
Genetic Variability for Mesocotyl Length in Maize †



Abstract
:1. Introduction
2. Experiments
2.1. Genetic Materials
2.2. Experimental Setup
2.3. Data Collection
- Mesocotyl length
- Seed biochemical attributes
2.4. Data Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Niu, L.; Hao, R.; Wu, X.; Wang, W. Maize mesocotyl: Role in response to stress and deep-sowing tolerance. Plant Breed. 2020, 139, 1–8. [Google Scholar] [CrossRef]
- Zhao, G.W.; Wang, J.H. Effect of auxin on mesocotyl elongation of dark grown maize under different seeding depths. Russ. J. Plant Physiol. 2010, 57, 79–86. [Google Scholar] [CrossRef]
- Pan, B.R.; Zhong, T.L.; Zhao, G.W. Promoting deep-sowing germinability of corn (Zea mays) by seed soaking with gibberellic acid. Arch. Agron. Soil Sci. 2017, 63, 1314–1323. [Google Scholar] [CrossRef]
- Zhao, G.W.; Wang, J.H. Effect of gibberellin and uniconazole on mesocotyl elongation of dark-grown maize under different seeding depths. Plant Prod. Sci. 2008, 11, 423–429. [Google Scholar] [CrossRef]
- Kutschera, U.; Wang, Z.-Y. Growth-limiting proteins in maize coleoptiles and the auxin-brassinosteroid hypothesis of mesocotyl elongation. Protoplasma 2016, 253, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Sivasubramanian, S.; Menon, M. Heterosis and inbreeding depression in rice. Madras Agric. J. 1973, 60, 1139. [Google Scholar]
- Johnson, H.W.; Robinson, H.; Comstock, R.F. Estimates of Genetic and Environmental Variability in Soybean. Agron. J. 1955, 47, 314–318. [Google Scholar] [CrossRef]
- Olayiwola, M.O.; Soremi, P.A.S. Variability for dry fodder yield and component traits in cowpea [Vigna unguiculata (L.) Walp]. Electron. J. Plant Breed. 2014, 5, 58–62. [Google Scholar]
- Ogunniyan, D.J.; Olakojo, S.A. Genetic variation, heritability, genetic advance and agronomic character association of yellow elite inbred lines of maize (Zea mays L.). Niger. J. Genet. 2015, 28, 24–28. [Google Scholar] [CrossRef] [Green Version]
- Mostafavi, K.; Shoahosseini, M.; Geive, G.S. Multivariate analysis of variation among traits of corn hybrids traits under drought stress. Int. J. Agric. Sci. 2011, 1, 416–422. [Google Scholar]
