
Evaluation of the Inhibitory Potential of Apigenin and Related Flavonoids on Various Proteins Associated with Human Diseases Using AutoDock

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
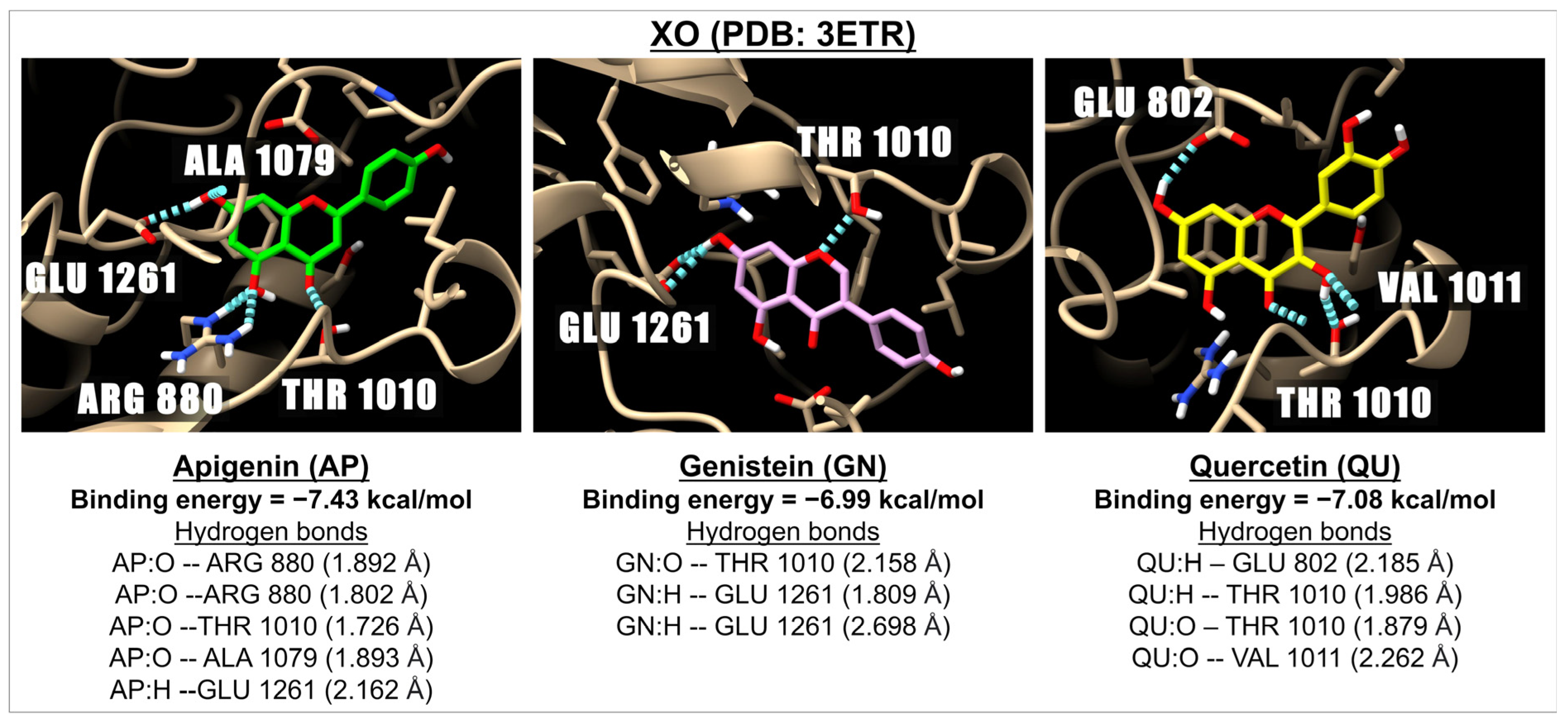
2.1. Binding Energy of Apigenin and Related Compounds with Proteins Known to Be Involved in Oxidative Stress
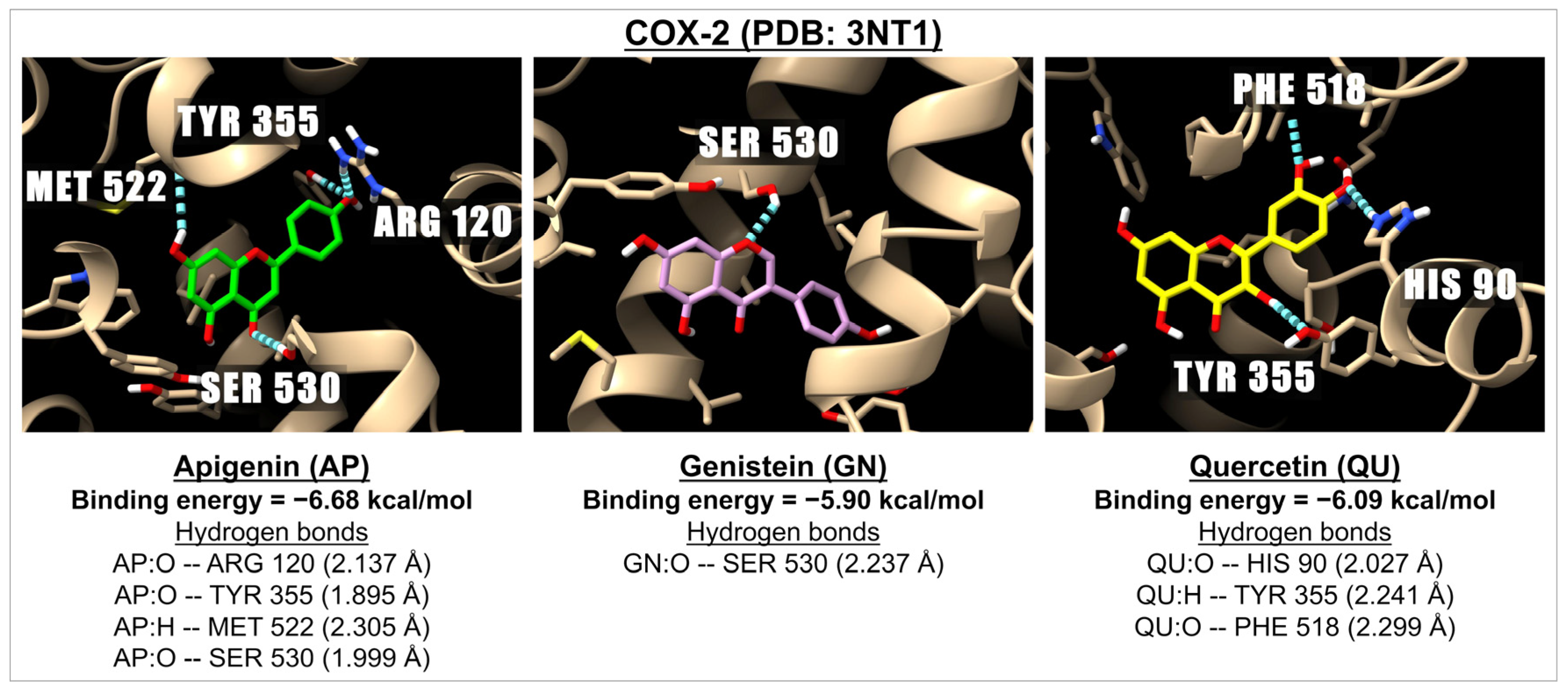
2.2. Binding Energies of Apigenin and Related Compounds with Proteins Known to Be Involved in Inflammation
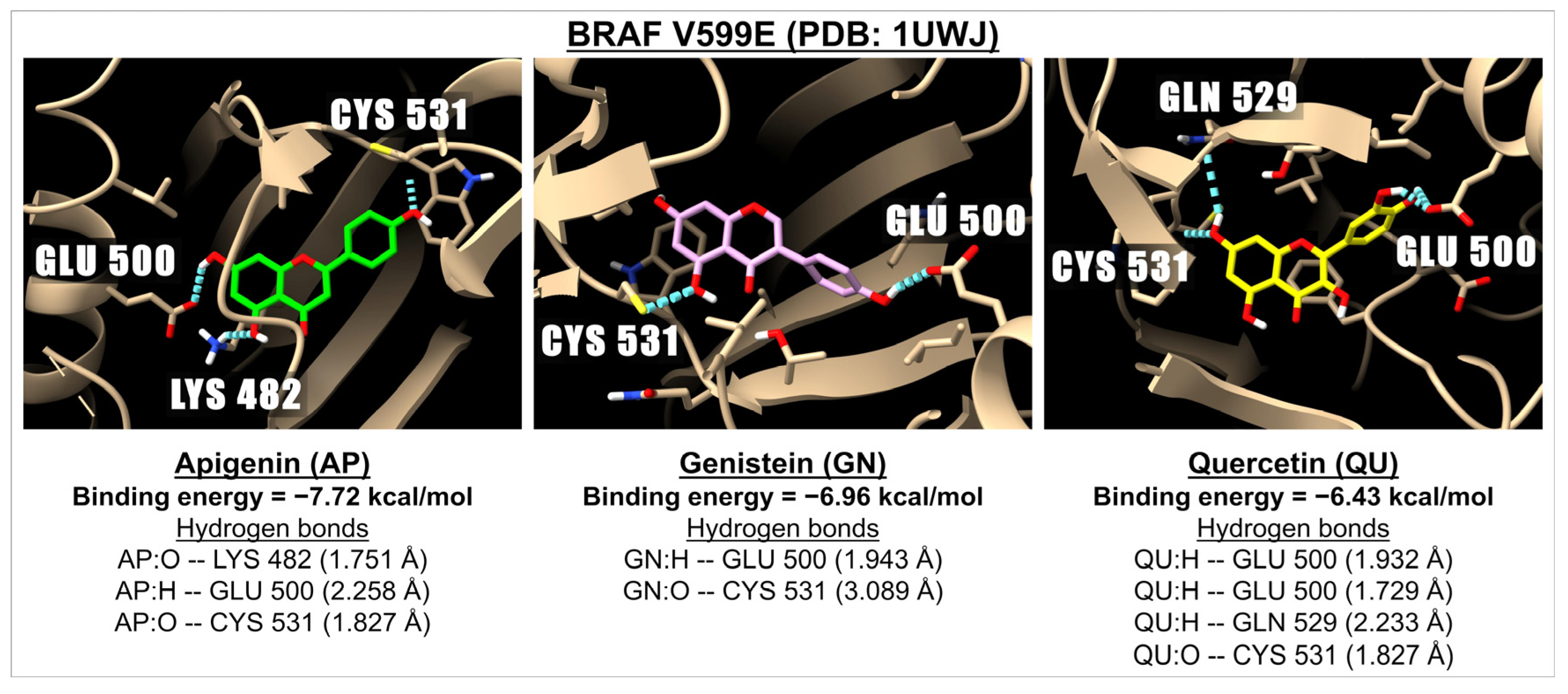
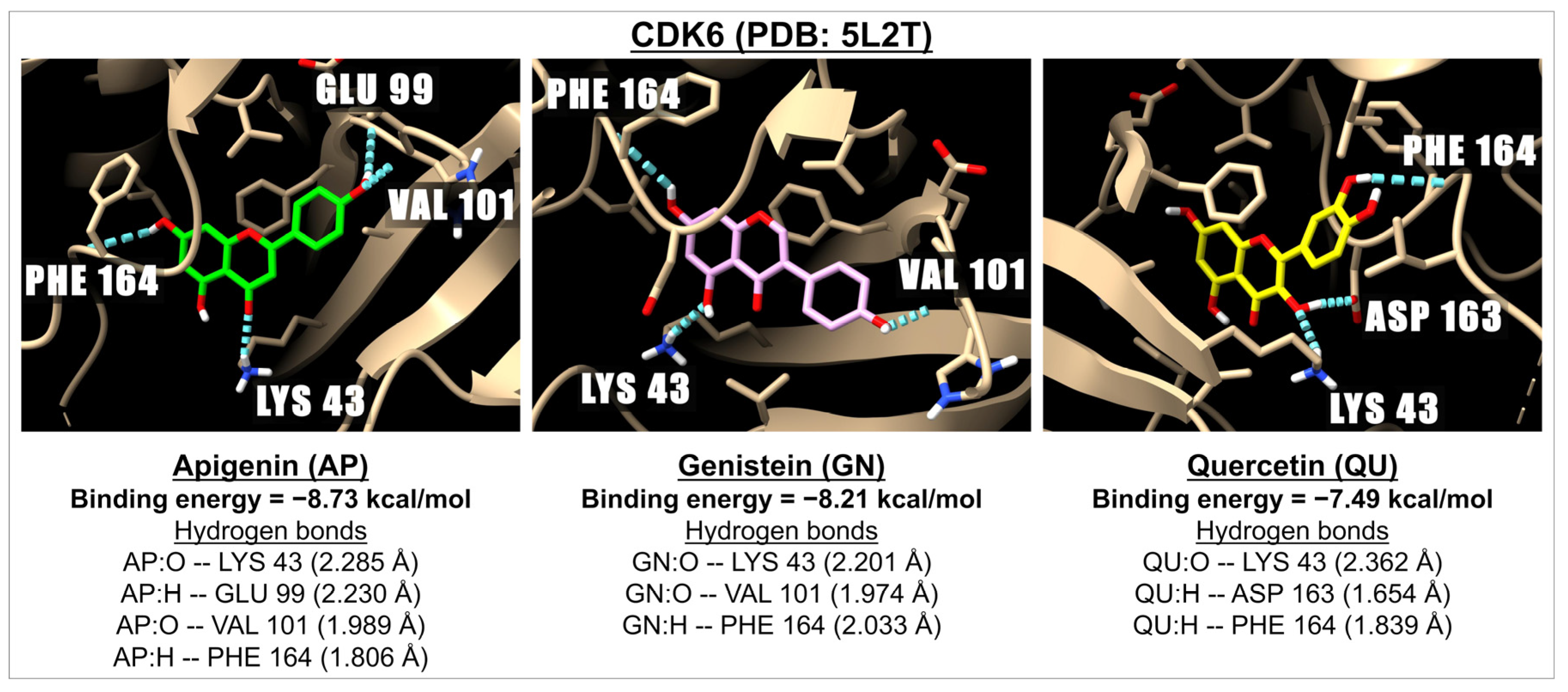
2.3. Binding Energy of Apigenin and Related Compounds with Proteins Known to Be Involved in Carcinogenesis
2.4. Binding Energies and Inhibition Constants (Ki) of Apigenin and Related Compounds with Proteins Known to Be Involved in Bacterial Infection
3. Discussion
4. Materials and Methods
4.1. Selection of Ligands (Flavonoids)
4.2. Ligand Preparation
4.3. Receptor Preparation
4.4. Molecular Docking
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.B.; Novellino, E.; et al. The therapeutic potential of apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Important flavonoids and their role as a therapeutic agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef] [PubMed]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Futur. J. Pharm. Sci. 2021, 7, 25. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, Z.; Luo, Y.; Li, X.; Huang, G.; Chen, H.; Li, A.; Qin, S. The role of flavonoids in the osteogenic differentiation of mesenchymal stem Ccls. Front. Pharmacol. 2022, 13, 849513. [Google Scholar] [CrossRef]
- Moonkum, N.; Chaichana, A.; Kantakam, P.; Malimart, C.; Piyachon, C.; Chananpanich, N.; Mankhetkorn, S. Siamois polyphenols as circulating endogenous stem cell regulator: Primordial source for repair and regeneration of tissue In Vivo. Curr. Biomark. 2018, 8, 1–16. [Google Scholar] [CrossRef]
- Peanlikhit, T.; Honikel, L.; Zimmerman, T.; Rithidech, K. Countermeasure efficacy of apigenin for silicon-ion-induced early damage in blood and bone marrow of exposed C57BL/6J mice. Life Sci. Space Res. 2022, 35, 44–52. [Google Scholar] [CrossRef]
- Siddiqui, N.; Abidin, L.; Nisar, N.; Ahmad, I.; Siddiqui, A.N. Flavonoids Targeting Cancer Stem Cells: A Paradigm to Anticancer Efficacy. In Polyphenols-Based Nanotherapeutics for Cancer Management; Tabrez, S., Imran Khan, M., Eds.; Springer: Singapore, 2021; pp. 239–287. [Google Scholar]
- Meerson, A.; Khatib, S.; Mahajna, J. Natural products targeting cancer stem cells for augmenting cancer therapeutics. Int. J. Mol. Sci. 2021, 22, 13044. [Google Scholar] [CrossRef]
- Rithidech, K.N.; Tungjai, M.; Whorton, E.B. Protective effect of apigenin on radiation-induced chromosomal damage in human lymphocytes. Mutat. Res. 2005, 585, 96–104. [Google Scholar] [CrossRef]
- Rithidech, K.N.; Tungjai, M.; Reungpatthanaphong, P.; Honikel, L.; Simon, S.R. Attenuation of oxidative damage and inflammatory responses by apigenin given to mice after irradiation. Mutat. Res. 2012, 749, 29–38. [Google Scholar] [CrossRef]
- Begum, N.; Rajendra Prasad, N.; Kanimozhi, G.; Agilan, B. Apigenin prevents gamma radiation-induced gastrointestinal damages by modulating inflammatory and apoptotic signalling mediators. Nat. Prod. Res. 2022, 36, 1631–1635. [Google Scholar] [CrossRef]
- Liu, D.; Peng, R.; Chen, Z.; Yu, H.; Wang, S.; Dong, S.; Li, W.; Shao, W.; Dai, J.; Li, F.; et al. The protective effects of apigenin against radiation-induced intestinal injury. Dose Response 2022, 20, 15593258221113791. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Shukla, S.; Gupta, S. Apigenin and cancer chemoprevention: Progress, potential and promise (review). Int. J. Oncol. 2007, 30, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Saeed, M.; Kadioglu, O.; Khalid, H.; Sugimoto, Y.; Efferth, T. Activity of the dietary flavonoid, apigenin, against multidrug-resistant tumor cells as determined by pharmacogenomics and molecular docking. J. Nutr. Biochem. 2015, 26, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yu, Y.; Lv, H.; Zhang, H.; Liang, T.; Zhou, G.; Huang, L.; Tian, Y.; Liang, W. Apigenin in cancer therapy: From mechanism of action to nano-therapeutic agent. Food Chem. Toxicol. 2022, 168, 113385. [Google Scholar] [CrossRef]
- DeRango-Adem, E.F.; Blay, J. Does Oral Apigenin Have Real Potential for a Therapeutic Effect in the Context of Human Gastrointestinal and Other Cancers? Front. Pharmacol. 2021, 12, 681477. [Google Scholar] [CrossRef]
