
Unveiling a Novel Zearalenone Biodegradation Pathway in Metarhizium anisopliae and Elucidating the Role of Cytochrome P450

 ,
,  , and
, and
Abstract
1. Introduction
2. Results
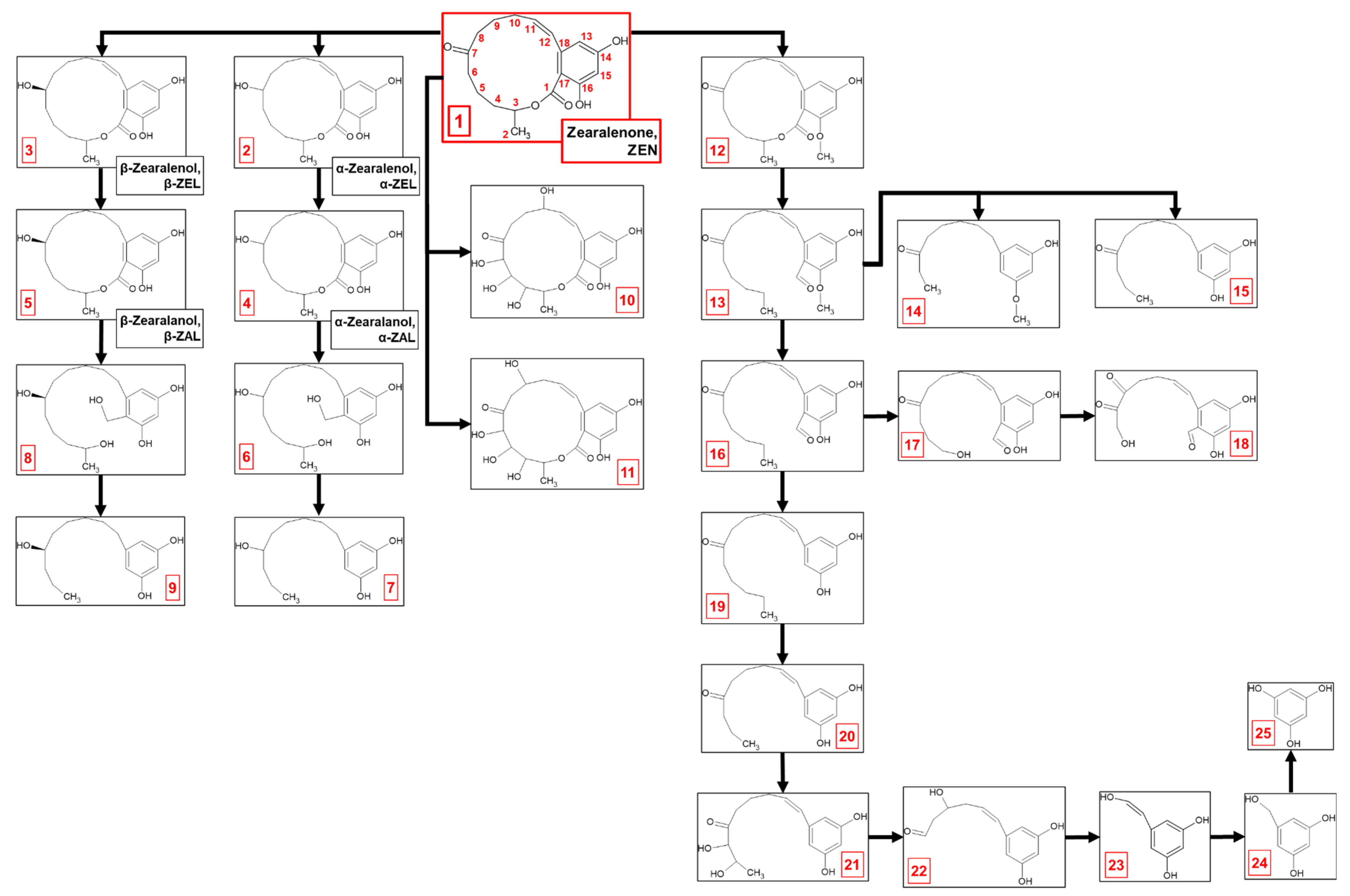
2.1. Zearalenone Biodegradation Pathway
2.2. Cytochrome P450 Involvement in ZEN Biodegradation
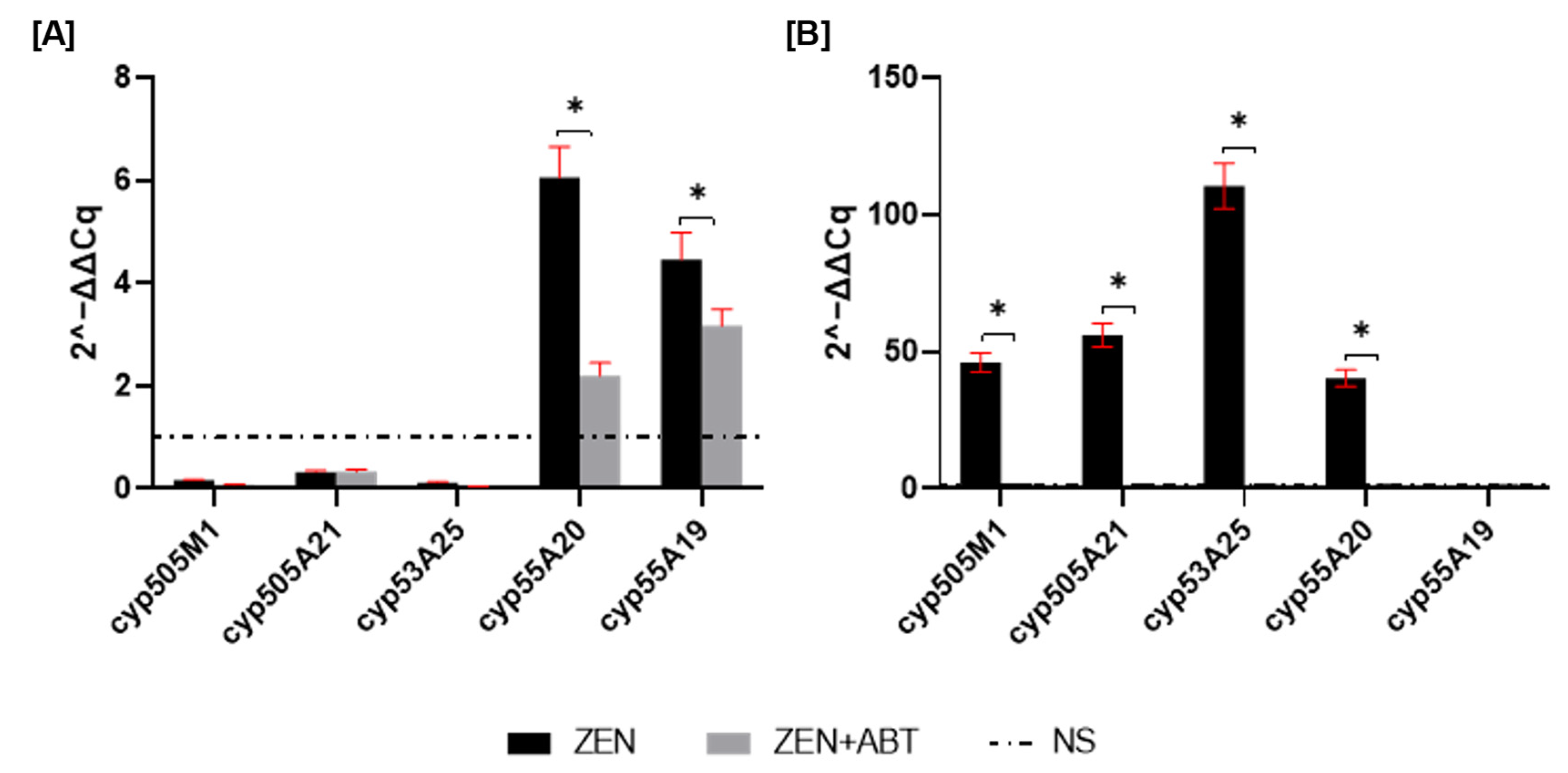
2.2.1. Cytochrome P450 Gene Expression Profiles in M. anisopliae During Zearalenone Degradation
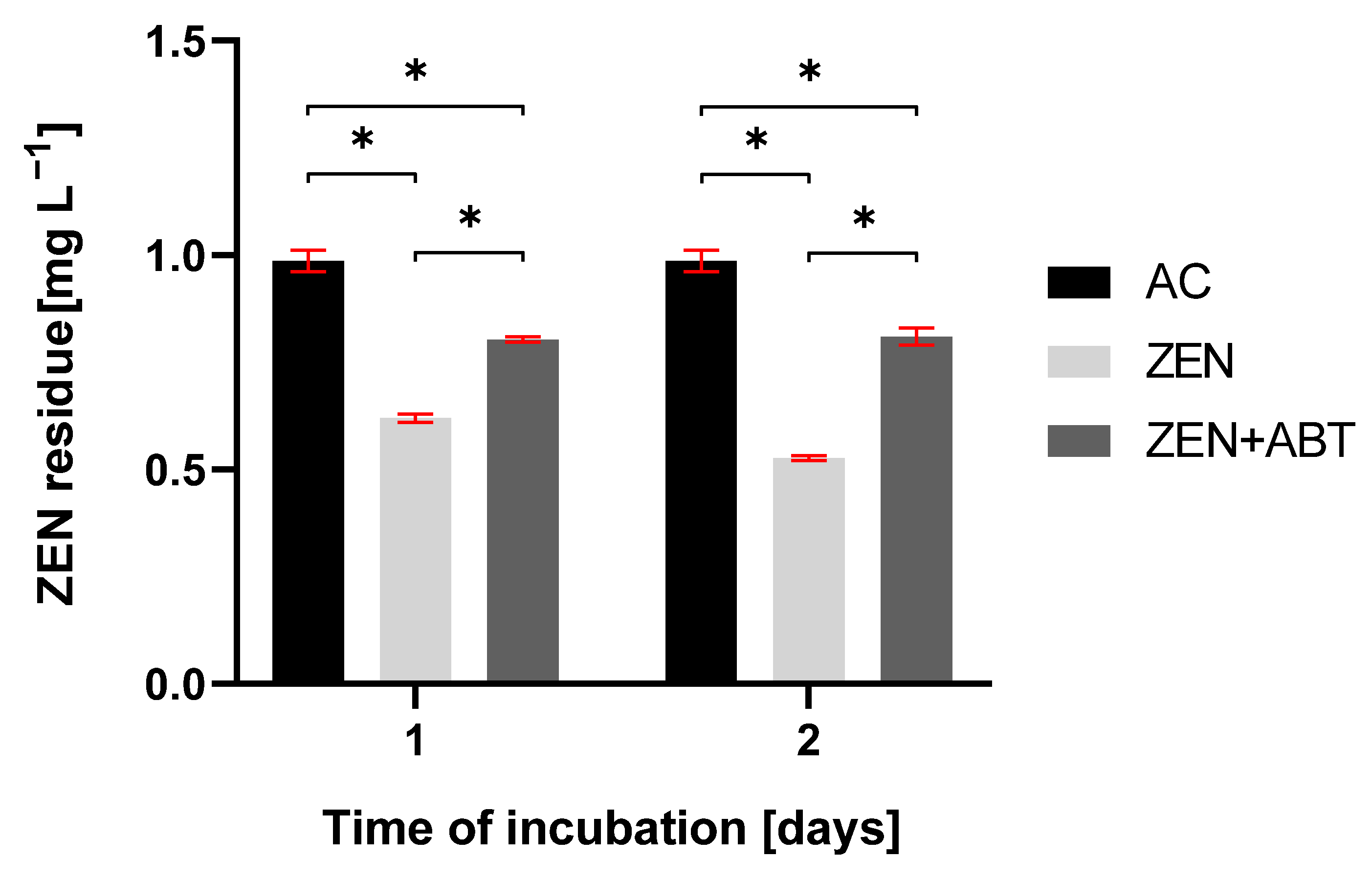
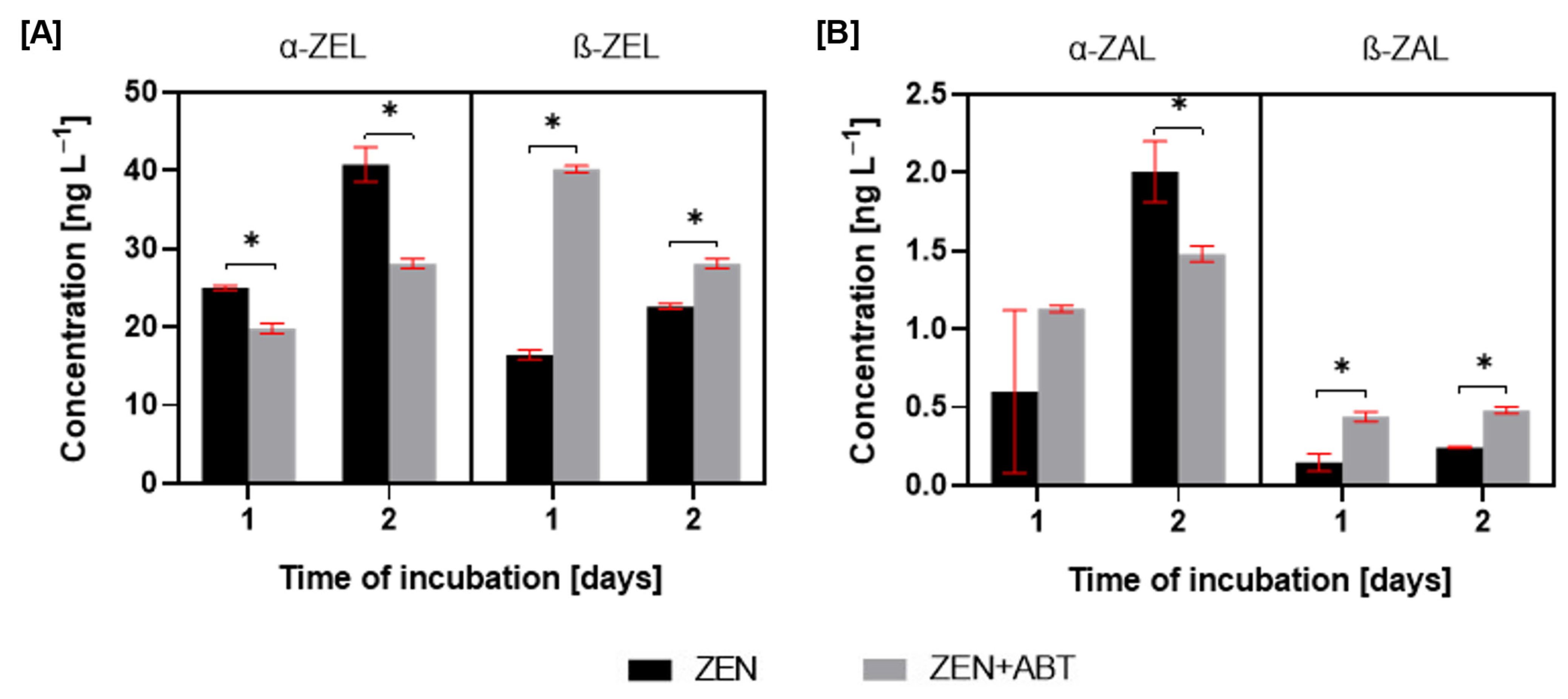
2.2.2. Changes in the Elimination Rate of Zearalenone and the Formation of Hydroxylated Derivatives After Inhibitory Assay with 1-Aminobenzotriazole
3. Discussion
4. Materials and Methods
4.1. Fungal Strain and Chemicals
4.2. Biodegradation Experiment
4.3. Sample Extraction and Preparation
4.4. Cytochrome P450 Activity
4.4.1. Cytochrome P450 Inhibitor Assay
4.4.2. RNA Extraction, cDNA Synthesis and Quantitative RT-PCR (RT-qPCR)
4.5. Analytical Methods
4.5.1. Quantitative Analysis of Zearalenone
4.5.2. Quantitative Analysis of α/β-Zearalenol and α/β-Zearalanol
4.5.3. Qualitative Analysis of Zearalenone Biodegradation and Biotransformation Derivatives
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brancini, G.T.P.; Hallsworth, J.E.; Corrochano, L.M.; Braga, G.Ú.L. Photobiology of the keystone genus Metarhizium. J. Photochem. Photobiol. B Biol. 2022, 226, 112374. [Google Scholar] [CrossRef] [PubMed]
- Leger, R.J.S. The evolution of complex Metarhizium-insect-plant interactions. Fungal Biol. 2024, 128, 2513–2528. [Google Scholar] [CrossRef]
