
Localization of Flavan-3-ol Species in Peanut Testa by Mass Spectrometry Imaging



Abstract
1. Introduction
2. Results
2.1. Mass Spectra Obtained from Peanut Section
2.2. Visualization of Flavan-3-ols in Peanut Sections
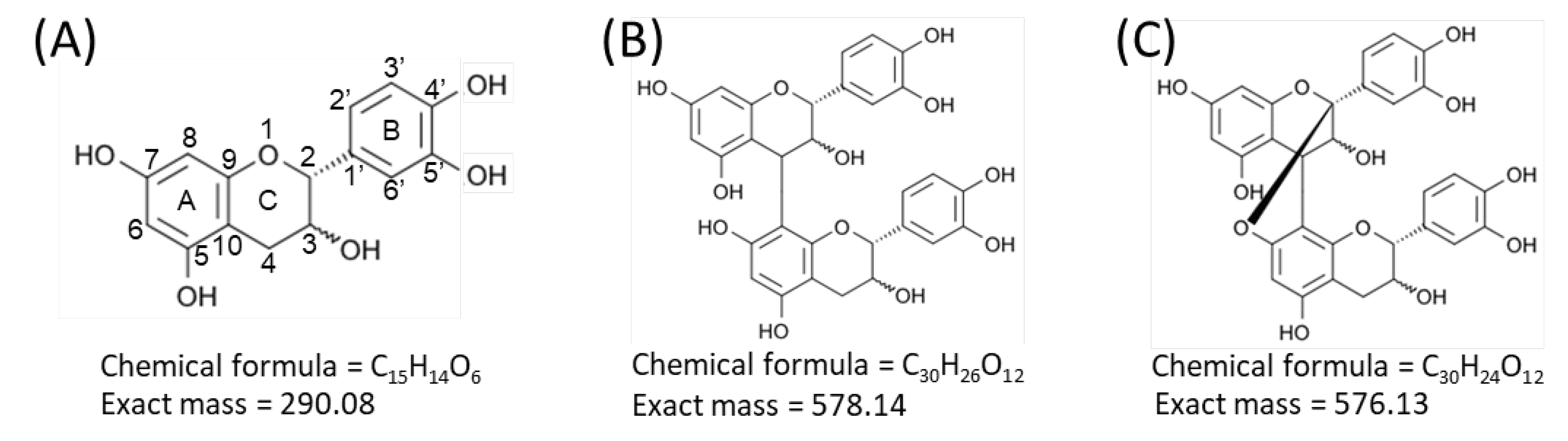
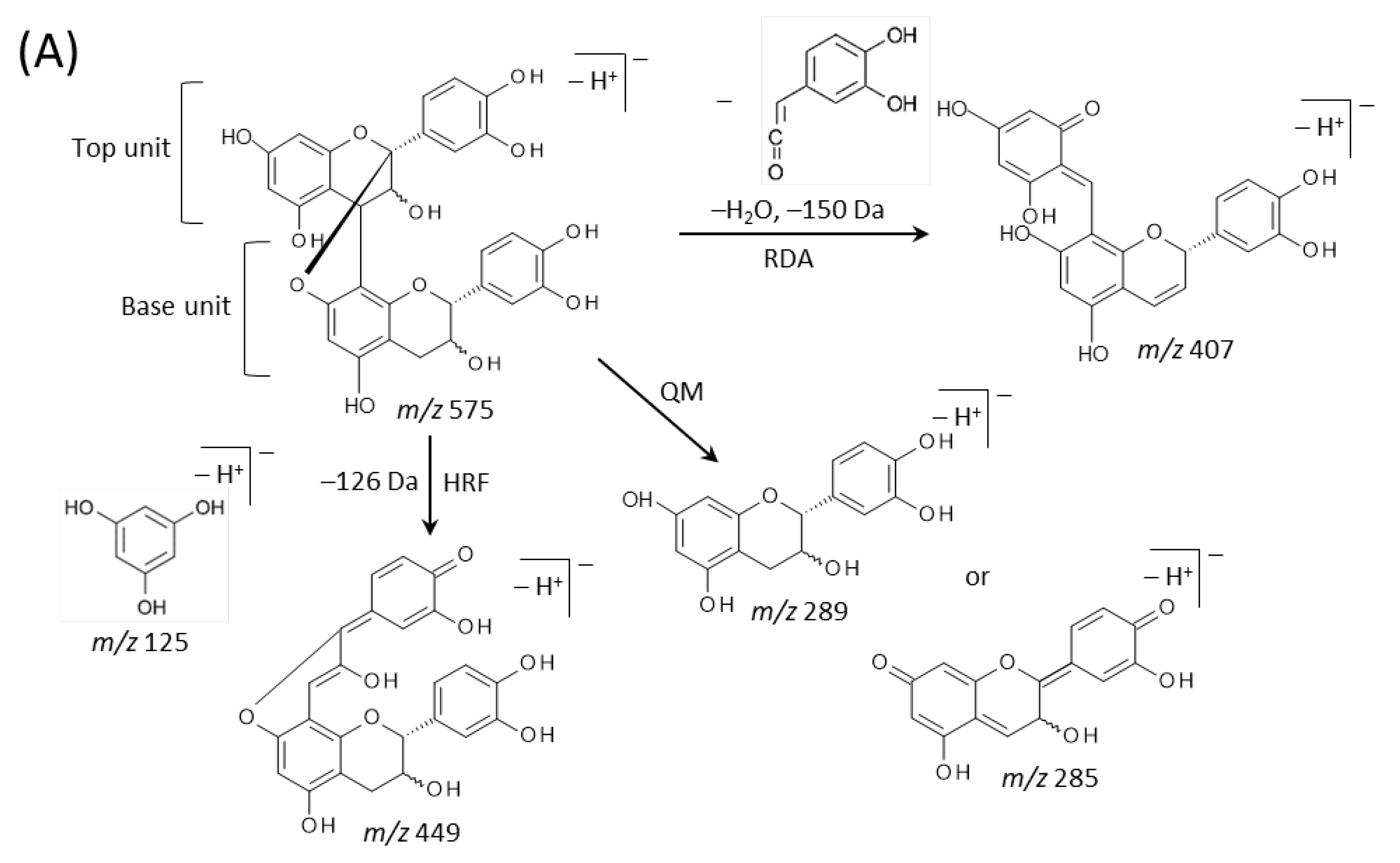
2.3. Identification of Flavan-3-ols in Peanut Sections
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Peanut (Arachis hypogaea L.) Samples
4.3. Preparation of Peanut Sections
4.4. Matrix Coating
4.5. MALDI-MSI Analysis of Peanut Sections
4.6. MALDI-MS/MS Analysis of Peanut Sections
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, F.; Pan, Q.H.; Shi, Y.; Duan, C.Q. Biosynthesis and genetic regulation of proanthocyanidins in plants. Molecules 2008, 13, 2674–2703. [Google Scholar] [CrossRef]
- Rue, E.A.; Rush, M.D.; van Breemen, R.B. Procyanidins: A comprehensive review encompassing structure elucidation via mass spectrometry. Phytochem. Rev. 2018, 17, 1–16. [Google Scholar] [CrossRef]
- Yu, J.; Ahmedna, M.; Goktepe, I.; Dai, J. Peanut skin procyanidins: Composition and antioxidant activities as affected by processing. J. Food Compost. Anal. 2006, 19, 364–371. [Google Scholar] [CrossRef]
- Tamura, T.; Inoue, N.; Ozawa, M.; Shimizu-Ibuka, A.; Arai, S.; Abe, N.; Koshino, H.; Mura, K. Peanut-skin polyphenols, procyanidin A1 and epicatechin-(4 β → 6)-epicatechin-(2 β → O → 7, 4 β → 8)-catechin, exert cholesterol micelle-degrading activity in vitro. Biosci. Biotechnol. Biochem. 2013, 77, 1306–1309. [Google Scholar] [CrossRef]
- Tomochika, K.; Shimizu-Ibuka, A.; Tamura, T.; Mura, K.; Abe, N.; Onose, J.; Arai, S. Effects of peanut-skin procyanidin A1 on degranulation of RBL-2H3 cells. Biosci. Biotechnol. Biochem. 2011, 75, 1644–1648. [Google Scholar] [CrossRef]
- Dudek, M.K.; Gliński, V.B.; Davey, M.H.; Sliva, D.; Kaźmierski, S.; Gliński, J.A. Trimeric and Tetrameric A-Type Procyanidins from Peanut Skins. J. Nat. Prod. 2017, 80, 415–426. [Google Scholar] [CrossRef]
- Woodroof, J.G.; Leahy, J.F. Microscopical studies of peanuts with reference to processing. Georgia Exp. Sta. Bull. 1940, 205, 1–39. [Google Scholar]
- Caprioli, R.M.; Farmer, T.B.; Gile, J. Molecular imaging of biological samples: Localization of peptides and proteins using MALDI–TOF MS. Anal. Chem. 1997, 69, 4751–4760. [Google Scholar] [CrossRef]