- Adel, M.; Zahmatkeshan, M.; Akbarzadeh, A.; Rabiee, N.; Ahmadi, S.; Keyhanvar, P.; Rezayat, S.M.; Seifalian, A.M. Chemotherapeutic effects of Apigenin in breast cancer: Preclinical evidence and molecular mechanisms; enhanced bioavailability by nanoparticles. Biotechnol. Rep. 2022, 34, e00730. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Bakhoda, M.R.; Bahmanpour, Z.; Ilkhani, K.; Zarrabi, A.; Makvandi, P.; Khan, H.; Mazaheri, S.; Darvish, M.; Mirzaei, H. Apigenin as tumor suppressor in Cancers: Biotherapeutic activity, nanodelivery, and mechanisms with emphasis on pancreatic cancer. Front. Chem. 2020, 8, 829. [Google Scholar] [CrossRef]
- Abid, R.; Ghazanfar, S.; Farid, A.; Sulaman, S.M.; Idrees, M.; Amen, R.A.; Muzammal, M.; Shahzad, M.K.; Mohamed, M.O.; Khaled, A.A.; et al. Pharmacological properties of 4′, 5, 7-trihydroxyflavone (Apigenin) and its impact on cell signaling pathways. Molecules 2022, 27, 4304. [Google Scholar] [CrossRef]
- Begum, N.; Prasad, N.R. Apigenin, a dietary antioxidant, modulates gamma radiation-induced oxidative damages in human peripheral blood lymphocytes. Biomed. Prev. Nutr. 2012, 2, 16–24. [Google Scholar] [CrossRef]
- Tong, X.; Van Dross, R.T.; Abu-Yousif, A.; Morrison, A.R.; Pelling, J.C. Apigenin prevents UVB-induced cyclooxygenase 2 expression: Coupled mRNA stabilization and translational inhibition. Mol. Cell. Biol. 2007, 27, 283–296. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Afzal, M. Antigenotoxic effect of apigenin against mitomycin C induced genotoxic damage in mice bone marrow cells. Food Chem. Toxicol. 2009, 47, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Siddique, Y.H.; Ara, G.; Beg, T.; Afzal, M. Anticlastogenic effect of apigenin in human lymphocytes treated with ethinylestradiol. Fitoterapia 2010, 81, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Thayalan, K.; Prasad, N. Apigenin protects gamma-radiation induced oxidative stress, hematological changes and animal survival in whole body irradiated Swiss albino mice. J. Nutr. Pharmacol. Neuro. Dis. 2012, 2, 45–52. [Google Scholar] [CrossRef]
- Rithidech, K.N.; Peanlikhit, T.; Honikel, L.; Li, J.; Liu, J.; Karakach, T.; Zimmerman, T.; Welsh, J. Consumption of apigenin prevents radiation-induced gut dysbiosis in male C57BL/6J mice exposed to silicon ions. Radiat. Res. 2024, 201, 317–329. [Google Scholar] [CrossRef]
- Bugel, S.M.; Bonventre, J.A.; Tanguay, R.L. Comparative developmental toxicity of flavonoids using an integrative zebrafish system. Toxicol. Sci. 2016, 154, 55–68. [Google Scholar] [CrossRef]
- Cvetanović, A. Chapter 24—Apigenin. In A Centum of Valuable Plant Bioactives; Mushtaq, M., Anwar, F., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 545–562. [Google Scholar]
- El Shoubaky, G.A.; Abdel-Daim, M.M.; Mansour, M.H.; Salem, E.A. Isolation and identification of a flavone apigenin from marine red alga Acanthophora spicifera with antinociceptive and anti-Inflammatory activities. J. Exp. Neurosci. 2016, 10, 21–29. [Google Scholar] [CrossRef]
- Agu, P.C.; Afiukwa, C.A.; Orji, O.U.; Ezeh, E.M.; Ofoke, I.H.; Ogbu, C.O.; Ugwuja, E.I.; Aja, P.M. Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management. Sci. Rep. 2023, 13, 13398. [Google Scholar] [CrossRef]
- Meng, X.Y.; Zhang, H.X.; Mezei, M.; Cui, M. Molecular docking: A powerful approach for structure-based drug discovery. Curr. Comput. Aided Drug Des. 2011, 7, 146–157. [Google Scholar] [CrossRef]
- Pinzi, L.; Rastelli, G. Molecular docking: Shifting paradigms in drug discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef]
- Ivanova, L.; Karelson, M. The impact of Software used and the type of target protein on molecular docking accuracy. Molecules 2022, 27, 9041. [Google Scholar] [CrossRef]
- Terefe, E.M.; Ghosh, A. Molecular Docking, Validation, Dynamics Simulations, and Pharmacokinetic Prediction of Phytochemicals Isolated from Croton dichogamus Against the HIV-1 Reverse Transcriptase. Bioinform. Biol. Insights 2022, 16, 11779322221125605. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nguyen, T.H.; Pham, T.N.H.; Huy, N.T.; Bay, M.V.; Pham, M.Q.; Nam, P.C.; Vu, V.V.; Ngo, S.T. Autodock Vina adopts more accurate binding poses but autodock4 forms better binding affinity. J. Chem. Inf. Model. 2020, 60, 204–211. [Google Scholar] [CrossRef]
- Zhu, C.; Guan, X.; Zhang, X.; Luan, X.; Song, Z.; Cheng, X.; Zhang, W.; Qin, J.J. Targeting KRAS mutant cancers: From druggable therapy to drug resistance. Mol. Cancer 2022, 21, 159. [Google Scholar] [CrossRef]