- Nowak, M.; Soboń, A.; Litwin, A.; Różalska, S. 4-n-nonylphenol degradation by the genus Metarhizium with cytochrome P450 involvement. Chemosphere 2019, 220, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, R.; Kuśmierska, A.; Bernat, P. Ametryn removal by Metarhizium brunneum: Biodegradation pathway proposal and metabolic background revealed. Chemosphere 2018, 190, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, R.; Różalska, S.; Mironenka, J.; Bernat, P. Atrazine biodegradation by mycoinsecticide Metarhizium robertsii: Insights into its amino acids and lipids profile. J. Environ. Manag. 2020, 262, 110304. [Google Scholar] [CrossRef]
- Wu, C.; Tang, D.; Dai, J.; Tang, X.; Bao, Y.; Ning, J.; Zhen, Q.; Song, H.; Leger, R.J.S.; Fang, W. Bioremediation of mercury-polluted soil and water by the plant symbiotic fungus Metarhizium robertsii. Proc. Natl. Acad. Sci. USA 2022, 119, e2214513119. [Google Scholar] [CrossRef]
- Stolarek, P.; Różalska, S.; Bernat, P. Lipidomic adaptations of the Metarhizium robertsii strain in response to the presence of butyltin compounds. Biochim. Biophys. Acta—Biomembr. 2019, 1861, 316–326. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef]
- Han, X.; Huangfu, B.; Xu, T.; Xu, W.; Asakiya, C.; He, X.; Huang, K. Research Progress of Safety of Zearalenone: A Review. Toxins 2022, 14, 386. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, H.; Sun, F.; De Ruyck, K.; Zhang, J.; Jin, Y.; Li, Y.; Wang, Z.; Zhang, S.; De Saeger, S.; et al. Metabolic Profile of Zearalenone in Liver Microsomes from Different Species and Its in Vivo Metabolism in Rats and Chickens Using Ultra High-Pressure Liquid Chromatography-Quadrupole/Time-of-Flight Mass Spectrometry. J. Agric. Food Chem. 2017, 65, 11292–11303. [Google Scholar] [CrossRef]
- Malekinejad, H.; Colenbrander, B.; Fink-Gremmels, J. Hydroxysteroid dehydrogenases in bovine and porcine granulosa cells convert zearalenone into its hydroxylated metabolites α-zearalenol and β-zearalenol. Vet. Res. Commun. 2006, 30, 445–453. [Google Scholar] [CrossRef]
