abiotic elicitors on cell growth and tanshinone accumulation in Salvia miltiorrhiza cell cultures. Appl. Microbiol. Biotechnol. 2010, 87, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Muthusamy, A.; Prasad, H.N.N.; Sanjay, E.R.; Rao, M.R.; Satyamoorthy, K. Impact of precursors and plant growth regulators on in vitro growth, bioactive lignans, and antioxidant content of Phyllanthus species. Vitr. Cell. Dev. Biol. Plant 2016, 52, 598–607. [Google Scholar] [CrossRef]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell. Tissue Organ Cult. 2018, 132, 239–265. [Google Scholar] [CrossRef]
- Nikolaeva, T.N.; Zagoskina, N.V.; Zaprometov, M.N. Production of phenolic compounds in callus cultures of tea plant under the effect of 2, 4-D and NAA. Russ. J. Plant Physiol. 2009, 56, 45–49. [Google Scholar] [CrossRef]
- Unander, D.W. Callus induction in Phyllanthus species and inhibition of viral DNA polymerase and reverse transcriptase by callus extracts. Plant Cell Rep. 1991, 10, 461–466. [Google Scholar] [CrossRef]
- Lee, N.Y.S.; Khoo, W.K.S.; Adnan, M.A.; Mahalingam, T.P.; Fernandez, A.R.; Jeevaratnam, K. The pharmacological potential of Phyllanthus niruri. J. Pharm. Pharmacol. 2016, 68, 953–969. [Google Scholar] [CrossRef]
- Teramoto, J.R.S.; Oliveira, R.F.; Santos, A.S.; Rehder, V.L.G. Avaliação dos teores das lignanas filantina, hipofilantina e nirantina em quebra-pedra (Phyllanthus amarus Schumach. & Thonn.), sob condições de deficiência hídrica. Rev. Bras. Plantas Med. 2010, 4, 67–75. [Google Scholar]
- Elfahmi; Batterman, S.; Koulman, A.; Hackl, T.; Bos, R.; Kayser, O.; Woerdenbag, H.J.; Quax, W.J. Lignans from cell suspension cultures of Phyllanthus niruri, an Indonesian medicinal plant. J. Nat. Prod. 2006, 69, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Al-Qurainy, F.; Ram, M.; Ahmad, S.; Abdin, M.Z. Phyllanthin biosynthesis in Phyllanthus amarus: Schum and Thonn growing at different altitudes. J. Med. Plants Res. 2010, 4, 41–48. [Google Scholar]
- De Saliba, E.O.S.; Rodriguez, N.M.; de Morais, S.A.L.; Piló-Veloso, D. Ligninas: Métodos de obtenção e caracterização química. Ciência Rural 2001, 31, 917–928. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Y.; Cheng, Q.; Niu, M.; Liang, H.; Yan, H.; Zhang, X.; Teixeira da Silva, J.A.; Ma, G. Plant regeneration via direct and callus-mediated organogenesis from leaf explants of Chirita swinglei (Merr.) W. T. Wang. Vitr. Cell. Dev. Biol. Plant 2016, 52, 521–529. [Google Scholar] [CrossRef]
- Huang, R.-L.; Huang, Y.-L.; Ou, J.-C.; Chen, C.-C.; Hsu, F.-L.; Chang, C. Screening of 25 compounds isolated fromPhyllanthus species for anti-human hepatitis B virusin vitro. Phyther. Res. 2003, 17, 449–453. [Google Scholar] [CrossRef]
- Liu, S.; Wei, W.; Shi, K.; Cao, X.; Zhou, M.; Liu, Z. In vitro and in vivo anti-hepatitis B virus activities of the lignan niranthin isolated from Phyllanthus niruri L. J. Ethnopharmacol. 2014, 155, 1061–1067. [Google Scholar] [CrossRef]
- Anuar, N.; Markom, M.; Khairedin, S.; Johari, N.A. Production and extraction of Quercetin and (+)-Chatechin from Phyllanthus niruri callus culture. Int. J. Biol. Sci. 2012, 6, 1240–1243. [Google Scholar]