- Oyetunde, O.A.; Badu-Apraku, B.; Ariyo, O.J.; Alake, C.O. Efficiencies of Heterotic Grouping Methods for Classifying Early Maturing Maize Inbred Lines. Agronomy 2020, 10, 1198. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Gedil, M.; Annor, B.; Talabi, A.O.; Akaogu, I.C.; Oyekunle, M.; Akinwale, R.O.; Fasanmade, T.Y. Heterotic patterns of IITA and CIMMYT Early maturing Yellow Maize Inbreds under Contrasting Environments. Agron. J. 2016, 108, 1321–1336. [Google Scholar] [CrossRef] [Green Version]
- Muhammad, K. Nutrient Seed Priming Improves Abiotic Stress Tolerance in Zea mays L. and Glycine max L. Ph.D. Thesis, University of Hohenheim, Stuttgart, Germany, 2012. [Google Scholar]
- Martínez-Ballesta, M.C.; Egea-Gilabert, C.; Conesa, E.; Ochoa, J.; Vicente, M.J.; Franco, J.A.; Bañon, S.; Martínez, J.J.; Fernández, J.A. The Importance of Ion Homeostasis and Nutrient Status in Seed Development and Germination. Agronomy 2020, 10, 504. [Google Scholar] [CrossRef] [Green Version]


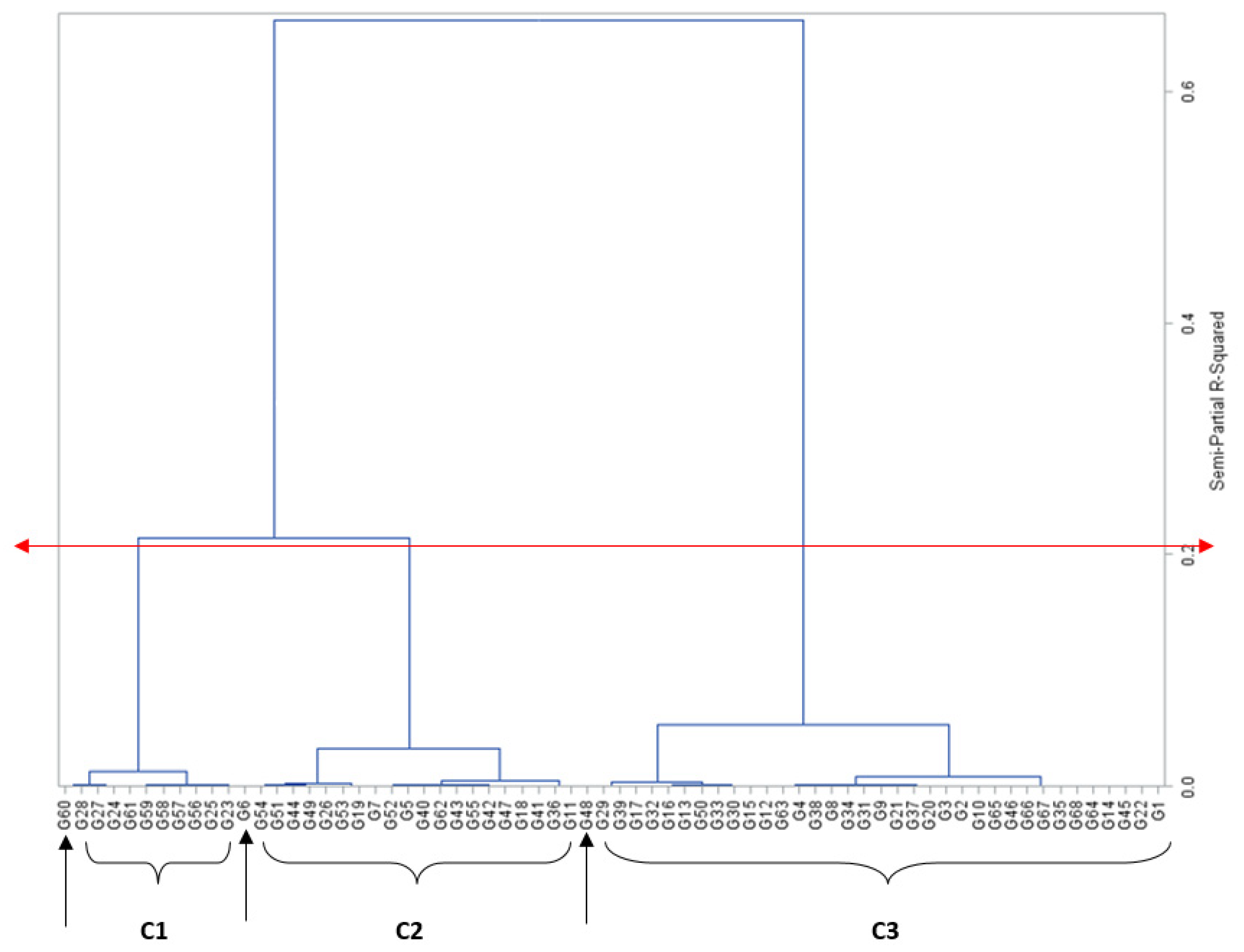{kind=link}
| Source of Variation | DF | Mean Square |
|---|---|---|
| Genotype | 67 | 13.19 ** |
| Error | 68 | 4.40 |
| R2 | 0.75 | |
| Genotypic variance | 8.79 | |
| Environmental variance | 4.40 | |
| Phenotypic variance | 13.19 | |
| Broad-sense heritability | 0.67 | |
| Genotypic coefficient of variation | 0.57 | |
| Phenotypic coefficient of variation | 0.70 | |
| Genetic advance (% of Mean) | 0.96 | |
| ID | Genotype | Mean | Std | Cluster | ID | Genotype | Mean | Std | Cluster |
|---|---|---|---|---|---|---|---|---|---|
| G25 | A1804-14 | 0 | 0 | 1 | G2 | PVAEH 30 | 5.99 | 1.57 | 3 |
| G56 | LY1001-18 | 0 | 0 | 1 | G37 | LY1901-11 | 6.08 | 3.11 | 3 |
| G57 | LY1409-14 | 0 | 0 | 1 | G20 | Oba super 2 | 6.12 | 1.27 | 3 |
| G58 | LY1312-11 | 0 | 0 | 1 | G9 | †PVAEH 41 | 6.2 | 0.83 | 3 |
| G59 | LY1901-15 | 0 | 0 | 1 | G21 | LY1312-23 | 6.2 | 0.99 | 3 |
| G61 | A1804-67 | 0 | 0 | 1 | G8 | †PVAEH 43 | 6.28 | 1.22 | 3 |
| G23 | A1312-12 | 0.58 | 0.19 | 1 | G34 | LY1901-23 | 6.3 | 2.12 | 3 |
| G24 | LY1501-7 | 1.18 | 0.22 | 1 | G31 | LY1501-8 | 6.31 | 0.86 | 3 |
| G27 | LY1901-14 | 1.36 | 0.55 | 1 | G38 | †PVAEH 40 | 6.45 | 2.81 | 3 |
| G28 | LY1913-3 | 1.74 | 1.02 | 1 | G4 | PVAEH 28 | 6.47 | 1.54 | 3 |
| G60 | A1804-66 * | 2.02 | 2.86 | - | G63 | A1736-6 | 6.57 | 1.51 | 3 |
| G19 | †PVAEH 36 | 2.45 | 3.46 | 2 | G66 | LY1001-23 | 6.77 | 2.73 | 3 |
| G53 | Local check | 2.65 | 3.75 | 2 | G67 | Mkt-cultivar A | 6.88 | 1.27 | 3 |
| G51 | M1124-31 | 3.06 | 3.62 | 2 | G35 | LY1901-20 | 6.93 | 3.15 | 3 |
| G54 | LY1501-6 | 3.23 | 1.1 | 2 | G68 | Mkt-cultivar B | 6.98 | 0.76 | 3 |