- Bravin, F.; Duca, R.C.; Balaguer, P.; Delaforge, M. In Vitro Cytochrome P450 Formation of a Mono-Hydroxylated Metabolite of Zearalenone Exhibiting Estrogenic Activities: Possible Occurrence of This Metabolite In Vivo. Int. J. Mol. Sci. 2009, 10, 1824–1837. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Barregard, L.; Bignami, M.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; et al. Appropriateness to set a group health-based guidance value for zearalenone and its modified forms. EFSA J. 2016, 14, e04425. [Google Scholar] [CrossRef]
- Ji, J.; Yu, J.; Ye, Y.; Sheng, L.; Fang, J.; Yang, Y.; Sun, X. Biodegradation methods and product analysis of zearalenone and its future development trend: A review. Food Control 2023, 145, 109469. [Google Scholar] [CrossRef]
- Brodehl, A.; Möller, A.; Kunte, H.J.; Koch, M.; Maul, R. Biotransformation of the mycotoxin zearalenone by fungi of the genera Rhizopus and Aspergillus. FEMS Microbiol. Lett. 2014, 359, 124–130. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Kakeya, H.; Takahashi-Ando, N.; Kimura, M.; Onose, R.; Yamaguchi, I.; Osada, H. Biotransformation of the Mycotoxin, Zearalenone, to a Non-estrogenic Compound by a Fungal Strain of Clonostachys sp. Biosci. Biotechnol. Biochem. 2002, 66, 2723–2726. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.; Soboń, A.; Bernat, P.; Różalska, S. Entomopathogenic fungi of the genus Cordyceps biotransform zearalenone—Metabolomic and proteomic backgrounds. Int. Biodeterior. Biodegrad. 2023, 179, 105572. [Google Scholar] [CrossRef]
- Nowak, M.; Bernat, P.; Różalska, S. Mutual interaction of the entomopathogenic and endophytic fungus Metarhizium anisopliae with zearalenone as a native component of crude Fusarium extract. Sci. Rep. 2024, 14, 22493. [Google Scholar] [CrossRef]
