Abstract
The phosphorylation state of 20 types of intracellular proteins in the presence of the protein phosphatase 1 (PP1)- and PP2A-specific Ser/Thr phosphatase inhibitor calyculin A or the Tyr phosphatase inhibitor pervanadate was visualized by Phos-tag SDS-PAGE followed by immunoblotting. All blots showed a Phos-tag pattern indicating increased phosphorylation in the presence of one or both phosphatase inhibitors. The increase in phosphorylation stoichiometry per protein tends to be greater for Ser/Thr phosphatase inhibition than for Tyr phosphatase inhibition. This is consistent with the fact that the number of Ser/Thr kinase genes in the human genome is greater than that of Tyr kinases and with the fact that the phospho-Ser/phospho-Thr ratio in the actual human phosphoproteome is far greater than that of phospho-Tyr ratio. This suggests that cellular proteins are routinely and randomly phosphorylated by different kinases with no biological significance, simply depending on the frequency of substrate encounters. Phosphatase is responsible for routinely removing these unwanted phosphate groups systematically and maintaining the dynamic equilibrium of physiological protein phosphorylation. Phos-tag SDS-PAGE visualized that the kinase reaction involves many incidental phosphorylation and that phosphatases play broader roles besides being strict counterparts to kinases.
1. Introduction
Protein phosphorylation plays a critical role in the regulation of fundamental cellular processes in all living cells [1]. The phosphorylation state of proteins is constantly maintained in dynamic equilibrium by the action of kinases and phosphatases [2,3,4]. Protein kinases are well characterized, whereas protein phosphatases have been less discussed. This may be because phosphorylation by kinases is a signaling switch-on reaction, whereas phosphatases are thought to have the passive function of signaling switch-off by dephosphorylation.
Previously, our phosphorylation analysis of intracellular proteins using Phos-tag two-dimensional fluorescence difference gel electrophoresis (2D DIGE) showed that many proteins switch to a hyperphosphorylated state in the presence of the Ser/Thr phosphatase inhibitor calyculin A [5]. Calyculin A is a cell-permeable inhibitor of protein phosphatase 1 (PP1) and protein phosphatase 2A (PP2A) which binds to the catalytic subunit of PP1 and PP2A [6]. The IC50s for PP1 and PP2A are 2 nM and 0.5–1 nM, respectively, with potent cytotoxicity at the nanomolar level [7]. Among the Ser/Thr phosphatases, PP1 and PP2A are ubiquitously and abundantly expressed in various types of eukaryotic cells and are involved in many of the universal biological activities, such as the cell cycle, metabolism, cytoskeletal regulation, ion channel and membrane receptor regulation, transcription, cell signaling, and cell differentiation [8]. 2D DIGE suggested that many proteins, despite routinely, randomly, and accidentally acting as substrates for various Ser/Thr kinases, are returned to their normal phosphorylation state by PP1 and PP2A, which are responsible for removing unintended phosphate groups. Mass spectrometry of gel spots after 2D DIGE showed that cytoskeletal proteins such as lamins, keratins, and vimentin are hyperphosphorylated by calyculin A [5]. While 2D DIGE only visualized proteins with high intracellular abundance, the present study used Phos-tag SDS-PAGE followed by immunoblotting to investigate the changes in phosphorylation states of a larger number of proteins with low intracellular abundance, such as proteins involved in signal transduction. In this study, we discuss the broader role of PP1 and PP2A, beyond simply switching off proteins activated by kinases.
To further discuss the role of Tyr phosphatase, changes in the phosphorylation state of intracellular proteins by the Tyr phosphatase inhibitor pervanadate were also examined in a similar manner. Protein tyrosine phosphorylation regulates cellular signaling pathways underlying a wide range of fundamental physiological processes [9]. Tyr kinases and Tyr phosphatases work in a coordinated manner to regulate reversible phosphorylation reactions that occur within seconds to minutes. The human genome encodes 90 Tyr kinases [2] and 107 putative protein Tyr phosphatases [10]. The almost identical number of Tyr kinase and Tyr phosphatase genes suggests similar levels of complexity between the two families. The diversity of Tyr phosphatases suggests that they are highly specific in function and substrate recognition in the regulation of signaling. Protein tyrosine phosphatases are a highly diverse family of enzymes, defined by the active-site signature motif His-Cys-X-X-X-X-X-Arg, in which the Cys acts as a nucleophile and is essential for catalysis [9]. In vivo Tyr phosphatase activity is irreversibly inhibited by pervanadate, which oxidizes Cys at the catalytic site [11]. In this paper, we discuss the role of Tyr phosphatases in contrast to the role of Ser/Thr phosphatases.
2. Results
2.1. Selection of Target Proteins
The total lysate of HeLa cells was subjected to conventional SDS-PAGE and Phos-tag SDS-PAGE followed by immunoblotting with different types of specific antibodies for cellular proteins. Twenty antibodies that specifically detected the protein of interest with an excellent signal-to-noise ratio were selected (Figure S1). Therefore, although the proteins analyzed in this study were randomly selected, they were categorized into the following nine types: (1) proteins related to the MAPK pathway, namely, A-Raf, ATF2, JNK1, MAPKAPK2, MEK1, p38 MAPK, and p42 MAPK; (2) proteins related to the JAK-STAT pathway, namely, JAK2, STAT1, STAT3, and STAT6; (3) proteins related to the mTOR pathway, namely, mTOR, Raptor, and Rictor; (4) a Wnt signaling pathway-related protein, β-catenin; (5) a cell cycle-related protein, CDK2, (6) a glycogen synthesis-related protein, GSK-3β; (7) a tumor suppressor protein, p53; (8) a phosphoinositide 3-kinase, PI3 kinase p110β; and (9) serum response factor, SRF. The number of phosphorylation sites and the estimated upstream kinases in human cells registered in the online database of post-transcriptional modifications, PhosphoSitePlus, for the 20 proteins are summarized in Table 1.

Table 1.
Number of potential phosphorylation sites and putative upstream kinases in human cells registered in PhosphoSitePlus database 1, for the 20 proteins analyzed.
2.2. Phosphorylation State of MAPK Pathway-Related Proteins in the Presence of the Phosphatase Inhibitor
The Phos-tag SDS-PAGE patterns of MAPK pathway-related proteins, namely, A-Raf, ATF2, JNK1/3, MAPKAPK2, MEK1, p38 MAPK, and p42 MAPK, in the presence of calyculin A or pervanadate are shown in Figure 1. Each blot is described below.
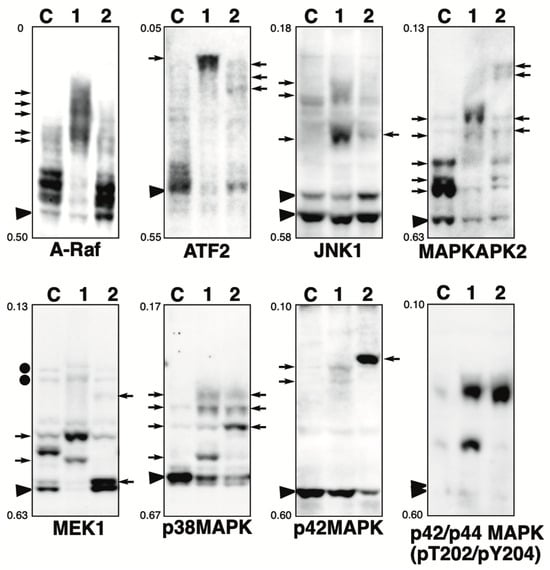
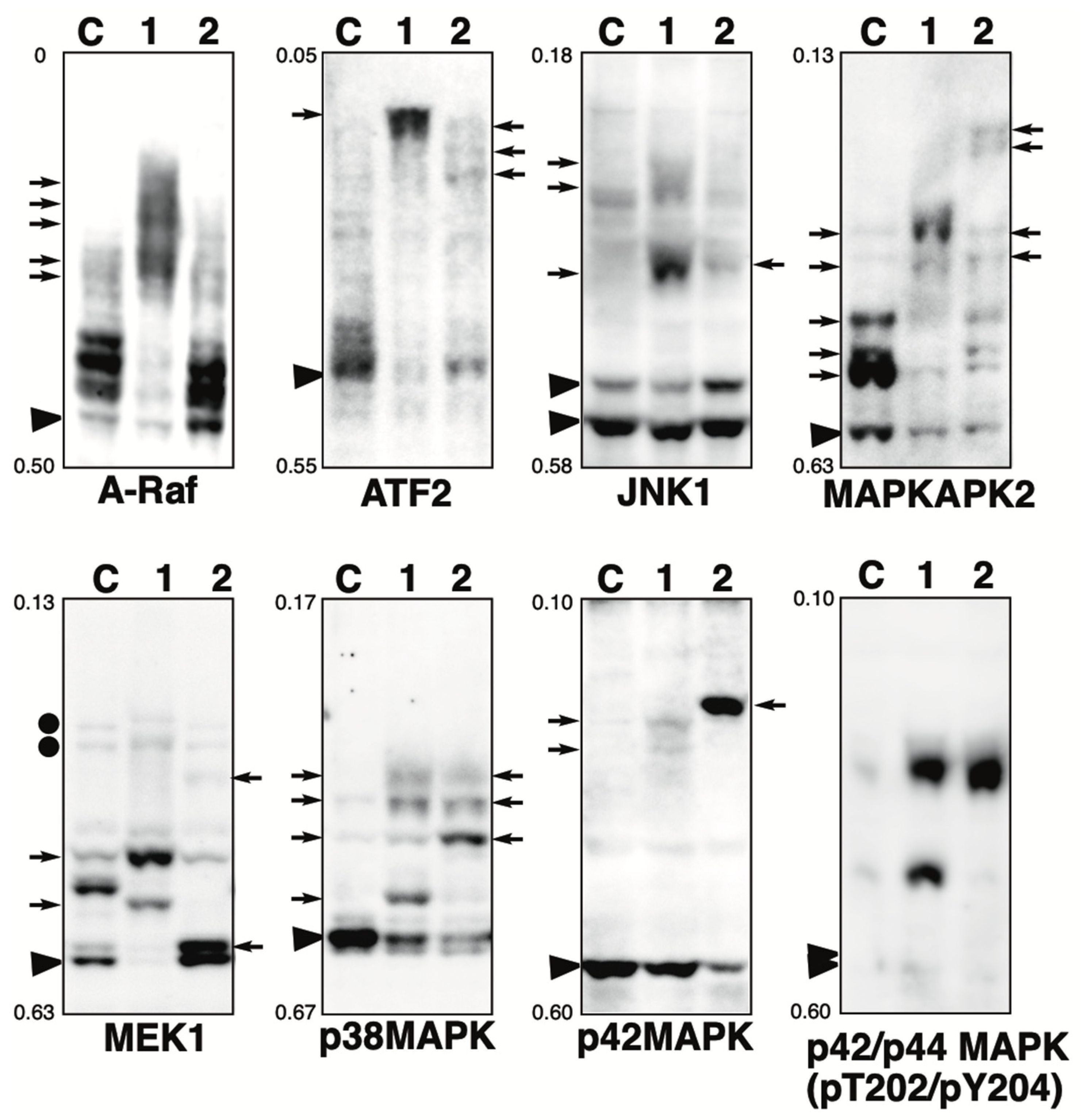
Figure 1.
