
Anti-Inflammatory Activity of the Wild Mushroom, Echinodontium tinctorium, in RAW264.7 Macrophage Cells and Mouse Microcirculation

 , , ,
, , , 
Abstract
1. Introduction
2. Results and Discussion
2.1. Identification of Echinodontium tinctorium
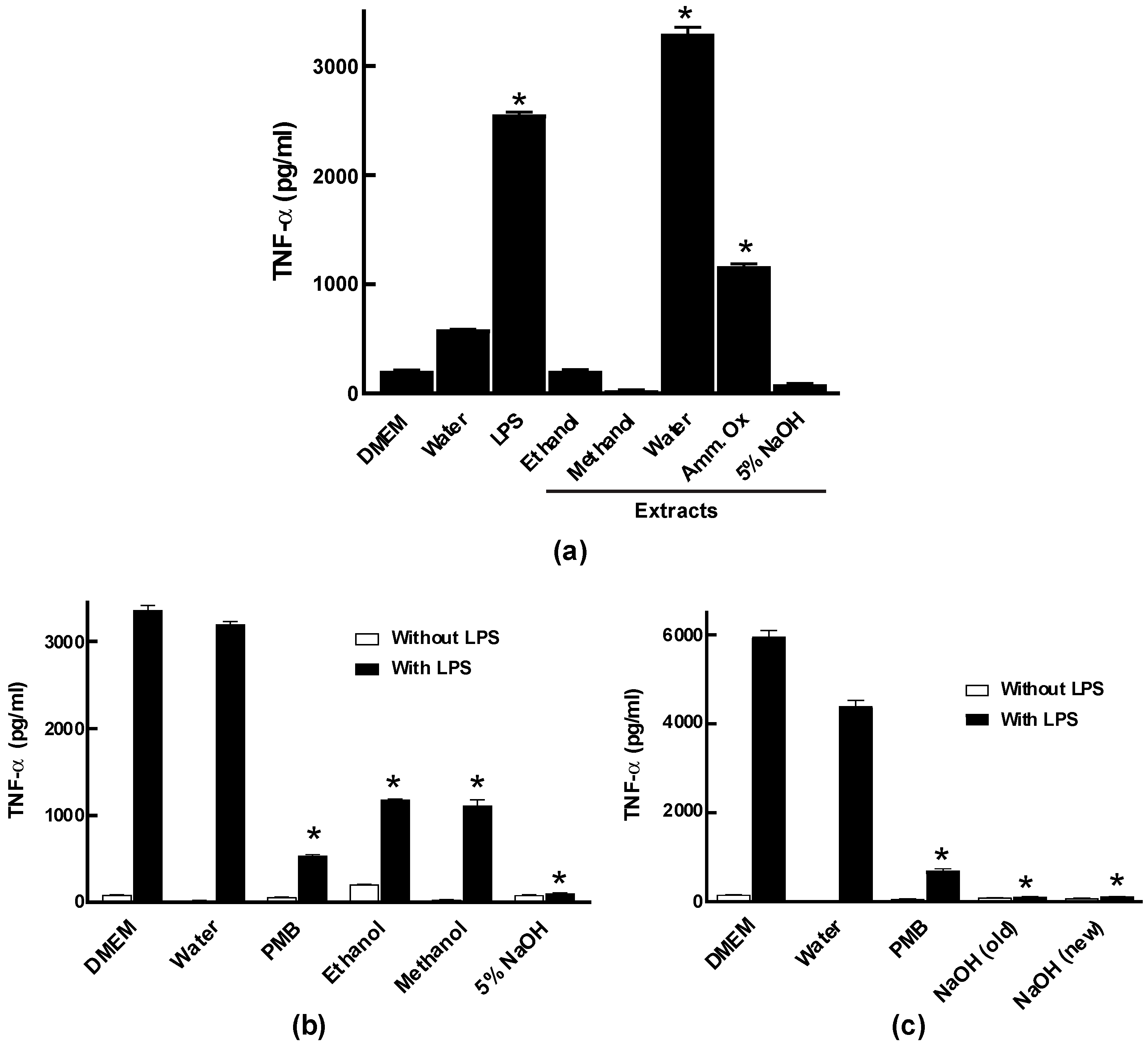
2.2. Chemical Extraction and Assessment of Crude Extracts of E. tinctorium for Bio-Activities
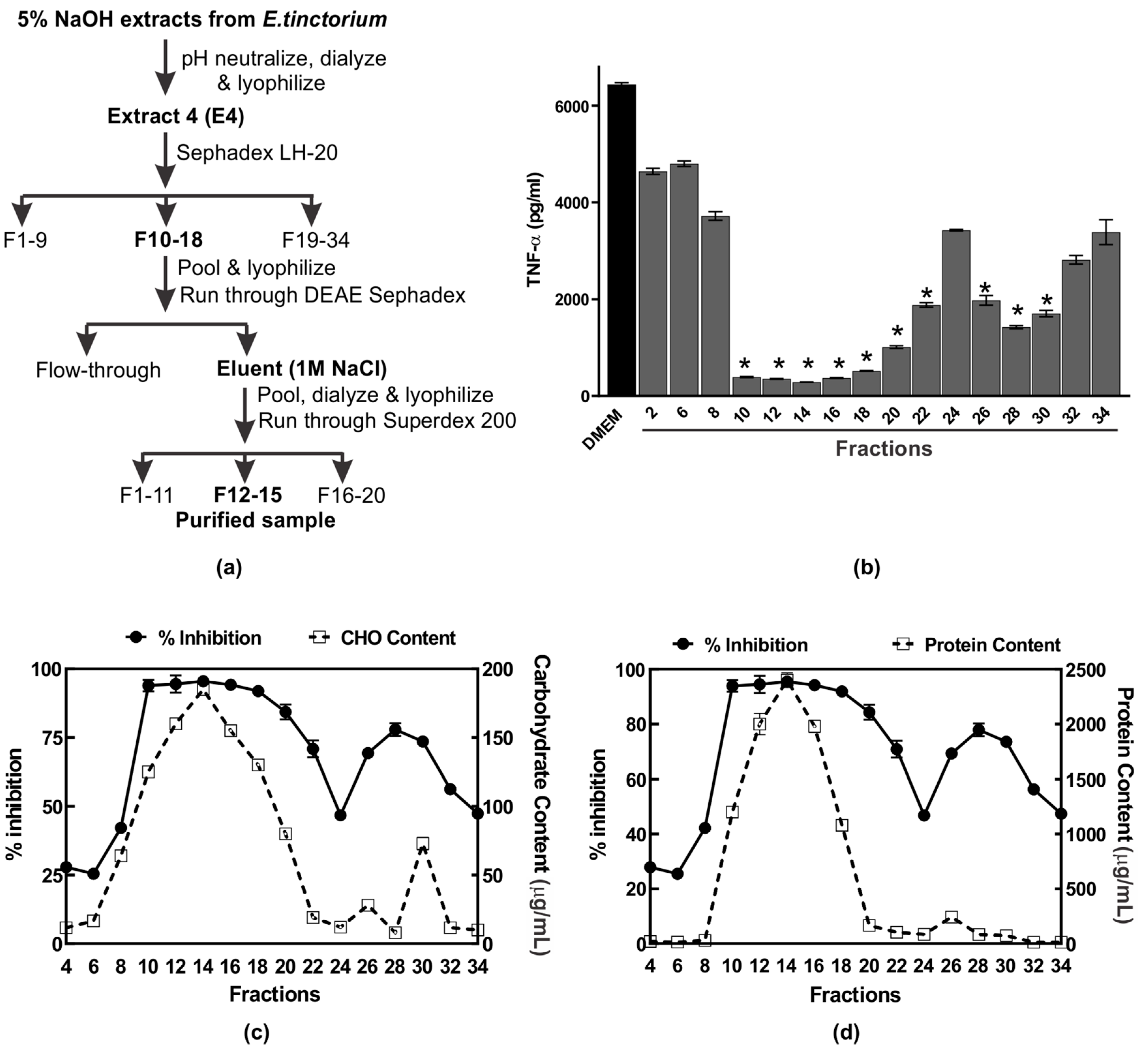
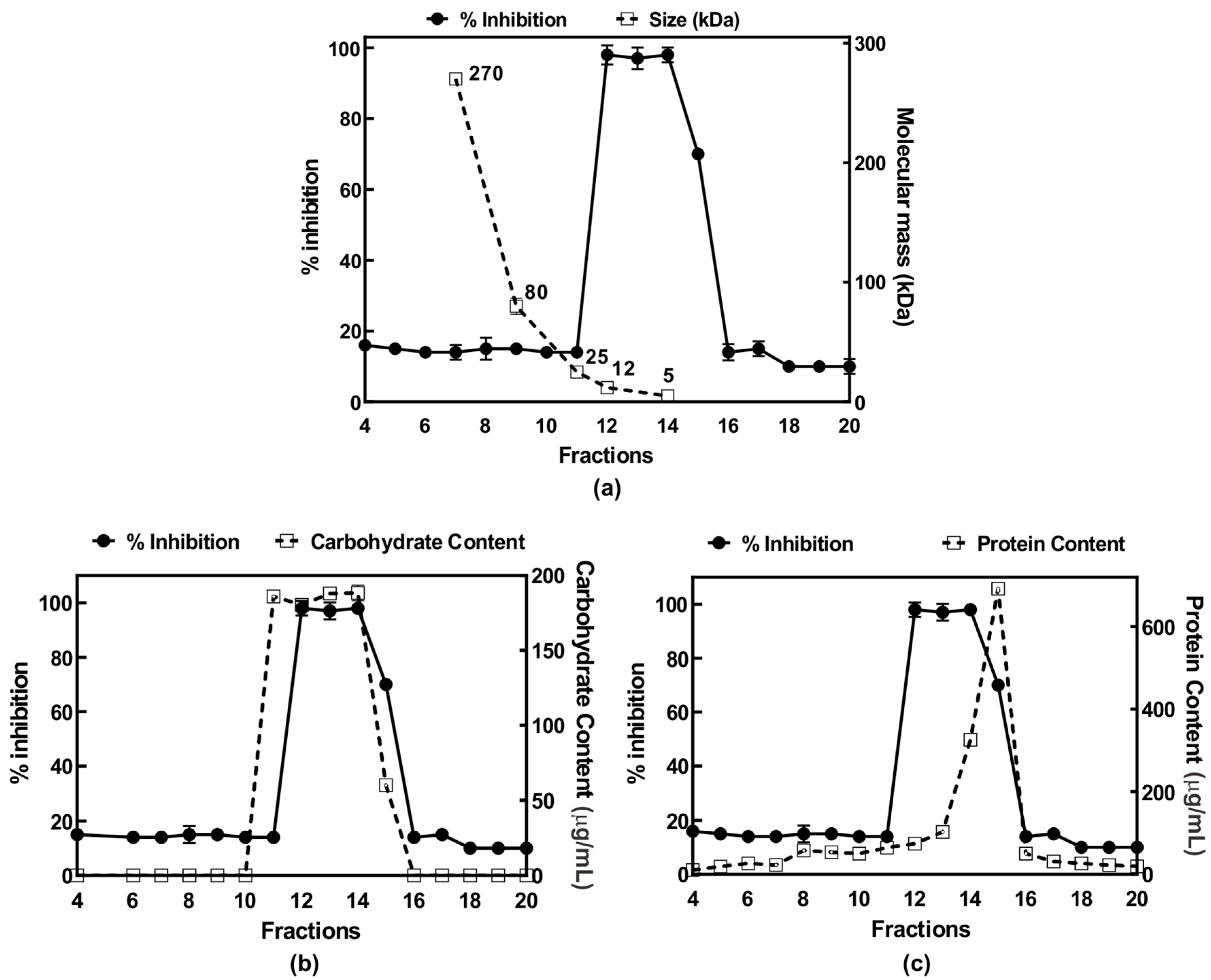
2.3. Purification of Anti-Inflammatory Polysaccharide from E. tinctorium
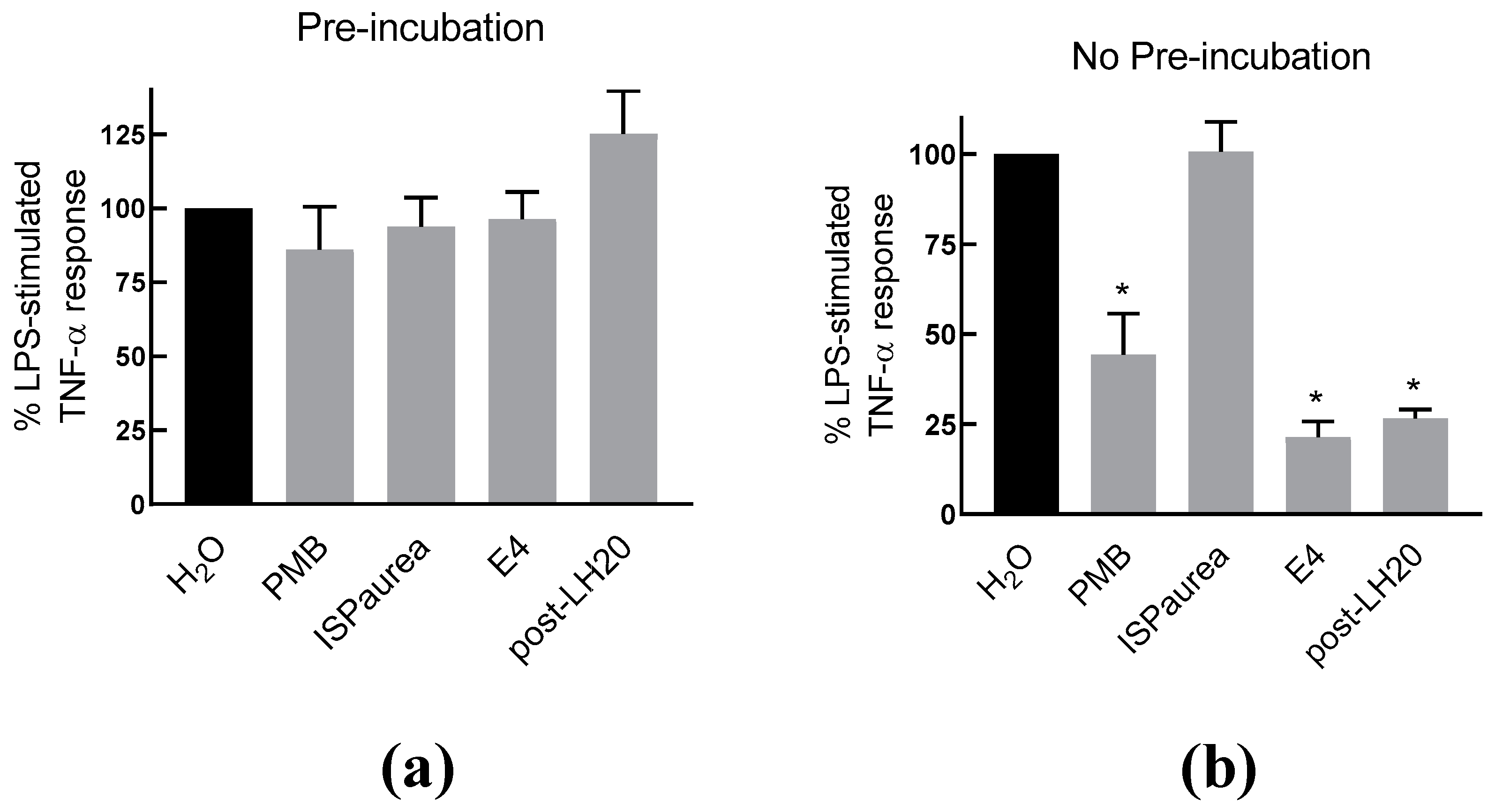
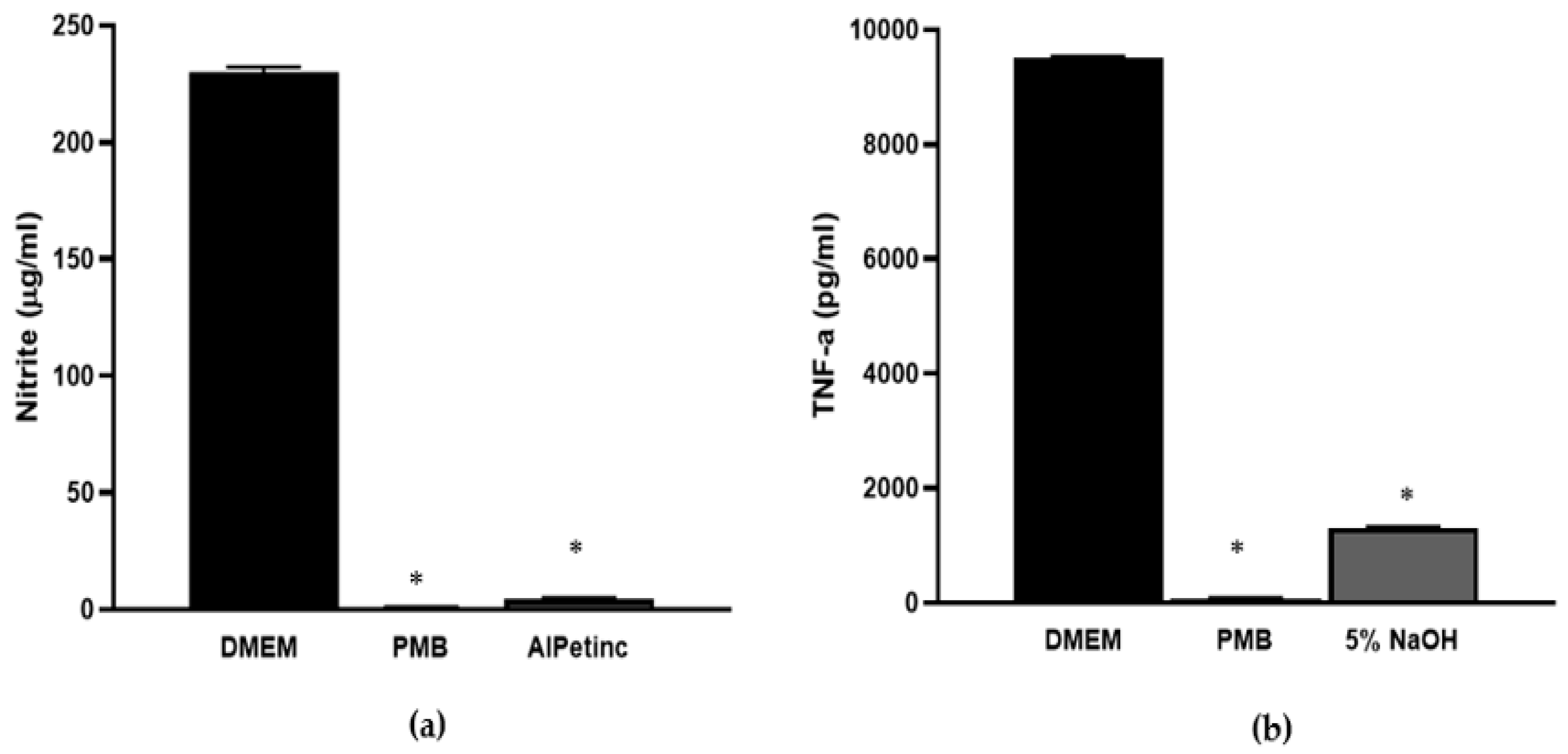
2.4. AIPetinc Inhibited LPS-Induced NO Production and Histamine-Induced TNF-α Production in RAW264.7 Macrophage Cells
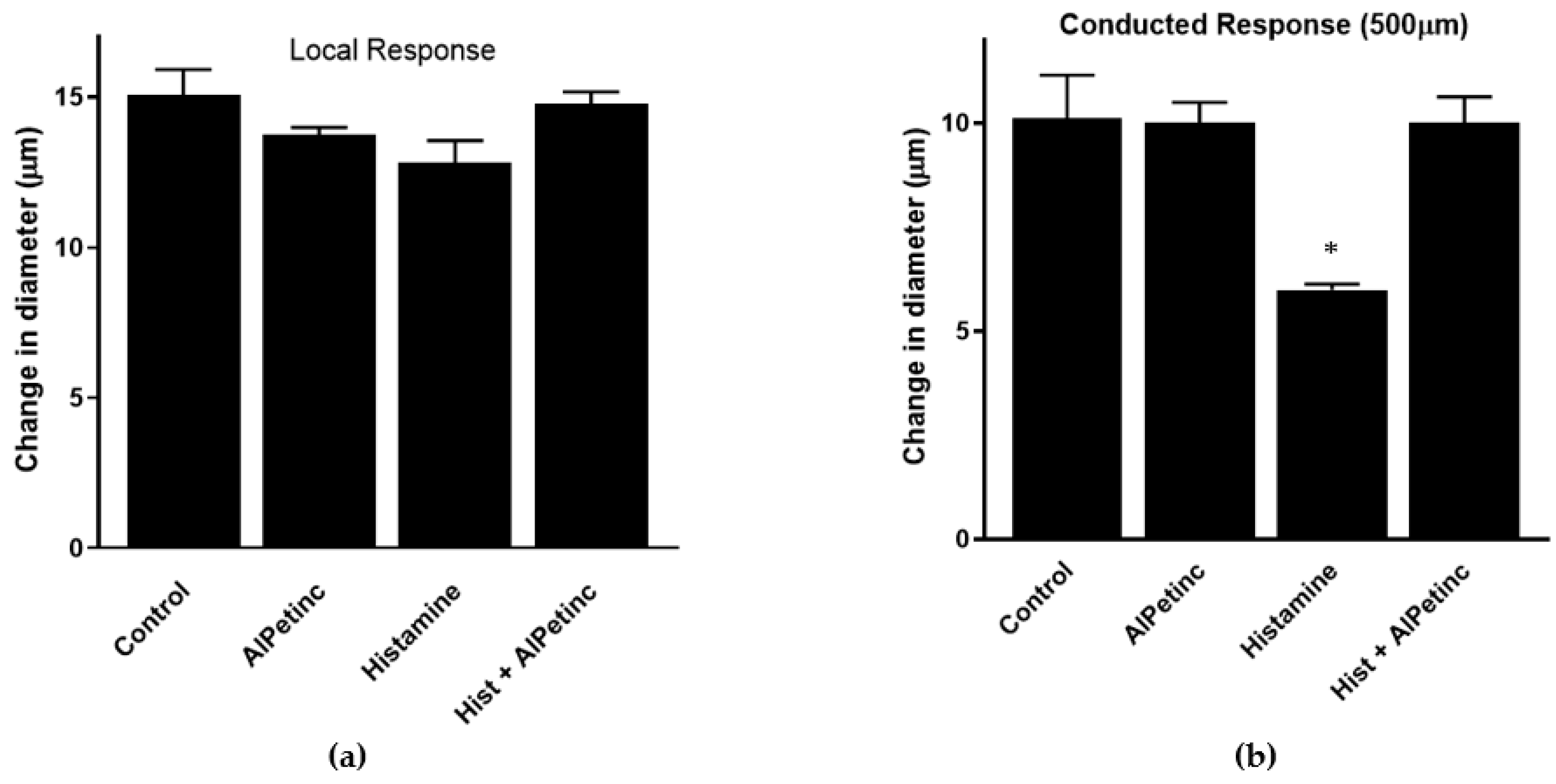
2.5. AIPetinc Blocked Histamine-Induced Inflammatory Event in Mice Microcirculation
2.6. Chemical Analysis of the Anti-Inflammatory Polysaccharide from E. tinctoirum
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Identification of Mushroom Species
3.3. Extraction and Purification of Anti-Inflammatory Polysaccharide
3.4. Monosaccharide Composition Analysis
3.5. Determining the Percentage of α- and β-Glucans in AIPetinc
3.6. Cell Line and Assessment for Immuno-Stimulatory Activity
