Abstract
Although sessile serrated adenoma/polyps (SSA/Ps) may arise through a pathway different from the traditional adenoma–carcinoma sequence, details of SSA/P tumorigenesis still remain unclear. Fusobacterium nucleatum (Fn) is frequently detected in colorectal cancer (CRC) tissues and may play a pivotal role in colorectal carcinogenesis. Here, we investigated the relationship between Fn and the β-catenin/REG Iα axis in SSA/Ps and their involvement in the proliferation of these lesions. Fn was detected in SSA/Ps by fluorescence in situ hybridization using a Fn-targeted probe, and expression of β-catenin, REG Iα and Ki67 was examined using immunohistochemistry. Sixteen of 30 SSA/P lesions (53.3%) were positive for Fn. Eighteen SSA/P lesions (60%) showed β-catenin immunoreactivity in the tumor cell nuclei. A significant majority of Fn-positive lesions showed nuclear expression of β-catenin (87.5%) and higher REG Iα scores and Ki67 labeling indices relative to Fn-negative lesions. The SSA/P lesions expressing β-catenin in nuclei had significantly higher REG Iα scores and Ki67 labeling indices than those expressing β-catenin on cytomembranes. The REG Iα score was positively correlated with the Ki67 labeling index in SSA/P lesions. The treatment with Wnt agonist SKL2001 promoted nuclear β-catenin translocation and enhanced REG Ia expression in Caco2 cells. Fn may play a role in the proliferation of SSA/P lesions through promotion of β-catenin nuclear translocation and REG Iα expression.
Keywords:
Fusobacterium nucleatum; sessile serrated adenoma; polyp; β-catenin; REG; Ki67; proliferation 1. Introduction
Although it is widely accepted that colorectal cancers (CRCs) arise from adenomas [1], the pathway responsible has recently been shown to be far from simple [2]. Among colorectal adenomas, sessile serrated adenoma/polyps (SSA/Ps) are classified as a subgroup exhibiting a specific saw-toothed colonic crypt morphology and genetic alterations such as BRAF mutation and microsatellite instability [3,4]. Furthermore, SSA/Ps lesions show high-labeling index for Ki67 expression [5], suggesting that those lesions have high ability in cell proliferation. These morphologic and genetic alterations are quite different from those in conventional adenomas, from which CRCs arise through accumulation of APC, KRAS and p53 mutations in multiple steps [1]. Thus, the molecular mechanism whereby CRCs arise and progress from adenomas is not fully clear. In addition to genetic alterations, the role of the gut microbiome in colorectal carcinogenesis has been highlighted; microbiome imbalance (dysbiosis) is closely associated with the development and progression of CRCs through promotion of chronic inflammatory conditions and production of carcinogenic metabolites [6]. Among various candidate pathogenic bacteria, Fusobacterium nucleatum (Fn) has been gathering the most attention, since numerous studies have reported that a higher abundance of Fn is associated with a more advanced stage, a higher risk of recurrence, and shorter survival in patients with CRC [7,8,9]. However, it is still debatable whether Fn directly plays a role in the pathogenesis of CRC patients although Fn may promote colorectal carcinogenesis by activating β-catenin signaling in vitro experiments [10,11].
The regenerating gene (REG) was first isolated from rat regenerating pancreatic islets [12], and since then, many REG-related genes have been isolated, currently constituting a family with multiple members (types I-IV) [13,14]. We and others have reported that REG family proteins play some roles in not only inflammatory [15,16,17,18] but also neoplastic [19,20,21] diseases of the gastrointestinal (GI) tract. In fact, REG Iα protein, as well as other REG family proteins [22,23,24], are suggested to exert a cell proliferative and/or anti-apoptotic effect in inflamed and neoplastic lesions in the GI tract [25,26,27]. In relation to the up-regulation of REG Iα expression in GI neoplasia, we have previously clarified that STAT3-associated cytokines play a pivotal role in REG Iα overexpression in gastric cancer cells [28]. However, the mechanism by which REG Iα overexpression is regulated in neoplastic lesions remains unclear. On the other hand, it is well known that β-catenin signaling is one of the major pathways of carcinogenesis in colorectal tumors, including SSA/Ps [29,30]. In this regard, it is interesting to speculate that β-catenin mutation and aberrant β-catenin expression may be linked to REG Iα overexpression in hepatocellular carcinoma, colorectal tumors, or salivary tumors [31,32]. In the present study, therefore, we focused on the relationship between Fn abundance and the β-catenin/REG Iα axis and investigated its significance for the proliferative ability of SSA/P lesions.
2. Results
2.1. Detection of Fusobacterium Nucleatum in SSA/P Lesions
The FISH signal for Fn was detected not only on the luminal surface but also the lamina propria of SSA/P lesions (Figure 1). We evaluated the density of the signal population (magnification ×200) in both SSA/P lesions and in the adjacent non-neoplastic regions. As the density in the non-neoplastic regions was 0.81 ± 0.15 (maximum 2.50), we considered any SSA/P lesion to be positive for Fn if the signal density was more than 3.0. Based on this definition, 16 of 30 SSA/P lesions (53.3%) were positive for Fn in the samples examined. None of the determined parameters—age, gender, tumor location, or endoscopic morphology—showed a significant relationship to Fn positivity.
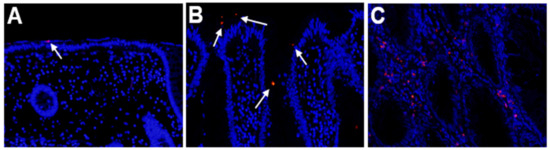
Figure 1.
