Abstract
As differences in seed dormancy between Actinidia species have not been reported previously, in this study we characterized the variation in the dormancy of seeds in 13 kiwifruit species that originated from different regions of China and Taiwan, and for which mature plants are now growing in New Zealand orchards. Dormancy-breaking treatments, including cold-moist stratification, seed coat scarification and soaking in water and gibberellic acid (GA3), were tested for their efficacy in alleviating dormancy and improving final germination and germination rates. In addition, we assessed seed viability using RNA integrity analysis to distinguish dead seeds from dormant seeds. This study identified that dormancy type in Actinidia seeds is species-specific and can be morphological, morphophysiological or a combination of physiological and physical, and that seed RNA integrity is a useful metric to incorporate into seed dormancy studies. Our results also suggest that species originating from colder climates that experience large differences between winter minimum and summer maximum temperatures exhibit physiological dormancy and require cold-moist stratification, contrasting with species originating in milder climates. Interestingly, although not all seeds from all the species were dormant, the proportion of dormant seeds in each species did not correlate to the climatic data of the region from which they originated. These findings provide new insights into mechanisms of seed dormancy in kiwifruit.
1. Introduction
The deciduous vine kiwifruit (Actinidia spp.) originated in China and neighboring countries and was introduced to the global market by New Zealand growers during the 1950s [1]. Kiwifruit has emerged as a high-value crop for its unique flavor, wide climatic adaptability and nutritional value [2]. Consequently, multiple breeding programs have been established, focusing on developing cultivars fit for both producers and consumers. This in turn has led to the rapid expansion of commercial kiwifruit-growing areas. In 2017, global kiwifruit production areas had surpassed 247,794 ha, with China (165,728 ha), Italy (26,403 ha) and New Zealand (11,705 ha) being the leading producers [3]. New Zealand’s exports of kiwifruit in 2023 made a substantial contribution to the country’s economy, at $NZ 2.6 billion [4], with the growth anticipated to continue. This sustained growth has been further fueled by industry resilience during the COVID-19 pandemic, driven by increased consumer preferences for health-oriented food selections and the fruit’s wide availability in supermarkets [5].
The New Zealand Kiwifruit Breeding Centre (KBC) was established in 2021 as a joint venture between the New Zealand Institute for Plant and Food Research Limited and Zespri Group Limited to support a large kiwifruit breeding program, utilizing genotypes from more than 20 kiwifruit species. Tens of thousands of seeds are germinated annually for the breeding program. Production of the required numbers of high-quality seedlings must be timed to match the outdoor planting season. However, this goal can be hindered by no, low or non-uniform seed germination, which are frequently associated with seed dormancy. Additionally, as kiwifruit is a clonally propagated crop new to cultivation, there has been little selection for uniform seed germination. Germination within and between families can be quite variable, with some seed families observed to require several spells of cool-moist stratification, i.e., a single 8–10 week stratification will result in some germination, with a subsequent period of cool-moist stratification resulting in more seedlings.
Seed dormancy is a physiological state that inhibits or prevents germination, even when environmental conditions are favorable. It also determines the range of environmental conditions necessary for germination to occur [6]. In contrast, a non-dormant seed can germinate across a wide range of favorable conditions [7,8]. A seed’s dormancy type is categorized into one or more of the main classifications: physiological dormancy (PD), morphological dormancy (MD), morphophysiological dormancy (MPD), physical dormancy (PY) and combinational dormancy (PY + PD) [7,8]. Currently, there is a general assumption that seeds of all kiwifruit species have physiological dormancy, with recommendations for germination typically involving cold-moist stratification at 2–4 °C with a 16 h low light (1–3 µmol m−2s−1) period over four to six weeks, which is supported by published data [9]. Data and observations on seed dormancy in kiwifruit have been previously reported, although the focus has been limited to Actinidia chinensis var. chinensis and A. chinensis var. deliciosa [10]. Thus, the questions remain of whether all kiwifruit species have dormant seeds and, if so, do they have the same dormancy type? As breeders explore more of the diversity in the genus, knowledge of the germination requirements for a wider range of species will be sought.
Interpreting the results of germination assays is complicated when working with dormant seeds, and particularly so for seeds that, like kiwifruit, are known to have poor vigor [10]. Should seeds fail to germinate, one may conclude the dormancy-breaking treatment was ineffective, or one may equally conclude the seeds were dead. Seed death resulting from deterioration or poor vigor, as opposed to physical damage, is currently considered to be a consequence of changes at the molecular level. Damage to lipids, proteins, DNA and RNA have all been associated with seed deterioration and death. Quantifying this damage in seeds that fail to germinate should differentiate between still-dormant seeds, with little-to-no damage, and dead seeds, markedly damaged. In particular, quantifying damage to RNA has been demonstrated as an effective method for rapidly determining seed health, independent of its ability to germinate [11].
This study investigated variations in seed dormancy for 13 kiwifruit species by testing the effects of different dormancy-breaking treatments (DBTs) on both seed germination and vigor (rate of germination). This study additionally provided an opportunity to evaluate the use of RNA integrity analysis for distinguishing dormant and dead seeds. This study also investigated if New Zealand-grown plants of the species tested retain seed dormancy requirements based on their geographic origin, or if these requirements are more plastic and perhaps reflect their maternal growing environments. A previous study has identified environmental maternal effects as significant factors explaining the variation in germination timing and dormancy of different plant species [12]. There is no relevant information available in the literature on the geographic distribution of dormancy in natural kiwifruit species populations. Although we do not have directly comparable data for dormancy classification, we hypothesized that the kiwifruit species that originated in temperate, cold and elevated environments would display physiological dormancy and would require cold stratification to break their dormancy. By contrast, species that originated from tropical (hot and humid) environments would be expected to show mild or no dormancy.
This study is expected to provide knowledge to improve the efficiency of seedling propagation by increasing the reliability of germination.