3.7. Anti-Inflammatory Activity In Vitro
3.8. Nitric Oxide Determination Assay
3.9. Intravital Microscopy and Measurement of Conducted Vasodilation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, A.; Javed, S.; Barad, A.; Myhre, V.; Li, W.M.; Reimer, K.; Massicotte, H.B.; Tackaberry, L.E.; Payne, G.W.; Egger, K.N.; et al. Growth-inhibitory and immunomodulatory activities of wild mushrooms from North-Central British Columbia (Canada). Int. J. Med. Mushrooms. 2017, 19, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Deo, G.; Khatra, J.; Buttar, S.; Li, W.M.; Tackaberry, L.E.; Massicotte, H.B.; Egger, K.N.; Reimer, K.; Lee, C.H. Anti-proliferative, immuno-stimulatory and anti-inflammatory activities of extracts derived from mushrooms collected in Haida Gwaii, British Columbia, Canada. Int. J. Med. Mushrooms. 2019, 21, 629–643. [Google Scholar] [CrossRef]
- Allen, E.A.; Morrison, D.J.; Wallis, G.W. Common Tree Diseases of British Columbia; Pacific Forestry Center: Hull, QC, Canada, 1996. [Google Scholar]
- Gilbertson, R.L.; Ryvarden, L. North American Polypores; Fungiflora: Oslo, Norway, 1986; Volume 1, p. 433. [Google Scholar]
