Definition
Fungi produce a wide range of secondary metabolites. Some of these metabolites are toxic to humans and cause various health disorders, ranging from acute poisoning to chronic diseases. Contrary to this, some fungal metabolites are valuable sources in therapeutics, such as penicillin. Herein, researchers briefly highlight the role played by different fungal metabolites in human health and diseases and give an overview of the most common fungal genera.
Keywords:
fungi; secondary metabolites; mycotoxin; Aspergillus; Penicillium; Fusarium; antibiotic; anticancer 1. Introduction
Fungi are extraordinary in their ability to produce numerous natural products known as fungal secondary metabolites, which exhibit various biological activities [1,2]. Considering the reported biological properties of fungi toward human health, they can be classified as toxic “mycotoxins” or non-toxic metabolites [3]. Mycotoxins are natural contaminants of food commodities and pose a measurable health risk for animal and human health [4]. Health risks are classified based on organs that are affected by the toxins, for instance, they possess carcinogenic, immunosuppressive, hepatotoxic, nephrotoxic, and neurotoxic effects [4,5]. Hundreds of mycotoxins have been identified [6,7]. The mycotoxins that receive the most significant concern from scientists are aflatoxin B1 and M1, cyclopiazonic acid, ochratoxin A, patulin, T-2 toxin, deoxynivalenol, zearalenone, ergot alkaloids, and macrocyclic trichothecenes, due to their health and economic effects. For instance, aflatoxin B1 is classified as the most potent hepatocarcinogen and mutagen [8,9]. However, human exposure to mycotoxins is still underestimated as scientists only focus on economically important mycotoxins. Therefore, it is important to intensify efforts to discover more toxic fungal metabolites that contribute to diseases of unknown aetiology.
On the contrary, some fungal metabolites are beneficial for human health and are used to treat many human diseases, such as antibiotics. Therefore, researchers from different fields work together towards understanding the fungal secondary metabolites in terms of their regulation, function, and applications, and evaluate their toxicity [10,11,12,13]. These fungal metabolites are characterized by their chemical structures and effects on human health [1]. A recent review mentioned that more than 1,500 fungal metabolites had been isolated and half of them have been used for primary health care. The other half is still under safety and toxicity investigation [1].
The scientific community has recently celebrated the 90th anniversary of Sir Alexander Fleming’s discovery of penicillin, which was the starting point of the antibiotic era [10]. Since then, scientists worldwide have started to consider and use innovative approaches to study fungi due to their health and biotechnological significance. Although drug discovery from fungal metabolites is increased due to the development of bioinformatics and genomics [14], researchers expressed the need to ramp up the discovery of fungal metabolites to improve food quality or discover novel antibiotics. This was obvious during the pandemic when researchers initially struggled to find drugs to treat the SARS-CoV-2.
In the coming section, researchers will briefly discuss the importance of different fungal metabolites in human health and diseases with an overview of the fungal genera most frequently involved.
2. Mycotoxin Involved in Human Diseases
Mycotoxins are produced by hundreds of thousands of species of fungi, such as Aspergillus spp., Penicillium spp., Fusarium spp., Claviceps spp., and Stachybotrys spp. [15,16]. Therefore, mycologists classified mycotoxins based on the fungal species that produce them, see Table 1. These toxins can cause several health issues, such as liver and kidney cancers [6].

Table 1.
Common toxigenic species of fungi and their chronic effects.
2.1. Mycotoxin Involved in Liver Diseases
Aflatoxins are difuranocumarin derivatives produced by many species of Aspergilli; particularly A. flavus, A. parasiticus, A. bombycis, A. coracles, A. nomius and A. pseudotamarii [6,15,37]. These fungi grow on particular foods under favourable temperature and humidity conditions and produce aflatoxins before and after harvesting [17,38]. Aflatoxins are primarily found in cereals, oilseeds, tree nuts, spices, milk, and dairy products [39,40,41]. A. flavus is the most common producer of aflatoxins. It was first isolated and characterized following the death of more than 100,000 turkeys due to an unidentified disease, named turkey X disease. The toxin was traced to a mould-contaminated peanut meal consumed by the turkeys [6]. Eighteen types of aflatoxins have been identified, with the major four being AFB1, AFB2, AFG1 and AFG2 (Figure 1A, 1–4, respectively). Due to high toxicity, aflatoxin B1 was classified as a Group 1A human carcinogen [8,42]. Aflatoxin B1 (AFB1) is the most potent natural toxin and may cause liver cancer [16]. The Food and Agriculture Organization emphasized that children under 5 years in Africa are at a high risk of consuming milk contaminated with aflatoxin M1 and M2 which cause growth retardation [17]. AFM1 and AFM2 are metabolism products of AFB1, and thus can be found in milk. Therefore, several regulations have been implemented worldwide to control aflatoxin consumption [15,16].