- Van Herwaarden, A.E.; Wagenaar, E.; Van Der Kruijssen, C.M.M.; Van Waterschoot, R.A.B.; Smit, J.W.; Song, J.Y.; Van Der Valk, M.A.; Van Tellingen, O.; Van Der Hoorn, J.W.A.; Rosing, H.; et al. Knockout of cytochrome P450 3A yields new mouse models for understanding xenobiotic metabolism. J. Clin. Investig. 2007, 117, 3583–3592. [Google Scholar] [CrossRef]
- Lin, L.; Fang, W.; Liao, X.; Wang, F.; Wei, D.; Leger, R.J.S. The MrCYP52 Cytochrome P450 Monoxygenase Gene of Metarhizium robertsii Is Important for Utilizing Insect Epicuticular Hydrocarbons. PLoS ONE 2011, 6, e28984. [Google Scholar] [CrossRef]
- Mahato, D.K.; Devi, S.; Pandhi, S.; Sharma, B.; Maurya, K.K.; Mishra, S.; Dhawan, K.; Selvakumar, R.; Kamle, M.; Mishra, A.K.; et al. Occurrence, Impact on Agriculture, Human Health, and Management Strategies of Zearalenone in Food and Feed: A Review. Toxins 2021, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Zhang, J.; Chen, Y.; Zhu, J. Zearalenone and Its Masked Forms in Cereals and Cereal-Derived Products: A Review of the Characteristics, Incidence, and Fate in Food Processing. J. Fungi 2022, 8, 976. [Google Scholar] [CrossRef]
- Metzler, M.; Pfeiffer, E.; Hildebrand, A.A. Zearalenone and its metabolites as endocrine disrupting chemicals. World Mycotoxin J. 2010, 3, 385–401. [Google Scholar] [CrossRef]
- Litwin, A.; Nowak, M.; Różalska, S. Entomopathogenic fungi: Unconventional applications. Rev. Environ. Sci. Bio/Technol. 2020, 19, 23–42. [Google Scholar] [CrossRef]
- Falkauskas, R.; Bakutis, B.; Jovaišienė, J.; Vaičiulienė, G.; Gerulis, G.; Kerzienė, S.; Jacevičienė, I.; Jacevičius, E.; Baliukonienė, V. Zearalenone and Its Metabolites in Blood Serum, Urine, and Milk of Dairy Cows. Animals 2022, 12, 1651. [Google Scholar] [CrossRef]