2. Materials and Methods
2.1. Seed Source
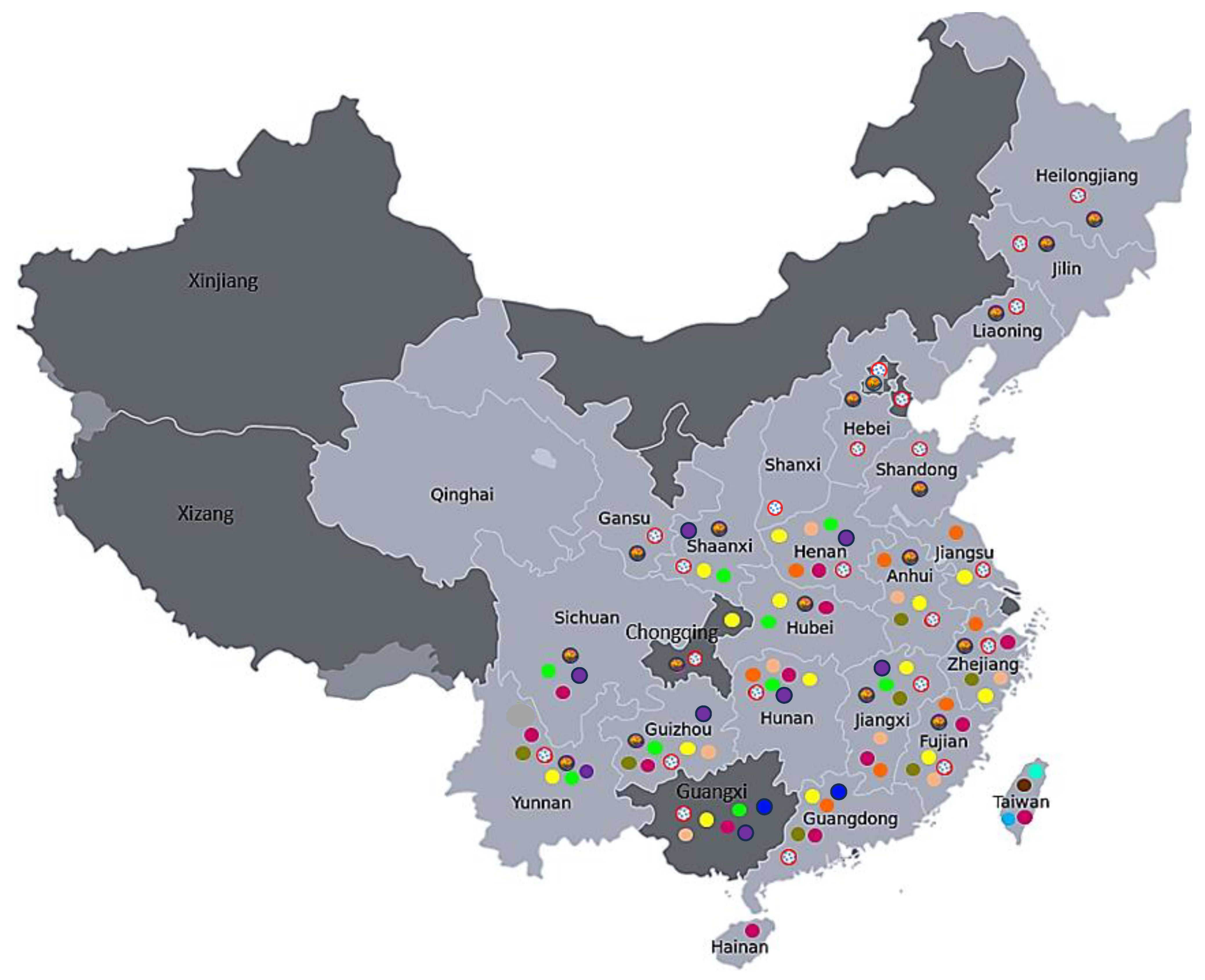
Ripened fruit from open-pollinated flowers of 13 kiwifruit species (Table 1) were sourced from Plant and Food Research orchards located in Motueka (South Island) and Te Puke (North Island), New Zealand in May 2018. The origins of the species used in this study are distributed in various regions of China and Taiwan, ranging from the warm climate in the south and southwest to the cold climate in the north and northeast ([13,14]; Figure 1; Table 2). Where available, the climatic data (minimum, maximum and mean annual temperature) for these geographic origins are summarized in Table 2 and compared with the same data for two New Zealand growing regions (Te Puke in the North Island and Motueka in the South Island).

Table 1.
List of the Actinidia species (and species codes) in which seed dormancy was investigated. The seeds were extracted from ripened fruit collected from Plant and Food Research orchards and derived from open pollination.
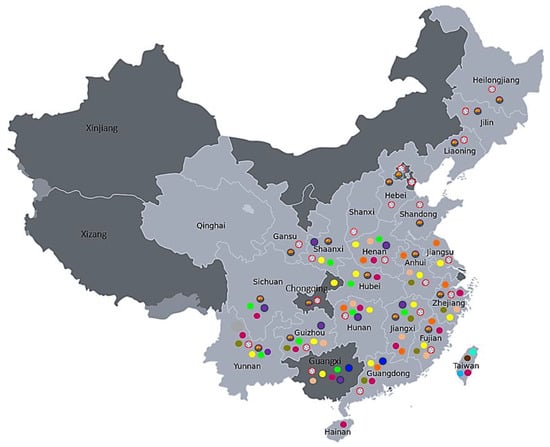
Figure 1.
Origin and geographic distribution of the 13 kiwifruit (Actinidia) species investigated in this study (using data compiled from [13,14]). AA (A. arguta var. arguta)  , AE (A. arguta)
, AE (A. arguta)  , CK (A. chinensis var. chinensis)
, CK (A. chinensis var. chinensis)  , DA (A. chinensis var. deliciosa)
, DA (A. chinensis var. deliciosa)  , DD (A. deliciosa var. coloris)
, DD (A. deliciosa var. coloris)  , EA (A. eriantha)
, EA (A. eriantha)  , GD (A. fortunatii)
, GD (A. fortunatii)  , LB (A. lanceolata)
, LB (A. lanceolata)  , LC (A. latifolia)
, LC (A. latifolia)  , PC (A. polygama)
, PC (A. polygama)  , RE (A. rufa)
, RE (A. rufa)  , SB (A. setosa)
, SB (A. setosa)  , VA (A. valvata)
, VA (A. valvata)  . China provinces map downloaded from free access File:China_provinces_highlighted.svg—Wikiwand. (https://www.wikiwand.com/en/File:China_provinces_highlighted.svg, accessed on 1 November 2023).
. China provinces map downloaded from free access File:China_provinces_highlighted.svg—Wikiwand. (https://www.wikiwand.com/en/File:China_provinces_highlighted.svg, accessed on 1 November 2023).
, AE (A. arguta) , CK (A. chinensis var. chinensis) , DA (A. chinensis var. deliciosa) , DD (A. deliciosa var. coloris) , EA (A. eriantha) , GD (A. fortunatii) , LB (A. lanceolata) , LC (A. latifolia) , PC (A. polygama) , RE (A. rufa) , SB (A. setosa) , VA (A. valvata) . China provinces map downloaded from free access File:China_provinces_highlighted.svg—Wikiwand. (https://www.wikiwand.com/en/File:China_provinces_highlighted.svg, accessed on 1 November 2023).
Table 2.
Annual minimum (min), maximum (max) and mean temperatures, and climate description for the geographic regions in which the 13 Actinidia species are found ([13,14]; Climate data source: chinamaps.org and cliflo.niwa.co.nz, accessed on 1 November 2023); alt. = altitude; lat. = latitude.
2.2. Seed Extraction, Sterilization, Drying and Temporary Storage
Seeds were extracted from vine-ripened fruit using Lafazym® extract (Laffort, France) solution (5 g/L) by soaking the fruit pulp over a period of 24 h at room temperature. The seeds were then separated from the undigested fruit pulp by sieving with tap water, taking care to ensure all fruit pulp was removed from the seed. The rinsed seeds were surface-sterilized by washing in a 1% (v/v) sodium hypochlorite (Janola™) solution for 20 min at room temperature, followed by three rinses with sterile reverse osmosis (RO) water. The sterilized seeds were then air dried on filter paper in a laminar flow hood. The seeds were stored in paper envelopes and kept in a refrigerator at 5 °C until used. The initial seed moisture content of the fresh seeds, calculated based on the ISTA Standards [16], was c. 12% (fresh-weight basis).
2.3. Initial Seed Germination
Initial seed germination (“Fresh control” in Table 3) was carried out on fresh seeds immediately following seed extraction and surface sterilization. Seeds were germinated on water agar (7 g/L agar) in Petri dishes at either constant (24 °C) or alternating temperature (24/12 °C), both with a 16 h photoperiod at a photosynthetic flux density of 30–50 μmol s−1 m−2 provided by cool-white fluorescent tubes. Seeds were considered germinated when a complete seedling was formed with full opening of both cotyledonary leaves. Germination results were recorded twice a week over an 8 week period. Seeds that germinated in the initial germination test were considered not dormant and seeds that did not germinate were considered as potentially dormant. Nevertheless, all the species were further evaluated for germination following various DBTs as described below.