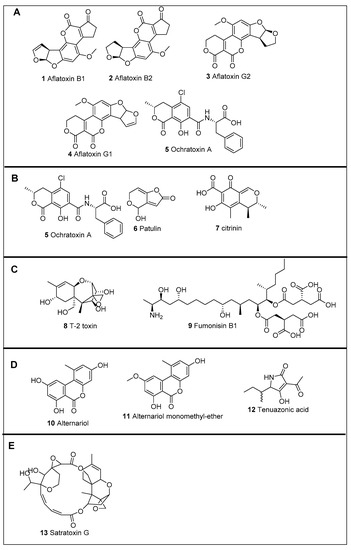
Figure 1.
Common mycotoxins—(A). Aspergillus spp. (B). Penicillium spp. (C). Fusarium spp. (D). Alternaria spp. (E). Stachybotrys spp.
2.2. Mycotoxins Involved in Kidney Diseases
Members of the ochratoxin family have been found in varied species of Aspergillus, in particular A. alliaceus, A. melleus, A. cabonarius, A. glaucus, and A. niger, and other related fungal species, including Penicillium viridicatum [6,43]. These toxins are found in various cereal grains, dried fruits, wine, and coffee. Ochratoxin A (OTA) (Figure 1A, 5) is the most toxic compound in the ochratoxin family [19]. Early studies have connected OTA exposure within human diseases, such as Balkan endemic nephropathy (BEN), chronic interstitial nephropathy (CIN), and other renal diseases. The effect of OTA was reported by IARC and is considered nephrotoxic, teratogenic, immunotoxic, and has been classified by IARC as a class 2B carcinogen, and a possible human carcinogen [20,44]. However, a recent European Food Safety Authority report showed that ochratoxins cause nephrotoxicity and renal tumours in various animal species. The information related to the effect of this toxin on human health is still limited, and more clinical studies are needed to confirm the hazardous extent of ochratoxins [18].
2.3. Mycotoxins Involved in Genotoxicity, Immunotoxicity, and Neurotoxicity
Patulin (PAT) (Figure 1B, 6) is a secondary metabolite produced by Aspergillus, Byssochlamys, and Penicillium spp. [25,45] and is common infestation of fruit- and vegetable-based products. The risk of human exposure to patulin through the consumption of juices and jams occurs at an early age [24].
Patulin is an α, β-unsaturated γ-lactone that has broad-spectrum antimicrobial activity but has been classified as mycotoxin [25]. Patulin has chronic health effects, including genotoxicity, immunotoxicity, and neurotoxicity in rodents, but insufficient evidence exists for carcinogenicity. Therefore, the IARC has classified it as Group 3B, however, its effects on humans are not yet clear [24,25].
2.4. Mycotoxins Involved in Damage to Organs
Citrinin (Figure 1B, 7) is mycotoxin that was isolated from Penicillium cultures that contaminated stored food for long periods of time [21]. Initially, citrine aimed to be used as a drug before World War II; unfortunately, it also displayed toxicity, so it was discontinued [22]. Several Penicillium strains were found to produce citrinin such as P. citrinum, P. expansum, P. radicicola, P. verrucosum [21,22]. Chemically citrinin is a quinone methide with two intramolecular hydrogen bonds [23].
Moreover, numerous Aspergillus species were also able to produce it. Citrinin has also been isolated from Monascus ruber and Monascus purpureus, which are used industrially to produce pigments for the food industry as a preservative or for food colouring. Although citrinin mainly targets the kidney, reports indicated it targets other organs, such as the liver and bone marrow. However, regulating this toxin in food or animal feed would meet an acceptable level in many countries including Europe [21,22].