- Keller, L.; Abrunhosa, L.; Keller, K.; Rosa, C.A.; Cavaglieri, L.; Venâncio, A. Zearalenone and Its Derivatives α-Zearalenol and β-Zearalenol Decontamination by Saccharomyces cerevisiae Strains Isolated from Bovine Forage. Toxins 2015, 7, 3297. [Google Scholar] [CrossRef]
- Borzekowski, A.; Drewitz, T.; Keller, J.; Pfeifer, D.; Kunte, H.J.; Koch, M.; Rohn, S.; Maul, R. Biosynthesis and Characterization of Zearalenone-14-Sulfate, Zearalenone-14-Glucoside and Zearalenone-16-Glucoside Using Common Fungal Strains. Toxins 2018, 10, 104. [Google Scholar] [CrossRef]
- Kim, S.H.; Vujanovic, V. Biodegradation and biodetoxification of Fusarium mycotoxins by Sphaerodes mycoparasitica. AMB Express 2017, 7, 145. [Google Scholar] [CrossRef]
- Pfeiffer, E.; Hildebrand, A.; Damm, G.; Rapp, A.; Cramer, B.; Humpf, H.U.; Metzler, M. Aromatic hydroxylation is a major metabolic pathway of the mycotoxin zearalenone in vitro. Mol. Nutr. Food Res. 2009, 53, 1123–1133. [Google Scholar] [CrossRef]
- Fang, W.; Bidochka, M.J. Expression of genes involved in germination, conidiogenesis and pathogenesis in Metarhizium anisopliae using quantitative real-time RT-PCR. Mycol. Res. 2006, 110, 1165–1171. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubation Time [Days] | ZEN Residue [mg L−1] | |
|---|---|---|
| AC | ZEN | |
| 1 | 0.99 ± 0.03 | 0.63 ± 0.01 |
| 2 | 0.99 ± 0.01 | 0.53 ± 0.00 |
| 3 | 0.99 ± 0.02 | 0.47 ± 0.00 |
| 4 | 0.99 ± 0.02 | 0.37 ± 0.00 |
| 5 | 0.99 ± 0.02 | 0.21 ± 0.01 |
| 6 | 0.99 ± 0.01 | 0.18 ± 0.00 |
| 7 | 0.98 ± 0.02 | 0.11 ± 0.00 |
| 10 | 0.98 ± 0.03 | 0.11 ± 0.00 |
| 14 | 0.99 ± 0.02 | 0.09 ± 0.01 |
| ID | Chemical Formula | RT [min] | Neutral Mass | Theoretical m/z | Measured m/z | ppm | Transformation | Fragments, CID (Maximum 6 Most Intensive) |