Table 3.
Description of dormancy-breaking treatments (DBTs) investigated in this study.
2.4. Germination Rate Assessments
In addition to the final germination percentage assessment, we also assessed the germination rate of the seeds using the following two criteria: mean germination time (MGT) and germination rate index (GRI). The MGT and GRI were calculated based on the method described in Kader 2005 [17]. A lower MGT indicates faster germination of a seed population, whereas the GRI reflects the percentage of germination on each day of the germination period. Higher GRI values indicate higher and faster germination [17].
2.5. Dormancy-Breaking Treatments
The DBTs were applied to both non-stratified (fresh) and stratified seeds to break PD, PY or combinations of both dormancy types. These treatments are summarized in Table 3. All seeds that received DBTs were germinated at alternating temperatures and evaluated for germination on water agar as described previously.
2.6. Seed Vigor Assessment
In parallel to assessing the final germination, we also assessed the seed vigor by comparing the number of seeds with radicle emergence with the number of seeds that completed germination, i.e., full seedling formation (radicle and shoot with both cotyledonary leaves). This assessment was carried out on species that showed dormancy, irrespective of the DBTs tested, i.e., by calculating the cumulative number of seeds with radicle emergence and germination out of the total number of seeds used for the entire experiment.
2.7. RNA Fragmentation Analysis
The RNA integrities of the seeds of three species, GD to represent dormant seeds, EA to represent non-dormant seeds, and A. chrysantha as dead seeds (control), were assessed at the National Laboratory for Genetic Resources Preservation, USDA, Fort Collins, CO, USA. For each species, 5–10 surface-sterilized dry seeds were ground in a Qiagen TissueLyser (Hilden, Germany) for 1 min in liquid nitrogen at 30 oscillations/s. RNA was extracted from the ground tissue with a Qiagen Plant RNeasy kit (Hilden, Germany) as per the recommendations, but modified by repeating the final wash with Buffer RPE to reduce guanidine hydrochloride carry-over. RNA purity was assessed with a NanoDrop 1000 spectrophotometer (Thermo Fisher, Wilmington, DE, USA), and RNA integrity was quantified for each sample using an Agilent Bioanalyzer (Waldbronn, Germany) running RNA 6000 Pico chips and the Plant RNA Pico assay (software version B.02.08.SI648 R3) according to the manufacturer’s protocols. Agilent 2100 software was used to draw electropherogram baselines and assess peak areas of different fragment sizes [11]. RNA Integrity Numbers (RINs) ranging from 0 to 10 were calculated as described by Schroeder et al. [18], with 0 being completely degraded and 10 being completely intact.
2.8. Data Analysis
The proportions of seeds that germinated from each species and treatment were analyzed using a binomial generalized linear model (GLM) with a logit link. Specific comparisons among treatments were made using contrasts. All analysis was carried out using Genstat 21st edition (VSN International Ltd., Hemel Hempstead, UK).
3. Results
3.1. Initial Seed Germination
For the initial seed germination, we compared two temperature regimes, i.e., constant (24 °C) or alternating temperature (24/12 °C). The results indicated that most species (except AA and EA) showed some germination in alternating temperatures while none of these species (except for LB with 5% germination) germinated in constant temperature (Table 4). RE was the only species with significant germination (around 85%), indicating this species may not be dormant.
Table 4.
Germination (%) of fresh seeds from open-pollinated fruit of 13 kiwifruit species germinated using two temperature regimes: constant (24 °C) or alternating temperature (24/12 °C), both with a 16 h photoperiod. *** p < 0.001 for comparison between germination at Constant and Alternating temperatures. Species codes are spelled out in Table 1.
3.2. Germination following Dormancy-Breaking Treatments
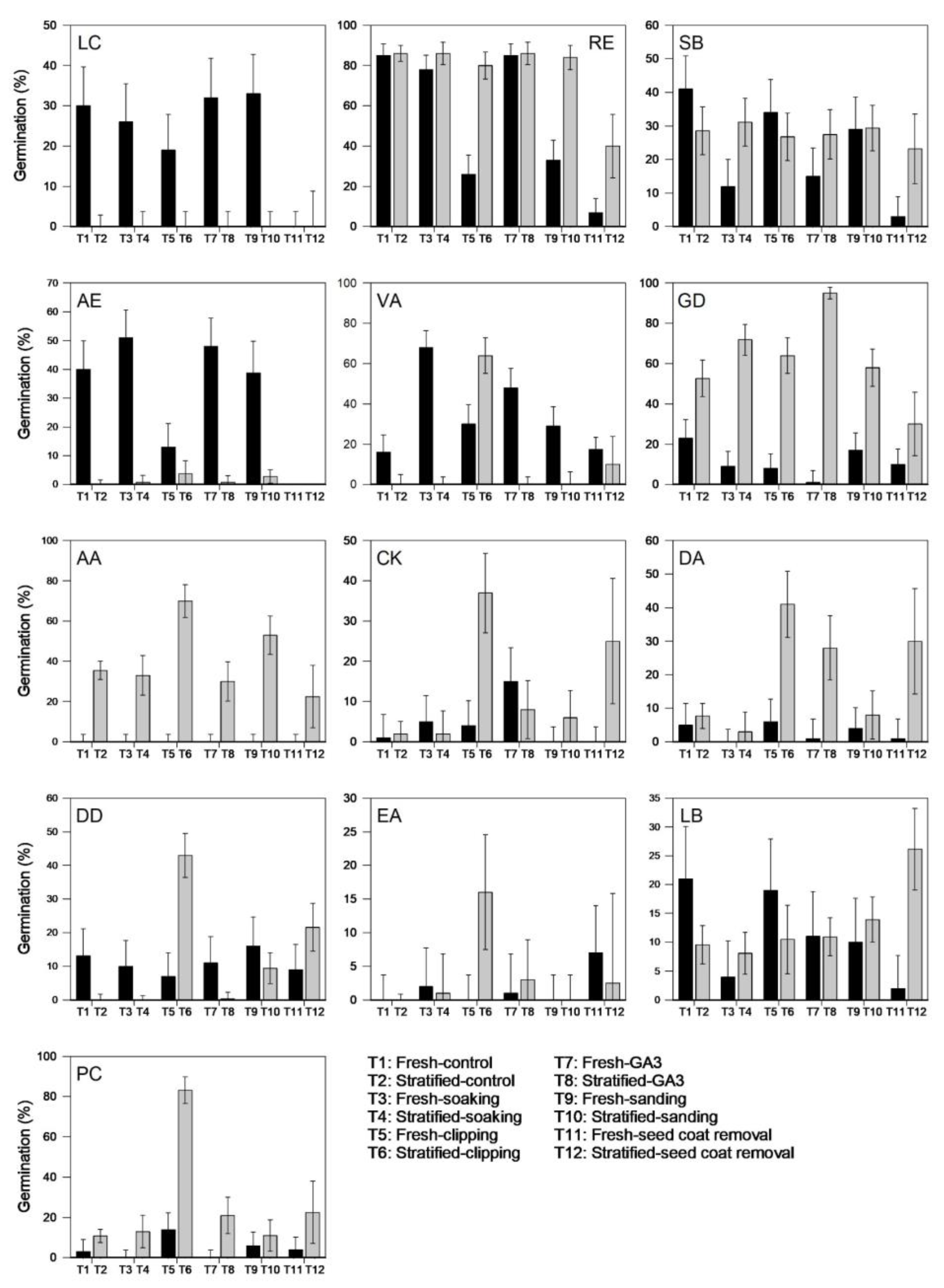
We assessed germination for all the species following the various DBTs using an alternating (24/12 °C) temperature regime and with or without prior stratification. As predicted, germination of RE seeds did not improve with the application of the DBTs and therefore the seeds of this species are confirmed as non-dormant (Table 5). Two species, AE and VA, produced their highest germination rates (51% and 68%, respectively) without stratification following soaking in water for 24 h, suggesting that these species exhibit only morphological dormancy. Although RE also had relatively high germination (c. 78%) after 24 h soaking in water, the germination percentage was lower than that achieved for the control treatment, i.e., 85%. The remaining species all had improved germination rates following cold-moist stratification. Nevertheless, stratification alone did not have a significant positive impact on most species, and the best germination results were achieved when stratification was carried out in combination with other DBTs. Apart from GD and LB seeds, which required stratification in combination with GA3 and seed coat removal, respectively, for their best germination, the remaining species produced their highest germination rates following combined stratification and clipping treatments (Table 5). It is important to note that seeds of EA consistently exhibited poor germination, which we attributed to either poor seed quality or seed immaturity. Figure 2 shows the species grouped into dormant and non-dormant species; the dormant species additionally are classified into different dormancy classes based on their responses to the various DBTs.