2.5. Mycotoxins Involved Gastrointestinal, Skin, Thyroid, and Bone Marrow Disorders
Trichothecenes are a large group of toxic fungal secondary metabolites produced by various fungi, including Fusarium spp., Myrothecium spp., Stachybotrys spp., and Trichoderma spp. [46,47]. These toxins cause many health issues for humans and animals [26,47]. Over 200 trichothecene toxins were reported. They share a common tricyclic 12,13-epoxytrichothec-9-ene (EPT) core structure and are classified into four groups A, B, C, and D based on the substitution pattern of EPT, with groups A and D recognised as the most toxic compounds [27,47].
T-2 toxin (Figure 1C, 8) is related to trichothecenes group A and is considered the most toxic fungal secondary metabolite produced by Fusarium species, including F. sporotrichioides, F. poae, and F. acuminatum. T-2 toxicity effects depend on several factors, such as the dosage, age, and exposure (oral, dermal, and aerosol). T-2 toxin causes a gastrointestinal disorder known as Alimentary Toxic Aleukia that affects humans after consuming moldy food. Several toxicological studies were conducted to evaluate the cytotoxic and genotoxic impact of T-2 toxin [26,27,46]. They found that this toxin could inhibit protein synthesis and has immunomodulatory insults [26,27,46]. This inhibitory effect is visible in proliferating cells, such as the gastrointestinal tract, skin, thyroid, and bone marrow cells [26,31].
2.6. Mycotoxins Involved in Cancer and Heart Failure
Fumonisins are mycotoxins produced by Fusarium spp. such as F. proliferatum and F. verticillioides. Fumonisins are classified into four major groups; FA, FB, FC, and FP, of which fumonisin B1 (Figure 1C, 9) is the most toxic member. FB1 contaminates corn, rice, oat, rye, barley and wheat [48]. However, the risk of contamination with these toxic fungal metabolites depends on several environmental factors, such as humidity and temperature [49].
FB1 (9) poses a health risk for animals and humans. It was reported that FB1 causes oxidative stress, cellular autophagy, apoptosis, and kidney diseases. Exposure to high levels of fumonisins in diet during early pregnancy induces a high risk of bearing a child with a brain or spinal cord deficiency. Therefore, worldwide regulations and legislation have restricted a maximum limit of fumonisin in human foods and animal feeds [28,49].
2.7. Mycotoxins Involved in Oesophageal Cancer and Hematologic Disorder
Alternaria spp. is a widespread fungal genus that infects various crops, causing economic loss [50]. Alternaria toxins are a large group of more than seventy toxins, however, only a few have been reported and chemically characterised, including alternariol (AOH), alternariol monomethyl-ether (AME), and tenuazonic acid (TeA) (Figure 1D, 10–12, respectively) [33,51,52]. These toxins contaminate food products, such as fruits, vegetables, cereals, and grains. According to EFSA, the information on the toxicological profile of Alternaria mycotoxins is not enough for a proper health hazard assessment [50,53]. Many studies described these toxins as having cytotoxic, genotoxic, mutagenic, fetotoxic and teratogenic exchange consequences among their effects [54]. Other studies reported these toxins to cause mutagenic, oestrogenic and clastogenic effects [50,55]. However, most of the published studies focus on a single toxin, although Alternaria toxins, in general, are produced simultaneously. Although one study investigated the effect of binary or ternary combinations in vitro on relevant human cell lines [34], more data is still needed to understand the impact of these toxins on human health.
2.8. Mycotoxins Involved in Cause Respiratory Illness
Satratoxins are macrocyclic trichothecene mycotoxins produced by Stachybotrys species. Satratoxin G (Figure 1E, 13) is one of the most known toxins in this family [56]. Stachybotrys chartarum is associated with dampness and is called saprophytic black mould [36,57]. Many researchers suggested that satratoxins contribute to sick building syndrome with different symptoms, such as upper respiratory tract symptoms including wheezing and coughing, and in some cases they may exacerbate chronic lung diseases, such as asthma [35].