- Prior, I.A.; Lewis, P.D.; Mattos, C. A comprehensive survey of Ras mutations in cancer. Cancer Res. 2012, 72, 2457–2467. [Google Scholar] [CrossRef]
- Hallin, J.; Bowcut, V.; Calinisan, A.; Briere, D.M.; Hargis, L.; Engstrom, L.D.; Laguer, J.; Medwid, J.; Vanderpool, D.; Lifset, E.; et al. Anti-tumor efficacy of a potent and selective non-covalent KRASG12D inhibitor. Nat. Med. 2022, 28, 2171–2182. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Maitra, A.; Choudhury, S. Selumetinib: The first ever approved drug for neurofibromatosis-1 related inoperable plexiform neurofibroma. Curr. Med. Res. Opin. 2021, 37, 789–794. [Google Scholar] [CrossRef]
- Solares, I.; Viñal, D.; Morales-Conejo, M.; Rodriguez-Salas, N.; Feliu, J. Novel molecular targeted therapies for patients with neurofibromatosis type 1 with inoperable plexiform neurofibromas: A comprehensive review. ESMO Open 2021, 6, 100223. [Google Scholar] [CrossRef]
- Li, D.; Wang, L.; Jing, Y.; Jiang, B.; Zhao, L.; Miao, Y.; Xin, S.; Ge, C. Exploring Molecular Targets and Mechanisms of Apigenin in the Treatment of Papillary Thyroid Carcinoma Based on Network Pharmacology and Molecular Docking Analysis. Nat. Product Commun. 2022, 17, 1934578X221135435. [Google Scholar] [CrossRef]
- Keerthanaa, T.; Boobalan, S.; Kamalanathan, D.; Karunakaran, G.; Sudha, K.G.; Aarthi, M.; Prasanna Rajeshkumar, M. Elicitation of apigenin in green leafy vegetable plants and its molecular docking evaluation for effective anticancer applications. Plant Cell Tissue Organ Cult. 2022, 150, 459–478. [Google Scholar] [CrossRef]
- Kashyap, P.; Thakur, M.; Singh, N.; Shikha, D.; Kumar, S.; Baniwal, P.; Yadav, Y.S.; Sharma, M.; Sridhar, K.; Inbaraj, B.S. In Silico evaluation of natural elavonoids as a potential inhibitor of Coronavirus Disease. Molecules 2022, 27, 6374. [Google Scholar] [CrossRef]
- Rathod, S.; Shinde, K.; Porlekar, J.; Choudhari, P.; Dhavale, R.; Mahuli, D.; Tamboli, Y.; Bhatia, M.; Haval, K.P.; Al-Sehemi, A.G.; et al. Computational Exploration of Anti-cancer Potential of Flavonoids against Cyclin-Dependent Kinase 8: An In Silico Molecular Docking and Dynamic Approach. ACS Omega 2023, 8, 391–409. [Google Scholar] [CrossRef]
- Essa, A.F.; Teleb, M.; El-Kersh, D.M.; El Gendy, A.E.-N.G.; Elshamy, A.I.; Farag, M.A. Natural acylated flavonoids: Their chemistry and biological merits in context to molecular docking studies. Phytochem. Rev. 2022, 22, 1469–1508. [Google Scholar] [CrossRef]
- Munafò, F.; Donati, E.; Brindani, N.; Ottonello, G.; Armirotti, A.; De Vivo, M. Quercetin and luteolin are single-digit micromolar inhibitors of the SARS-CoV-2 RNA-dependent RNA polymerase. Sci. Rep. 2022, 12, 10571. [Google Scholar] [CrossRef]
- Sepay, N.; Mondal, R.; Al-Muhanna, M.K.; Saha, D. Identification of natural flavonoids as novel EGFR inhibitors using DFT, molecular docking, and molecular dynamics. New J. Chem. 2022, 46, 9735–9744. [Google Scholar] [CrossRef]
- Moulishankar, A.; Lakshmanan, K. Data on molecular docking of naturally occurring flavonoids with biologically important targets. Data Brief. 2020, 29, 105243. [Google Scholar] [CrossRef]
- Adeoye, A.O.; Olanlokun, J.O.; Tijani, H.; Lawal, S.O.; Babarinde, C.O.; Akinwole, M.T.; Bewaji, C.O. Molecular docking analysis of apigenin and quercetin from ethylacetate fraction of Adansonia digitata with malaria-associated calcium transport protein: An in silico approach. Heliyon 2019, 5, e02248. [Google Scholar] [CrossRef]
- Yang, J.-y.; Li, M.; Zhang, C.-l.; Liu, D. Pharmacological properties of baicalin on liver diseases: A narrative review. Pharmacol. Rep. 2021, 73, 1230–1239. [Google Scholar] [CrossRef]
- Invernizzi, P.; Carbone, M.; Jones, D.; Levy, C.; Little, N.; Wiesel, P.; Nevens, F.; the study investigators. Setanaxib, a first-in-class selective NADPH oxidase 1/4 inhibitor for primary biliary cholangitis: A randomized, placebo-controlled, phase 2 trial. Liver Int. 2023, 43, 1507–1522. [Google Scholar] [CrossRef]
- Love, B.L.; Barrons, R.; Veverka, A.; Snider, K.M. Urate-lowering therapy for gout: Focus on febuxostat. Pharmacotherapy 2010, 30, 594–608. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Collins, A. Polyphenols and DNA damage: A mixed blessing. Nutrients 2016, 8, 785. [Google Scholar] [CrossRef] [PubMed]