- Koohang, A.; Majewski, N.D.; Szotek, E.L.; Mar, A.A.; Eiznhamer, D.A.; Flavin, M.T.; Xu, Z.Q. Synthesis and cytotoxicity of 2-cyano-28-hydroxy-lup-1-en-3-ones. Bioorg. Med. Chem. Lett. 2009, 19, 2168–2171. [Google Scholar] [CrossRef]
- Aye Mar, A.; Koohang, A.; Majewski, N.D.; Szotek, E.L.; Eiznhamer, D.A.; Flavin, M.T.; Xu, Z.Q. Synthesis and cytotoxicity of 28-carboxymethoxy lupane triterpenoids. Preference of 28-O-acylation over 28-O-alkylation of betulin by ethyl bromoacetate. Chin. Chem. Lett. 2009, 20, 1141–1144. [Google Scholar] [CrossRef]
- Grishko, V.V.; Tarasova, E.V.; Ivshina, I.B. Biotransformation of betulin to betulone by growing and resting cells of the actinobacterium Rhodococcus rhodochrous IEGM 66. Process Biochem. 2013, 48, 1640–1644. [Google Scholar] [CrossRef]
- Bębenek, E.; Kadela-Tomanek, M.; Chrobak, E.; Wietrzyk, J.; Sadowska, J.; Boryczka, S. New acetylenic derivatives of betulin and betulone, synthesis and cytotoxic activity. Med. Chem. Res. 2017, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bębenek, E.; Kadela-Tomanek, M.; Chrobak, E.; Latocha, M.; Boryczka, S. Novel triazoles of 3-acetylbetulin and betulone as anticancer agents. Med. Chem. Res. 2018, 27, 2051–2061. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| 2,4-D | Fresh Weight | BA/Callus Diameter (cm) Mean ± SD | ||
|---|---|---|---|---|
| (mg/L) | (g/callus) Mean ± SD | 0 mg/L | 1 mg/L | 2 mg/L |
| 0 | 0.00 ± 0.00 c | 0.00 ± 0.00 cA | 0.00 ± 0.00 cA | 0.00 ± 0.00 bA |
| 0.25 | 2.00 ± 0.23 a | 2.58 ± 0.30 aA | 2.00 ± 0.30 aB | 1.76 ± 0.37 aB |
| 0.50 | 1.41 ± 0.30 b | 2.23 ± 0.37 bA | 1.59 ± 0.21 bB | 1.57 ± 0.04 aB |
| 0.75 | 1.24 ± 0.20 b | 1.99 ± 0.58 bA | 1.43 ± 0.20 bB | 1.56 ± 0.29 aB |
| F (2,4-D) 1 | 103.07 ** | 294.05 ** | ||
| F (BA) | 5.57 ** | 31.10 ** | ||
| F (2,4-D × BA) | 0.94 ns | 4.39 ** | ||
| CV (%) | 11.55 | 21.73 | ||
| 2,4-D | BA | BA | ||||
|---|---|---|---|---|---|---|
| (mg/L) | 0 mg/L | 1 mg/L | 2 mg/L | 0 mg/L | 1 mg/L | 2 mg/L |
| Callus Diameter (cm) Mean ± SD | Fresh Weight (g) Mean ± SD | |||||
| 0.00 | 1.01 ± 0.23 cA | 1.11 ± 0.36 bA | 1.09 ± 0.41 dA | 0.17 ± 0.31 bA | 0.32 ± 0.24 bA | 0.30 ± 0.41 bA |
| 0.25 | 1.05 ± 0.26 cB | 2.03 ± 0.26 aA | 2.17 ± 0.56 aA | 0.16 ± 0.37 bB | 1.79 ± 0.34 aA | 2.01 ± 0.24 aA |
| 0.50 | 2.39 ± 0.28 aA | 1.85 ± 0.29 aB | 1.73 ± 0.31 bB | 1.89 ± 0.44 aA | 1.29 ± 0.28 aA | 1.37 ± 0.44 aA |
| 0.75 | 2.04 ± 0.41 bA | 1.66 ± 0.58 aB | 1.50 ± 0.17 cB | 2.30 ± 0.33 aA | 1.15 ± 0.38 aB | 0.86 ± 0.31 bB |
| F (2,4-D) 1 | 35.30 ** | 31.97 ** | ||||
| F (BA) | 0.16 ns | 0.31 ns | ||||
| F (2,4-D × BA) | 14.35 ** | 14.34 ** | ||||
| CV (%) | 24.39 | 14.34 | ||||
| BA (mg/L) 1 | Fresh Weight (g/callus) Mean ± SD |
|---|---|
| 0.0 | 1.19 ± 0.44 d |
| 1.0 | 1.50 ± 0.31 c |
| 2.0 | 1.92 ± 0.46 a |
| 3.0 | 1.76 ± 0.38 b |
| 4.0 | 1.69 ± 0.33 b |
| F | 133.74 ** |
| CV (%) | 3.36 |
| Elicitors 1 | 0.5 mg/L 2,4-D + Darkness | 2.0 mg/L BA + Light | |
|---|---|---|---|
| Diameter (cm) Mean ± SD | Fresh Weight (g/callus) Mean ± SD | Fresh Weight (g/callus) Mean ± SD | |