Table 5.
Final Germination Percentage (FGP, %) from the six fresh seed dormancy-breaking treatments on 13 kiwifruit species incubated in an alternating temperature regime. For each species and stratification combination, treatment comparisons were made with the control (no dormancy-breaking treatment): † p < 0.10, * p < 0.05, ** p < 0.01, *** p < 0.001 (significance indicators beside each mean). Additionally, for each species and dormancy-breaking treatment, comparisons were made between fresh and stratified seeds (significance indicators above the means).
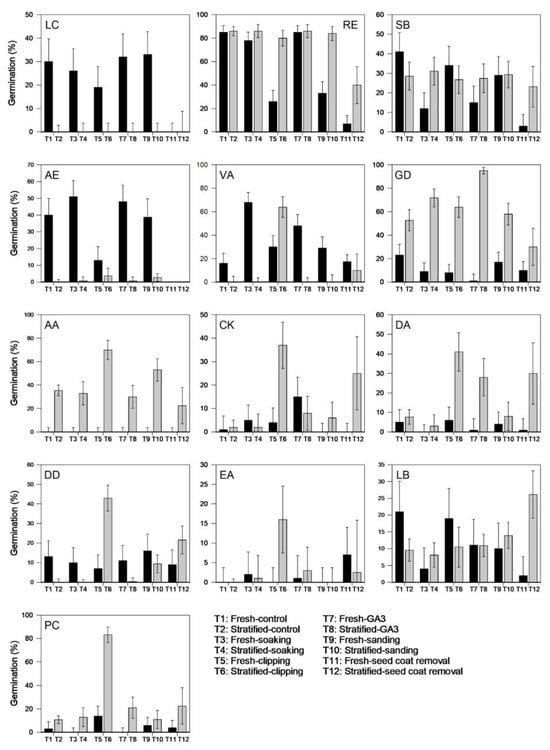
Figure 2.
Germination (%) of 13 kiwifruit species following various dormancy-breaking treatments. Seeds were germinated on water agar at alternating temperatures (24/12 °C) with a 16 h photoperiod. LC, RE and SB with no dormancy; AE and VA with morphological dormancy (MD); GD with morpho-physiological dormancy (MDP); AA, CK, DA, DD, EA, LB and PC with combined physiological and physical dormancy (PY + PD). Error bars are 95% confidence intervals. Species codes are spelled out in Table 1. GA3 = gibberellic acid.
3.3. Assessment of Germination Rate following Dormancy-Breaking Treatments
In addition to identifying treatments with the highest final germination percentages for each species, further evaluation was made to assess the rate of germination by calculating their mean germination time (MGT) and germination rate index (GRI) (Supplementary Table S1). The most effective (best) DBTs for each species, indicated by the highest final germination percentage, lowest MGTs and highest GRIs, are summarized in Table 6. Apart from AE, the rest of the species each had one best DBT. For AE, where the highest germination rate was not represented by the lowest MGT and highest GRI, we found that the second highest germination rate corresponded with the lowest MGT and highest GRI. Hence, for AE, dormancy can be alleviated by soaking in either water or GA3 for 24 h. We also calculated the proportion of dormant seeds as the difference in percentages of seed germination between the fresh control treatment and the best DBT for each of the species (Table 6). AA, GD and PC had the highest proportion (≥70%) of dormant seeds, while for VA, about half the seeds were dormant. For CK, DA and DD, about one third of the seeds was dormant, followed by EA (c. 16%), with AE and LB having the lowest proportions of dormant seeds (<10%) (Table 6).
Table 6.
The most effective seed dormancy-breaking treatment for the 10 dormant kiwifruit (Actinidia) species based on highest final germination percentage, lowest mean germination time (MGT) and highest germination rate index (GRI). Values in brackets are the standard errors of the means. Species codes are spelled out in Table 1. DBT: dormancy-breaking treatment, FGP: final germination percentage, MGT: mean germination time, GRI: germination rate index. PD = physiological dormancy; PY = physical dormancy; MD = morphological dormancy; MPD = morpho-physiological dormancy. GA3 = gibberellic acid.