Detection of Fusobacterium nucleatum (Fn) by fluorescence in situ hybridization. (A) Non-neoplastic colonic mucosa adjacent to a sessile serrated adenoma/polyp (SSA/P) lesion. (B,C) SSA/P lesions. The signals (red dots) for Fn are evident at the surface (B) or lamina propria (C) of the SSA/P lesions but are hardly evident in the non-neoplastic mucosa. Signals are indicated by white arrows.
2.2. Expression of REG Iα and β-Catenin in SSA/P Lesions
REG Iα was expressed in a few epithelial cells in non-neoplastic crypts in the colonic mucosa (Figure 2A). In SSA/P lesions, REG Iα was expressed mainly in goblet cells, and we scored its expression as described in materials and methods. All of the SSA/P lesions showed apparently higher scores for REG Iα expression than their adjacent non-neoplastic areas (Figure 2A–D). The REG Iα expression score was significantly higher in SSA/Ps on the right side of the colon (1.71 ± 0.11, p < 0.05) than in those on the left side of the colon (0.90 ± 0.31). None of the remaining parameters had any significant relationship to the REG Iα expression score.
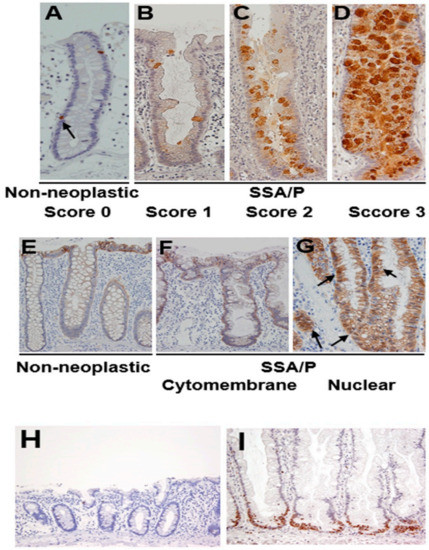
Figure 2.
Expression of REG Iα, β-catenin and Ki67 in SSA/P and its adjacent non-neoplastic mucosa in the colon. (A–D) Grading of REG Iα expression in SSA/P lesions and non-neoplastic colonic mucosa. (E–G) β-catenin expression pattern in SSA/P lesions and non-neoplastic colonic mucosa. Arrows indicate nuclear expression of β-catenin. (H,I) Expression of Ki67 in SSA/P lesions and non-neoplastic colonic mucosa.
Immunoreactivity for β-catenin was detected in the cytomembranes of non-neoplastic epithelial cells in the colonic mucosa (Figure 2E). In SSA/P lesions, β-catenin immunoreactivity was detected in not only cytomembranes (Figure 2F) but also the nuclei of serrated epithelial cells (Figure 2G). Twelve of the 30 SSA/P lesions (40%) showed a cytomembrane-type β-catenin staining pattern, whereas 18 of them (60%) showed nuclear-type immunostaining. None of the clinicopathological parameters examined had a significant relationship to the β-catenin expression pattern.
Ki67 was sparsely expressed in the lower portion of non-neoplastic crypts in the colonic mucosa (Figure 2H). On the other hand, Ki67 immunoreactivity was apparently increased, mainly at the bottom of serrated glands (Figure 2I). None of the clinicopathological parameters examined showed a significant relationship to the Ki67 labeling index.
2.3. Relationships among REG Iα/β-Catenin Expression, Fusobacterium Nucleatum Abundance, and Proliferative Activity in SSA/P Lesions
We investigated the relationship between Fn positivity and REG Iα/β-catenin expression in SSA/P lesions (Table 1). The REG Iα expression score was significantly higher in Fn-positive SSA/P lesions. Fn-positive SSA/P lesions showed a significant preponderance of nuclear-type β-catenin expression, whereas Fn-negative SSA/P lesions tended to show cytomembrane-type expression. In terms of cell proliferative ability, the Ki67 labeling index was significantly higher in Fn-positive than in negative SSA/P lesions.

Table 1.
Association between Fn presence and β-catenin, REG Iα, or Ki67 expression in SSA/P lesions.
We then investigated the correlation between REG Iα/β-catenin expression and the Ki67 labeling index. As shown in Figure 3A, the REG Iα expression score was significantly correlated with the Ki67 labeling index. In addition, SSA/P lesions with nuclear β-catenin expression showed a significantly higher Ki67 labeling index than lesions with cytomembrane expression (Figure 3B). These findings suggested that expression of REG Iα and nuclear β-catenin was linked to the proliferative ability of SSA/P lesions. It was also evident that SSA/P lesions with nuclear β-catenin expression had a significantly higher REG Iα score than lesions with cytomembrane expression (Figure 3C).
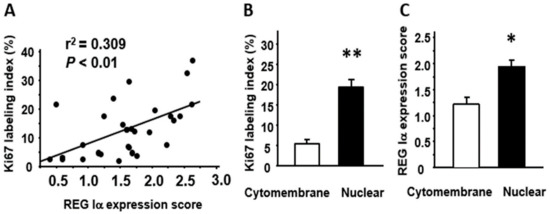
Figure 3.