3. Fungal Metabolites against Human Disease
3.1. Fungal Metabolites Used as Antibiotics
An antibiotic is a chemical substance generally produced by microorganisms and able to inhibit the growth or destroy different microbial pathogens. Literature indicated that most filamentous fungi could produce different antibiotic compounds against pathogenic bacteria, fungi, etc. [58]. Most currently used antibiotics are derived from Streptomyces species and other actinobacteria, with relatively few compound classes derived from fungi. This includes Penicillium-derived penicillins (Figure 2, 14) and Cephalosporium-derived cephalosporins (Figure 2, 15), which belong to the β-glucan antibiotics, as well as the steroid fusidic acid (Figure 2, 16) from Fusidium coccineum and pleuromutilin (Figure 2, 17) [59,60,61].
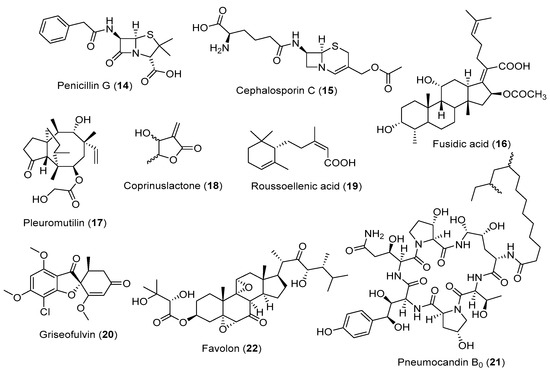
Figure 2.
Selected fungal metabolites with antibiotic effect.
The discovery of the penicillin- and cephalosporin-type β-lactams has shaped the modern pharmaceutical industry, and they have remained the most important antibiotics for around 70 years after their discovery. Afterwards, thousands of fungal metabolites were discovered; however, only a few have entered clinical development, which could be attributed to the exit of most of the Big Pharma companies from the field of novel antibiotics discovery [62].
Although penicillin G was the first fungal natural product to develop the first antibiotic, mycophenolic acid was discovered even earlier in 1893, from a Penicillium brevicompactum strain identified to inhibit the growth of Bacillus anthracis. It was claimed to be the first purified natural product antibiotic [63]. Due to its toxicity, it was recently dismissed as an antibiotic but developed as the immunosuppressant drug mycophenolate mofetil.
Recently, the antibacterial antibiotics pipeline showed that most compounds under development had been optimised from old classes with known modes of action [64]. Hence, antibiotic resistance is likely to happen once launched. The inhibition of biofilm formation is one of the most promising strategies to discover antibiotics that combat resistant pathogens. Two recent examples are coprinuslactone (Figure 2, 18) derived from the mushroom Coprinus comatus against Pseudomonas aeruginosa biofilm [11], and roussoellenic acid (Figure 2, 19) from a Roussoella spp., against Staphylococcus aureus biofilm [12].
While resistant bacterial pathogens are a pressing topic, relatively low attention is paid to the resistant pathogenic fungi. There are only a few antimycotic compound classes on the market, including the oldest griseofulvin (Figure 2, 20), which was discovered from Penicillium griseofulvum in 1952 [13]. The newest class of fungi-derived antifungal agents are the echinocandin lipopeptides, such as pneumocandin B0 (Figure 2, 21) derived from Glarea lozoyensis [65]. Another example is favolon (Figure 2, 22), isolated from Favolaschia calocera and exhibiting a potent antifungal effect [66]. Strobilurin is also considered an antifungal agent. Strobilurin A and B are extracted from Strobilurus tenacellus and used in agriculture as fungicides [67].
3.2. Fungal Metabolites Used as Anticancer Agents
Although most anticancer agents were discovered from plants and bacteria, fungi are also considered as an essential source for anticancer drug discovery. Illudin S (Figure 3, 23) is a fungal toxin isolated from Omphalotus illudens (Jack O’Lantern mushroom) and showed promising anticancer effects against different cancer cell lines [68]. Its semi-synthetic derivative irofulven (Figure 3, 24) interferes with DNA replication complexes and cell division in DNA synthesis. It passed phase II clinical trials with promising effects against brain, breast, blood, colon, prostate, lungs, ovarian, and pancreatic cancer cell lines [69]. Aphidicolin (Figure 3, 25) is a tetracyclic diterpene isolated from Cephalosporium aphidicola (currently changed to Akanthomyces muscarius) with antimitotic properties that acts via the specific binding on DNA polymerases and is still under intensive clinical trials [70].