- Hansen, E.M.; Lewis, K.J.; Chastagner, G.A. Compendium of Conifer Diseases, 2nd ed.; APS Press, the American Phytopathological Society: St. Paul, MN, USA, 2018; p. 224. [Google Scholar]
- Ginns, J.H. Polypores of British Columbia (Fungi: Basidiomycota); Province of British Columbia: Victoria, BC, USA, 2017. [Google Scholar]
- Gross, H.L. The Echinodontiaceae. Mycopathologia et Mycologia Applicata. 1964, 24, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-L.; Zhao, Y.; Dai, Y.-C.; Nakasone, K.K.; He, S.-H. Phylogeny and taxonomy of Echinodontium and related genera. Mycologia 2017, 109, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Tabata, M.; Harrington, T.C.; Chen, W.; Abe, Y. Molecular phylogeny of species in the genera Amylostereum and Echinodontium. Mycoscience 2000, 41, 585–593. [Google Scholar] [CrossRef]
- Ye, Y.; Josten, I.; Arnold, N.; Steffan, B.; Steglich, W. Isolation of a fluorone pigment from the Indian paint fungus Echinodontium tinctorium and Pyrofomes albomarginatus. Tetrahedron 1996, 52, 5793–5798. [Google Scholar] [CrossRef]
- Bond, F.T.; Fullerton, D.S.; Sciuchetti, L.A.; Catalfomo, P. The isolation and structure of echinodol, a triterpene acetate. J. Amer. Chem. Soc. 1966, 88, 3882–3883. [Google Scholar] [CrossRef] [PubMed]