Phosphorylation state of MAPK pathway-related proteins in the presence of phosphatase inhibitor. Total lysates of HeLa cells treated with calyculin A (lane 1) or pervanadate (lane 2) were subjected to Phos-tag SDS-PAGE together with untreated control lysate (lane C). The antibody used for the blot is indicated below each panel. Rf values indicating mobility are shown to the left of each panel. The arrowhead indicates the position of the non-phosphorylated form. The arrows on the left or right of each panel indicate newly observed bands or changes in band intensity in lane 1 or lane 2, respectively, compared to lane C. Phos-tag SDS-PAGE (8% w/v polyacrylamide, 20 µM Zn2+–Phos-tag) was performed using a neutral pH gel system buffered with Bis-Tris–HCl.
A-Raf: A-Raf, B-Raf, and C-Raf are the Ser/Thr kinases, which are the main effectors recruited by GTP-bound Ras to activate the MEK-MAPK pathway [12]. In untreated cells (lane C), multiple bands were detected, indicating that several sites are constitutively phosphorylated under homeostatic conditions. In calyculin A-treated cells (lane 1), only a few bands remained in the same position as in control cells, and several significantly up-shifted bands were detected. This suggests that multiple sites are phosphorylated by multiple Ser/Thr kinases. Putative upstream Ser/Thr kinases in vivo have not been deposited in PhosphoSitePlus (see Table 1). However, since A-Raf is similar in sequence and function to C-Raf, and several kinases are involved in the phosphorylation of the activation sites [13], it is likely that there are several upstream Ser/Thr kinases similar to C-Raf. In the pervanadate-treated cells (lane 2), the banding pattern was almost the same as in the control cells. This suggests that A-Raf is not phosphorylated by any Tyr kinases.
ATF2: cyclic AMP-dependent transcription factor 2 (ATF2) interacts with a variety of viral oncoproteins and cellular tumor suppressors and is a target of the SAPK/JNK and p38 MAP kinase signaling pathways [14]. ATF2 is phosphorylated by activated p38 MAP kinase. In untreated cells (lane C), multiple bands were detected, indicating that several sites are constitutively phosphorylated under homeostatic conditions. In cyclin A-treated cells (lane 1), few bands remained in the same position as in control cells, and one exaggeratedly shifted band was detected. This suggests that multiple sites are phosphorylated by multiple Ser/Thr kinases. In pervanadate-treated cells (lane 2), three up-shifted bands were detected. The Tyr phosphorylation sites were not registered in PhosphoSitePlus (Table 1), whereas the protein may contain potential substrates for multiple tyrosine kinases and phosphorylated by multiple Tyr kinases.
JNK1: Stress-activated protein kinase (SAPK)/Jun-amino-terminal kinase (JNK) is potently and preferentially activated by a variety of environmental stresses [15]. Activation of JNK1 occurs via the phosphorylation of Thr183/Tyr185 by MKK4 and MKK7 [16]. In untreated cells (lane C), distinct bands due to the non-phosphorylated 46 kDa and 54 kDa isoforms and several up-shifted low-intensity bands were detected. In the calyculin A-treated cells (lane 1), several newly appearing phosphorylated bands were detected. This suggests that multiple sites are phosphorylated by multiple Ser/The kinases. In the pervanadate-treated cells (lane 2), one new phosphorylated band was detected. JNK1 would be phosphorylated by several Tyr kinases, whereas the stoichiometry of Tyr phosphorylation at a particular site is less than 10%, as estimated from the intensity of the phosphorylated band in lane 2.
MAPKAPK2: MAP kinase-activated protein kinase 2 (MAPKAPK2) is rapidly phosphorylated by p38 MAPK and activated in response to cytokines, stress, and chemotactic factors [17]. Activation of MAPKAPK2 occurs through the phosphorylation of Thr222, Ser272, and Thr334 by p38 MAPK. In untreated cells (lane C), multiple bands were detected, indicating that several sites are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in control cells and two newly phosphorylated bands were detected. In the pervanadate-treated cells (lane 2), four newly appeared phosphorylated bands were detected. This suggests that MAPKAPK2 is phosphorylated by multiple Ser/The and Tyr kinases.
MEK1: Mitogen-activated ERK-regulated kinase 1 (MEK1) is a dual-specificity protein kinase that functions in an MAPK cascade to control cell growth and differentiation [18,19,20]. MEK1 is activated by the phosphorylation of Ser218/Ser222 by C-Raf [19,21]. In untreated cells (lane C), multiple bands were detected, indicating that multiple sites are constitutively phosphorylated under homeostatic conditions. This is consistent with our previous study using Phos-tag SDS-PAGE, which showed that multiple phosphorylation variants of MEK1 are constitutively present in typical human cells [22]. In the calyculin A-treated cells (lane 1), few bands remained at the position of the non-phosphorylated form, and several newly appeared phosphorylated bands were detected. In the pervanadate-treated cells (lane 2), a stronger signal was detected at a position just above the non-phosphorylated band, although several bands observed in the control cells appeared to be shifted down. The degree of migration in the Phos-tag gel is not defined by the number of phosphorylation sites but by the phosphorylation state, so this band pattern indicates the change in the phosphorylation state in the presence of pervanadate. This suggests that MEK1 is phosphorylated by several Ser/Thr and Tyr kinases. In our previous studies, we showed that MEK1 is phosphorylated by epidermal growth factor (EGF) treatment and that the intensity of the bands corresponding to active MEK1 (indicated by black circles on the left of the panel) increases, as detected by anti-phosphoMEK1 (Ser218/Ser222) [22]. The band patterns of lanes 1 and 2 were clearly different from the pattern after EGF stimulation, suggesting that they were phosphorylated by kinases unrelated to the activation of the MAPK pathway.
p38 MAPK: p38 MAPK is involved in a signaling cascade that controls cellular responses to cytokines and stress, similar to SAPK/JNK [17,23,24,25]. Activation of p38 MAPK occurs through the phosphorylation of Thr180/Tyr182 by MKK3, MKK4, and MKK6 [26]. Four isoforms of p38 MAPK have been identified: p38α, β, γ, and δ. In untreated cells (lane C), a clear band of the non-phosphorylated form and several up-shifted low-intensity bands were detected. Meanwhile, in the calyculin A-treated cells (lane 1), four distinct phosphorylated bands were detected. In addition, in the pervanadate-treated cells (lane 2), three phosphorylated bands were clearly detected. This suggests that specific sites are phosphorylated by multiple Ser/Thr and Tyr kinases.
p42 MAPK: The p44/42 MAPK signaling pathway can be activated in response to a variety of extracellular stimuli, including mitogens, growth factors, and cytokines [27]. Activation of p42 MAPK occurs through the phosphorylation of Thr183/Tyr185 by MEK1/2 [18,28]. In untreated cells (lane C), a non-phosphorylated band was detected. In the calyculin A-treated cells (lane 1), two up-shifted low-intensity bands were detected, while most proteins remained in the non-phosphorylated form. p42 MAPK is randomly phosphorylated by several Ser/Thr kinases, with the stoichiometry at a particular site being less than 10%, as estimated from the intensity of the phosphorylated band in lane 1. In the pervanadate-treated cells (lane 2), few bands remained in the same position as in the control cells, and one clearly up-shifted band was detected. This suggests that p42MAPK is phosphorylated by several Tyr kinases. Estimated from the intensity of the phosphorylated bands in lane 2, the stoichiometry of Tyr phosphorylation at a particular site is more than 50%. In our previous studies, we showed that p42 MAPK is phosphorylated by EGF treatment and that there is an increase in the intensity of the bands corresponding to active p42 MAPK, as detected by anti-phospho-p42/p44 MAPK (Thr202/Tyr204, Thr202, or Tyr204) [5]. Activation of p42 MAPK by treatment with calyculin A and pervanadate was examined by immunoblotting with an anti-phospho-p42/p44 MAPK (Thr202/Tyr204), as shown to the right of the p42 MAPK panel. Signals indicating the activation of p42 MAPK were detected in lanes 1 and 2, but at a different position from the phosphorylated band detected by anti-p42 MAPK in lane 2. This suggests that p42 MAPK is randomly phosphorylated in the presence of pervanadate by Tyr kinases unrelated to the MAPK pathway.