Relationships among REG Iα, β-catenin, and Ki67 expression in SSA/P lesions. (A) Correlation between the REG Iα expression score and the Ki67 labeling index in SSA/P lesions. (B) Comparison of Ki67 labeling index between SSA/P lesions with cytomembrane-type β-catenin expression and those with nuclear-type expression. (C) Comparison of REG Iα expression score between SSA/P lesions with cytomembrane-type β-catenin expression and those with nuclear-type expression. Results are expressed as the mean ± SE. * p < 0.005, ** p < 0.001 vs. cytomembrane-type group.
2.4. Relationship among Histology, Tumor Size, and REG Iα/β-catenin/Ki67 Expression in SSA/P Lesions
Since β-catenin nuclear translocation may occur in SSA/P at the transition to dysplasia [29], we investigated the relationships among pathological features and REG Iα/β-catenin/Ki67 expression in SSA/P lesions. Subsequently, the SSA/Ps with dysplastic change significantly showed nuclear β-catenin expression and, moreover, had a significantly higher score for REG Iα expression (Table 2). In addition, those SSA/Ps tended to show higher Ki67 labeling index compared with ones without. Regarding the size of SSA/P, we found no significant relations to β-catenin (cytomembrane vs. nuclear: 23.4 ± 2.5 vs. 19.5 ± 1.8 mm), REG Iα (r2 = 0.02, p = 0.45), or Ki67 (r2 = 0.03, p = 0.88) expression in this study.
Table 2.
Association between dysplastic change and β-catenin, REG Iα or Ki67 expression in SSA/P lesions.
2.5. Effect of Wnt Signaling on Nuclear β-Catenin Translocation and REG Iα Expression in Caco2 Cells
When Caco2 cells were stimulated with Wnt agonist SKL2001, the immunoreactivity for β-catenin was detected not only at the cytomembrane but also in the cytoplasm and the nuclei in the cells, suggesting that nuclear β-catenin translocation was promoted in those cells (Figure 4A). Furthermore, the expression of REG Iα was significantly enhanced in Caco2 cells by the treatment with Wnt agonist SKL2001 (Figure 4B).
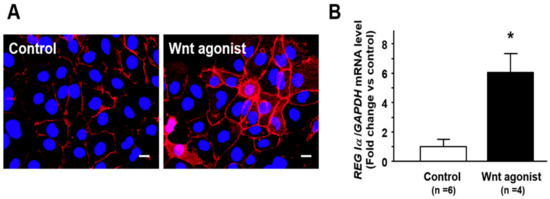
Figure 4.
Effect of Wnt agonist SKL2001 on nuclear β-catenin translocation and REG Iα expression in Caco2 cells. (A) Representative photograph showing nuclear β-catenin (red) translocation in Caco2 cells treated with Wnt agonist SKL2001. Caco2 cells (5 × 104) were cultured in 12-well plates for 24 h and then treated with Wnt agonist SKL2001 (40 mM) for 24 h. After stimulation, the cells were fixed with methanol for 10 min at room temperature, incubated with primary anti-β-catenin antibody for 1 h, followed by TRITC-labeled secondary anti-rabbit antibodies for 1 h, nuclear counterstained using Antifade Mountant with DAPI (blue) and observed under Olympus BX53 microscope. Bar = 20 mm. (B) Change in REG Iα expression in in Caco2 cells treated with Wnt agonist SKL2001. Results are expressed as the mean ±SE. * p < 0.05 vs. control group.
3. Discussion
The mechanism of CRC development has been largely studied by focusing on genetic alterations, but recent investigators have begun to recognize the role of the gut microbiome in this respect [6], similarly to the role of Helicobacter pylori in gastric cancer development [33]. So far, comprehensive analyses of the gut microbiome have identified several candidate bacteria that may play a role in the development of CRC, and among them, Fn has received special attention [7,8,9]. In the present study, we were able to detect the presence of Fn in approximately half of the SSA/P lesions we examined, in agreement with a previous report [34]. One limitation in this study was that we were unable to address the mechanism whereby Fn affects the carcinogenesis of CRC. However, since SSA/P lesions are known to show high proliferative ability [5], we investigated the relationship between Fn positivity and proliferative ability in SSA/P lesions and subsequently clarified that Fn-positive lesions had higher proliferative ability than Fn-negative lesions. In addition to proliferative ability, Fn infection appears to accelerate inflammation and DNA damage in colonic epithelial cells, and those accelerations may be regulated by a specific DNA glycosylase [35]. These findings suggest that not only cell proliferation, but also inflammation-associated DNA damage may be a key to understand the effect of Fn infection on the progression of malignant potential in SSA/P lesions.
Of note, β-catenin nuclear translocation is likely to occur in SSA/P lesions at the transition to dysplasia [29,36], being compatible with our obtained data in this study. This finding may suggest that nuclear β-catenin expression may be a marker of high risk of malignant progression in SSA/P lesions. Interestingly, recent studies have clarified that Fn is likely to promote CRC growth through the formation of a FadA-E-cadherin-annexin A1-β-catenin complex to activate the nuclear translocation of β-catenin [10,11]. Furthermore, another study has demonstrated that Fn may promote the nuclear translocation of β-catenin via a TLR4/P-PAK1 cascade in colorectal cancers [37]. In this respect, it was noteworthy in this study that Fn-positive SSA/P lesions showed significant nuclear immunoreactivity for β-catenin. Moreover, we found that SSA/P lesions with a nuclear β-catenin immunostaining pattern had a significantly higher Ki67 labeling index than lesions with cytomembrane immunostaining, indicating that SSA/P lesions with nuclear β-catenin expression had higher proliferative ability. As we demonstrated in this study, β-catenin is normally localized on the cytomembranes of non-neoplastic epithelial cells in the colonic mucosa. However, nuclear translocation of β-catenin is often observed in various tumors, and such translocated β-catenin acts as a transcriptional factor to regulate the expression of its target genes [38,39]. Although we cannot explain exactly why SSA/P lesions with a nuclear β-catenin immunostaining pattern have higher proliferative ability, we speculate that Fn-associated β-catenin nuclear translocation may play at least some role in the proliferation of SSA/P lesions. Moreover, it has been known that APC mutations and APC-related abnormalities are less common in SSA/Ps [40,41]. Conversely, this suggests that APC-unrelated β-catenin activation, such as Fn-associated ones, may play a pivotal role in progression of SSA/P lesions.