| G6 | PVAQEH-4 * | 3.28 | 1.64 | - | G14 | PVAQEH-6 | 7.02 | 2.88 | 3 |
| G44 | LY1901-12 | 3.57 | 3.01 | 2 | G64 | LY1901-24 | 7.02 | 0.68 | 3 |
| G49 | A1802-4 | 3.67 | 0.61 | 2 | G45 | A1706-2 | 7.04 | 2.88 | 3 |
| G26 | A1802-12 | 3.7 | 0.95 | 2 | G22 | LY1901-22 | 7.05 | 3.1 | 3 |
| G36 | LY1901-17 | 4.1 | 0.11 | 2 | G1 | PVAEH 26 | 7.06 | 2.43 | 3 |
| G41 | LY1409-21 | 4.11 | 2.42 | 2 | G65 | LY1501-9 | 7.19 | 0.24 | 3 |
| G11 | PVAQEH-3 | 4.4 | 0.62 | 2 | G46 | Ife hybrid-4 | 7.29 | 4.26 | 3 |
| G18 | †PVAEH 37 | 4.65 | 6.58 | 2 | G48 | LY1914-14* | 7.67 | 3.95 | - |
| G47 | LY1901-19 | 4.78 | 4.1 | 2 | G29 | LY1501-5 | 7.68 | 0.99 | 3 |
| G62 | LY1501-1 | 4.89 | 1.99 | 2 | G39 | †PVAEH 33 | 7.87 | 0.52 | 3 |
| G43 | LY1901-25 | 4.94 | 1.92 | 2 | G17 | †PVAEH 44 | 7.89 | 0.78 | 3 |
| G55 | LY1901-21 | 5.05 | 1.53 | 2 | G33 | A1736-13 | 8.18 | 0.85 | 3 |
| G42 | LY1901-16 | 5.11 | 1.2 | 2 | G50 | A1804-15 | 8.22 | 2.8 | 3 |
| G40 | Ife hybrid-3 | 5.24 | 0.62 | 2 | G30 | LY1913-16 | 8.38 | 2.38 | 3 |
| G5 | Check (RE) | 5.39 | 0.69 | 2 | G15 | PVAEH 27 | 8.42 | 0.28 | 3 |
| G52 | LY1901-13 | 5.39 | 0.87 | 2 | G12 | Local check | 8.52 | 0.51 | 3 |
| G7 | PVAEH 29 | 5.56 | 0.81 | 2 | G13 | †PVAEH 31 | 8.73 | 1.6 | 3 |
| G3 | †PVAEH 32 | 5.95 | 2.5 | 3 | G16 | †PVAEH 34 | 8.94 | 0.03 | 3 |
| G10 | PVAQEH-5 | 5.98 | 0.72 | 3 | G32 | LY1901-18 | 9.02 | 0.42 | 3 |
| Biochemical Content | ML |
|---|---|
| Iron | 0.31 * |
| Zinc | 0.29 * |
| Selenium | 0.01 ns |
| Crude protein | 0.33 ** |
| Free fatty acid | 0.26 * |
| Oil | 0.30 * |
| Linoleic acid | 0.28 * |
| Amylase | −0.31 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oyetunde, O.A.; Godonu, K.G.; Adeboye, K.A. Genetic Variability for Mesocotyl Length in Maize. Biol. Life Sci. Forum 2021, 4, 26. https://doi.org/10.3390/IECPS2020-08781
Oyetunde OA, Godonu KG, Adeboye KA. Genetic Variability for Mesocotyl Length in Maize. Biology and Life Sciences Forum. 2021; 4(1):26. https://doi.org/10.3390/IECPS2020-08781
Chicago/Turabian StyleOyetunde, Oyeboade Adebiyi, Kolawole Gbemavo Godonu, and Kehinde Adewole Adeboye. 2021. "Genetic Variability for Mesocotyl Length in Maize" Biology and Life Sciences Forum 4, no. 1: 26. https://doi.org/10.3390/IECPS2020-08781
APA StyleOyetunde, O. A., Godonu, K. G., & Adeboye, K. A. (2021). Genetic Variability for Mesocotyl Length in Maize. Biology and Life Sciences Forum, 4(1), 26. https://doi.org/10.3390/IECPS2020-08781