2.3. Effect of Hydrogen Peroxide Treatment on MAPKs
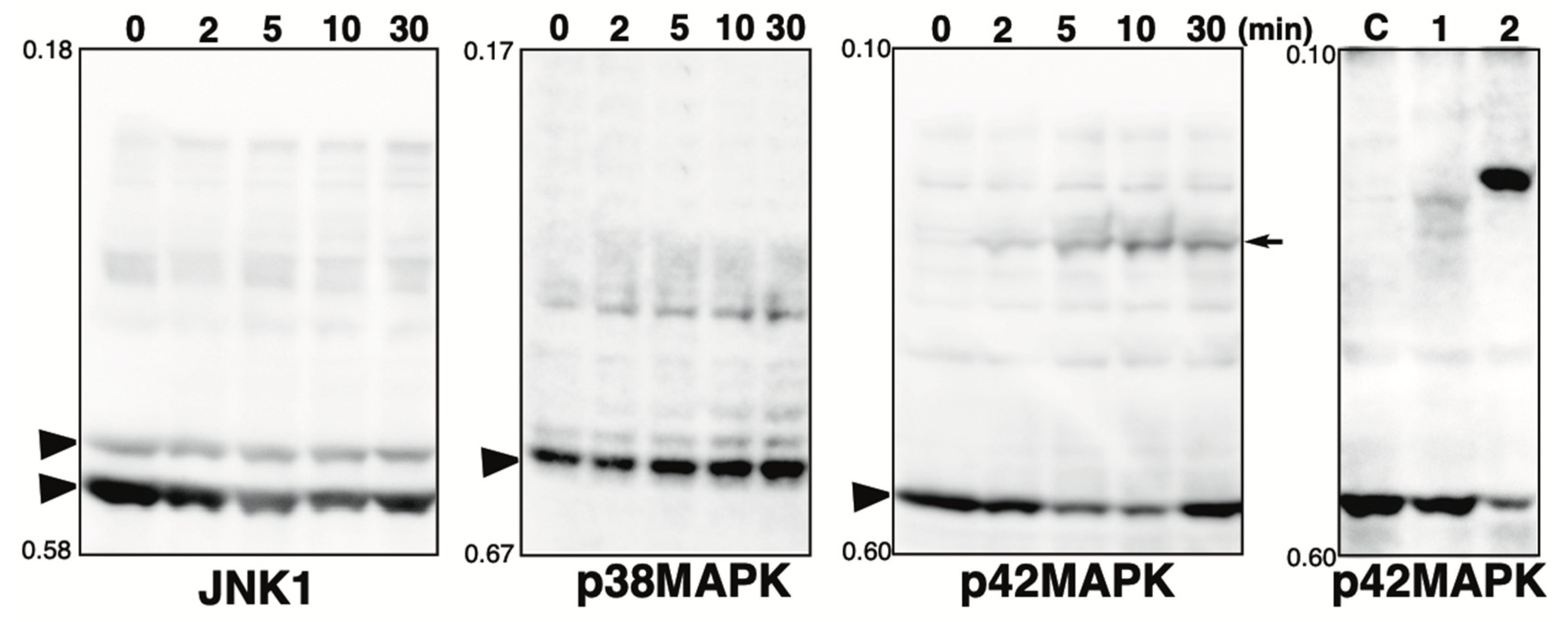
The MAPKs are known to be activated by oxidative stress, such as hydrogen peroxide [29]. Pervanadate, the tyrosine phosphatase inhibitor used in this study, was prepared as a mixture of vanadate and hydrogen peroxide. Changes in the phosphorylation state of MAPKs (p38, JNK1, and p42 MAPK) upon treatment with hydrogen peroxide alone were assessed by Phos-tag SDS-PAGE (Figure 2). The phosphorylation state of each protein was monitored from 0 to 30 min after the addition of hydrogen peroxide. No change in the phosphorylation state of JNK1 and p38MAPK was observed with hydrogen peroxide, whereas one phosphorylated band was detected for p42MAPK. The position of this band is different from that of the phosphorylated band observed for cells treated with pervanadate. For comparison, the same panel as that for p42 MAPK shown in Figure 1 is shown on the right. The results suggest that the changes in the Phos-tag pattern of MAPK in pervanadate-treated cells shown in Figure 1 reflect the inhibition of tyrosine phosphatase by pervanadate and not activation of the MAPK pathway by hydrogen peroxide. A-Raf, ATF2, MAPKAPK2, and MEK1 associated with the MAPK pathway may also be phosphorylated by hydrogen peroxide treatment, although, in the present study, it was assumed that the effect was not sufficient to significantly alter the Phos-tag pattern.
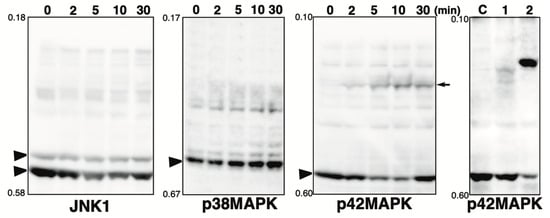
Figure 2.
Phos-tag pattern of MAPKs (JNK1, p38MAPK, and p42MAPK) treated with hydrogen peroxide. Total lysates of HeLa cells treated with 3 mM hydrogen peroxide (H2O2) for 0, 2, 5, 10, or 30 min were subjected to Phos-tag SDS-PAGE. The used antibody or the blot is indicated below each panel. Rf values indicating mobility are shown to the left of each panel. The arrowhead indicates the position of the non-phosphorylated form. The arrow indicates the newly appeared band phosphorylated by hydrogen peroxide treatment. For comparison, the panel of p42 MAPK shown in Figure 1 is presented at the extreme right.
2.4. Phosphorylation State of JAK-STAT Pathway-Related Proteins in the Presence of the Phosphatase Inhibitors
The Phos-tag SDS-PAGE patterns of JAK-STAT pathway-related proteins, namely, JAK2, STAT1, STAT3, and STAT6, in the presence of calyculin A or pervanadate are shown in Figure 3. Each blot is described below.
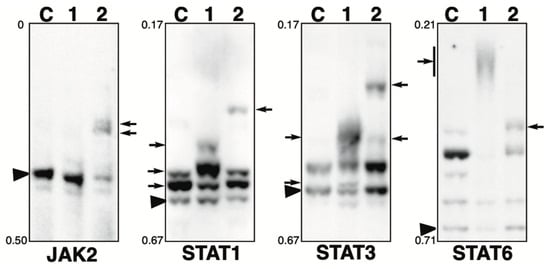
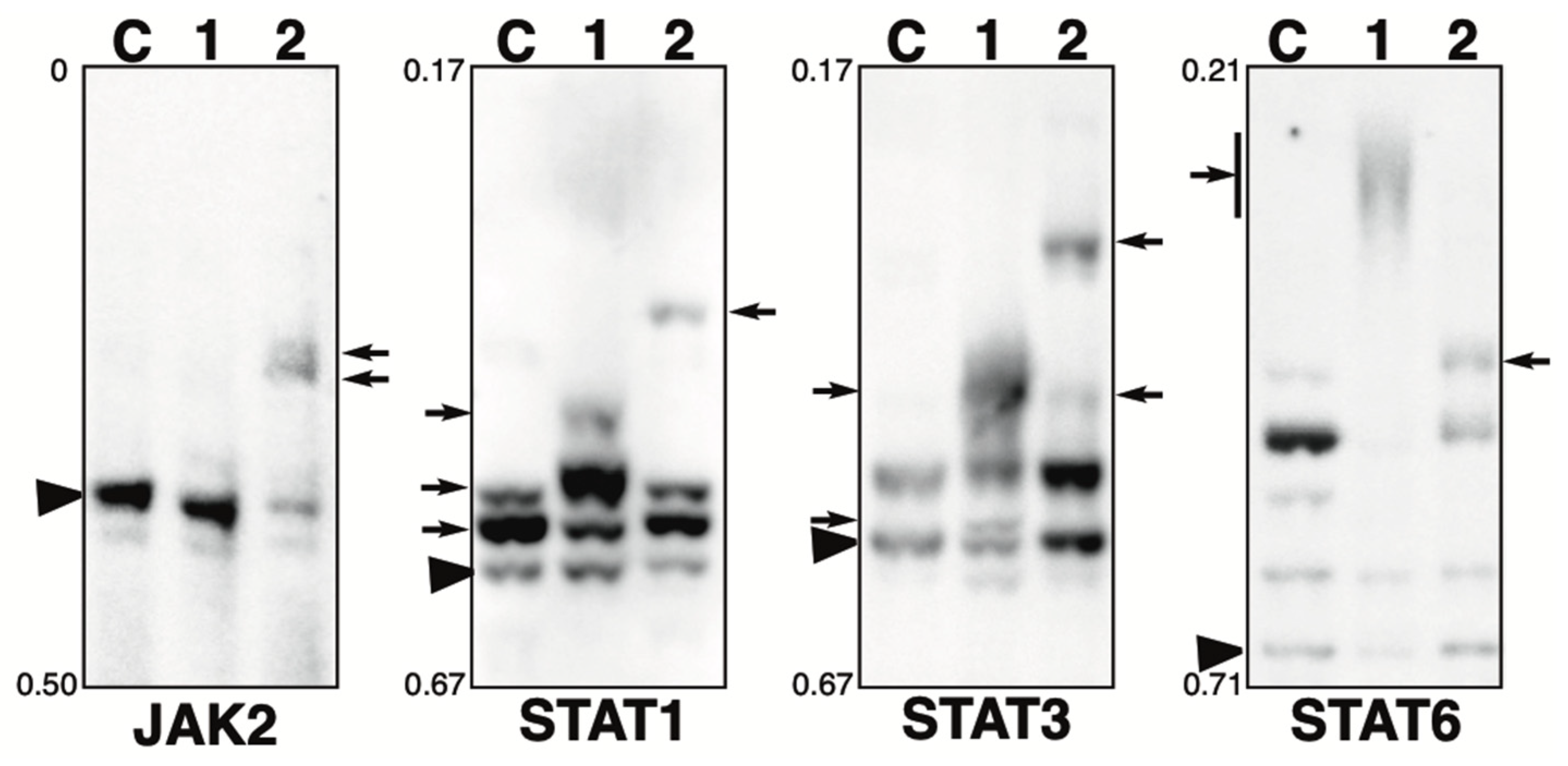
Figure 3.