Vigorous mucin production is one characteristic of SSA/P lesions [41,42]. Of note, REG Iα protein is overexpressed in precancerous metaplasia and adenoma that actively express various mucin phenotypes [19,26,43]. Moreover, REG Iα protein functions as a growth and/or anti-apoptotic factor in gastrointestinal tumors [26,27,43], which prompted us to investigate REG Iα expression in SSA/P lesions. In non-neoplastic crypts, REG Iα is expressed in a few endocrine cells with an ovoid or pyramidal morphology, whereas all of the present SSA/P lesions apparently overexpressed REG Iα protein. Interestingly, the REG Iα expression score was positively correlated with the Ki67 labeling index in SSA/P lesions. This suggests that REG Iα expression is associated with the proliferative ability of these lesions, which would be reasonable in view of the known cell growth effect of REG Iα protein. However, one might be concerned whether REG Iα overexpression is specific in SSA/Ps that predominantly arise in the right-side colon. In this regard, a few studies have reported that a proportion of colonic conventional adenomas also overexpress REG Iα protein [44,45]. Therefore, we may have to investigate REG Iα expression in not only SSA/Ps but also conventional adenomas while analyzing the relationship between REG Iα expression and various clinicopathological features.
Finally, we investigated the relationship between β-catenin and REG Iα protein in SSA/P lesions and clarified for the first time that Fn positivity was positively correlated with not only nuclear β-catenin expression but also the REG Iα expression score. Moreover, it was noteworthy that SSA/P lesions with nuclear β-catenin expression had a significantly higher REG Iα expression score. These finding suggest that nuclear translocation of β-catenin may be linked to enhancement of REG Iα expression in SSA/P lesions. In this context, previous in vitro studies have shown that β-catenin signaling is responsible for REG Iα expression [46], and that β-catenin mutation may be linked to aberrant REG Iα overexpression in hepatocellular carcinoma [31]. Thus, since both β-catenin and REG Iα are certainly involved in the progression of CRCs, the β-catenin/REG Iα axis may play a pivotal role in the growth of SSA/P lesions. One limitation of the present study was that signaling from Fn to the β-catenin/REG Iα axis was not fully investigated. However, we preliminarily showed that nuclear β-catenin translocation by Wnt signaling activation may be closely associated with the enhancement of REG Iα expression in colon cancer cells in vitro. Interestingly, recent studies have suggested that Wnt signaling may be a key pathway for Fn-associated cell growth in colon cancer [11,47,48]. Thus, since REG Iα is able to function as a growth factor [25,26], we would like to speculate that Fn-associated β-catenin-nuclear translocation may play a role in the growth of SSA/P lesions, at least in part, via the growth-promoting effect of REG Iα protein.
In summary, we have demonstrated that nuclear β-catenin expression and REG Iα overexpression are simultaneously evident in Fn-positive SSA/P lesions and that these are commonly correlated with the proliferative ability in such lesions. Although we were unable to investigate the pathway involved in signaling from Fn to the β-catenin/REG Iα axis, the present findings at least suggest that Fn affects the proliferation of SSA/P lesions accompanied by promotion of β-catenin nuclear translocation and REG Iα expression.
4. Materials and Methods
4.1. Tissue Specimens and Clinicopathological Examination
Thirty patients with SSA/P who underwent endoscopic submucosal dissection at Hyogo College of Medicine Hospital between 2016 and 2019 were enrolled. The SSA/P lesions examined were collected by endoscopic submucosal dissection (n =16), endoscopic mucosal resection (n = 10) or polypectomy (n = 4). The tissue specimens obtained were fixed in 10% buffered formalin and embedded in paraffin, then cut into sections for pathological examination and immunohistochemistry. The characteristics of the patients and their lesions are listed in Table 3. The histological features of dysplastic change were assessed according to the previous descriptions [29,36]. This work was done with the approval of the Ethics Committee of Hyogo College of Medicine.
Table 3.
Clinicopathological features of patients with SSA/Ps.
4.2. Immunohistochemistry
Immunohistochemical staining for REG Iα, Ki67 and β-catenin was performed with an Envision Kit (Dako Agilent Technologies, Tokyo, Japan) as described previously [49], using anti-REG Iα antibody (dilution; 1:2000), anti-Ki67 antibody (Dako Agilent Technologies, dilution; 1:50), and anti-β-catenin antibody (Cell Signaling Technology, Danvers, MA, USA; dilution; 1:500). In brief, the rehydrated sections were treated by microwave heating for 20 min in 1×Dako REAL Target Retrieval Solution (Dako Agilent Technologies) and then preincubated with 0.3% H2O2 in methanol for 20 min at room temperature to quench endogenous peroxidase activity. The sections were then incubated with primary antibodies for 60 min at room temperature, washed in PBS, and incubated with horseradish peroxidase-conjugated secondary antibody for 30 min. The slides were visualized using 3,3′-diaminobenzidine tetrahydrochloride with 0.05% H2O2 for 3 min and then counterstained with Mayer’s hematoxylin.