- Stoeckli, M.; Chaurand, P.; Hallahan, D.E.; Caprioli, R.M. Imaging mass spectrometry: A new technology for the analysis of protein expression in mammalian tissues. Nat. Methods 2001, 7, 493–496. [Google Scholar] [CrossRef]
- Cornett, D.S.; Reyzer, M.L.; Chaurand, P.; Caprioli, R.M. MALDI imaging mass spectrometry: Molecular snapshots of biochemical systems. Nat. Methods 2007, 4, 828–833. [Google Scholar] [CrossRef]
- Enomoto, H.; Kotani, M.; Ohmura, T. Novel blotting method for mass spectrometry imaging of metabolites in strawberry fruit by desorption/ionization using through hole alumina membrane. Foods 2020, 9, 408. [Google Scholar] [CrossRef]
- Nizioł, J.; Misiorek, M.; Ruman, T. Mass spectrometry imaging of low molecular weight metabolites in strawberry fruit (Fragaria x ananassa Duch.) cv. Primoris with 109Ag nanoparticle enhanced target. Phytochemistry 2019, 159, 11–19. [Google Scholar] [CrossRef]
- Cooks, R.G.; Ouyang, Z.; Takats, Z.; Wiseman, J.M. Ambient mass spectrometry. Science 2006, 311, 1566–1570. [Google Scholar] [CrossRef]
- Enomoto, H.; Sensu, T.; Yumoto, E.; Yokota, T.; Yamane, H. Derivatization for detection of abscisic acid and 12-oxo-phytodienoic acid using matrix-assisted laser desorption/ionization imaging mass spectrometry. Rapid Commun. Mass Spectrom. 2018, 32, 1565–1572. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Goto-lnoue, N.; Moriyama, T.; Zaima, N. Significant advancement of mass spectrometry imaging for food chemistry. Food Chem. 2016, 210, 200–211. [Google Scholar] [CrossRef]
- Zaima, N.; Hayasaka, T.; Goto-Inoue, N.; Setou, M. Matrix-assisted laser desorption/ionization imaging mass spectrometry. Int. J. Mol. Sci. 2010, 11, 5040–5055. [Google Scholar] [CrossRef]
- Zaima, N.; Goto-Inoue, N.; Hayasaka, T.; Enomoto, H.; Setou, M. Authenticity assessment of beef origin by principal component analysis of matrix-assisted laser desorption/ionization mass spectrometric data. Anal. Bioanal. Chem. 2011, 400, 1865–1871. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Enomoto, H.; Moriyama, T.; Kawamura, Y.; Setou, M.; Zamia, N. Visualization of anthocyanin species in rabbiteye blueberry Vaccinium ashei by matrix-assisted laser desorption/ionization imaging mass spectrometry. Anal. Bioanal. Chem. 2012, 403, 1885–1895. [Google Scholar] [CrossRef]