- Biechonski, S.; Gourevich, D.; Rall, M.; Aqaqe, N.; Yassin, M.; Zipin-Roitman, A.; Trakhtenbrot, L.; Olender, L.; Raz, Y.; Jaffa, A.J.; et al. Quercetin alters the DNA damage response in human hematopoietic stem and progenitor cells via TopoII- and PI3K-dependent mechanisms synergizing in leukemogenic rearrangements. Int. J. Cancer 2017, 140, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Elmegerhi, S.; Su, C.; Buglewicz, D.J.; Aizawa, Y.; Kato, T.A. Effect of hydroxyl group position in flavonoids on inducing single-stranded DNA damage mediated by cupric ions. Int. J. Mol. Med. 2018, 42, 658–664. [Google Scholar] [CrossRef]
- Das, A.; Majumder, D.; Saha, C. Correlation of binding efficacies of DNA to flavonoids and their induced cellular damage. J. Photochem. Photobiol. B Biol. 2017, 170, 256–262. [Google Scholar] [CrossRef]
- Jiang, M.; Zhu, M.; Wang, L.; Yu, S. Anti-tumor effects and associated molecular mechanisms of myricetin. Biomed. Pharmacother. 2019, 120, 109506. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Nile, S.H.; Keum, Y.S.; Nile, A.S.; Jalde, S.S.; Patel, R.V. Antioxidant, anti-inflammatory, and enzyme inhibitory activity of natural plant flavonoids and their synthesized derivatives. J. Biochem. Mol. Toxicol. 2018, 32, e22002. [Google Scholar] [CrossRef]
- Hinz, M.; Scheidereit, C. The IκB kinase complex in NF-κB regulation and beyond. EMBO Rep. 2014, 15, 46–61. [Google Scholar] [CrossRef]
- Saha, R.N.; Jana, M.; Pahan, K. MAPK p38 regulates transcriptional activity of NF-kappaB in primary human astrocytes via acetylation of p65. J. Immunol. 2007, 179, 7101–7109. [Google Scholar] [CrossRef]
- Huang, L.; Guo, Z.; Wang, F.; Fu, L. KRAS mutation: From undruggable to druggable in cancer. Signal Transduct. Target. Ther. 2021, 6, 386. [Google Scholar] [CrossRef] [PubMed]
- Bandaru, P.; Kondo, Y.; Kuriyan, J. The interdependent activation of Son-of-Sevenless and Ras. Cold Spring Harb. Perspect. Med. 2019, 9, a031534. [Google Scholar] [CrossRef] [PubMed]
- Kazi, A.; Ranjan, A.; Kumar, M.V.V.; Agianian, B.; Garcia Chavez, M.; Vudatha, V.; Wang, R.; Vangipurapu, R.; Chen, L.; Kennedy, P.; et al. Discovery of KRB-456, a KRAS G12D Switch-I/II Allosteric Pocket Binder That Inhibits the Growth of Pancreatic Cancer Patient-derived Tumors. Cancer Res. Commun. 2023, 3, 2623–2639. [Google Scholar] [CrossRef] [PubMed]
- Harwood, S.J.; Smith, C.R.; Lawson, J.D.; Ketcham, J.M. Selected Approaches to Disrupting Protein-Protein Interactions within the MAPK/RAS Pathway. Int. J. Mol. Sci. 2023, 24, 7373. [Google Scholar] [CrossRef]
- Mohanty, A.; Nam, A.; Srivastava, S.; Jones, J.; Lomenick, B.; Singhal, S.S.; Guo, L.; Cho, H.; Li, A.; Behal, A.; et al. Acquired resistance to KRAS G12C small-molecule inhibitors via genetic/nongenetic mechanisms in lung cancer. Sci. Adv. 2023, 9, eade3816. [Google Scholar] [CrossRef]
- Desnoyers, A.; Nadler, M.B.; Kumar, V.; Saleh, R.; Amir, E. Comparison of treatment-related adverse events of different Cyclin-dependent kinase 4/6 inhibitors in metastatic breast cancer: A network meta-analysis. Cancer Treat. Rev. 2020, 90, 102086. [Google Scholar] [CrossRef]
- Puri, P.; Wajih, Z.; Fadia, M.; Jiang, S. Letrozole-induced acute interstitial nephritis. J. Clin. and Med. Case Rep. 2020, 1, 2–3. [Google Scholar] [CrossRef]
- Almutlaq, R.N.; Newell-Fugate, A.E.; Evans, L.C.; Fatima, H.; Gohar, E.Y. Aromatase inhibition increases blood pressure and markers of renal injury in female rats. Am. J. Physiol. Renal Physiol. 2022, 323, F349–F360. [Google Scholar] [CrossRef]
- Durrant, J.D.; McCammon, J.A. Molecular dynamics simulations and drug discovery. BMC Biol. 2011, 9, 71. [Google Scholar] [CrossRef]
- Wang, X.; Cao, Y.; Chen, S.; Lin, J.; Bian, J.; Huang, D. Anti-inflammation activity of flavones and their structure-activity relationship. J. Agric. Food Chem. 2021, 69, 7285–7302. [Google Scholar] [CrossRef]
- Talib, W.H.; Abuawad, A.; Thiab, S.; Alshweiat, A.; Mahmod, A.I. Flavonoid-based nanomedicines to target tumor microenvironment. OpenNano 2022, 8, 100081. [Google Scholar] [CrossRef]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminformatics 2012, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub- Family | Flavonoids | NOX PDB: 2CDU | XO PDB: 1FIQ | XO PDB: 3ETR | iNOS PDB: 4NOS | ||||
|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
| Flavones | Apigenin | −6.83 | 9.90 | −7.74 | 2.12 | −7.43 | 3.56 | −7.05 | 6.85 |
| Luteolin | −6.21 | 27.88 | −7.54 | 2.99 | −6.92 | 8.50 | −5.87 | 49.73 | |
| Chrysin | −6.35 | 22.17 | −8.23 | 0.92 | −8.05 | 1.25 | −7.03 | 7.08 | |