|---|---|---|---|---|---|---|---|---|
| 1 | C18H22O5 | 12.35 | 318.146 | 317.1394 | 317.1387 | −2.4 | parent, zearalenone | 317.1 (100), 273.1 (20), 187.0 (20), 175.0 (53), 160.0 (26), 131.1 (63) |
| 2 | C18H24O5 | 11.71 | 320.1618 | 319.1551 | 319.1545 | −1.9 | + (H2), α-zearalenol | 319.2 (100), 275.2 (24), 188.0 (9), 174.0 (14), 160.0 (20), 130.0 (18) |
| 3 | C18H24O5 | 10.99 | 320.1608 | 319.1551 | 319.1549 | −0.6 | + (H2), β-zearalenol | 319.2 (100), 301.1 (7), 275.2 (21), 188.0 (7), 174.0 (11), 160.0 (17) |
| 4 | C18H26O5 | 11.56 | 322.177 | 321.1707 | 321.1697 | −3.3 | + (H4), α-zearalanol | 321.2 (100), 303.2 (19), 277.2 (46), 205.1 (6), 191.1 (6), 161.1 (5) |
| 5 | C18H26O5 | 10.78 | 322.1769 | 321.1707 | 321.1696 | −3.6 | + (H4), β-zearalenol | 321.2 (100), 303.2 (19), 293.2 (3), 277.2 (46), 205.1 (6), 161.1 (5) |
| 6 | C18H30O5 | 13.14 | 326.2078 | 325.202 | 325.2006 | −4.6 | + (H8) | 181.1 (54), 165.1 (15), 143.1 (100), 138.1 (24), 121.1 (6), 97.1 (12) |
| 7 | C15H24O3 | 13.08 | 252.1717 | 251.1653 | 251.1644 | −3.3 | + (H2) − (C3O2) | 251.2 (54), 207.1 (54), 205.1 (26), 193.1 (67), 179.1 (100), 122.0 (54) |
| 8 | C18H30O5 | 9.66 | 326.2082 | 325.202 | 325.2009 | −3.5 | + (H8) | 181.1 (34), 165.1 (25), 143.1 (100), 138.1 (34), 121.1 (5), 97.1 (10) |
| 9 | C15H24O3 | 11.73 | 252.1714 | 251.1653 | 251.1641 | −4.7 | + (H2) − (C3O2) | 233.2 (36), 207.1 (73), 179.1 (100), 122.0 (90), 109.0 (82), 73.0 (46) |
| 10 | C18H22O9 | 5.96 | 382.1252 | 381.1191 | 381.1179 | −3.2 | + (O4) | 177.1 (100), 167.0 (51), 159.0 (90), 153.1 (75), 147.0 (72), 123.0 (48) |
| 11 | C18H22O9 | 5.66 | 382.1253 | 381.1191 | 381.118 | −2.8 | + (O4) | 177.1 (100), 159.0 (55), 153.1 (64), 147.0 (38), 123.0 (29), 99.0 (30) |