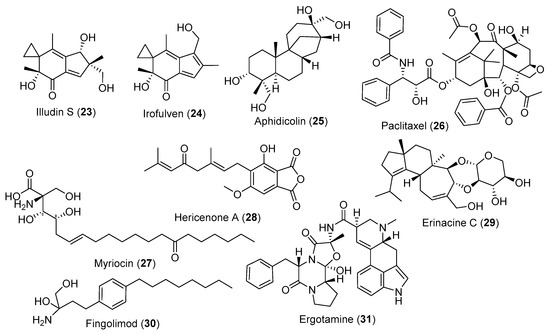
Figure 3.
Selected fungal metabolites with anticancer and CNS-related effects.
In 1971, a novel diterpene was isolated from the bark of the northwest Pacific yew tree, Taxus brevifolia and named taxol or paclitaxel (Figure 3, 26). It demonstrated moderate in vivo activity against different cancer cell lines. After more than 20 years of its initial report, it was FDA-approved against breast and ovarian cancers with a unique mechanism of action. However, the supply to the market was a real problem due to its very low yield. With many different trials to solve this issue, Stierle et al. [71] focused on discovering a new production source; endophytes that colonize the yew tree. They isolated 50 ng/L of the drug for the first time from Taxomyces andreanae [70]. The continuous search for paclitaxel led to the discovery of over 20 endophytic fungal genera that could produce either paclitaxel or its analogues [72].
3.3. Fungal Metabolites Used as CNS-Disease-Related Agents
Hericium erinaceus is a medical mushroom that produces two terpenoid classes, hericenones, such as hericenone A (Figure 3, 27), and erinacines, such as erinacine C (Figure 3, 28), which can stimulate the synthesis of the nerve growth factor and nerve regeneration in vitro and in vivo [73]. During the synthesis process to produce myriocin (Figure 3, 29) derivatives, a compound produced by the insect-associated ascomycete Isaria sinclairii, fingolimod (Figure 3, 30) was one of the final products. Fingolimod is a potent immunosuppressant that was FDA-approved in 2010 as a new treatment for multiple sclerosis [74]. Ergotamine (Figure 3, 31) is the principal alkaloid produced by the ergot fungus Claviceps purpurea. It has a similar structure to a few neurotransmitters and exhibits a vasoconstrictor effect. It is used to prevent post-partum haemorrhage (bleeding after childbirth) and is currently used for treating acute migraine attacks [75].
3.4. Fungal Metabolites Control Cardiovascular Diseases
Cardiovascular diseases include the malfunctioning of the heart and blood vessels due to elevated plasma cholesterol levels. Cardiovascular diseases are the leading cause of death worldwide, with more than eighteen million deaths yearly. Inhibition of de novo synthesis of cholesterol through 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase inhibitors is an effective method to reduce plasma cholesterol and hence cardiovascular diseases. The fungal metabolites statins are the most important class of HMG-CoA reductase inhibitors. Compactin or mevastatin (Figure 4, 32) was isolated from Penicillium brevicompactum and exhibited promising hypocholesterolemic activity, and since then has been known under the name “ML-236B” [76,77]. Later, the first FDA-approved statin as an HMG-CoA reductase inhibitor was lovastatin (Figure 4, 33). Although different fungal species produce it, the commercial product is derived from Aspergillus terreus [78].
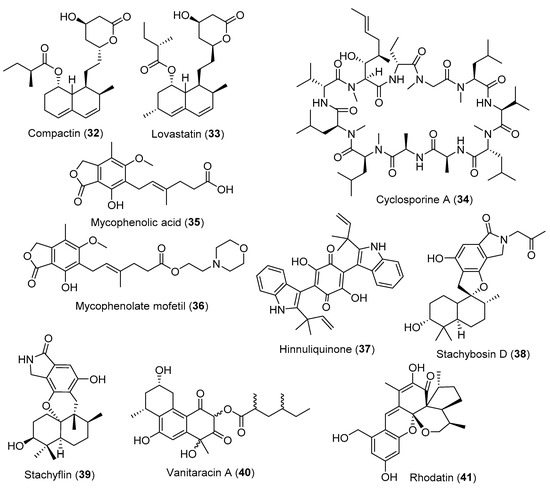
Figure 4.