| Baicalein | −6.07 | 35.56 | −8.48 | 0.61 | −8.25 | 0.90 | −6.98 | 7.62 | |
| Baicalin | −7.16 | 5.68 | −6.33 | 23.03 | −6.27 | 25.44 | −7.30 | 4.47 | |
| Vitexin | −6.23 | 26.95 | −6.96 | 7.95 | −6.12 | 32.86 | −6.64 | 13.63 | |
| Isovitexin | −6.16 | 30.52 | −7.15 | 5.75 | −7.29 | 4.55 | −7.16 | 5.66 | |
| Orientin | −6.01 | 39.20 | −6.53 | 16.31 | −6.09 | 34.17 | −6.00 | 39.70 | |
| Isoorientin | −5.55 | 85.34 | −6.92 | 8.50 | −7.15 | 5.70 | −6.57 | 15.30 | |
| Hispidulin | −6.02 | 38.48 | −7.63 | 2.53 | −7.19 | 5.34 | −5.99 | 40.61 | |
| Tricin | −5.68 | 69.19 | −7.82 | 1.84 | −7.41 | 3.67 | −5.94 | 44.46 | |
| Isoflavones | Genistein | −6.38 | 21.13 | −7.27 | 4.72 | −6.99 | 7.50 | −6.40 | 20.48 |
| Daidzein | −6.64 | 13.47 | −7.36 | 4.05 | −6.99 | 7.52 | −7.60 | 2.66 | |
| Flavonols | Quercetin | −5.51 | 91.56 | −7.53 | 3.01 | −7.08 | 6.41 | −5.55 | 85.60 |
| Kaempferol | −5.99 | 40.73 | −7.66 | 2.44 | −7.65 | 2.49 | −5.77 | 58.58 | |
| Fisetin | −6.45 | 18.82 | −8.77 | 0.37 | −8.70 | 0.42 | −5.85 | 51.23 | |
| Myricetin | −5.03 | 205.46 | −6.75 | 11.26 | −6.60 | 14.60 | −5.05 | 200.07 | |
| NOX inhibitor (setanaxib) | −8.35 | 0.76 | na | na | na | na | na | na | |
| XO inhibitor (febuxostat) | na | na | −6.94 | 8.18 | −5.89 | 48.45 | na | na | |
| iNOS inhibitor(iNOS inhibitor-10) | na | na | na | na | na | na | −11.04 | 0.008 | |
| Sub- Family | Flavonoids | IKK PDB: 4KIK | p38 MAPK PDB: 4DLI | NF-kB (p50–p65) PDB: 1VKX | NF-kB (p52–p65) PDB: 3DO7 | COX-2 PDB: 3NT1 | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
| Flavones | Apigenin | −7.45 | 3.47 | −8.21 | 0.96 | −5.54 | 87.04 | −6.16 | 30.54 | −6.68 | 12.72 |
| Luteolin | −6.75 | 11.19 | −8.21 | 0.95 | −5.61 | 77.81 | −6.34 | 22.43 | −6.79 | 10.48 | |
| Chrysin | −6.83 | 9.89 | −8.40 | 0.70 | −5.50 | 92.88 | −6.33 | 22.84 | −6.84 | 9.66 | |
| Baicalein | −6.96 | 7.92 | −8.31 | 0.80 | −5.47 | 98.11 | −6.15 | 31.29 | −6.69 | 12.57 | |
| Baicalin | −7.98 | 1.42 | −9.91 | 0.05 | −4.72 | 344.90 | −5.75 | 61.09 | −5.49 | 94.28 | |
| Vitexin | −7.63 | 2.55 | −9.29 | 0.15 | −6.15 | 31.19 | −5.72 | 63.95 | −7.00 | 7.40 | |
| Isovitexin | −7.85 | 1.77 | −7.41 | 3.71 | −5.14 | 172.13 | −5.64 | 72.90 | −6.34 | 22.52 | |
| Orientin | −7.51 | 3.13 | −9.25 | 0.17 | −5.61 | 77.81 | −5.83 | 53.14 | −6.73 | 11.65 | |
| Isoorientin | −7.70 | 2.27 | −6.85 | 9.45 | −6.10 | 33.82 | −5.20 | 158.18 | −5.40 | 109.50 | |
| Hispidulin | −7.26 | 4.74 | −7.99 | 1.38 | −5.40 | 109.57 | −6.21 | 27.93 | −6.04 | 37.66 | |
| Tricin | −6.84 | 9.65 | −8.44 | 0.65 | −5.24 | 143.80 | −5.41 | 109.01 | −6.70 | 12.27 | |
| Isoflavones | Genistein | −6.94 | 8.21 | −8.08 | 1.19 | −6.05 | 36.78 | −5.89 | 48.27 | −5.90 | 47.13 |
| Daidzein | −7.60 | 2.66 | −7.76 | 2.03 | −5.51 | 91.38 | −5.96 | 42.65 | −6.56 | 15.64 | |
| Flavonols | Quercetin | −6.74 | 11.43 | −8.43 | 0.67 | −5.04 | 203.17 | −5.63 | 74.70 | −6.09 | 34.53 |
| Kaempferol | −7.04 | 6.88 | −8.19 | 0.99 | −5.29 | 131.97 | −5.95 | 43.51 | −5.76 | 60.14 | |
| Fisetin | −6.71 | 12.10 | −8.14 | 1.08 | −5.24 | 143.20 | −5.94 | 44.59 | −6.72 | 11.84 | |
| Myricetin | −6.44 | 18.88 | −8.34 | 0.77 | −4.72 | 344.66 | −5.30 | 130.94 | −5.29 | 132.73 | |
| MAPK p38 inhibitor (adezmapimod) | na | na | −9.96 | 0.05 | na | na | na | na | na | na | |
| NF-kB inhibitor (sulfasalazine) | na | na | na | na | −6.99 | 7.49 | −7.42 | 3.62 | na | na | |
| COX-2 inhibitor (naproxen) | na | na | na | na | na | na | na | na | −7.61 | 2.62 | |
| Sub- Family | Flavonoids | EGFR (T790M/V948R) PDB: 6Z4B | KRAS G12C PDB: 4LV6 Switch I | KRAS G12C PDB: 4LV6 Switch II | KRAS G12D PDB: 6GJ8 Switch I | KRAS G12D PDB: 6GJ8 Switch II | KRAS G12V PDB:8AZZ Switch I | KRAS G12V PDB: 8AZZ Switch II | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
| Flavones | Apigenin | −7.41 | 3.67 | −7.68 | 2.35 | −6.47 | 17.97 | −7.86 | 1.72 | −5.68 | 69.12 | −7.22 | 5.14 | −6.57 | 15.24 |
| Luteolin | −7.64 | 2.50 | −7.69 | 2.31 | −6.37 | 21.47 | −7.47 | 3.37 | −5.25 | 142.33 | −6.81 | 10.18 | −6.65 | 13.36 | |
| Chrysin | −7.35 | 4.07 | −7.75 | 2.09 | −6.56 | 15.58 | −7.55 | 2.90 | −5.74 | 61.73 | −7.44 | 3.52 | −7.19 | 5.41 | |
| Baicalein | −6.76 | 11.08 | −7.07 | 6.52 | −6.67 | 12.81 | −7.94 | 1.50 | −5.27 | 136.05 | −7.34 | 4.15 | −7.37 | 3.98 | |
| Baicalin | −8.08 | 1.19 | −7.85 | 1.76 | −8.28 | 0.849 | −6.95 | 8.01 | −5.37 | 115.45 | −5.83 | 53.33 | −7.72 | 2.21 | |
| Vitexin | −8.04 | 1.27 | −7.78 | 1.97 | −6.76 | 11.08 | −6.27 | 25.42 | −5.72 | 64.64 | −5.83 | 53.71 | −6.65 | 13.29 | |