Phosphorylation state of JAK-STAT pathway-related proteins in the presence of phosphatase inhibitor. Total lysates of HeLa cells treated with calyculin A (lane 1) or pervanadate (lane 2) were subjected to Phos-tag SDS-PAGE together with untreated control lysate (lane C). The antibody used for the blot is indicated below each panel. Rf values indicating mobility are shown to the left of each panel. The arrowhead indicates the position of the non-phosphorylated form. The arrows on the left or right of each panel indicate newly observed bands or changes in band intensity in lane 1 or lane 2, respectively, compared to lane C. Phos-tag SDS-PAGE (5% w/v polyacrylamide, 20 µM Zn2+–Phos-tag) was performed using a neutral pH gel system buffered with Tris-acetate for JAK2. Phos-tag SDS-PAGE (8% w/v polyacrylamide, 20 µM Zn2+–Phos-tag) was performed using a neutral pH gel system buffered with Bis-Tris HCl for STAT1, 3, and 6.
JAK2: JAK2 is a tyrosine kinase that is activated by ligand binding to a number of associated cytokine receptors [30]. Upon cytokine receptor activation, JAK2 is autophosphorylated at Tyr1007/Tyr1008 and phosphorylates its associated proteins such as STATs [31]. In untreated cells (lane C), most of the protein was detected in a non-phosphorylated state. In the calyculin A-treated cells (lane 1), little change was observed compared to the findings for control cells. Meanwhile, in the pervanadate-treated cells (lane 2), new phosphorylated bands were detected. Estimated from the intensity of the phosphorylated bands in lane 2, the stoichiometry of Tyr phosphorylation at a particular site is more than 50%. This suggests that JAK2 is phosphorylated by several Tyr kinases, whereas it is not phosphorylated by Ser/Thr kinases.
STAT1: STAT1 is a signal transducer and activator of transcription that mediates cellular responses to interferons, cytokines, and growth factors [32,33,34,35]. In untreated cells (lane C), one non-phosphorylated and two phosphorylated bands were detected, indicating that several sites are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), one newly appeared phosphorylated band was detected, and the intensity of two phosphorylated bands observed at the same position as for the control cells also changed, indicating a change in phosphorylation state. This suggests that several sites are phosphorylated by several Ser/Thr kinases. In the pervanadate-treated cells (lane 2), a newly phosphorylated band was detected. STAT1 is phosphorylated by several Tyr kinases, whereas the stoichiometry of Tyr phosphorylation at a particular site is less than 10%, as estimated from the intensity of the phosphorylated band in lane 2.
STAT3: STAT3 is a signal transducer and activator of transcription that mediates cellular responses to interferons, cytokines, and growth factors [32]. In untreated cells (lane C), one non-phosphorylated band and one phosphorylated band were detected. In the calyculin A-treated cells (lane 1), two new bands were detected. In the pervanadate-treated cells (lane 2), two new up-shifted bands were detected. This suggests that STAT3 is phosphorylated by multiple Ser/Thr and Tyr kinases.
STAT6: STAT6 is a signal transducer and activator of transcription involved in interleukin-mediated signaling [35,36]. In untreated cells (lane C), multiple bands were detected, indicating that several sites are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in control cells and smear-shifted bands were detected. In the pervanadate-treated cells (lane 2), a newly appeared phosphorylated band was detected. This suggests that STAT6 is phosphorylated by several Ser/Thr and Tyr kinases.
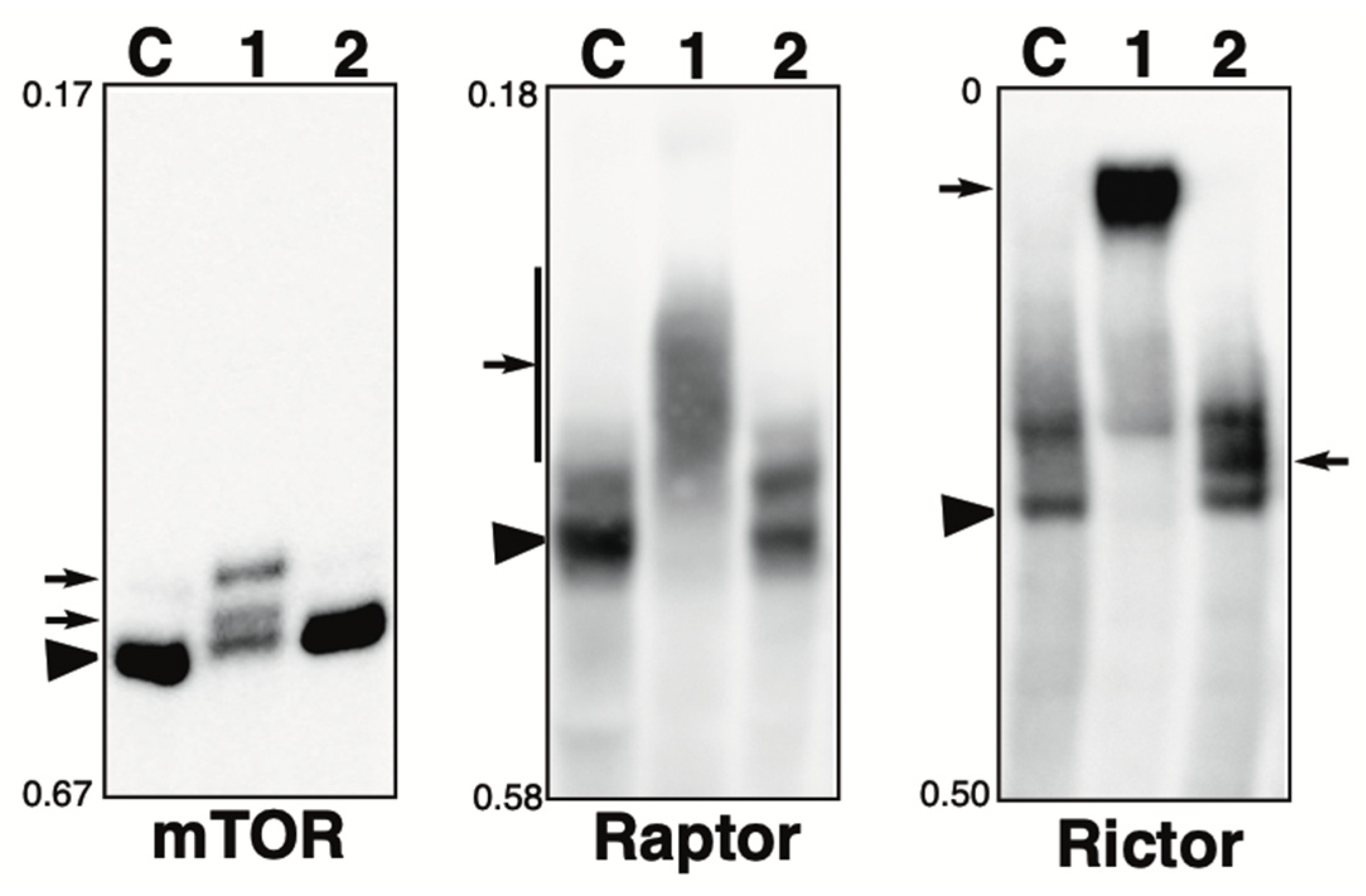
2.5. Phosphorylation State of mTOR Pathway-Related Proteins in the Presence of the Phosphatase Inhibitors
The Phos-tag SDS-PAGE patterns of mTOR pathway-related proteins, namely, mTOR, Raptor, and Rictor, in the presence of calyculin A or pervanadate are shown in Figure 4. Each blot is described below.