4.3. Evaluation of Immunostaining
For evaluation of immunohistochemical expression, crypts that were well oriented perpendicularly from the bottom to the surface of the colorectal epithelium were selected. The expression of REG Iα was graded as previously described [50] with minor modification. Thus, it was scored according to the percentage of positive cells in a crypt as follows: score 0, a few cells; score 1, <10%; score 2, 10–50%; score 3, >50%. The Ki67 labeling index was expressed as the percentage of positive cells in a crypt. At least five different visual fields for each SSA/P lesion were observed, and the average REG Iα score and Ki67 labeling index were calculated. β-catenin immunoreactivity was detected in the cytomembranes and nuclei of neoplastic cells in SSA/P lesions. When an SSA/P lesion showed β-catenin immunoreactivity not only in the cytomembranes but also the nucleus, it was classified as the nuclear type, whereas if β-catenin immunoreactivity was evident in the cytomembranes but not in the nucleus, the lesion was classified as the cytomembrane type.
4.4. Fluorescence In Situ Hybridization (FISH)
The sequence of the Fn-targeted probe, FUS664 (Cy3 labeled), was 5′-CTTGTAGTT CCGC(C/T)TACCTC-3′-(Chromosome Science Labo Inc., Sapporo, Japan). The 16 rRNA-targeted oligonucleotide probe was obtained from probe-Base (http://www.microbial-ecology.net/probebase/) [34,51]. The tissue sections were deparaffinized and then hybridized with the Fn-targeted probe at 46 ℃ for 2 h in accordance with the manufacturer’s protocol. After hybridization, the slides were washed in buffer (40% formamide, 0.9 M NaCl, 0.01% SDS, 20 mM Tris-HCl) for 10 min, then in phosphate-buffered saline, and counterstained with DAPI (Thermo Fisher Scientific, Tokyo, Japan).
SSA/P lesions were observed using a fluorescence microscope (DP72; Olympus, Tokyo, Japan; magnification ×200) and fluorescent signals were recorded throughout each lesion (at least five different visual fields). The number of signal dots for Fn was counted in each field and the average was calculated. When the average Fn signal count was greater than 3.0/field, the lesion was considered to be positive.
4.5. Stimulation for Caco2 Cells by Wnt Agonist SKL2001
Human intestinal epithelial cell line Caco2 was obtained from ATCC (Manassas, VA, USA) and cultured in RPMI 1640 medium (Invitrogen, Carlsbad, CA, USA) with 10% fetal bovine serum (Biowest, Nuaillé, France) in a humidified incubator at 37 °C with an atmosphere of 5% CO2. The cells were treated with Wnt agonist SKL2001 (40 µM, Selleck, Houston, USA) for 24 h.
Total RNA was isolated from the cells and reverse-transcribed using oligo-dT primer (Applied Biosystems, Branchburg, NJ). Thereafter, real-time RT-PCR was performed as previously reported [18]. In brief, the following sets of primers for human REG Iα and glyceraldehydes-3-phosphate dehydrogenase (GAPDH) were prepared: human REG Iα 5′-CTAGAGGCAACTGGAAAATACATGTCT-3′ (sense), 5′-GTTGGAGAGATGGTCCGGTTT-3′ (antisense), human GAPDH 5′-GAGTCAACGGATTTGGTCGT-3′ (sense), 5′-TTGATTTTGGAGGGATCTCG-3′ (antisense). The intensity of the fluorescent dye was determined, and the expression levels of REG Iα mRNA were normalized to those of GAPDH mRNA.
The treated Caco2 cell were also subjected to immunostaining of β-catenin. Details are mentioned in the Figure 4 legend. Briefly, the cells were fixed with methanol, incubated with anti-β-catenin antibody (dilution 1:100; Cell Signaling), followed by TRITC-labeled secondary anti-rabbit antibodies (dilution 1:1000; DAKO), and counterstained using Antifade Mountant with DAPI (Life Technologies, Carlsbad, CA, USA).
4.6. Statistical Analysis
All values were expressed as the mean ± SE. Significance of differences between two animal groups was analyzed by Mann–Whitney U-test. Correlations between two parameters were assessed by linear regression analysis. Differences were considered to be significant at p < 0.05.
Author Contributions
Conceptualization, H.N. and H.F.; experimentation, H.N., H.F., X.W., N.E., and T.N.; methodology, H.N., H.F., X.W., N.E., and T.N.; data acquisition and analysis, H.N., H.F., X.W., N.E., and T.N.; resources, H.N., H.F., X.W., N.E., T.N., T.T., T.O., and S.H.; writing—original draft preparation, N.E. and H.F.; writing—review and editing, N.E., H.F., T.T., T.O., S.H., and H.M.; supervision, T.T., T.O., S.H., and H.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by Grants-in-aid for Scientific Research 17K09363 from the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of Hyogo College of Medicine (Protocol Identification Number: 3216).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study, and the opt out for this observational study was informed from Hyogo College of Medicine.