3.4. Seed Vigor Assessment Comparing Radicle Emergence and Final Germination
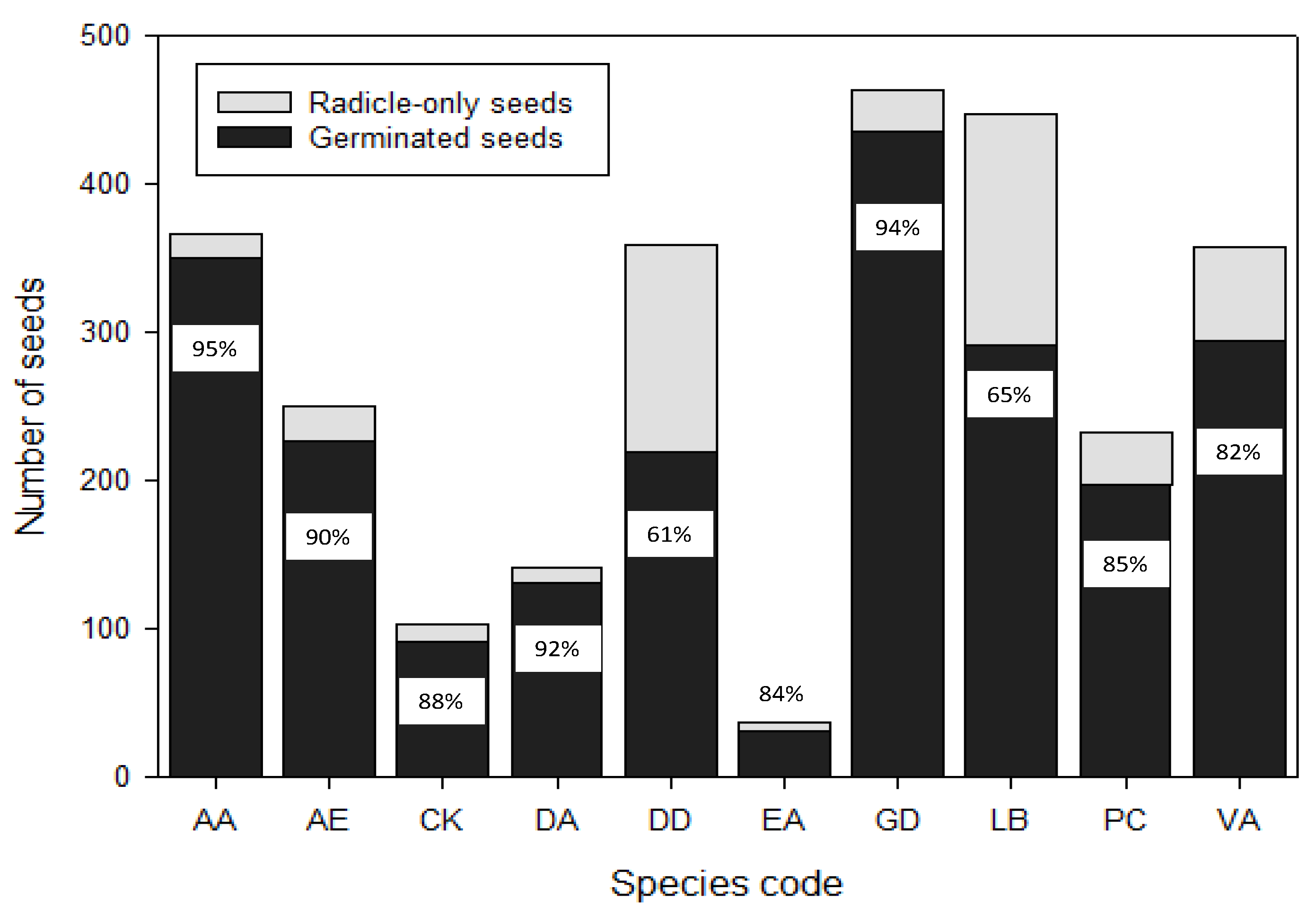
In this assessment, we compared radicle emergence of the seeds with the total final germination, i.e., when the full seedling was established, for the 10 dormant species. We aimed to confirm that all seeds that showed radicle emergence did eventually grow into complete seedlings. For AA, AE, DA and GD, almost 90% of the seeds that produced radicles did grow into complete seedlings (Figure 3). CK, EA, PC and VA had above 80% complete seedling formation, whereas for DD and LB, only around 60% of the seeds with radicle emergence developed into complete seedlings (Figure 3), indicating that radicle emergence alone is not the best predictor for seed vigor.
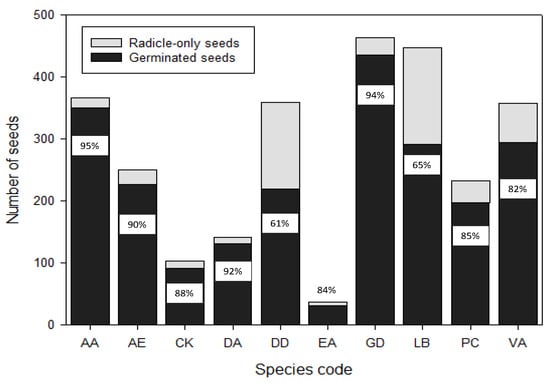
Figure 3.
Total number of seeds with radicle emergence in comparison to number reaching full seedling formation (radicle and shoot with both cotyledonary leaves) for 13 kiwifruit (Actinidia) species. Species codes are spelled out in Table 1.
3.5. RNA Integrity Analysis for Seed Health
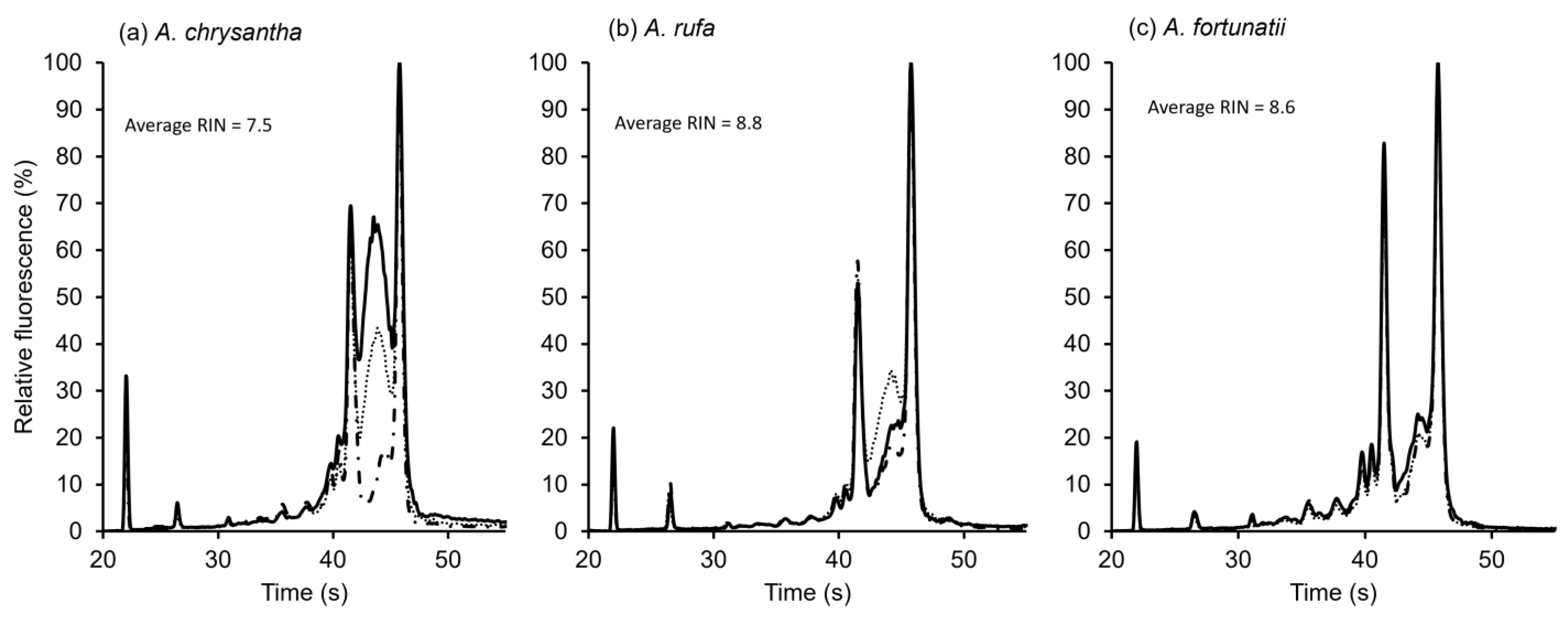
RNA fragmentation assays were completed on three species, providing clear results showing dormant seeds could be distinguished from dead seeds. A. chrysantha seeds were dead upon receipt in the laboratory. The RNA fragmentation analysis showed that the RNA for these seeds had disintegrated, with a relatively low RNA Integrity Number (RIN) of 7.5, as shown in Figure 4a. RE had non-dormant seeds with very high germination immediately after extraction (fresh) and showed high RNA integrity (RIN = 8.8; Figure 4b). GD had dormant seeds which showed over 95% germination following stratification and GA3 treatment and had a high RNA integrity (RIN = 8.6; Figure 4c).