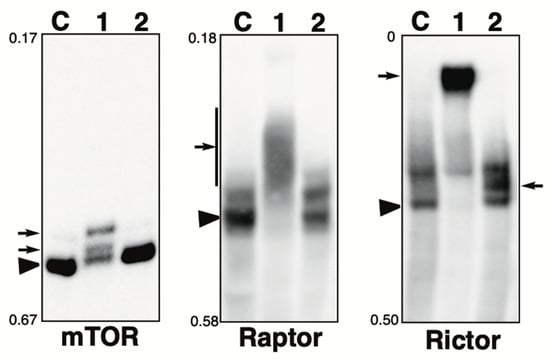
Figure 4.
Phosphorylation state of mTOR pathway-related proteins in the presence of phosphatase inhibitor. Total lysates of HeLa cells treated with calyculin A (lane 1) or pervanadate (lane 2) were subjected to Phos-tag SDS-PAGE together with untreated control lysate (lane C). The antibody used for the blot is indicated below each panel. Rf values indicating mobility are shown to the left of each panel. The arrowhead indicates the position of the non-phosphorylated form. The arrows on the left or right of each panel indicate newly observed bands or changes in band intensity in lane 1 or lane 2, respectively, compared to lane C. Phos-tag SDS-PAGE (3.5% w/v polyacrylamide, 20 µM Zn2+–Phos-tag) was performed using a neutral pH gel system buffered with Tris-acetate.
mTOR: A Ser/Thr kinase, a mammalian target of rapamycin (mTOR), functions as a sensor of ATP and amino acids to control cell growth [37]. When sufficient nutrients are available, mTOR is phosphorylated at the Ser2448 residue by the PI3kinase/Akt signaling pathway and then autophosphorylated at the Ser2481 residue. In untreated cells (lane C), most of the protein was detected in a non-phosphorylated state. In the calyculin A-treated cells (lane 1), two new phosphorylated bands were detected. In the pervanadate-treated cells (lane 2), the band pattern was almost the same as that of control cells. This suggests that mTOR is phosphorylated by several Ser/Thr kinases, whereas it is not phosphorylated by Tyr kinases. In our previous studies, we showed that mTOR was phosphorylated by treatment with fetal bovine serum (FBS) and that phosphorylated bands appeared, as detected by anti-phospho-mTOR (Ser2448 or Ser2481) [38]. The band patterns of lane 1 were clearly different from the pattern after FBS treatment, suggesting that they were phosphorylated by kinases unrelated to activation of the mTOR pathway.
Raptor: Raptor is an adaptor protein of mTOR and a member of the mTORC1 complex [39]. This complex promotes ribosome production and protein biosynthesis, suppresses proteolysis, and stimulates cell growth by providing information on nutrients, energy, and redox status [40]. The activity of mTORC1 is inhibited by rapamycin [41]. In untreated cells (lane C), one non-phosphorylated and one phosphorylated band were detected. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in the control cells, and smear-shifted bands were detected. Meanwhile, in the pervanadate-treated cells (lane 2), the band pattern was almost the same as in the control cells. This suggests that multiple sites are phosphorylated by multiple Ser/Thr kinases, whereas phosphorylation by Tyr kinases does not occur.
Rictor: Rictor is an adaptor protein of mTOR and a member of the mTORC2 complex. Similar to mTORC1, mTORC2 is regulated by growth factors and nutritional status and its activity is not inhibited by rapamycin [42]. In untreated cells (lane C), one non-phosphorylated band and one phosphorylated band were detected. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in the control cells, and an exaggeratedly up-shifted band was detected. In the pervanadate-treated cells (lane 2), a newly appeared phosphorylated band was detected. This suggests that Rictor is phosphorylated by several Ser/Thr and Tyr kinases.
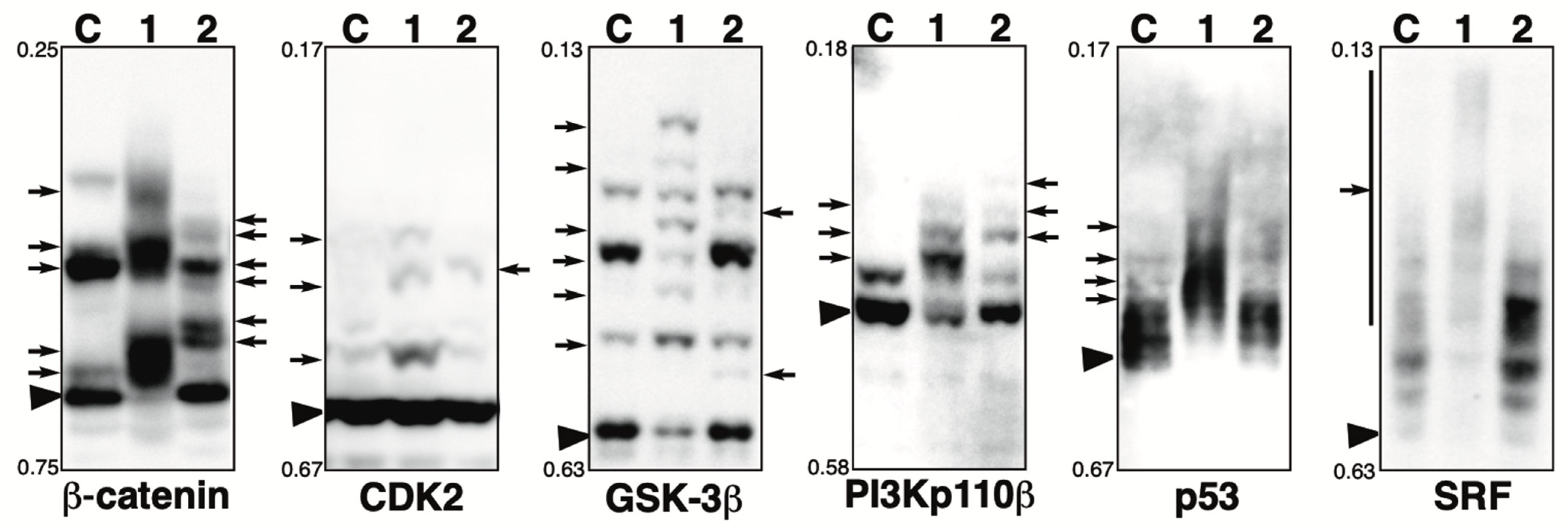
2.6. Phosphorylation State of Other Proteins in the Presence of the Phosphatase Inhibitor
The Phos-tag SDS-PAGE patterns of β-catenin, CDK2, GSK-3β, p53, PI3 kinase p110β, and SRF in the presence of calyculin A or pervanadate are shown in Figure 5. Each blot is described below.
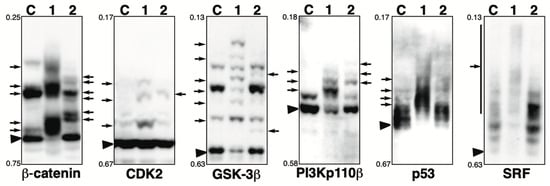
Figure 5.
Phosphorylation state of β-catenin, CDK2, GSK-3β, p53, PI3 kinase p110β, and SRF in the presence of phosphatase inhibitor. Total lysates of HeLa cells treated with calyculin A (lane 1) or pervanadate (lane 2) were subjected to Phos-tag SDS-PAGE together with untreated control lysate (lane C). The antibody used for the blot is indicated below each panel. Rf values indicating mobility are shown to the left of each panel. The arrowhead indicates the position of the non-phosphorylated form. The arrows on the left or right of each panel indicate newly observed bands or changes in band intensity in lane 1 or lane 2, respectively, compared to lane C. Phos-tag SDS-PAGE (8% w/v polyacrylamide, 20 µM Zn2+–Phos-tag) was performed using a neutral pH gel system buffered with Bis-Tris HCl.
β-Catenin: β-Catenin is a key downstream effector in the Wnt signaling pathway [43]. In untreated cells (lane C), multiple bands were detected, indicating that multiple sites are constitutively phosphorylated under homeostatic conditions. This is consistent with our previous study using Phos-tag SDS-PAGE, which showed that multiple phosphorylation variants of β-catenin are constitutively present in typical human cells [44]. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in control cells, and several newly appeared bands were detected due to changes in phosphorylation states. In the pervanadate-treated cells (lane 2), several newly appeared bands were also observed due to changes in phosphorylation states, whereas some bands remained in the same position as in the control cells. These findings suggest that β-catenin is phosphorylated by several Ser/Thr and Tyr kinases.