Data Availability Statement
The data presented in this study are available upon reasonable request from the corresponding author via hfukui@hyo-med.ac.jp.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Powell, S.M.; Zilz, N.; Beazer-Barclay, Y.; Bryan, T.M.; Hamilton, S.R.; Thibodeau, S.N.; Vogelstein, B.; Kinzler, K.W. APC mutations occur early during colorectal tumorigenesis. Nature 1992, 359, 235–237. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.J.; Yang, S.; Mack, C.; Xu, H.; Huang, C.S.; Mulcahy, E.; Amorosino, M.; Farraye, F.A. Comparison of microsatellite instability, CpG island methylation phenotype, BRAF and KRAS status in serrated polyps and traditional adenomas indicates separate pathways to distinct colorectal carcinoma end points. Am. J. Surg. Pathol. 2006, 30, 1491–1501. [Google Scholar] [CrossRef] [PubMed]
- Leggett, B.; Whitehall, V. Role of the serrated pathway in colorectal cancer pathogenesis. Gastroenterology 2010, 138, 2088–2100. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Kitamura, S.; Kimura, T.; Nakagawa, T.; Sogabe, M.; Miyamoto, H.; Muguruma, N.; Takayama, T. Clinicopathological characteristics of serrated polyps as precursors to colorectal cancer: Current status and management. J. Gastroenterol. Hepatol. 2017, 32, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Shida, Y.; Ichikawa, K.; Fujimori, T.; Fujimori, Y.; Tomita, S.; Fujii, T.; Sano, Y.; Oda, Y.; Goto, H.; Ohta, A.; et al. Differentiation between sessile serrated adenoma/polyp and non-sessile serrated adenoma/polyp in large hyper plastic polyp: A Japanese collaborative study. Mol. Clin. Oncol. 2013, 1, 53–58. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Song, M.; Chan, A.T.; Sun, J. Influence of the gut microbiome, diet, and environment on risk of colorectal cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Castellarin, M.; Warren, R.L.; Freeman, J.D.; Dreolini, L.; Krzywinski, M.; Strauss, J.; Barnes, R.; Watson, P.; Allen-Vercoe, E.; Moore, R.A.; et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012, 22, 299–306. [Google Scholar] [CrossRef]
- Flanagan, L.; Schmid, J.; Ebert, M.; Soucek, P.; Kunicka, T.; Liska, V.; Bruha, J.; Neary, P.; Dezeeuw, N.; Tommasino, M.; et al. Fusobacterium nucleatum associates with stages of colorectal neoplasia development, colorectal cancer and disease outcome. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1381–1390. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium nucleatum promotes colorectal cancer by inducing Wnt/β-catenin modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef]
- Terazono, K.; Yamamoto, H.; Takasawa, S.; Shiga, K.; Yonemura, Y.; Tochino, Y.; Okamoto, H. A novel gene activated in regenerating islets. J. Biol. Chem. 1988, 263, 2111–2114. [Google Scholar] [CrossRef]
- Unno, M.; Yonekura, H.; Nakagawara, K.; Watanabe, T.; Miyashita, H.; Moriizumi, S.; Okamoto, H.; Itoh, T.; Teraoka, H. Structure, chromosomal localization, and expression of mouse reg genes, reg I and reg II. A novel type of reg gene, reg II, exists in the mouse genome. J. Biol. Chem. 1993, 268, 15974–15982. [Google Scholar] [CrossRef]
- Takasawa, S. Regenerating gene (REG) product and its potential clinical usage. Expert Opin. Ther. Targets 2016, 20, 541–550. [Google Scholar] [CrossRef]
- Sekikawa, A.; Fukui, H.; Suzuki, K.; Karibe, T.; Fujii, S.; Ichikawa, K.; Tomita, S.; Imura, J.; Shiratori, K.; Chiba, T.; et al. Involvement of the IL-22/REG Iα axis in ulcerative colitis. Lab. Invest. 2010, 90, 496–505. [Google Scholar] [CrossRef]
- Van Beelen Granlund, A.; Østvik, A.E.; Brenna, Ø.; Torp, S.H.; Gustafsson, B.I.; Sandvik, A.K. REG gene expression in inflamed and healthy colon mucosa explored by in situ hybridization. Cell Tissue Res. 2013, 352, 639–646. [Google Scholar] [CrossRef]
- Sun, C.; Fukui, H.; Hara, K.; Kitayama, Y.; Eda, H.; Yang, M.; Yamagishi, H.; Tomita, T.; Oshima, T.; Watari, J.; et al. Expression of Reg family genes in the gastrointestinal tract of mice treated with indomethacin. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G736–G744. [Google Scholar] [CrossRef]
- Xu, X.; Fukui, H.; Ran, Y.; Wang, X.; Inoue, Y.; Ebisudani, N.; Nishimura, H.; Tomita, T.; Oshima, T.; Watari, J.; et al. The link between type III Reg and STAT3-associated cytokines in inflamed colonic tissues. Mediat. Inflamm. 2019, 2019, 7859460. [Google Scholar] [CrossRef]