| 12 | C19H24O5 | 10.39 | 332.1614 | 331.1551 | 331.1541 | −2.9 | + (C1H2) | 331.2 (72), 207.1 (100), 165.1 (65), 151.0 (36), 139.0 (19), 115.0 (26) |
| 13 | C19H26O4 | 11.97 | 318.1819 | 317.1758 | 317.1747 | −3.7 | + (C1H4) − (O1) | 317.2 (76), 191.1 (100), 177.1 (27), 166.1 (28), 162.0 (16), 112.0 (14) |
| 14 | C15H22O3 | 13.13 | 250.1557 | 249.1496 | 249.1485 | −4.6 | – (C3O2) | 249.1 (64), 205.2 (34), 203.1 (100), 191.1 (13), 177.1 (14) |
| 15 | C15H22O3 | 9.57 | 250.1563 | 249.1496 | 249.149 | −2.4 | − (C3O2) | 213.1 (10), 141.1 (100), 139.1 (49), 109.1 (24), 59.0 (24), 57.0 (18) |
| 16 | C18H24O4 | 13.75 | 304.1663 | 303.1602 | 303.159 | −3.9 | +(H2) − (O1) | 189.1 (6), 165.1 (12), 161.1 (23), 83.0 (6), 81.0 (100), 57.0 (7) |
| 17 | C16H20O5 | 10.23 | 292.1301 | 291.1238 | 291.1228 | −3.5 | − (C2H2) | 163.1 (20), 125.0 (100), 85.0 (29), 65.0 (44) |
| 18 | C14H14O6 | 7.21 | 278.0788 | 277.0718 | 277.0715 | −0.9 | − (C4H8) + (O1) | 175.0 (37), 149.1 (87), 133.0 (41), 121.1 (45), 109.0 (100), 107.1 (77) |
| 19 | C17H24O3 | 11.71 | 276.1725 | 275.1653 | 275.1652 | −0.3 | + (H2) − (C1O2) | 275.2 (80), 187.1 (100), 109.0 (78), 59.0 (98) |
| 20 | C15H20O3 | 11.87 | 248.1404 | 247.134 | 247.1331 | −3.3 | – (C3H2O3) | 203.1 (100), 133.1 (93), 123.1 (64) |
| 21 | C15H20O5 | 7.36 | 280.1299 | 279.1238 | 279.1226 | −4.1 | – (C3H2) | 190.1 (34), 163.1 (100), 107.0 (52), 79.1 (37) |
| 22 | C12H14O4 | 8.33 | 222.089 | 221.0819 | 221.0817 | −1.1 | – (C6H8O1) | 179.1 (8), 135.0 (23), 107.1 (100), 105.1 (10), 91.1 (88), 65.0 (8) |
| 23 | C8H8O3 | 9.29 | 152.047 | 151.0401 | 151.0397 | −2.3 | – (C10H14O2) | 107.1 (100), 106.0 (19), 92.0 (17) |
| 24 | C7H8O3 | 6.87 | 140.0468 | 139.0401 | 139.0396 | −3.6 | – (C11H14O2) | 95.0 (100), 69.0 (34) |
| 25 | C6H6O3 | 5.37 | 126.0317 | 125.0244 | 125.0245 | 0.3 | – (C12H16O2) | 83.0 (21), 81.0 (100), 65.0 (7), 63.0 (8) |