CDK2: Cyclin-dependent kinase 2 (CDK2) is an important component of the cell cycle machinery, and its kinase activity is regulated by its phosphorylation state [45]. In untreated cells (lane C), one phosphorylated band with low signal intensity was detected, whereas most proteins were unphosphorylated. In the calyculin A-treated cells (lane 1), three newly appeared phosphorylated bands were detected, whereas most proteins remained in the non-phosphorylated form. In the pervanadate-treated cells (lane 2), one newly appeared phosphorylated band was detected, whereas most proteins remained in the non-phosphorylated form. These findings suggest CDK2 is phosphorylated by several Ser/Thr and Tyr kinases, whereas the stoichiometry of random phosphorylation at a particular site is less than 10%, as estimated from the intensity of phosphorylated bands in lanes 1 and 2.
GSK-3β: Glycogen synthase kinase-3β (GSK-3β) regulates glycogen synthesis in response to insulin by phosphorylating and inactivating glycogen synthase [46]. In untreated cells (lane C), multiple bands were detected, indicating that several sites are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), several newly appeared phosphorylated bands were detected. This suggests that specific sites of GSK-3β are phosphorylated by several Ser/Thr kinases. In the pervanadate-treated cells (lane 2), two new phosphorylated bands were detected. These findings suggest GSK-3β is phosphorylated by several Tyr kinases, whereas the stoichiometry of Tyr phosphorylation at particular sites is less than 10%, as estimated from the intensity of newly appeared phosphorylated bands in lane 2.
PI3Kp110β: Phosphoinositide 3-kinase (PI3K) catalyzes the production of phosphatidylinositol-3,4,5-triphosphate. Growth factors and hormones trigger this phosphorylation event, which in turn coordinates cell growth, cell cycle entry, cell migration, and cell survival [47]. Four isoforms of the PI3K catalytic subunit, p110α, p110β, p110γ, and p110δ, have been identified. In untreated cells (lane C), one non-phosphorylated and one phosphorylated band were detected, indicating that several sites of PI3 kinase p110β are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in the control cells, and three new phosphorylated bands were detected. In the pervanadate-treated cells (lane 2), three new phosphorylated bands were detected. This suggests that specific sites of PI3 kinase p110β are phosphorylated by multiple Ser/Thr and Tyr kinases.
p53: The p53 tumor-suppressor protein plays an important role in the cellular response to DNA damage and other genomic aberrations. p53 is activated by phosphorylation to induce either cell cycle arrest and DNA repair or apoptosis [48,49,50]. In untreated cells (lane C), multiple bands were detected, indicating that several sites of p53 are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in the control cells, and multiple significantly up-shifted bands were detected. In pervanadate-treated cells (lane 2), the band pattern was almost the same as in control cells. This suggests that multiple sites of p53 are phosphorylated by multiple Ser/Thr kinases, whereas it is not phosphorylated by Tyr kinases.
SRF: Serum response factor (SRF) is a phospho-protein that, together with auxiliary factors, modulates the transcription of immediate early genes containing serum response elements at their promoters [51,52]. In untreated cells (lane C), multiple bands were detected, indicating that several sites of SRF are constitutively phosphorylated under homeostatic conditions. In the calyculin A-treated cells (lane 1), few bands remained in the same position as in the control cells, and smear-shifted bands were detected. In the pervanadate-treated cells (lane 2), the band pattern was almost the same as in the control cells. This suggests that multiple sites of SRF are phosphorylated by multiple Ser/Thr kinases, whereas it is not phosphorylated by Tyr kinases.
3. Discussion
The phosphorylation state of 20 types of intracellular proteins in the presence of the PP1 and PP2A inhibitor calyculin A or the tyrosine phosphatase inhibitor pervanadate was analyzed by Phos-tag SDS-PAGE followed by immunoblotting. The Ser/Thr phosphatases PP1 and PP2A which are specifically inhibited by calyculin A are ubiquitous and abundant phosphatases involved in universal cellular activity. Meanwhile, pervanadate is an irreversible inhibitor of the active center common to all Tyr phosphatases. All immunoblots showed a Phos-tag pattern indicating increased phosphorylation in the presence of one or both phosphatase inhibitors. This suggests that Ser/Thr and Tyr kinases constantly phosphorylate various proteins on a routine basis. Protein phosphorylation in the presence of these phosphatase inhibitors would be random and incidental with no biological significance due to the disordered kinase reaction with loss of reversibility.
In calyculin A-treated cells, phosphorylation was increased in all blots except JAK2 compared with the findings in control cells. Ten blots (A-Raf, ATF2, MAPKAPK2, MEK1, STAT6, Raptor, Rictor, β-catenin, PI3Kp110β, and SRF) showed a shift to the hyperphosphorylated state with few remaining bands showing the same phosphorylation state as control cells (Figure 1, Figure 3, Figure 4 and Figure 5). Two blots (p42 MAPK and CDK2) showed several new low-intensity phosphorylated bands, indicating a slight increase in phosphorylation (Figure 1 and Figure 5). In pervanadate-treated cells, phosphorylation was increased in 16 blots compared with the findings for control cells, except for A-Raf, mTOR, p53, and SRF (Figure 1, Figure 3 and Figure 5). Two blots (p42 MAPK and JAK2) showed a shift to a hyperphosphorylated state with few of the bands observed in control cells remaining (Figure 1 and Figure 2). Eight blots (JNK1, STAT1, STAT3, STAT6, Raptor, Rictor, CDK2, and GSK-3β) showed new low-intensity phosphorylated bands, indicating a slight increase in phosphorylation (Figure 1, Figure 3, Figure 4 and Figure 5).
The increase in phosphorylation stoichiometry per protein tended to be greater for Ser/Thr phosphatase inhibition than for Tyr phosphatase inhibition for the proteins analyzed in the 20 blots. This could be explained by the fact that the number of Ser/Thr kinases in the human genome is 428, which significantly outnumbers the 90 Tyr kinases [2,10]. Furthermore, based on proteomic data from more than 2000 phosphoproteins, the frequencies of pTyr, pThr, and pSer sites are 1.8%, 11.8%, and 86.4%, respectively [53]. Comparing the number of pSer/pThr sites with the number of pTyr sites registered in PhosphoSitePlus for the proteins analyzed in the 20 blots, the number of pSer/pThr sites is higher for all except JAK2 (Table 1). JAK2 alone had more pTyr sites (n = 23) than pSer/pThr sites (n = 6) and was phosphorylated at a higher stoichiometry by pervanadate treatment, whereas calyculin A treatment did not change its phosphorylation state (Figure 2). Considering our results from the number of Ser/Thr and Tyr kinase genes on the genome and the proteomic data, it is suggested that many kinase reactions are random, simply depending on the frequency of substrate encounters, irrespective of their biological significance. Recent advances in MS-based phosphoproteomics have enabled the measurement of extremely low-abundance phosphorylated proteins and have led to an increasing number of publications on studies involving high-throughput analysis of phosphorylation sites [54]. The biological significance and upstream kinases of most of these sites have not been determined, which could be explained by substantial incidental phosphorylation having been captured.
If many of the kinase reactions are random, phosphatase must perform systematic dephosphorylation to maintain proteins in a physiological phosphorylated state. Tyr phosphatases have been identified to be encoded by 107 genes, more than for Tyr kinases [10], suggesting that Tyr phosphatases are highly specific in function and substrate recognition in the regulation of signaling. For example, in the mechanism by which the MAPK pathway is inactivated, MAPK phosphatases, the dual-specificity Tyr phosphatases for pTyr and pSer/Thr, display distinct patterns of subcellular localization and specificity for individual MAPKs, thereby forming a phosphatase response network that functions in the attenuation of MAPK-dependent signaling pathways [55]. In this study, we have shown that p42 MAPK is highly phosphorylated at the Tyr which is not the site of phosphorylation upon activation of the MAPK pathway under conditions not regulated by any Tyr phosphatase (Figure 2). This suggests that the Tyr phosphatases are not only strict counterparts to individual Tyr kinases but also responsible for removing random unintended phosphate groups. The human genome contains 428 genes encoding Ser/Thr kinases, but far fewer, approximately 25, encoding Ser/Thr phosphatases have been identified [56]. A relatively small number of Ser/Thr phosphatases control many specific dephosphorylation reactions. In the case of PP1 and PP2A, this is explained by the formation of many different holoenzymes from a shared catalytic subunit and numerous regulatory subunits to achieve broad substrate specificity [57]. PP1 and PP2A have not been characterized as well as kinases due to their broad substrate specificity, but since the discovery of specific inhibitors such as calyculin A, it has become clear that they actively regulate specific protein functions. For example, the hyperphosphorylation of tau, a neuronal microtubule-binding protein, in Alzheimer’s disease is due to a defect in PP2A [58]. However, PP1 and PP2A must also be constantly working to remove random and unintended phosphate groups, as shown in this study. The stoichiometry of random Ser/Thr phosphorylation per protein would be far greater than that of Tyr phosphorylation. The broad substrate specificity achieved by the existence of several different holoenzymes from a small gene family makes logical sense for more diverse dephosphorylation reactions, in contrast to the Tyr phosphatase gene family.
In conclusion, we have discussed that kinase reactions involve a large amount of incidental phosphorylation, and that the physiological phosphorylation state of proteins is maintained by phosphatases removing these unwanted phosphate groups. This may seem like a waste of energy, but maintaining this dynamic equilibrium would allow biologically important reactions such as signal transduction to take place in the right place at the right time.