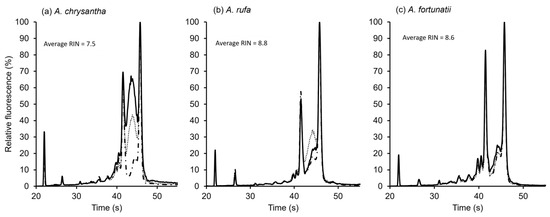
Figure 4.
Comparative RNA fragmentation analysis for (a) Actinidia chrysantha (dead seed); (b) A. rufa (RE) (non-dormant seed); and (c) A. fortunatii (GD) (dormant seed) showing RNA disintegration in dead seeds but not in viable dormant and non-dormant seeds. RIN = RNA Integrity Number.
4. Discussion
To our knowledge, studies on the seed dormancy of kiwifruit seeds are limited and typically focused on Actinidia chinensis var. chinensis and A. chinensis var. deliciosa [9]. In this study, we performed experiments to characterize the dormancy types and to optimize DBTs for seeds of 13 Actinidia species. We also explored the possibility of using RNA integrity analysis to distinguish viable dormant and non-dormant seeds from dead seeds. In addition, we investigated if dormancy traits in kiwifruit seeds could be based on their original species geographic distribution or if there is some adaptation of these traits to their maternal growing environment.
4.1. Environmental Control of Seed Germination
First, we looked at the temperature requirements for germination of the kiwifruit seeds. For this, we tested seed germination under two temperature regimes, i.e., constant (24 °C) and alternating (24/12 °C) temperatures, both with a 16 h photoperiod. Constant temperature was ineffective for the germination of most of the species. Almost all species showed some germination under alternating temperatures, indicating that temperature fluctuation is a critical requirement for the germination of kiwifruit seeds. In general, daily fluctuations in temperature have been shown to facilitate the imbibition of water and thereby improve germination [19]. Fluctuating light and temperature are reported as two critical environmental factors that can promote seed germination [20]. According to Koller [21], the amplitude of daily fluctuating temperatures can be sensed by seeds, and it is generally considered as an indicator of depth of burial: the wider the temperature range occurring near the surface, the stronger the signal to terminate dormancy. Our findings agree with previous studies on kiwifruit seeds indicating that fluctuating temperatures are required to obtain high germination [8,9,14].
4.2. Assessment of Seed Dormancy
Our initial germination test investigated if any of the studied kiwifruit species had non-dormant seeds, i.e., seeds capable of germination without the application of dormancy-breaking treatment. Out of the 13 species tested, three species (LC, RE and SB) had the highest initial germination rates, with their germination not improved following application of DBTs. Notably, RE exhibited remarkably high initial germination, c. 85%. Hence, we concluded that these three species are not dormant. Neither AE nor VA required stratification, but only soaking in water or GA3 for 24 h, for germination. These species could be classified as morphologically dormant. MD is associated with seeds that are dispersed with undeveloped or partly grown embryos that need to grow before they can germinate [22]. Extending the after-ripening period allows the embryo to grow inside the seed prior to germination. Plant hormones, in particular abscisic acid (ABA) and gibberellins (GA), mediate plant developmental processes and regulate whether the seed remains dormant or germinates [23]. ABA is known for inhibiting germination, whereas GA is known to suppress ABA and promote seed germination [24]. While we did not measure the embryo growth of these two (AE and VA) species, the fact that they did not reach their highest potential germination rates during their initial germination (fresh control) test, but showed their highest germination rates following soaking in water or GA3 for 24 h, reconfirms their MD. Seeds with MD can also have PD [7]. In general, if embryo growth and radicle emergence are completed in about 30 days under suitable conditions, the seeds are considered to exhibit only MD [7]. If germination takes more than 30 days, and seeds require additional DBTs such as stratification and GA3 treatment to germinate, they are considered to have MPD [7]. GD provided a classic example of MPD in our study, where its seeds required both stratification and application of GA3 to break dormancy.
The other seven species, AA, CK, DA, DD, EA, PC and LB, needed stratification along with clipping or seed coat removal to reach their highest germination rates, indicating a combination of PY and PD. Some seeds have impermeable seed coats that prevent entry of water and oxygen, causing PY [15]. The presence of a cuticle layer, suberin, lignin or cutin all contribute to hard seed coats [25]. In natural habitats, PY is alleviated by temperature fluctuations due to freezing and melting of soil water, soil microorganism activities, forest fires or animal digestion [26]. Seed PY can be overcome by mechanical scarification, exposure to extreme changes in temperature or by chemical scarification [9,10]. PD is the most common type of dormancy and responds to seasonal cues to ensure that germination occurs only after specific environmental events [27]. The response of PD seeds to their growing environment is highly specific, and germination can be triggered in response to a particular set of temperature, chemical or light signals [6]. All the seven species are confirmed to exhibit combinational dormancy (PY + PD), as seed coat permeability alone was insufficient to break their dormancy. Previous reports [9,10,28,29] on Actinidia chinensis (var. chinensis and A. chinensis var. deliciosa) indicated that this species requires stratification and fluctuating temperatures to break its dormancy, which agrees with our current findings.
4.3. Effect of Species Origin and Geographical Distribution on Their Seed Dormancy
All the Actinidia species studied here originated from China and Taiwan; however, the seeds used in this study were from mature plants growing in New Zealand orchards. As New Zealand experiences a much milder climate than some of the regions from which kiwifruit originate in China, it would be interesting to know if the seed dormancy traits in these species are highly conserved or if there is some phenotypic plasticity that allows rapid adaptation to their maternal environment; in this case, the milder New Zealand climate. We postulated that species from higher altitude and cold climate environments should possess some form of seed dormancy to survive their harsher growing conditions and shorter seasons, and would therefore require a period of cold stratification to break their seed dormancy.
Interestingly, we found that three species in this study (LC, RE and SB) are not dormant, and all three originated from Taiwan [13]. Taiwan has a subtropical humid monsoon climate that experiences mild winter and high summer temperatures, with about a 16 °C difference between the annual minimum and maximum temperatures. While RE and SB are exclusively present in Taiwan, LC is widespread from central to southern China, which experiences a milder climate. AE is another species that is found only in Taiwan and exhibits MD, with only 9% of the seeds being dormant. Originating from a warmer climate, it is not surprising that these four species do not require stratification to germinate.
Both AA and PC species had a high proportion of dormant seeds (70% and 80% respectively) and a combination of PY and PD. While these species are more prominent in the cold temperate climates of northern and northeast China, they are also widely distributed from cold-temperate to tropical-humid monsoon climates and from lower to higher altitudes. AA is found at elevations ranging from 600 to 2600 m. PC belongs to a small group of cold-hardy Actinidia species found in the northern and northwestern regions of China [13]. Both these species require stratification for their germination. Interestingly, GD (with MPD), which also had a high proportion of dormant seeds, is found only in Yunnan, in a region in the southwest of China that experiences a subtropical and tropical highland humid monsoon climate. VA, with about 50% dormant seeds and exhibiting MD, is spread from central to southern China, growing in regions with a comparatively mild climate, and is restricted to elevations between 200 and 600 m. The remaining five species (CK, DA, DD, EA and LB) had 5–36% dormant seeds, all showing combination PY + PD dormancy, and are distributed from central to southern China. While these species originated from a milder climate region, CK, DA and DD are distributed in mountainous regions, from Shaanxi, Hubei, Hunan and Chongqing in the center to Sichuan and Guizhou provinces in the southwest, with altitudes ranging from 200 to 2200 m. Both EA and LB have a similar distribution range, with altitudes ranging from 200 to 2000 m. Their distribution into higher altitudes could be the reason for their requirement for cold-moist stratification for dormancy breaking. While two of the cold-climate-growing species (AA and PC) had higher proportions of dormant seeds, we noted that the proportion of dormant seeds did not correlate with climatic data, as GD and VA, which thrive only in warm climates, also had higher proportions of dormant seeds. It has been reported that the proportion of dormant seeds, depth of dormancy and requirement for dormancy-releasing mechanisms could vary among individuals, populations and genotypes within a species [8,30,31,32] and that this variation is a hereditary component in many species [8,33].