| Control | 0.50 ± 0.24 a | 1.51 ± 0.45 a | 1.78 ± 0.41 a |
| Salicylic acid | 0.35 ± 0.38 a | 0.90 ± 0.38 c | 1.83 ± 0.33 a |
| Chitosan | 0.40 ± 0.44 a | 1.31 ± 0.36 b | 1.77 ± 0.48 a |
| F 1 | 0.33 ns | 24.59 ** | 0.07 ns |
| CV (%) | 13.17 | 15.89 | 21.16 |
| Rt (min) | Formula | Molar Weight | Biomolecule | Greenhouse Plants | Callus | References | |||
|---|---|---|---|---|---|---|---|---|---|
| BA + Light (Relative Content %) | 2,4-D + Dark | ||||||||
| (g·mol−1) | (Leaves + Stem) | Control | Salicylic Acid | Chitosan | Salicylic Acid | ||||
| 10.86 | C15H24 | 204 | alpha-muurolene | 1.33 | 2.23 | NIST-11 | |||
| ni | 0.61 | ||||||||
| 298 | ni | 13.90 | |||||||
| 300 | ni | 3.97 | 2.49 | 14.69 | |||||
| 314 | ni | 0.81 | |||||||
| 316 | ni | 1.50 | |||||||
| 386 | ni | 3.15% | |||||||
| 392 | ni | 0.50 | |||||||
| 394 | ni | 0.59 | |||||||
| 16.40 | C23H30O6 | 402 | 5-demethoxy-niranthin | 23.34% | Library CPQBA | ||||
| 16.91 | C24H34O6 | 418 | Phyllanthin | 30.03% | Library CPQBA | ||||
| 17.02 | C24H32O6 | 416 | Phyltetralin | 6.19% | Library CPQBA | ||||
| 17.34 | C23H28O6 | 400 | 5-demethoxy-nirtetralin | 10.01% | Library CPQBA | ||||
| 18.26 | C24H30O7 | 430 | Nirtetralin | 3.07% | Library CPQBA | ||||
| 19.85 | C24H32O7 | 432 | Niranthin | 18.92% | Library CPQBA | ||||
| 24.96 | C29H48O | 412 | Stigmasterol | 5.22 | 8.14 | 5.45 | 8.02 | NIST-11 | |
| 25.04 | C30H50 | 410 | Squalene | 0.73 | NIST-11 | ||||
| 26.86 | C29H50O | 414 | γ-Sitosterol | 2.56 | 3.35 | 5.14 | 6.72 | NIST-11 | |
| 28.42 | C30H50O | 426 | β-Amyrin | 2.58 | 2.57 | NIST-11 | |||
| 422 | ni | 9.93 | 8.95 | 7.90 | 29.10 | ||||
| ni | |||||||||
| ni | 4.74 | 4.57 | 2.60 | ||||||
| ni | 2.34 | ||||||||
| ni | 6.2 | ||||||||
| 32.67 | C30H48O2 | 440 | Betulin aldehyde or betulone | 62.5 | 60.92 | 75.98 | 4.69 | NIST-11 | |
| ni | 4.29 | 2.32 | |||||||
| ni | 2.88 | ||||||||
| Total Molecules | 7 | 10 | 10 | 7 | 11 | ||||
Sample Availability: Samples of the compounds are available from the authors. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira, M.E.B.S.; Sartoratto, A.; Carlos Cardoso, J. In Vitro Calli Production Resulted in Different Profiles of Plant-Derived Medicinal Compounds in Phyllanthus amarus. Molecules 2020, 25, 5895. https://doi.org/10.3390/molecules25245895
de Oliveira MEBS, Sartoratto A, Carlos Cardoso J. In Vitro Calli Production Resulted in Different Profiles of Plant-Derived Medicinal Compounds in Phyllanthus amarus. Molecules. 2020; 25(24):5895. https://doi.org/10.3390/molecules25245895
Chicago/Turabian Stylede Oliveira, Maria Eduarda B. S., Adilson Sartoratto, and Jean Carlos Cardoso. 2020. "In Vitro Calli Production Resulted in Different Profiles of Plant-Derived Medicinal Compounds in Phyllanthus amarus" Molecules 25, no. 24: 5895. https://doi.org/10.3390/molecules25245895
APA Stylede Oliveira, M. E. B. S., Sartoratto, A., & Carlos Cardoso, J. (2020). In Vitro Calli Production Resulted in Different Profiles of Plant-Derived Medicinal Compounds in Phyllanthus amarus. Molecules, 25(24), 5895. https://doi.org/10.3390/molecules25245895