- Arnone, A.; Brambilla, U.; Nasini, G.; Pava, O.V.D. Isolation and structure elucidation of tsugicolines A-D, novel protoilludane sesquiterpenes from Laurilia tsugicola. Tetrahedron 1995, 51, 13357–13364. [Google Scholar] [CrossRef]
- Arnone, A.; Gregorio, C.D.; Meille, S.V.; Nasini, G.; Sidoti, G. Tsugicoline E, a new polyoxygenated protoilludane sesquiterpene from the fungus Laurilia tsugicola. J. Nat. Prod. 1999, 62, 51–53. [Google Scholar] [CrossRef]
- Arnone, A.; Gregorio, C.D.; Nasini, G.; Pava, O.V.D. Isolation and structure elucidation of tsugicolines F-H, novel furosesquiterpenes, and tsugicoline I from the fungus Laurilia tsugicola. Tetrahedron 1998, 54, 10199–10204. [Google Scholar] [CrossRef]
- Suzuki, S.; Murayama, T.; Shiono, Y. Illudalane sesquiterpenoids, echinolactones A and B, from a mycelial culture of Echinodontium japonicum. Phytochemistry 2005, 66, 2329–2333. [Google Scholar] [CrossRef] [PubMed]
- Taofiq, O.; Martins, A.; Barreiro, M.F.; Ferreira, I.C. Anti-inflammatory potential of mushroom extracts and isolated metabolites. Trends Food Sci. Technol. 2016, 50, 193–210. [Google Scholar] [CrossRef]
- Sevinsky, R.; Stewart, D.; Harirforoosh, S. Nonsteroidal anti-inflammatory drugs: Is there a link between cardiovascular and renal adverse effects? J. Integr. Nephrol. Androl. 2017, 4, 1–2. [Google Scholar]