| Isovitexin | −7.79 | 1.94 | −7.14 | 5.85 | −7.77 | 2.01 | −7.89 | 1.64 | −5.62 | 76.58 | −6.21 | 28.03 | −7.49 | 3.22 | |
| Orientin | −7.88 | 1.68 | −7.87 | 1.70 | −7.18 | 5.42 | −6.75 | 11.29 | −4.66 | 381.43 | −5.79 | 56.79 | −6.21 | 28.00 | |
| Isoorientin | −7.68 | 2.33 | −6.83 | 9.89 | −6.85 | 9.48 | −6.80 | 10.44 | −5.08 | 188.42 | −5.93 | 45.13 | −7.19 | 5.37 | |
| Hispidulin | −7.03 | 7.08 | −6.25 | 26.34 | −6.03 | 38.27 | −7.76 | 2.06 | −5.00 | 215.17 | −6.31 | 23.52 | −6.63 | 13.87 | |
| Tricin | −6.32 | 23.29 | −6.95 | 7.99 | −6.73 | 11.70 | −6.89 | 8.85 | −4.56 | 453.29 | −5.93 | 45.15 | −5.90 | 47.32 | |
| Isoflavone | Genistein | −6.55 | 15.76 | −6.94 | 8.13 | −6.53 | 16.33 | −8.83 | 0.34 | −5.31 | 127.81 | −6.84 | 9.68 | −7.95 | 1.50 |
| Daidzein | −7.06 | 6.63 | −6.90 | 8.73 | −6.97 | 7.73 | −8.66 | 0.45 | −5.20 | 154.75 | −6.93 | 7.69 | −7.26 | 4.76 | |
| Flavonols | Quercetin | −7.03 | 6.99 | −7.09 | 6.39 | −6.36 | 21.93 | −7.62 | 2.59 | −5.01 | 213.68 | −6.17 | 29.81 | −6.77 | 10.92 |
| Kaempferol | −7.28 | 4.62 | −7.09 | 6.32 | −6.79 | 10.50 | −7.71 | 2.24 | −5.13 | 174.36 | −6.26 | 25.99 | −6.82 | 10.08 | |
| Fisetin | −6.45 | 18.68 | −6.72 | 11.86 | −6.48 | 17.91 | −7.85 | 1.76 | −5.40 | 110.47 | −6.79 | 10.62 | −7.36 | 4.02 | |
| Myricetin | −6.61 | 14.21 | −6.60 | 14.42 | −6.12 | 32.43 | −7.94 | 1.52 | −4.49 | 514.89 | −6.07 | 35.71 | −6.55 | 15.87 | |
| EGFR inhibitor (erlotinib) | −8.86 | 0.32 | na | na | na | na | na | na | na | na | na | na | na | na | |
| EGFR inhibitor (gefitinib) | −8.20 | 0.97 | na | na | na | na | na | na | na | na | na | na | na | na | |
| KRAS inhibitor (adagrasib) | na | na | −11.31 | 0.005 | −11.67 | 0.003 | na | na | na | na | na | na | na | na | |
| KRAS inhibitor (sotorasib) | na | na | −9.40 | 0.128 | −8.95 | 0.28 | na | na | na | na | na | na | na | na | |
| KRAS inhibitor (MRTX1133) | na | na | na | na | na | na | −7.67 | 2.39 | −7.49 | 3.22 | na | na | na | na | |
| KRAS inhibitor (BI-2865) | na | na | na | na | na | na | na | na | na | na | −6.97 | 7.74 | −11.12 | 0.007 | |
| Sub- Family | Flavonoids | BRAF V599E PDB: 1UWJ | BRAF V600E PDB: 3OG7 | MEK-1 C121S PDB: 7F2X | ERK-2 G169D PDB: 6D5Y | ||||
|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
| Flavones | Apigenin | −7.72 | 2.20 | −6.93 | 8.36 | −6.58 | 15.00 | −5.66 | 71.20 |
| Luteolin | −6.99 | 7.50 | −7.07 | 6.52 | −6.23 | 27.28 | −5.69 | 67.27 | |
| Chrysin | −7.76 | 2.05 | −6.89 | 8.93 | −6.46 | 18.34 | −5.31 | 129.07 | |
| Baicalein | −7.54 | 2.99 | −7.08 | 6.44 | −6.47 | 18.11 | −5.49 | 94.13 | |
| Baicalin | −9.23 | 0.17 | −7.72 | 2.21 | −6.97 | 7.81 | −6.41 | 20.08 | |
| Vitexin | −8.09 | 1.17 | −7.25 | 4.82 | −6.11 | 33.08 | −6.27 | 25.48 | |
| Isovitexin | −8.10 | 1.16 | −7.54 | 2.97 | −6.85 | 9.54 | −5.34 | 122.47 | |
| Orientin | −7.31 | 4.35 | −6.83 | 9.81 | −6.12 | 32.74 | −6.32 | 23.33 | |
| Isoorientin | −7.85 | 1.75 | −5.97 | 42.07 | −6.91 | 8.65 | −5.25 | 141.59 | |
| Hispidulin | −6.64 | 13.53 | −6.48 | 17.69 | −6.25 | 26.08 | −5.44 | 103.72 | |
| Tricin | −6.88 | 9.05 | −6.03 | 37.97 | −7.07 | 6.56 | −5.38 | 114.13 | |
| Isoflavone | Genistein | −6.96 | 7.93 | −6.51 | 16.97 | −6.78 | 10.73 | −5.24 | 144.57 |
| Daidzein | −6.90 | 8.70 | −6.93 | 8.34 | −6.86 | 9.32 | −5.43 | 105.32 | |
| Flavonols | Quercetin | −6.43 | 19.31 | −5.59 | 79.26 | −6.14 | 31.51 | −5.49 | 95.16 |
| Kaempferol | −6.79 | 10.48 | −5.99 | 40.34 | −6.12 | 32.40 | −5.33 | 123.20 | |
| Fisetin | −7.06 | 6.73 | −5.77 | 58.80 | −6.42 | 19.80 | −5.32 | 126.37 | |
| Myricetin | −6.82 | 10.00 | −5.05 | 197.92 | −5.74 | 61.89 | −5.14 | 171.93 | |
| RAF inhibitor (dabrafenib) | −13.27 | 1.87 × 10−4 | −11.76 | 0.002 | na | na | na | na | |
| RAF inhibitor (encorafenib) | −10.57 | 0.02 | −7.85 | 1.77 | na | na | na | na | |
| MEK inhibitor (trametinib) | na | na | na | na | −8.96 | 0.270 | na | na | |
| MEK inhibitor (selumetinib) | na | na | na | na | −6.81 | 10.23 | na | na | |
| ERK inhibitor (ulixertinib) | na | na | na | na | na | na | −7.85 | 1.75 | |
| Sub- Family | Flavonoids | PI3K PDB: 1E7U | AKT PDB: 3O96 | mTOR PDB: 4JT6 | |||
|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
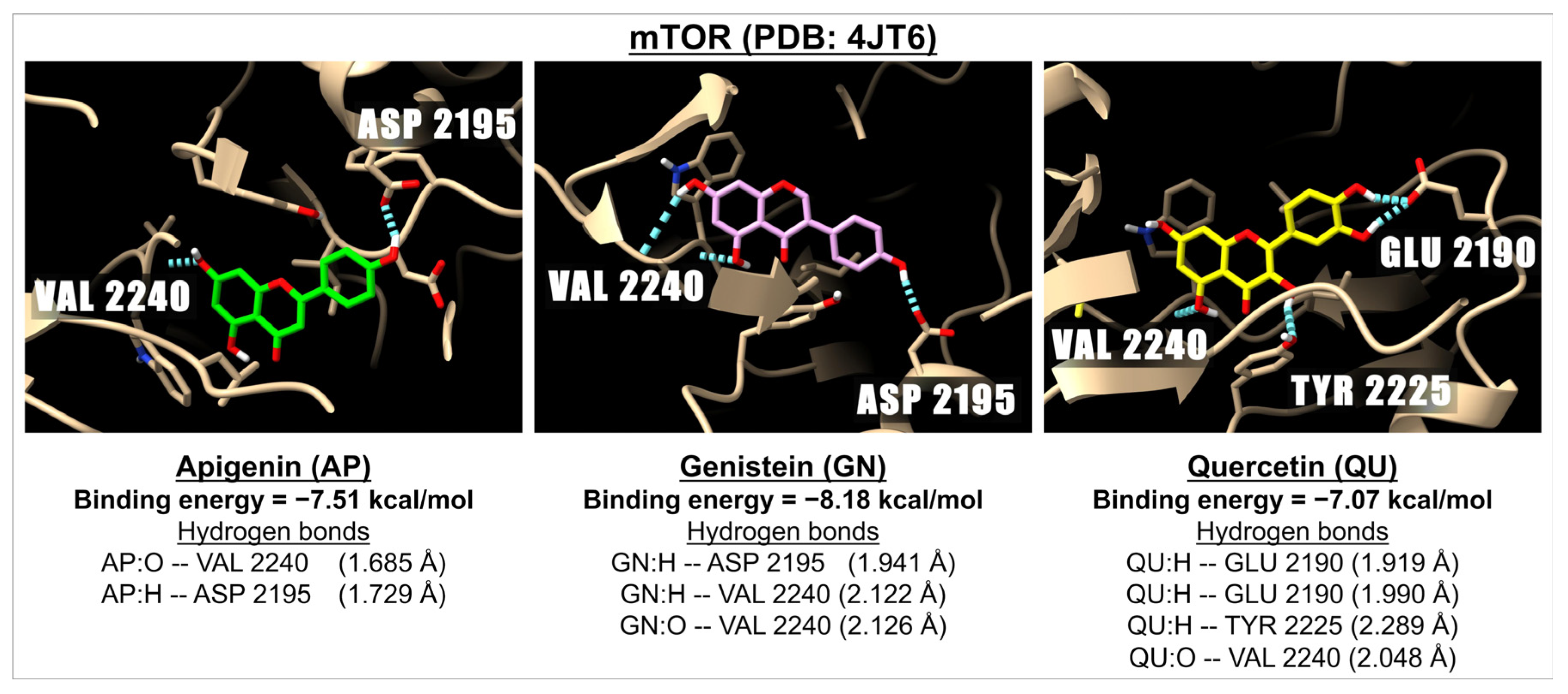
| Flavones | Apigenin | −7.05 | 6.81 | −6.57 | 15.26 | −7.51 | 3.12 |
| Luteolin | −7.27 | 4.72 | −6.71 | 12.12 | −7.80 | 1.93 | |
| Chrysin | −6.69 | 12.58 | −6.80 | 10.40 | −7.74 | 2.12 | |
| Baicalein | −7.20 | 5.25 | −6.42 | 19.64 | −7.87 | 1.69 | |