| Gene Name | Primer Sequence (5′-3′) |
|---|---|
| cyp53A25 | Forward: ACCATGCATCACTCCAAGGA Reverse: TCTCCATCTCTGCCACGTTT |
| cyp55A20 | Forward: CTCCAAGGCCGATGTTGTTC Reverse: CCCCGATTTCAATGTCCGTC |
| cyp55A19 | Forward: CAAGTTGCCACCCGATCAAA Reverse: AAGCAGGAGGAAGGCAATCT |
| cyp505A21 | Forward: TGTTGACAGCACGCTAGAGA Reverse: TGGCCTCTTCGACATCTTGT |
| cyp505M1 | Forward: TACCATCTCGTCCTCTTCGC Reverse: ACCGTGTCGAATTCCTCACT |
| ubi | Forward: AAGGGCGACATCAAGAAGGA Reverse: CGTTGACACCCATGACGTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowak, M.; Kozłowska, E.; Agier, J.; Góralczyk-Bińkowska, A.; Różalska, S. Unveiling a Novel Zearalenone Biodegradation Pathway in Metarhizium anisopliae and Elucidating the Role of Cytochrome P450. Int. J. Mol. Sci. 2025, 26, 2547. https://doi.org/10.3390/ijms26062547
Nowak M, Kozłowska E, Agier J, Góralczyk-Bińkowska A, Różalska S. Unveiling a Novel Zearalenone Biodegradation Pathway in Metarhizium anisopliae and Elucidating the Role of Cytochrome P450. International Journal of Molecular Sciences. 2025; 26(6):2547. https://doi.org/10.3390/ijms26062547
Chicago/Turabian StyleNowak, Monika, Elżbieta Kozłowska, Justyna Agier, Aleksandra Góralczyk-Bińkowska, and Sylwia Różalska. 2025. "Unveiling a Novel Zearalenone Biodegradation Pathway in Metarhizium anisopliae and Elucidating the Role of Cytochrome P450" International Journal of Molecular Sciences 26, no. 6: 2547. https://doi.org/10.3390/ijms26062547
APA StyleNowak, M., Kozłowska, E., Agier, J., Góralczyk-Bińkowska, A., & Różalska, S. (2025). Unveiling a Novel Zearalenone Biodegradation Pathway in Metarhizium anisopliae and Elucidating the Role of Cytochrome P450. International Journal of Molecular Sciences, 26(6), 2547. https://doi.org/10.3390/ijms26062547