4.4. Putative Impacts of Maternal and Paternal Parents on Seed Dormancy
An important point to note is that the seeds studied here were obtained from open-pollinated flowers in orchards where plants of multiple species were growing. Thus, while we have the information on the seed parents (maternal), the pollen parents (paternal) could not be identified. Different seed dormancy types could be imposed by both the maternal and paternal parents [34]. The seed parent’s contribution can occur through nuclear and cytoplasmic genetic effects; maternal environments; regulation of nutrient resources from endosperm to embryo; synthesis and regulation of ABA and nitrates that either promote seed development and prevent precocious germination, or act as a signal in overcoming seed dormancy, respectively [35,36,37]; and development of the seed coat [34]. For example, the physiological dormancy in the genus Chenopodium was reportedly due to seed coat thickness, which is affected by environmental signals experienced by the mother plant during seed maturation [38,39]. On the other hand, the influence of the pollen parent on dormancy traits could occur through nuclear (genotype) and non-nuclear genetic material [40] and paternal environmental effects [41,42]. Hence, it can be postulated that the pollen parent might have influence in seed morphological dormancy that is related to embryo development, while physical (seed coat related) and physiological (endosperm related) dormancies could be imposed by the seed parent [43]. Therefore, conclusions from this study need some caution as we emphasize a need for careful consideration to distinguish between the effects mediated by the seed parent, by the pollen parent, and those caused directly by the environment [12]. Penfield and MacGregor [12] suggested that these factors can be differentiated through physiological and mechanistic studies coupled with appropriate genetic analysis to demonstrate if the maternal genotype or the immediate environmental effects are the key to seed response.
4.5. Seed Vigor Assessment
The assessment of seed vigor revealed an interesting discrepancy between radicle emergence and completed germination (radicle and shoot with both cotyledonary leaves). The observation that a portion of seeds with radicle emergence did not progress to full germination underscores the importance of considering multiple parameters for accurate vigor assessment. Our study also showed that while the emergence of the radicle from the seed coat is considered the first visible sign of germination, it does not necessarily predict successful germination and subsequent seedling growth. Nonogaki suggests that in most species, the appearance of the embryo, specifically the radicle, marks the start of germination and subsequent seedling growth [44]. In agriculture, seedling emergence from the soil is often referred to as “germination” to signify successful seedling establishment, and is defined by Bewley and Black as the emergence of both the shoot and the root system [45].
4.6. RNA Integrity Analysis
This study also revealed the potential of RNA integrity analysis for distinguishing between dormant and dead seeds, offering a rapid and precise alternative to conventional viability staining assessments. This assay was developed at the USDA [11] for studying dormancy in tomato and brassica seeds and provides a quick method for determining dormancy, as results are available within 5 min of RNA sample preparation. When non-dormant, dormant and dead seeds are assayed for germinability, dormant and dead seeds may show the same response (no germination). A cut test can show an ungerminated seed was probably alive but does not provide quantitative data. However, our study showed that when non-dormant, dormant and dead seeds are assayed for RNA integrity, non-dormant and dormant seeds show the same response (high RNA integrity), while dead seeds (low RNA integrity) and dormant seeds can be distinguished by quantifiable differences in RNA integrity. This RNA fragmentation assay provides a fast and precise method to distinguish dead and dormant seed lots—this could save time and money compared with the conventional viability staining assessment. RNA extraction and RNA quantification are easily scaled up for high-throughput applications and have great potential for application in breeding and propagation.
5. Conclusions
In conclusion, this study advances our understanding of kiwifruit seed dormancy and germination. The findings underscore the need for tailored dormancy-breaking treatments, emphasize the importance of regional considerations in breeding efforts and introduce a novel approach to seed health assessment through RNA integrity analysis. These insights contribute significantly to the field of kiwifruit seed research and have broader implications for optimizing seed propagation in other horticultural crops.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/seeds3020014/s1, Table S1: Comparative Mean Germination Time (MGT) and Germination Rate Index (GRI) calculated for 13 Actinidia species following various dormancy-breaking treatments.
Author Contributions
Conceptualization and methodology, J.N., A.E., M.F., L.M. and E.M.; data collection, J.N., A.E., L.M., C.N., R.K. and M.F.; data analysis, A.M. and M.F.; revision and discussion, J.N., A.E., C.N., R.K., A.M., L.M., M.F. and E.M.; manuscript written by J.N., A.E., C.N., R.K., A.M., L.M., M.F. and E.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the New Zealand Institute for Plant and Food Research Limited (PFR) Kiwifruit Royalty Investment Programme (KRIP) for Germplasm Programme.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Acknowledgments
The authors express their gratitude to Andrew Mullan and Belinda Diepenheim for providing laboratory support; a Catalyst Seeding grant (Contract number 34877) from the Royal Society Te Apārangi for supporting the RNA fragmentation study in collaboration with the National Center for Genetic Resources Preservation (NCGRP), USDA, Fort Collins; Keith Funnel and Annette Richardson for providing an internal review of the manuscript; and the Plant & Food Research Science Publishing Office for editorial support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barboni, T.; Cannac, M.; Chiaramonti, N. Effect of cold storage and ozone treatment on physicochemical parameters, soluble sugars and organic acids in Actinidia deliciosa. Food Chem. 2010, 121, 946–951. [Google Scholar] [CrossRef]
- Poudel, K.; Shah, M.; Mandal, J. Fruit Quality Analysis of Kiwifruit Cultivars Cultivated in Eastern Mid-Hills in Nepal. J. Agric. Environ. 2019, 20, 217–225. [Google Scholar] [CrossRef]
- Rey, M.; Ferradas, Y.; Martinez, O.; Gonzalez, M.V. Actinidia spp. Kiwifruit; CABI Digital Library: Wallingford, UK, 2020; pp. 1–18. [Google Scholar]
- Zespri-Annual-Report. Available online: https://www.zespri.com/content/dam/zespri/nz/publications/annual-reports/Zespri-Annual-Report-2021-22.pdf (accessed on 5 June 2023).
- MPI. Situation and Outlook for Primary Industry New Zealand; Ministry for Primary Industries: Wellington, New Zealand, 2023.