| Baicalin | −7.32 | 4.33 | −8.27 | 0.86 | −8.61 | 0.49 | |
| Vitexin | −6.70 | 12.17 | −8.64 | 0.46 | −7.28 | 4.63 | |
| Isovitexin | −5.40 | 110.84 | −8.17 | 1.03 | −7.07 | 6.53 | |
| Orientin | −7.11 | 6.13 | −8.19 | 0.99 | −7.26 | 4.79 | |
| Isoorientin | −5.39 | 112.16 | −8.02 | 1.31 | −7.37 | 3.97 | |
| Hispidulin | −7.14 | 5.84 | −6.49 | 17.57 | −7.91 | 1.58 | |
| Tricin | −7.37 | 3.96 | −6.32 | 23.26 | −7.40 | 3.79 | |
| Isoflavone | Genistein | −6.68 | 12.70 | −6.46 | 18.53 | −8.18 | 1.01 |
| Daidzein | −7.02 | 7.13 | −6.67 | 12.84 | −7.52 | 3.05 | |
| Flavonols | Quercetin | −6.93 | 8.31 | −6.12 | 32.39 | −7.07 | 6.62 |
| Kaempferol | −6.80 | 10.42 | −6.27 | 25.57 | −6.96 | 7.90 | |
| Fisetin | −6.83 | 9.88 | −6.73 | 11.58 | −7.86 | 1.73 | |
| Myricetin | −6.34 | 22.69 | −6.03 | 38.21 | −6.88 | 9.09 | |
| PI3K inhibitor (buparlisib) | −7.16 | 5.64 | na | na | na | na | |
| PI3K inhibitor (copanlisib) | −8.86 | 0.32 | na | na | na | na | |
| AKT inhibitor (GSK690693) | na | na | −9.73 | 0.07 | na | na | |
| mTOR inhibitor (PI-103) | na | na | na | na | −9.22 | 0.15 | |
| Sub- Family | Flavonoids | CDK2 PDB: 1GIH | CDK2 PDB: 4ERW | CDK2 PDB: 1AQ1 | CDK4 PDB: 3G33 | CDK4 PDB: 2W96 | CDK6 PDB:1XO2 | CDK6 PDB: 5L2T | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
| Flavones | Apigenin | −7.21 | 5.17 | −6.62 | 14.07 | −6.88 | 9.10 | −5.34 | 121.78 | −6.64 | 13.66 | −7.80 | 1.75 | −8.73 | 0.40 |
| Luteolin | −7.18 | 5.48 | −6.42 | 19.74 | −6.85 | 9.53 | −5.75 | 60.80 | −6.53 | 16.42 | −7.92 | 1.57 | −8.14 | 1.07 | |
| Chrysin | −7.23 | 5.02 | −6.91 | 8.60 | −7.16 | 5.69 | −5.21 | 151.49 | −6.67 | 12.89 | −7.75 | 2.10 | −8.37 | 0.73 | |
| Baicalein | −7.03 | 6.98 | −6.55 | 15.79 | −6.99 | 7.51 | −5.75 | 60.86 | −6.50 | 17.07 | −8.10 | 1.15 | −8.40 | 0.70 | |
| Baicalin | −7.33 | 4.21 | −8.69 | 0.43 | −8.32 | 0.79 | −6.41 | 20.17 | −6.74 | 11.49 | −8.46 | 0.63 | −8.60 | 0.50 | |
| Vitexin | −8.78 | 0.37 | −7.67 | 2.40 | −6.91 | 8.58 | −6.45 | 18.78 | −6.75 | 11.27 | −9.77 | 0.07 | −7.14 | 5.86 | |
| Isovitexin | −8.20 | 0.97 | −8.26 | 0.88 | −7.18 | 5.50 | −4.82 | 294.02 | −6.33 | 23.03 | −8.95 | 0.27 | −9.77 | 0.07 | |
| Orientin | −8.46 | 0.63 | −7.70 | 2.25 | −7.25 | 4.88 | −6.21 | 27.89 | −6.49 | 17.57 | −9.35 | 0.14 | −6.92 | 8.41 | |
| Isoorientin | −7.79 | 1.95 | −7.92 | 1.51 | −7.38 | 3.89 | −5.14 | 170.04 | −5.89 | 47.98 | −8.97 | 0.27 | −8.81 | 0.35 | |
| Hispidulin | −6.97 | 7.80 | −6.78 | 10.78 | −6.89 | 8.85 | −5.54 | 87.47 | −5.97 | 41.76 | −8.05 | 1.25 | −8.39 | 0.71 | |
| Tricin | −7.48 | 3.28 | −6.92 | 8.40 | −6.61 | 14.24 | −5.24 | 144.80 | −7.04 | 6.90 | −7.62 | 2.61 | −7.27 | 4.72 | |
| Isoflavones | Genistein | −6.70 | 12.28 | −6.75 | 11.20 | −6.24 | 26.83 | −5.50 | 92.47 | −6.45 | 18.72 | −7.67 | 2.40 | −8.21 | 0.96 |
| Daidzein | −7.38 | 3.88 | −6.54 | 16.00 | −6.91 | 8.61 | −5.65 | 68.52 | −6.61 | 14.21 | −7.64 | 2.53 | −8.40 | 0.70 | |
| Flavonols | Quercetin | −6.58 | 15.03 | −6.35 | 22.27 | −6.64 | 13.56 | −5.82 | 54.35 | −5.89 | 48.55 | −8.13 | 1.11 | −7.49 | 3.21 |
| Kaempferol | −7.00 | 7.40 | −6.79 | 10.52 | −6.40 | 20.41 | −5.38 | 113.87 | −6.41 | 20.11 | −7.92 | 1.57 | −8.03 | 1.29 | |
| Fisetin | −7.03 | 7.05 | −6.64 | 13.50 | −6.99 | 7.50 | −5.90 | 47.35 | −6.50 | 17.33 | −8.55 | 0.54 | −6.99 | 7.50 | |
| Myricetin | −6.20 | 28.44 | −6.14 | 31.34 | −6.29 | 24.69 | −5.54 | 86.32 | −5.94 | 44.44 | −8.58 | 0.51 | −6.94 | 8.21 | |
| CDK 2 inhibitor (1PU) | −9.18 | 0.19 | −8.68 | 0.44 | −8.47 | 0.62 | na | na | na | na | na | na | na | na | |
| CDK 4 inhibitor (palbociclib) | na | na | na | na | na | na | −8.44 | 0.65 | −9.51 | 0.11 | na | na | na | na | |
| CDK 6 inhibitor (ribociclib) | na | na | na | na | na | na | na | na | na | na | −10.32 | 0.03 | −10.29 | 0.03 | |
| Sub- Family | Flavonoids | Aromatase PDB: 3EQM | Aromatase PDB: 5JKV | DNMT1 PDB: 3AV6 | HDAC1 PDB: 4BKX | HDAC2 PDB: 4LXZ | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
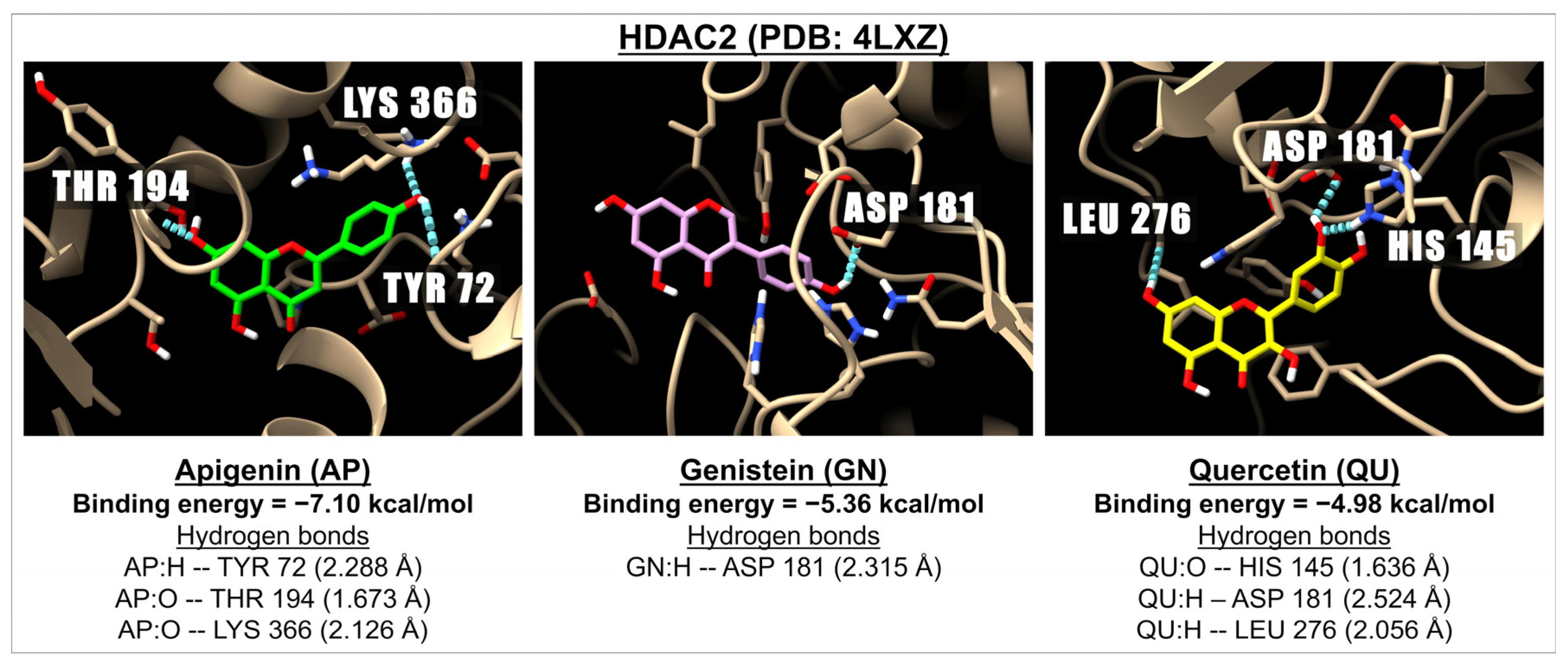
| Flavones | Apigenin | −6.51 | 16.90 | −6.50 | 17.33 | −7.80 | 1.92 | −6.82 | 10.08 | −7.10 | 6.25 |
| Luteolin | −6.37 | 21.47 | −6.19 | 28.89 | −7.66 | 2.41 | −6.85 | 9.50 | −6.42 | 19.63 | |
| Chrysin | −6.45 | 18.71 | −6.66 | 13.04 | −6.92 | 8.44 | −7.25 | 4.81 | −6.93 | 8.37 | |
| Baicalein | −6.83 | 9.84 | −6.36 | 21.96 | −7.01 | 7.27 | −6.68 | 12.61 | −5.95 | 43.19 | |
| Baicalin | −7.53 | 3.01 | −6.58 | 14.69 | −8.49 | 0.60 | −6.34 | 22.59 | −6.10 | 33.65 | |