- Mizuno, T.; Saito, H.; Nishitoba, T.; Kawagishi, H. Antitumor-Active Substances from Mushrooms. Food Rev. Int. 1995, 11, 23–61. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide synthase in innate and adaptive immunity: An update. Trends Immunol. 2015, 36, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.Z.; Baldwin, A.L. Transient venular permeability increase and endothelial gap formation induced by histamine. Am. J. Physiol. 1992, 262, H1238–H1247. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Granger, H.J.; Zawieja, D.C.; DeFily, D.V.; Chilian, W.M. Histamine increases venular permeability via a phospholipase C-NO synthase-guanylate cyclase cascade. Am. J. Physiol. 1993, 264, H1734–H1739. [Google Scholar] [CrossRef] [PubMed]
- Payne, G.W.; Madri, J.A.; Sessa, W.C.; Segal, S.S. Histamine inhibits conducted vasodilation through endothelium-derived NO production in arterioles of mouse skeletal muscle. FASEB J. 2004, 18, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Javed, S.; Mitchell, K.; Sidsworth, D.; Sellers, S.L.; Reutens-Hernandez, J.; Massicotte, H.B.; Egger, K.N.; Lee, C.H.; Payne, G.W. Inonotus obliquus attenuates histamine-induced microvascular inflammation. PLoS ONE 2019, 14, e0220776. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Rosendahl, A.; Wang, X.; Wu, D.; Andersson, R. Polysaccharide-K (PSK) in cancer-old story, new possibilities? Curr. Med. Chem. 2012, 19, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Barad, A.; Mackedenski, S.; Li, W.M.; Li, X.J.; Lim, B.C.C.; Rashid, F.; Tackaberry, L.E.; Massicotte, H.B.; Egger, K.N.; Reimer, K.; et al. Anti-proliferative activity of a purified polysaccharide isolated from the basidiomycete fungus Paxillus involutus. Carbohydr. Polym. 2018, 181, 923–930. [Google Scholar] [CrossRef]
- Du, B.; Lin, C.; Bian, Z.; Xu, B. An insight into anti-inflammatory effects of fungal beta-glucans. Trends Food Sci. Technol. 2015, 41, 49–59. [Google Scholar] [CrossRef]
- Muszynska, B.; Grzywacz-Kisielewska, A.; Kala, K.; Gdula-Argasinska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef] [PubMed]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Analytical Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Blakeney, A.B.; Harris, P.J.; Stone, B.A. A simple and rapid preparation of alditol acetates for monosaccharide analysis. Carbohydr. Res. 1983, 113, 291–299. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Shin, W.-B.; Dong, X.; Kim, E.-K.; Nawarathna, W.P.A.S.; Kim, H.; Park, P.-J. Anti-inflammatory effect of the extract from fermented Asterina pectinifera with Cordyceps militaris mycelia in LPS-induced RAW264.7 macrophages. Food Sci. Biotechnol. 2017, 26, 1633–1640. [Google Scholar] [CrossRef]
- Gunawardena, D.; Bennett, L.; Shanmugam, K.; King, K.; Williams, R.; Zabaras, D.; Head, R.; Ooi, L.; Gyengesi, E.; Münch, G. Anti-inflammatory effects of five commercially available mushroom species determined in lipopolysaccharide and interferon-γ activated murine macrophages. Food Chem. 2014, 148, 92–96. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monosaccharide Composition (%) | Anti-Inflammatory Polysaccharide from E. tinctorium | Growth-Inhibitory Polysaccharide from P. involutus [25] | Polysaccharide-K [24] |
|---|---|---|---|
| Glucose | 88.6 | 65.9 | 74.4 |
| Galactose | 4.0 | 20.8 | 7.1 |
| Mannose | 4.4 | 7.8 | 14.0 |
| Fucose | 0.7 | 3.2 | 1.6 |
| Xylose | 2.3 | 2.3 | 2.7 |
| Ribose | - | - | 0.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javed, S.; Li, W.M.; Zeb, M.; Yaqoob, A.; Tackaberry, L.E.; Massicotte, H.B.; Egger, K.N.; Cheung, P.C.K.; Payne, G.W.; Lee, C.H. Anti-Inflammatory Activity of the Wild Mushroom, Echinodontium tinctorium, in RAW264.7 Macrophage Cells and Mouse Microcirculation. Molecules 2019, 24, 3509. https://doi.org/10.3390/molecules24193509