- Enomoto, H.; Sato, K.; Miyamoto, K.; Ohtsuka, A.; Yamane, H. Distribution analysis of anthocyanins, sugars, and organic acids in strawberry fruits using matrix-assisted laser desorption/ionization-imaging mass spectrometry. J. Agric. Food Chem. 2018, 66, 4958–4965. [Google Scholar] [CrossRef]
- Enomoto, H.; Takahashi, S.; Takeda, S.; Hatta, H. Distribution of flavan-3-ol species in ripe strawberry fruit revealed by matrix-assisted laser desorption/ionization-mass spectrometry imaging. Molecules 2020, 25, 103. [Google Scholar] [CrossRef]
- Enomoto, H.; Takeda, S.; Hatta, H.; Zaima, N. Tissue-specific distribution of sphingomyelin species in pork chop revealed by matrix-assisted laser desorption/ionization-imaging mass spectrometry. J. Food Sci. 2019, 84, 1758–1763. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, H.; Furukawa, T.; Takeda, S.; Hatta, H.; Zaima, N. Unique distribution of diacyl-, alkylacyl-, and alkenylacyl-phosphatidylcholine species visualized in pork chop tissues by matrix-assisted laser desorption/ionization–mass spectrometry imaging. Foods 2020, 9, 205. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, H.; Sensu, T.; Sato, K.; Sato, F.; Paxton, T.; Yumoto, E.; Miyamoto, K.; Asahina, M.; Yokota, T.; Yamane, H. Visualisation of abscisic acid and 12-oxo-phytodienoic acid in immature Phaseolus vulgaris L. seeds using desorption electrospray ionisation-imaging mass spectrometry. Sci. Rep. 2017, 7, 42977. [Google Scholar] [CrossRef]
- Meisenbichler, C.; Doppler, C.; Bernhard, D.; Müller, T. Improved matrix coating for positive- and negative-ion-mode MALDI–TOF imaging of lipids in blood vessel tissues. Anal. Bioanal. Chem. 2019, 411, 3221–3227. [Google Scholar] [CrossRef]
- Shimma, S.; Takashima, Y.; Hashimoto, J.; Yonemori, K.; Tamura, K.; Hamada, A. Alternative two-step matrix application method for imaging mass spectrometry to avoid tissue shrinkage and improve ionization efficiency. J. Mass Spectrom. 2013, 48, 1285–1290. [Google Scholar] [CrossRef]