| Vitexin | −7.15 | 5.77 | −6.21 | 28.19 | −8.03 | 1.30 | −5.91 | 46.78 | −5.88 | 48.72 | |
| Isovitexin | −6.33 | 23.05 | −6.85 | 9.60 | −6.97 | 7.82 | −4.36 | 634.05 | −3.93 | 1320 | |
| Orientin | −5.82 | 53.85 | −6.61 | 14.20 | −6.98 | 7.68 | −5.81 | 55.45 | −4.96 | 232.05 | |
| Isoorientin | −5.62 | 75.78 | −5.03 | 206.28 | −6.26 | 25.60 | −4.45 | 551.34 | −4.03 | 1120 | |
| Hispidulin | −5.94 | 44.44 | −5.68 | 68.38 | −7.77 | 2.02 | −6.19 | 29.14 | −5.74 | 61.90 | |
| Tricin | −5.82 | 54.07 | −5.82 | 53.91 | −7.58 | 2.77 | −6.48 | 17.65 | −5.89 | 48.24 | |
| Isoflavones | Genistein | −6.51 | 16.96 | −6.50 | 17.10 | −7.34 | 4.17 | −5.50 | 93.27 | −5.36 | 118.61 |
| Daidzein | −6.70 | 12.32 | −6.63 | 13.87 | −7.06 | 6.74 | −6.29 | 24.48 | −6.59 | 14.73 | |
| Flavonols | Quercetin | −5.79 | 57.03 | −5.62 | 75.60 | −7.54 | 2.99 | −4.73 | 338.65 | −4.98 | 222.77 |
| Kaempferol | −5.87 | 50.02 | −5.87 | 49.55 | −7.57 | 2.84 | −5.45 | 100.60 | −5.19 | 156.74 | |
| Fisetin | −6.29 | 24.47 | −6.29 | 24.56 | −8.12 | 1.12 | −5.49 | 94.40 | −5.13 | 174.33 | |
| Myricetin | −5.31 | 128.69 | −5.02 | 207.32 | −7.41 | 3.69 | −4.38 | 617.88 | −5.21 | 152.93 | |
| Aromatase inhibitor (letrozole) | −7.77 | 2.03 | −7.77 | 2.01 | na | na | na | na | na | na | |
| DNMT 1 inhibitor (azacytidine) | na | na | na | na | −7.41 | 3.73 | na | na | na | na | |
| HDAC 1&2 inhibitor (vorinostat) | na | na | na | na | na | na | −7.49 | 3.22 | −9.19 | 0.18 | |
| Sub- Family | Flavonoids | DNA gyrase B PDB: 6F86 | DNA gyrase B PDB: 4PRV | FABZ PDB: 3CF9 Site A | FABZ PDB: 3CF9 Site B | ||||
|---|---|---|---|---|---|---|---|---|---|
| Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | Binding Energy (kcal/mol) | Ki (µM) | ||
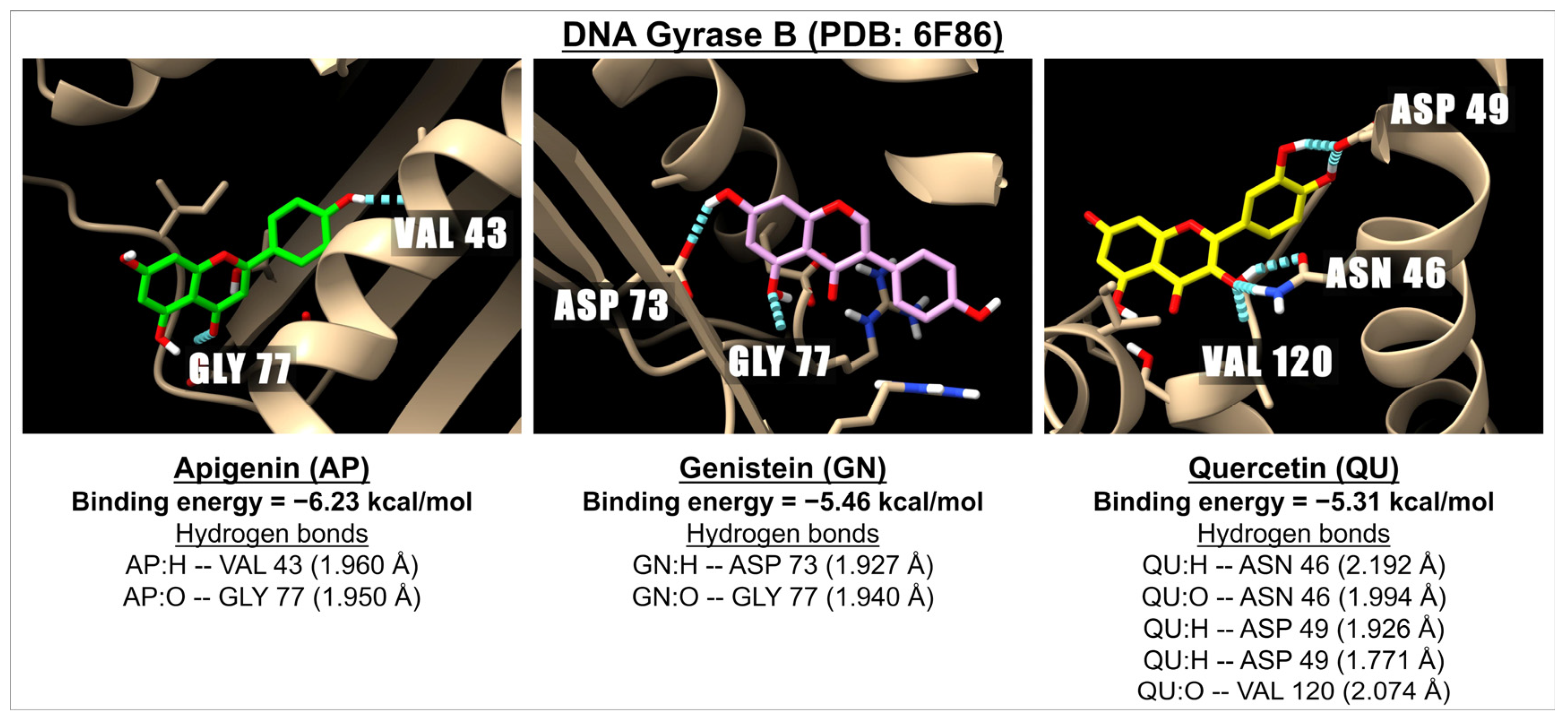
| Flavones | Apigenin | −6.23 | 27.21 | −6.51 | 16.97 | −7.70 | 2.27 | −7.98 | 1.42 |
| Luteolin | −5.74 | 61.92 | −6.39 | 20.80 | −8.19 | 0.98 | −7.74 | 2.11 | |
| Chrysin | −6.66 | 13.03 | −6.83 | 9.8 | −7.75 | 2.09 | −7.78 | 1.99 | |
| Baicalein | −6.24 | 26.84 | −6.43 | 19.45 | −7.85 | 1.77 | −7.45 | 3.45 | |
| Baicalin | −6.24 | 26.48 | −8.61 | 0.49 | −6.90 | 8.79 | −9.81 | 0.06 | |
| Vitexin | −5.37 | 115.48 | −7.34 | 4.18 | −7.52 | 3.07 | −9.44 | 0.12 | |
| Isovitexin | −6.00 | 39.75 | −7.29 | 4.51 | −7.32 | 4.30 | −8.36 | 0.74 | |
| Orientin | −5.13 | 173.59 | −7.13 | 5.94 | −6.99 | 7.50 | −8.96 | 0.27 | |
| Isoorientin | −5.25 | 142.45 | −7.75 | 2.07 | −7.05 | 6.84 | −8.89 | 0.30 | |
| Hispidulin | −6.10 | 33.55 | −6.52 | 16.65 | −7.72 | 2.20 | −7.48 | 3.27 | |
| Tricin | −6.81 | 10.14 | −6.22 | 27.59 | −7.13 | 5.96 | −8.06 | 1.23 | |
| Isoflavones | Genistein | −5.46 | 99.68 | −6.75 | 11.33 | −8.32 | 0.80 | −8.74 | 0.39 |
| Daidzein | −5.72 | 64.48 | −7.31 | 4.40 | −7.75 | 2.10 | −8.21 | 0.96 | |
| Flavonols | Quercetin | −5.31 | 128.9 | −6.64 | 13.69 | −7.36 | 4.01 | −7.29 | 4.57 |
| Kaempferol | −5.29 | 131.63 | −6.26 | 25.60 | −7.52 | 3.07 | −7.03 | 6.98 | |
| Fisetin | −5.83 | 53.49 | −7.26 | 4.77 | −7.53 | 3.05 | −7.55 | 2.90 | |
| Myricetin | −5.66 | 70.99 | −6.07 | 35.77 | −7.08 | 6.51 | −6.98 | 7.65 | |
| Antibiotic (ciprofloxacin) | −7.88 | 1.66 | −10.45 | 0.02 | −9.97 | 0.05 | −11.68 | 0.002 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peanlikhit, T.; Aryal, U.; Welsh, J.S.; Shroyer, K.R.; Rithidech, K.N. Evaluation of the Inhibitory Potential of Apigenin and Related Flavonoids on Various Proteins Associated with Human Diseases Using AutoDock. Int. J. Mol. Sci. 2025, 26, 2548. https://doi.org/10.3390/ijms26062548
Peanlikhit T, Aryal U, Welsh JS, Shroyer KR, Rithidech KN. Evaluation of the Inhibitory Potential of Apigenin and Related Flavonoids on Various Proteins Associated with Human Diseases Using AutoDock. International Journal of Molecular Sciences. 2025; 26(6):2548. https://doi.org/10.3390/ijms26062548
Chicago/Turabian StylePeanlikhit, Tanat, Uma Aryal, James S. Welsh, Kenneth R. Shroyer, and Kanokporn Noy Rithidech. 2025. "Evaluation of the Inhibitory Potential of Apigenin and Related Flavonoids on Various Proteins Associated with Human Diseases Using AutoDock" International Journal of Molecular Sciences 26, no. 6: 2548. https://doi.org/10.3390/ijms26062548
APA StylePeanlikhit, T., Aryal, U., Welsh, J. S., Shroyer, K. R., & Rithidech, K. N. (2025). Evaluation of the Inhibitory Potential of Apigenin and Related Flavonoids on Various Proteins Associated with Human Diseases Using AutoDock. International Journal of Molecular Sciences, 26(6), 2548. https://doi.org/10.3390/ijms26062548