- Yamagishi, H.; Fukui, H.; Sekikawa, A.; Kono, T.; Fujii, S.; Ichikawa, K.; Tomita, S.; Imura, J.; Hiraishi, H.; Chiba, T.; et al. Expression profile of REG family proteins REG Iα and REG IV in advanced gastric cancer: Comparison with mucin phenotype and prognostic markers. Mod. Pathol. 2009, 22, 906–913. [Google Scholar] [CrossRef]
- Zheng, H.C.; Sugawara, A.; Okamoto, H.; Takasawa, S.; Takahashi, H.; Masuda, S.; Takano, Y. Expression profile of the REG gene family in colorectal carcinoma. J. Histochem. Cytochem. 2011, 59, 106–115. [Google Scholar] [CrossRef]
- Sekikawa, A.; Fukui, H.; Zhang, X.; Maruo, T.; Tsumura, T.; Okabe, Y.; Wakasa, T.; Osaki, Y.; Chiba, T.; Tomita, T.; et al. REG Iα is a biomarker for predicting response to chemotherapy with S-1 plus cisplatin in patients with unresectable stage IV gastric cancer. Br. J. Cancer 2013, 108, 395–401. [Google Scholar] [CrossRef]
- Bishnupuri, K.S.; Luo, Q.; Korzenik, J.R.; Henderson, J.O.; Houchen, C.W.; Anant, S.; Dieckgraefe, B.K. Dysregulation of Reg gene expression occurs early in gastrointestinal tumorigenesis and regulates anti-apoptotic genes. Cancer Biol. Ther. 2006, 5, 1714–1720. [Google Scholar] [CrossRef]
- Nanakin, A.; Fukui, H.; Fujii, S.; Sekikawa, A.; Kanda, N.; Hisatsune, H.; Seno, H.; Konda, Y.; Fujimori, T.; Chiba, T. Expression of the REG IV gene in ulcerative colitis. Lab. Invest. 2007, 87, 304–314. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Wang, J.; Guo, J. Role of regenerating islet-derived protein 3A in gastrointestinal cancer. Front. Oncol. 2019, 9, 1449. [Google Scholar] [CrossRef]
- Fukui, H.; Kinoshita, Y.; Maekawa, T.; Okada, A.; Waki, S.; Hassan, S.; Okamoto, H.; Chiba, T. Regenerating gene protein may mediate gastric mucosal proliferation induced by hypergastrinemia in rats. Gastroenterology 1998, 115, 1483–1493. [Google Scholar] [CrossRef]
- Sekikawa, A.; Fukui, H.; Fujii, S.; Takeda, J.; Nanakin, A.; Hisatsune, H.; Seno, H.; Takasawa, S.; Okamoto, H.; Fujimori, T.; et al. REG Iα protein may function as a trophic and/or anti-apoptotic factor in the development of gastric cancer. Gastroenterology 2005, 128, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Sekikawa, A.; Fukui, H.; Fujii, S.; Nanakin, A.; Kanda, N.; Uenoyama, Y.; Sawabu, T.; Hisatsune, H.; Kusaka, T.; Ueno, S.; et al. Possible role of REG Iα protein in ulcerative colitis and colitic cancer. Gut 2005, 54, 1437–1444. [Google Scholar] [CrossRef] [PubMed]
- Sekikawa, A.; Fukui, H.; Fujii, S.; Ichikawa, K.; Tomita, S.; Imura, J.; Chiba, T.; Fujimori, T. REG Iα protein mediates an anti-apoptotic effect of STAT3 signaling in gastric cancer cells. Carcinogenesis 2008, 29, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Mitomi, H.; Saito, T.; Takahashi, M.; Sakamoto, N.; Fukui, N.; Yao, T.; Watanabe, S. Distinct WNT/β-catenin signaling activation in the serrated neoplasia pathway and the adenoma-carcinoma sequence of the colorectum. Mod. Pathol. 2015, 28, 146–158. [Google Scholar] [CrossRef]
- Nourbakhsh, M.; Mansoor, A.; Koro, K.; Zhang, Q.; Minoo, P. Expression profiling reveals involvement of Wnt pathway in the malignant progression of sessile serrated adenomas. Am. J. Pathol. 2019, 189, 1732–1743. [Google Scholar] [CrossRef]
- Cavard, C.; Terris, B.; Grimber, G.; Christa, L.; Audard, V.; Radenen-Bussiere, B.; Simon, M.T.; Renard, C.A.; Buendia, M.A.; Perret, C. Overexpression of regenerating islet-derived 1 alpha and 3 alpha genes in human primary liver tumors with beta-catenin mutations. Oncogene 2006, 25, 599–608. [Google Scholar] [CrossRef]
- Hakata, Y.; Fukui, H.; Sekikawa, A.; Yamagishi, H.; Ichikawa, K.; Tomita, S.; Imura, J.; Kawamata, H.; Imai, Y.; Fujimori, T. Expression of β-catenin and REG Iα in relation to cell proliferative ability in salivary gland tumors. Exp. Ther. Med. 2010, 1, 437–443. [Google Scholar] [CrossRef]
- Sayed, I.M.; Sahan, A.Z.; Venkova, T.; Chakraborty, A.; Mukhopadhyay, D.; Bimczok, D.; Beswick, E.J.; Reyes, V.E.; Pinchuk, I.; Sahoo, D.; et al. Helicobacter pylori infection downregulates the DNA glycosylase NEIL2, resulting in increased genome damage and inflammation in gastric epithelial cells. J. Biol. Chem. 2020, 295, 11082–11098. [Google Scholar] [CrossRef]
- Yu, J.; Chen, Y.; Fu, X.; Zhou, X.; Peng, Y.; Shi, L.; Chen, T.; Wu, Y. Invasive Fusobacterium nucleatum may play a role in the carcinogenesis of proximal colon cancer through the serrated neoplasia pathway. Int. J. Cancer 2016, 139, 1318–1326. [Google Scholar] [CrossRef]