Selected fungal metabolites with antiviral, immunomodulatory and cardiovascular-related effects.
3.5. Fungal Metabolites Used as Immunomodulatory Agents
In the global market, some of the crucial immunomodulatory drugs are biotechnologically produced by bacterial or fungal fermentation. Cyclosporine A (Figure 4, 34) was first isolated from the ascomycete Tolypocladium inflatum and exhibited a mild antifungal effect. After many years, it was discovered that this cyclopeptide possesses a precise mode of action by selectively binding to cyclophilin A. This protein inhibits calcineurin, which in turn inhibits interleukine II production cascade, and hence the immune response of the human body is suppressed [63]. This is usually used in crucial immunosuppressant therapy to prevent the rejection of organ transplants.
Mycophenolic acid (Figure 4, 35) was the first discovered meroterpenoid antibiotic and produced by Penicillium spp., including P. brevicompactum and P. roquefortii. However, it has never been marketed as an antibiotic [79]. Its use as an immunosuppressant became evident, as it selectively inhibits inosine monophosphate dehydrogenase (IMPDH), an essential enzyme for the T- and B-lymphocytes, and thereby suppresses the immune system [80]. It can also prevent peroxynitrite tissue damage through the depletion of tetrahydrobiopterin, a co-factor to induce nitric oxide synthase (iNOS). Consequently, its prodrug mycophenolate mofetil (Figure 4, 36) is used to prevent organ rejection and manage psoriasis and other immunological disorders.
3.6. Fungal Metabolites Used as Antiviral Agents
Viral diseases usually cause severe global epidemics/pandemics, especially when vaccines or antiviral drugs are not available. So, a continuous search is urgently needed for new antiviral agents from natural sources. A lot of potentially active fungal natural products have shown antiviral activity; however, none of them have reached the market. The bis-indolyl quinone hinnuliquinone (Figure 4, 37) isolated from an unknown fungus recovered from Quercus coccifera, inhibited the wild-type and clinically-resistant HIV-1 protease, a key enzyme in the replication and maturation of the HIV-1 virus [81]. Stachybosin D (Figure 4, 38), a phenylspirodrimane metabolite isolated from a sponge-derived fungus Stachybotrys chartarum, was able to inhibit HIV-1 replication in both NNRTIs-resistant strain and wild-type HIV-1 by targeting reverse transcriptase [82].
The terpenoid stachyflin (Figure 4, 39), isolated from a marine-derived isolate of Stachybotrys spp. showed good activity against the influenza A virus (H1N1) [83]. The novel tricyclic polyketide vanitaracin A (Figure 4, 40) isolated from a culture broth of Talaromyces spp., was reported to inhibit all hepatitis B genotypes (A–D) viral entry processes through direct interaction with the HBV entry receptor correlated to the hepatitis D virus [84]. Recently, the novel meroterpenoid rhodatin (Figure 4, 41) with its unprecedented carbon skeleton, was isolated from cultures of the rare basidiomycete Rhodotus palmatus and exhibited significant anti-HCV activities [85].
4. Summary
This review has emphasised the importance of fungal metabolites in human health and disease. The fungal metabolites are divided into mycotoxin and non-toxic metabolites. While mycotoxin contaminations are almost non-avoidable and thus constitute a serious risk for food safety, other non-toxic metabolites may even become beneficial for human health. This is further underlined by the recent pandemic that dragged attention to the need for the discovery of new metabolites that can be used as treatments in the future.