Javed S, Li WM, Zeb M, Yaqoob A, Tackaberry LE, Massicotte HB, Egger KN, Cheung PCK, Payne GW, Lee CH. Anti-Inflammatory Activity of the Wild Mushroom, Echinodontium tinctorium, in RAW264.7 Macrophage Cells and Mouse Microcirculation. Molecules. 2019; 24(19):3509. https://doi.org/10.3390/molecules24193509
Chicago/Turabian StyleJaved, Sumreen, Wai Ming Li, Mehreen Zeb, Almas Yaqoob, Linda E. Tackaberry, Hugues B. Massicotte, Keith N. Egger, Peter C.K. Cheung, Geoffrey W. Payne, and Chow H. Lee. 2019. "Anti-Inflammatory Activity of the Wild Mushroom, Echinodontium tinctorium, in RAW264.7 Macrophage Cells and Mouse Microcirculation" Molecules 24, no. 19: 3509. https://doi.org/10.3390/molecules24193509
APA StyleJaved, S., Li, W. M., Zeb, M., Yaqoob, A., Tackaberry, L. E., Massicotte, H. B., Egger, K. N., Cheung, P. C. K., Payne, G. W., & Lee, C. H. (2019). Anti-Inflammatory Activity of the Wild Mushroom, Echinodontium tinctorium, in RAW264.7 Macrophage Cells and Mouse Microcirculation. Molecules, 24(19), 3509. https://doi.org/10.3390/molecules24193509