- Sayed, I.M.; Chakraborty, A.; Abd El-Hafeez, A.A.; Sharma, A.; Sahan, A.Z.; Huang, W.J.M.; Sahoo, D.; Ghosh, P.; Hazra, T.K.; Das, S. The DNA glycosylase NEIL2 suppresses Fusobacterium-infection-induced inflammation and DNA damage in colonic epithelial cells. Cells 2020, 9, 1980. [Google Scholar] [CrossRef]
- Fujita, K.; Yamamoto, H.; Matsumoto, T.; Hirahashi, M.; Gushima, M.; Kishimoto, J.; Nishiyama, K.; Taguchi, T.; Yao, T.; Oda, Y. Sessile serrated adenoma with early neoplastic progression: A clinicopathologic and molecular study. Am. J. Surg. Pathol. 2011, 35, 295–304. [Google Scholar] [CrossRef]
- Chen, Y.; Peng, Y.; Yu, J.; Chen, T.; Wu, Y.; Shi, L.; Li, Q.; Wu, J.; Fu, X. Invasive Fusobacterium nucleatum activates beta-catenin signaling in colorectal cancer via a TLR4/P-PAK1 cascade. Oncotarget 2017, 8, 31802–31814. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Vishnoi, K.; Viswakarma, N.; Rana, A.; Rana, B. Transcription factors in cancer development and therapy. Cancers 2020, 12, 2296. [Google Scholar] [CrossRef]
- Yachida, S.; Mudali, S.; Martin, S.A.; Montgomery, E.A.; Iacobuzio-Donahue, C.A. Beta-catenin nuclear labeling is a common feature of sessile serrated adenomas and correlates with early neoplastic progression after BRAF activation. Am. J. Surg. Pathol. 2009, 33, 1823–1832. [Google Scholar] [CrossRef]
- Murakami, T.; Akazawa, Y.; Yatagai, N.; Hiromoto, T.; Sasahara, N.; Saito, T.; Sakamoto, N.; Nagahara, A.; Yao, T. Molecular characterization of sessile serrated adenoma/polyps with dysplasia/carcinoma based on immunohistochemistry, next-generation sequencing, and microsatellite instability testing: A case series study. Diagn. Pathol. 2018, 13, 88. [Google Scholar] [CrossRef]
- Murakami, T.; Sakamoto, N.; Nagahara, A. Clinicopathological features, diagnosis, and treatment of sessile serrated adenoma/polyp with dysplasia/carcinoma. J. Gastroenterol. Hepatol. 2019, 34, 1685–1695. [Google Scholar] [CrossRef]
- Chen, Z.; Downing, S.; Tzanakakis, E.S. Four decades after the discovery of regenerating islet-derived (Reg) proteins: Current understanding and challenges. Front. Cell Dev. Biol. 2019, 7, 235. [Google Scholar] [CrossRef]
- Zenilman, M.E.; Kim, S.; Levine, B.A.; Lee, C.; Steinberg, J.J. Ectopic expression of reg protein: A marker of colorectal mucosa at risk for neoplasia. J. Gastrointest. Surg. 1997, 1, 194–202. [Google Scholar] [CrossRef]
- Rechreche, H.; Montalto, G.; Mallo, G.V.; Vasseur, S.; Marasa, L.; Soubeyran, P.; Dagorn, J.C.; Iovanna, J.L. pap, reg I alpha and reg I beta mRNAs are concomitantly up-regulated during human colorectal carcinogenesis. Int. J. Cancer 1999, 81, 688–694. [Google Scholar] [CrossRef]
- Jing, D.; Kehoe, D.E.; Tzanakakis, E.S. Expression of Reg family proteins in embryonic stem cells and its modulation by Wnt/beta-catenin signaling. Stem Cells Dev. 2010, 19, 1307–1319. [Google Scholar] [CrossRef]
- Lin, R.; Han, C.; Ding, Z.; Shi, H.; He, R.; Liu, J.; Qian, W.; Zhang, Q.; Fu, X.; Deng, X.; et al. Knock down of BMSC-derived Wnt3a or its antagonist analogs attenuate colorectal carcinogenesis induced by chronic Fusobacterium nucleatum infection. Cancer Lett. 2020, 495, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, J.; Yu, T.; Fang, X.; Lou, L.; Xin, S.; Ji, L.; Jiang, F.; Lou, Y. Fusobacterium nucleatum promotes the progression of colorectal cancer through Cdk5-activated Wnt/β-catenin signaling. Front. Microbiol. 2021, 11, 545251. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.; Fukui, H.; Sun, C.; Kitayama, Y.; Eda, H.; Yamasaki, T.; Kondo, T.; Tomita, T.; Oshima, T.; Watari, J.; et al. Effect of REG Iα protein on angiogenesis in gastric cancer tissues. Oncol. Rep. 2015, 33, 2183–2189. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, K.; Fujimori, T.; Yamaguchi, T.; Ichikawa, K.; Tomita, S.; Sugai, T.; Imura, J.; Ohkura, Y.; Yao, T.; Fujii, S.; et al. Overexpression of regenerating gene Iα appears to reflect aberration of crypt cell compartmentalization in sessile serrated adenoma/polyps of the colon. Diagn. Pathol. 2013, 8, 187. [Google Scholar] [CrossRef]
- Loy, A.; Arnold, R.; Tischler, P.; Rattei, T.; Wagner, M.; Horn, M. ProbeCheck-a central resource for evaluating oligonucleotide probe coverage and specificity. Environ. Microbiol. 2008, 10, 2894–2898. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).