4. Materials and Methods
4.1. Materials
The acrylamide-pendant Phos-tag ligand (AAL-107) was purchased from Wako Pure Chemical Industries (Osaka, Japan). The HeLa cell line was obtained from the RIKEN BioResource Research Center (RIKEN BRC, Tsukuba, Japan). Calyculin A, sodium orthovanadate, and hydrogen peroxide were obtained from FUJIFILM Wako (Osaka, Japan). Anti-A-Raf antibody, anti-ATF2 antibody, anti-CDK2 antibody, anti-GSK-3β antibody, anti-CDK2 antibody, anti-JAK2 antibody, anti-CDK2 antibody, anti-MAPKAP2 antibody, anti-p38 MAPK antibody, anti-p42 MAP kinase antibody, anti-p53 antibody, anti-PI3 kinase p110β antibody, anti-Raptor antibody, anti-Rictor antibody, anti-mTOR antibody, anti-SRF antibody, anti-STAT1 antibody, anti-STAT3 antibody, and anti-STAT6 antibody were purchased from Cell Signaling Technology (Danvers, MA, USA). Anti-β-catenin antibody and anti-MEK1 antibody were purchased from BD Biosciences (San Jose, CA, USA). Anti-JNK1/3 antibody was purchased from Santa Cruz Biotechnology (Dallas, TX, USA).
4.2. Cell Culture and Phosphatase Inhibitor Treatment
HeLa cells were incubated in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% v/v fetal bovine serum, 100 units/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere of 5% CO2 and 95% air at 37 °C. For phosphatase treatment, 1 × 108 cells/2 mL were plated onto a dish for 12 h, and the cells were treated with calyculin A (100 nM) or pervanadate (0.1 mM) for 30 min. Pervanadate solution (10 mM) was prepared by adding 10 µL of 100 mM Na3VO4 and 30 µL of 300 mM H2O2 to 60 µL of water. The pervanadate solutions were used within 5 min. The cells were washed with Tris-buffered saline and immediately lysed with a sample-loading buffer for SDS-PAGE (65 mM Tris-HCl (pH 6.8), 1% w/v SDS, 10% v/v glycerol, 5% v/v 2-sulfanylethanol, and 0.03% w/v bromophenol blue).
4.3. Phos-Tag SDS-PAGE
Phos-tag SDS-PAGE for the analysis of proteins with a molecular weight below 100 kDa using a neutral-pH gel system buffered with Bis-Tris HCl was performed as previously reported [44]. Phos-tag SDS-PAGE for the analysis of high-molecular-weight proteins was performed using a neutral pH gel system buffered with Tris-acetate, as previously reported [5,59].
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/kinasesphosphatases2030014/s1, Figure S1: Immunoblotting analysis of whole lysate of HeLa cells with 20 types of antibodies.
Author Contributions
E.K.-K., conceptualization, methodology, resources, investigation, writing—original draft, writing—review and editing, and funding acquisition. K.N. (Kento Nishikawa), investigation. K.H., investigation. K.S., investigation. K.N. (Kenichi Nagase), writing—review and editing. E.K., writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Japan Society for the Promotion of Science (KAKENHI) Grants No. 21K06455 and No. 24K09710 to E.K.-K.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Acknowledgments
We thank the Natural Science Center for Basic Research and Development, Hiroshima University, Japan, for the use of its facilities (NBARD-00090).
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ATF | cyclic AMP-dependent transcription factor |
| CDK | cyclin-dependent kinase |
| DMEM | Dulbecco’s modified Eagle’s medium |
| EGF | epidermal growth factor |
| FBS | fetal bovine serum |
| GSK | glycogen synthase kinase |
| JAK | Janus kinase |
| JNK | Jun-amino-terminal kinase |
| MAPK | mitogen-activated protein kinases |
| MAPKAPK | mitogen-activated protein kinase-activated protein kinase |
| MEK | mitogen-activated ERK-regulated kinase |
| MKK | mitogen-activated protein kinase |
| mTOR | mammalian target of rapamycin |
| mTORC | mTOR complex |
| PI3 kinase | phosphoinositide 3-kinase |
| PP1 | protein phosphatase 1 |
| PP2A | protein phosphatase 2A |
| Raf | rapidly accelerated fibrosarcoma |
| SAPK | stress-activated protein kinase |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| STAT | signal transduction and activator of transcription |
| SRF | serum response factor |
| SUMO | small ubiquitin-related modifier |
| 2D DIGE | two-dimensional fluorescence difference gel electrophoresis |
References
- Arshad, O.A.; Danna, V.; Petyuk, V.A.; Piehowski, P.D.; Liu, T.; Rodland, K.D.; McDermott, J.E. An integrative analysis of tumor proteomic and phosphoproteomic profiles to examine the relationships between kinase activity and phosphorylation. Mol. Cell. Proteom. 2019, 18, S26–S36. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [PubMed]
- Turdo, A.; D’Accardo, C.; Glaviano, A.; Porcelli, G.; Colarossi, C.; Colarossi, L.; Mare, M.; Faldetta, N.; Modica, C.; Pistonek, G.; et al. Targeting Phosphatases and Kinases: How to Checkmate Cancer. Front. Cell Dev. Biol. 2021, 9, 690306. [Google Scholar] [CrossRef] [PubMed]
- Licheva, M.; Raman, B.; Kraft, C.; Reggiori, F. Phosphoregulation of the autophagy machinery by kinases and phosphatases. Autophagy 2022, 18, 104–123. [Google Scholar] [CrossRef]
- Kinoshita, E.; Kinoshita-Kikuta, E.; Koike, T. Phos-tag SDS-PAGE systems for phosphorylation profiling of proteins with a wide range of molecular masses under neutral pH conditions. Proteomics 2012, 12, 192–202. [Google Scholar] [CrossRef]
- Kita, A.; Matsunaga, S.; Takai, A.; Kataiwa, H.; Wakimoto, T.; Fusetani, N.; Isobe, M.; Miki, K. Crystal structure of the complex between calyculin A and the catalytic subunit of protein phosphatase 1. Structure 2002, 10, 715–724. [Google Scholar] [CrossRef]
- Suganuma, M.; Fujiki, H.; Furuya-Suguri, H.; Yoshizawa, S.; Yasumoto, S.; Kato, Y.; Fusetani, N.; Sugimura, T. Calyculin A, an inhibitor of protein phosphatases, a potent tumor promoter on CD-1 mouse skin. Cancer Res. 1990, 50, 3521–3525. [Google Scholar] [PubMed]
- Ceulemans, H.; Bollen, M. Functional diversity of protein phosphatase-1, a cellular economizer and reset button. Physiol. Rev. 2004, 84, 1–39. [Google Scholar] [CrossRef]
- Tonks, N.K. Protein tyrosine phosphatases: From genes, to function, to disease. Nat. Rev. Mol. Cell Biol. 2006, 7, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Sasin, J.; Bottini, N.; Friedberg, I.; Friedberg, I.; Osterman, A.; Godzik, A.; Hunter, T.; Dixon, J.; Mustelin, T. Protein tyrosine phosphatases in the human genome. Cell 2004, 117, 699–711. [Google Scholar] [CrossRef]
- Huyer, G.; Liu, S.; Kelly, J.; Moffat, J.; Payette, P.; Kennedy, B.; Tsaprailis, G.; Gresser, M.J.; Ramachandran, C. Mechanism of inhibition of protein-tyrosine phosphatases by vanadate and pervanadate. J. Biol. Chem. 1997, 272, 843–851. [Google Scholar] [CrossRef]
- Avruch, J.; Zhang, X.F.; Kyriakis, J.M. Raf meets Ras: Completing the framework of a signal transduction pathway. Trends Biochem. Sci. 1994, 19, 279–283. [Google Scholar] [CrossRef]
- Chong, H.; Lee, J.; Guan, K.L. Positive and negative regulation of Raf kinase activity and function by phosphorylation. EMBO J. 2001, 20, 3716–3727. [Google Scholar] [CrossRef] [PubMed]
- van Dam, H.; Wilhelm, D.; Herr, I.; Steffen, A.; Herrlich, P.; Angel, P. ATF-2 is preferentially activated by stress-activated protein kinases to mediate c-jun induction in response to genotoxic agents. EMBO J. 1995, 14, 1798–1811. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, H. From receptors to stress-activated MAP kinases. Oncogene 1999, 18, 6087–6093. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.; Cohen, P.; Trigon, S.; Morange, M.; Alonso-Llamazares, A.; Zamanillo, D.; Hunt, T.; Nebreda, A.R. A novel kinase cascade triggered by stress and heat shock that stimulates MAPKAP kinase-2 and phosphorylation of the small heat shock proteins. Cell 1994, 78, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Crews, C.M.; Alessandrini, A.; Erikson, R.L. The primary structure of MEK, a protein kinase that phosphorylates the ERK gene product. Science 1992, 258, 478–480. [Google Scholar] [CrossRef]