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Elsevier: Amsterdam, The Netherlands, 1998. [Google Scholar]
- Lawes, G.; Anderson, D. Influence of temperature and gibberellic acid on kiwifruit (Actinidia chinensis) seed germination. N. Z. J. Exp. Agric. 1980, 8, 277–280. [Google Scholar] [CrossRef]
- Windauer, L.B.; Insausti, P.; Biganzoli, F.; Benech-Arnold, R.; Izaguirre, M.M. Dormancy and germination responses of kiwifruit (Actinidia deliciosa) seeds to environmental cues. Seed Sci. Res. 2016, 26, 342–350. [Google Scholar] [CrossRef]
- Fleming, M.B.; Richards, C.M.; Walters, C. Decline in RNA integrity of dry-stored soybean seeds correlates with loss of germination potential. J. Exp. Bot. 2017, 68, 2219–2230. [Google Scholar] [CrossRef]
- Penfield, S.; MacGregor, D.R. Effects of environmental variation during seed production on seed dormancy and germination. J. Exp. Bot. 2016, 68, 819–825. [Google Scholar] [CrossRef]
- Huang, H. Kiwifruit: The Genus Actinidia; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Ferguson, A.R. The genus Actinidia. In Kiwifruit Science and Management, 1st ed.; Warrington, I.J., Weston, G.C., Eds.; New Zealand Society for Horticultural Science: Auckland, New Zealand, 1990; pp. 15–35. [Google Scholar]
- Li, J.-Q.; Li, X.-W.; Soejarto, D.D. Actinidiaceae. In Flora of China; Wu, Z.-Y., Raven, P.H., Hong, D.-Y., Eds.; Science Press: Beijing, China; Missouri Botanical Gardens: St. Louis, MO, USA, 2007; Volume 12, pp. 334–360. [Google Scholar]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Wallisellen, Switzerland, 2023. [Google Scholar]
- Kader, M.A. A comparison of seed germination calculation formulae and the associated interpretation of resulting data. J. Proc. R. Soc. N. S. W. 2005, 138, 65–75. [Google Scholar] [CrossRef]
- Schroeder, A.; Mueller, O.; Stocker, S.; Salowsky, R.; Leiber, M.; Gassmann, M.; Lightfoot, S.; Menzel, W.; Granzow, M.; Ragg, T. The RIN: An RNA integrity number for assigning integrity values to RNA measurements. BMC Mol. Biol. 2006, 7, 3. [Google Scholar] [CrossRef]
- Geneve, R.L. Impact of temperature on seed dormancy. HortScience 2003, 38, 336–340. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Sánchez, R.A.; Forcella, F.; Kruk, B.C.; Ghersa, C.M. Environmental control of dormancy in weed seed banks in soil. Field Crops Res. 2000, 67, 105–122. [Google Scholar] [CrossRef]
- Koller, D. Environmental control of seed germination. In Seed Biology; Kozlowski, T.T.E., Ed.; Academic Press: New York, NY, USA, 1972; pp. 2–101. [Google Scholar]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue, K.; Rubio de Casas, R.; The NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytol. 2014, 203, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.D.; Ocampo-Serna, D.M.; Martínez-Rubio, R.A.; Correa-Navarro, Y.M. Determination of gibberellic acid and abscisic acid in (Zea mays L.) (ICA-V305) seeds germinated using dynamic sonication assisted solvent extraction and maceration. MethodsX 2022, 9, 101821. [Google Scholar] [CrossRef] [PubMed]
- Kermode, A.R. Role of Abscisic Acid in Seed Dormancy. J. Plant Growth Regul. 2005, 24, 319–344. [Google Scholar] [CrossRef]
- Rolston, M.P. Water Impermeable Seed Dormancy. Bot. Rev. 1978, 44, 365–396. [Google Scholar] [CrossRef]
- Mayer, W.V. “Dormancy”. Encyclopedia Britannica. 18 August 2022. Available online: https://www.britannica.com/science/dormancy (accessed on 1 November 2023).
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014; p. 1600. [Google Scholar]
- Kim, I.; Hwang, J.; Han, K.; Lee, K. Studies on the germination of seeds in native Actinidia species. J. Korean Soc. Hortic. Sci. 1987, 28, 335–342. [Google Scholar]
- Smith, R.; Toy, S. Effects of stratification and alternating temperatures on seed germination of the Chinese gooseberry, Actinidia chinensis Planch. Proc. Am. Soc. Hortic. Sci. 1967, 90, 409–412. [Google Scholar]
- Donohue, K. Seeds and seasons: Interpreting germination timing in the field. Seed Sci. Res. 2005, 15, 175–187. [Google Scholar] [CrossRef]
- Sautu, A.; Baskin, J.M.; Baskin, C.C.; Condit, R. Studies on the seed biology of 100 native species of trees in a seasonal moist tropical forest, Panama, Central America. For. Ecol. Manag. 2006, 234, 245–263. [Google Scholar] [CrossRef]
- Wagmann, K.; Hautekèete, N.-C.; Piquot, Y.; Meunier, C.; Schmitt, S.E.; Van Dijk, H. Seed dormancy distribution: Explanatory ecological factors. Ann. Bot. 2012, 110, 1205–1219. [Google Scholar] [CrossRef]
- Meyer, S.E.; Allen, P.S. Ecological Genetics of Seed Germination Regulation in Bromus tectorum L. II. Reaction Norms in Response to a Water Stress Gradient Imposed during Seed Maturation. Oecologia 1999, 120, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Baskin, C.C. How much influence does the paternal parent have on seed germination? Seed Sci. Res. 2019, 29, 1–11. [Google Scholar] [CrossRef]
- Lee, K.P.; Piskurewicz, U.; Turecková, V.; Strnad, M.; Lopez-Molina, L. A seed coat bedding assay shows that RGL2-dependent release of abscisic acid by the endosperm controls embryo growth in Arabidopsis dormant seeds. Proc. Natl. Acad. Sci. USA 2010, 107, 19108–19113. [Google Scholar] [CrossRef]
- Kang, J.; Yim, S.; Choi, H.; Kim, A.; Lee, K.P.; Lopez-Molina, L.; Martinoia, E.; Lee, Y. Abscisic acid transporters cooperate to control seed germination. Nat. Commun. 2015, 6, 8113. [Google Scholar] [CrossRef] [PubMed]
- Penfield, S.; Li, Y.; Gilday, A.D.; Graham, S.; Graham, I.A. Arabidopsis ABA INSENSITIVE4 regulates lipid mobilization in the embryo and reveals repression of seed germination by the endosperm. Plant Cell 2006, 18, 1887–1899. [Google Scholar] [CrossRef]
- Karssen, C.M. The light promoted germination of the seeds of Chenopodium album L. III. Effect of the photoperiod during growth and development of the plants on the dormancy of the produced seeds. Plant Biol. 1970, 19, 81–94. [Google Scholar] [CrossRef]
- Pourrat, Y.; Jacques, R. The influence of photoperiodic conditions received by the mother plant on morphological and physiological characteristics of Chenopodium polyspermum L. seeds. Plant Sci. Lett. 1975, 4, 273–279. [Google Scholar] [CrossRef]
- Hansen, A.K.; Escobar, L.K.; Gilbert, L.E.; Jansen, R.K. Paternal, maternal, and biparental inheritance of the chloroplast genome in Passiflora (Passifloraceae): Implications for phylogenetic studies. Am. J. Bot. 2007, 94, 42–46. [Google Scholar] [CrossRef]
- Schmid, B.; Dolt, C. Effects of Maternal and Paternal Environment and Genotype on Offspring Phenotype in Solidago altissima L. Evolution 1994, 48, 1525–1549. [Google Scholar] [CrossRef]
- Herman, J.; Sultan, S. Adaptive Transgenerational Plasticity in Plants: Case Studies, Mechanisms, and Implications for Natural Populations. Front. Plant Sci. 2011, 2, 102. [Google Scholar] [CrossRef] [PubMed]
- De Giorgi, J.; Piskurewicz, U.; Loubery, S.; Utz-Pugin, A.; Bailly, C.; Mène-Saffrané, L.; Lopez-Molina, L. An Endosperm-Associated Cuticle Is Required for Arabidopsis Seed Viability, Dormancy and Early Control of Germination. PLoS Genet. 2015, 11, e1005708. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination—Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M. Seeds: Physiology of Development and Germination; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).