- Rush, M.D.; Rue, E.A.; Wong, A.; Kowalski, P.; Glinski, J.A.; van Breemen, R.B. Rapid determination of procyanidins using MALDI–ToF/ToF mass spectrometry. J. Agric. Food Chem. 2018, 66, 11355–11361. [Google Scholar] [CrossRef]
- Francisco, M.L.D.L.; Resurreccion, A.V.A. Functional components in peanuts. Crit. Rev. Food Sci. Nutr. 2008, 48, 715–746. [Google Scholar] [CrossRef]
- Chen, L.; Yuan, P.; Chen, K.; Jia, Q.; Li, Y. Oxidative conversion of B- to A-type procyanidin trimer: Evidence for quinone methide mechanism. Food Chem. 2014, 154, 315–322. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Precursor Ion [M − H]−, (m/z) | Flavan-3-ol Species | Number of Linkage | Fragment Ions for Assignment [M − H]−, (m/z) | |||
|---|---|---|---|---|---|---|
| A-Type | B-Type | |||||
| 289.1 | Catechin and/or epicatechin | 245, 125 | ||||
| QM | RDA | HRF | ||||
| 575.1 | A-type procyanidin dimers | 1 | 0 | 289, 285 | 407 | 449 |
| 577.1 | B-type procyanidin dimers | 0 | 1 | 289, 287 | 425, 407 | 451 |
| 861.2 | A-type procyanidin trimers | 2 | 0 | 575, 289, 285 | 693, 407 | 735, 449 |
| 863.2 | A-type procyanidin trimers | 1 | 1 | 575, 573, 289, 287, 285 | 711, 695, 693, 407 | 737, 451, 449 |
| 1149.3 | A-type procyanidin tetramers | 2 | 1 | 863, 861, 575, 573, 289, 287, 285 | 997, 981, 979, 711, 695, 693, 425, 407 | 1023, 737, 735, 451, 449 |
| 1151.3 | A-type procyanidin tetramers | 1 | 2 | 863, 861, 575, 573, 289, 287, 285 | 999, 981, 711, 695, 693, 425, 407 | 1025, 739, 737, 451, 449 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enomoto, H.; Nirasawa, T. Localization of Flavan-3-ol Species in Peanut Testa by Mass Spectrometry Imaging. Molecules 2020, 25, 2373. https://doi.org/10.3390/molecules25102373
Enomoto H, Nirasawa T. Localization of Flavan-3-ol Species in Peanut Testa by Mass Spectrometry Imaging. Molecules. 2020; 25(10):2373. https://doi.org/10.3390/molecules25102373
Chicago/Turabian StyleEnomoto, Hirofumi, and Takashi Nirasawa. 2020. "Localization of Flavan-3-ol Species in Peanut Testa by Mass Spectrometry Imaging" Molecules 25, no. 10: 2373. https://doi.org/10.3390/molecules25102373
APA StyleEnomoto, H., & Nirasawa, T. (2020). Localization of Flavan-3-ol Species in Peanut Testa by Mass Spectrometry Imaging. Molecules, 25(10), 2373. https://doi.org/10.3390/molecules25102373