- Alessi, D.R.; Saito, Y.; Campbell, D.G.; Cohen, P.; Sithanandam, G.; Rapp, U.; Ashworth, A.; Marshall, C.J.; Cowley, S. Identification of the sites in MAP kinase kinase-1 phosphorylated by p74raf-1. EMBO J. 1994, 13, 1610–1619. [Google Scholar] [CrossRef]
- Rosen, L.B.; Ginty, D.D.; Weber, M.J.; Greenberg, M.E. Membrane depolarization and calcium influx stimulate MEK and MAP kinase via activation of Ras. Neuron 1994, 12, 1207–1221. [Google Scholar] [CrossRef]
- Zheng, C.F.; Guan, K.L. Activation of MEK family kinases requires phosphorylation of two conserved Ser/Thr residues. EMBO J. 1994, 13, 1123–1131. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, E.; Kinoshita-Kikuta, E.; Kubota, Y.; Takekawa, M.; Koike, T. A Phos-tag SDS-PAGE method that effectively uses phosphoproteomic data for profiling the phosphorylation dynamics of MEK1. Proteomics 2016, 16, 1825–1836. [Google Scholar] [CrossRef]
- Han, J.; Lee, J.D.; Bibbs, L.; Ulevitch, R.J. A MAP kinase targeted by endotoxin and hyperosmolarity in mammalian cells. Science 1994, 265, 808–811. [Google Scholar] [CrossRef]
- Lee, J.C.; Laydon, J.T.; McDonnell, P.C.; Gallagher, T.F.; Kumar, S.; Green, D.; McNulty, D.; Blumenthal, M.J.; Heys, J.R.; Land Vatter, S.W.; et al. A protein kinase involved in the regulation of inflammatory cytokine biosynthesis. Nature 1994, 372, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Freshney, N.W.; Rawlinson, L.; Guesdon, F.; Jones, E.; Cowley, S.; Hsuan, J.; Saklatvala, J. Interleukin-1 activates a novel protein kinase cascade that results in the phosphorylation of Hsp27. Cell 1994, 78, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Ando, K.; Uemura, K.; Kuzuya, A.; Maesako, M.; Asada-Utsugi, M.; Kubota, M.; Aoyagi, N.; Yoshioka, K.; Okawa, K.; Inoue, H.; et al. N-cadherin regulates p38 MAPK signaling via association with JNK-associated leucine zipper protein: Implications for neurodegeneration in Alzheimer disease. J. Biol. Chem. 2011, 286, 7619–7628. [Google Scholar] [CrossRef]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef] [PubMed]
- Seger, R.; Ahn, N.G.; Posada, J.; Munar, E.S.; Munar, E.S.; Jensen, A.M.; Cooper, J.A.; Cobb, M.H.; Krebs, E.G. Purification and characterization of MAP kinase activator(s) from epidermal growth factor stimulated A431 cells. J. Biol. Chem. 1992, 267, 14373–14381. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.; Xu, B.; Wright, A.; Vanderbilt, C.; Cobb, M.H.; Chen, Z.; Gibson, T.B.; Robinson, F.; Silvestro, L. MAP kinases. Chem. Rev. 2001, 101, 2449–2476. [Google Scholar]
- Leonard, W.J.; O’Shea, J.J. Jaks and STATs: Biological implications. Annu. Rev. Immunol. 1998, 16, 293–322. [Google Scholar] [CrossRef]
- Darnell, J.E. STATs and gene regulation. Science 1997, 277, 1630–1635. [Google Scholar] [CrossRef]
- Heim, M.H. The Jak-STAT pathway: Cytokine signalling from the receptor to the nucleus. J. Recept. Signal Transduct. Res. 1999, 19, 75–120. [Google Scholar] [CrossRef]
- Durbin, J.E.; Hackenmiller, R.; Simon, M.C.; Levy, D.E. Targeted disruption of the mouse Stat1 gene results in compromised innate immunity to viral disease. Cell 1996, 84, 443–450. [Google Scholar] [CrossRef]
- Meraz, M.A.; White, J.M.; Sheehan, K.C.; Bach, E.A.; Rodig, S.J.; Dighe, A.S.; Kaplan, D.H.; Riley, J.K.; Greenlund, A.C.; Campbell, D.; et al. Targeted disruption of the Stat1 gene in mice reveals unexpected physiologic specificity in the JAK-STAT signaling pathway. Cell 1996, 84, 431–442. [Google Scholar] [CrossRef]
- Nelms, K.; Keegan, A.D.; Zamorano, J.; Ryan, J.J.; Paul, W.E. The IL-4 receptor: Signaling mechanisms and biologic functions. Annu. Rev. Immunol. 1999, 17, 701–738. [Google Scholar] [CrossRef] [PubMed]
- Malabarba, M.G.; Rui, H.; Deutsch, H.H.; Chung, J.; Kalthoff, F.S.; Farrar, W.L.; Kirken, R.A. Interleukin-13 is a potent activator of JAK3 and STAT6 in cells expressing interleukin-2 receptor-gamma and interleukin-4 receptor-alpha. Biochem. J. 1996, 319, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Dennis, P.B.; Jaeschke, A.; Saitoh, M.; Fowler, B.; Kozma, S.C.; Thomas, G. Mammalian TOR: A homeostatic ATP sensor. Science 2001, 294, 1102–1105. [Google Scholar] [CrossRef]
- Kinoshita, E.; Kinoshita-Kikuta, E.; Ujihara, H.; Koike, T. Mobility shift detection of phosphorylation on large proteins using a Phos-tag SDS-PAGE gel strengthened with agarose. Proteomics 2009, 9, 4098–4101. [Google Scholar] [CrossRef]
- Hara, K.; Maruki, Y.; Long, X.; Yoshino, K.; Oshiro, N.; Hidayat, S.; Tokunaga, C.; Avruch, J.; Yonezawa, K. Raptor, a binding partner of target of rapamycin (TOR), mediates TOR action. Cell 2002, 110, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Sarbassov, D.D.; Ali, S.M.; King, J.E.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery. Cell 2002, 110, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, N.; Yoshino, K.; Hidayat, S.; Tokunaga, C.; Hara, K.; Eguchi, S.; Avruch, J.; Yonezawa, K. Dissociation of raptor from mTOR is a mechanism of rapamycin-induced inhibition of mTOR function. Genes Cells 2004, 9, 359–366. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Ali, S.M.; Kim, D.H.; Guertin, D.A.; Latek, R.R.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. Rictor, a novel binding partner of mTOR, defines a rapamycin-insensitive and raptor-independent pathway that regulates the cytoskeleton. Curr. Biol. 2004, 14, 1296–1302. [Google Scholar] [CrossRef] [PubMed]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, E.; Kinoshita-Kikuta, E. Improved Phos-tag SDS-PAGE under neutral pH conditions for advanced protein phosphorylation profiling. Proteomics 2011, 11, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Morgan, D.O. Principles of CDK regulation. Nature 1995, 374, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Welsh, G.I.; Wilson, C.; Proud, C.G. GSK3: A SHAGGY frog story. Trends Cell Biol. 1996, 6, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Cantley, L.C. The phosphoinositide 3-kinase pathway. Science 2002, 296, 1655–1657. [Google Scholar] [CrossRef]
- Levine, A.J. p53, the cellular gatekeeper for growth and division. Cell 1997, 88, 323–331. [Google Scholar] [CrossRef]
- Meek, D.W. Post-translational modification of p53. Semin. Cancer Biol. 1994, 5, 203–210. [Google Scholar] [PubMed]
- Milczarek, G.J.; Martinez, J.; Bowden, G.T. p53 Phosphorylation: Biochemical and functional consequences. Life Sci. 1997, 60, 1–11. [Google Scholar] [CrossRef]
- Treisman, R. Journey to the surface of the cell: Fos regulation and the SRE. EMBO J. 1995, 14, 4905–4913. [Google Scholar] [CrossRef]
- Janknecht, R.; Cahill, M.A.; Nordheim, A. Signal integration at the c-fos promoter. Carcinogenesis 1995, 16, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Olsen, J.V.; Blagoev, B.; Gnad, F.; Macek, B.; Kumar, C.; Mortensen, P.; Mann, M. Global, in vivo, and site-specific phosphorylation dynamics in signaling networks. Cell 2006, 107, 635–648. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, N.; Imamura, H.; Ishihama, Y. Large-scale discovery of substrates of the human kinome. Sci. Rep. 2019, 9, 10503. [Google Scholar] [CrossRef] [PubMed]
- Farooq, A.; Zhou, M.M. Structure and regulation of MAPK phosphatases. Cell. Signal. 2004, 16, 769–779. [Google Scholar] [CrossRef]
- Plowman, G.D.; Sudarsanam, S.; Bingham, J.; Whyte, D.; Hunter, T. The protein kinases of Caenorhabditis elegans: A model for signal transduction in multicellular organisms. Proc. Natl. Acad. Sci. USA 1999, 96, 13603–13610. [Google Scholar] [CrossRef] [PubMed]
- Yigong, S. Serine/Threonine Phosphatases: Mechanism through Structure. Cell 2009, 139, 468–484. [Google Scholar]
- Gong, C.X.; Grundke-Iqbal, I.; Iqbal, K. Dephosphorylation of Alzheimer’s disease abnormally phosphorylated tau by protein phosphatase-2A. Neuroscience 1994, 61, 765–772. [Google Scholar] [CrossRef]
- Kinoshita, E.; Kinoshita-Kikuta, E.; Koike, T. Separation and detection of large phosphoproteins using Phos-tag SDS-PAGE. Nat. Protoc. 2009, 9, 513–1521. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).