Author Contributions
All authors were involved in writing—original draft preparation, review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Naranjo-Ortiz, M.; Gabaldón, T. Fungal evolution: Cellular, genomic and metabolic complexity. Biol. Rev. 2020, 95, 1198–1232. [Google Scholar] [CrossRef] [PubMed]
- Boruta, T. Uncovering the repertoire of fungal secondary metabolites: From Fleming’s laboratory to the International Space Station. Bioengineered 2017, 9, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Eshelli, M.; Qader, M.M.; Jambi, E.J.; Hursthouse, A.S.; Rateb, M.E. Current Status and Future Opportunities of Omics Tools in Mycotoxin Research. Toxins 2018, 10, 433. [Google Scholar] [CrossRef]
- Peraica, M.; Radić, B.; Lucić, A.; Pavlović, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. [Google Scholar]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- Liew, W.P.P.; Mohd-Redzwan, S. Mycotoxin: Its impact on gut health and microbiota. Front. Cell. Infect. Microbiol. 2018, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Stela, M.; Saluk-Bijak, J.; Siadkowski, A.; Bijak, M. Molecular Aspects of Mycotoxins—A Serious Problem for Human Health. Int. J. Mol. Sci. 2020, 21, 8187. [Google Scholar] [CrossRef] [PubMed]
- Magnussen, A.; A Parsi, M. Aflatoxins, hepatocellular carcinoma and public health. World J. Gastroenterol. 2013, 19, 1508–1512. [Google Scholar] [CrossRef] [PubMed]
- Tola, M.; Kebede, B. Occurrence, importance and control of mycotoxins: A review. Cogent Food Agric. 2016, 2, 1191103. [Google Scholar] [CrossRef]
- Karwehl, S.; Stadler, M. Exploitation of fungal biodiversity for discovery of novel antibiotics. Curr. Top. Microbiol. Immunol. 2016, 398, 303–338. [Google Scholar]
- De Carvalho, A.; do Amaral, M.W. Coprinuslactone Protects the Edible Mushroom Coprinus Comatus against Biofilm Infections by Blocking Both Quorum-sensing and MurA. Wiley Online Libr. 2016, 18, 4254–4264. [Google Scholar]
- Phukhamsakda, C.; Macabeo, A.P.G.; Yuyama, K.T.; Hyde, K.D.; Stadler, M. Biofilm Inhibitory Abscisic Acid Derivatives from the Plant-Associated Dothideomycete Fungus, Roussoella sp. Molecules 2018, 23, 2190. [Google Scholar] [CrossRef]
- Grove, J.F.; MacMillan, J.; Mulholland, T.P.C.; Rogers, M.A.T. 762. Griseofulvin. Part IV. Structure. J. Chem. Soc. 1952, 3977–3987. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A Global Concern for Food Safety, Human Health and Their Management. Front. Microbiol. 2017, 7, 2170. [Google Scholar] [CrossRef] [PubMed]
- Hamid, A.S.; Tesfamariam, I.G.; Zhang, Y.; Zhang, Z.G. Aflatoxin B1-induced hepatocellular carcinoma in developing countries: Geographical distribution, mechanism of action and prevention. Oncol. Lett. 2013, 5, 1087–1092. [Google Scholar] [CrossRef]
- Walte, H.-G.; Schwake-Anduschus, C.; Geisen, R.; Fritsche, J. Aflatoxin: Food chain transfer from feed to milk. J. Für Verbrauch. Und Lebensm. 2016, 11, 295–297. [Google Scholar] [CrossRef]
- Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.C.; Nebbia, C.S. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, e06113. [Google Scholar] [CrossRef]
- EL Khoury, A.; Atoui, A. Ochratoxin A: General Overview and Actual Molecular Status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef]
- Bui-Klimke, T.R.; Wu, F. Ochratoxin A and Human Health Risk: A Review of the Evidence. Crit. Rev. Food Sci. Nutr. 2013, 55, 1860–1869. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain (Contam). Scientific Opinion on the risks for public and animal health related to the presence of citrinin in food and feed. EFSA J. 2012, 10, 2605. [Google Scholar] [CrossRef]
- Silva, L.J.G.; Pereira, A.M.P.T.; Pena, A.; Lino, C.M. Citrinin in Foods and Supplements: A Review of Occurrence and Analytical Methodologies. Foods 2020, 10, 14. [Google Scholar] [CrossRef] [PubMed]
- Atapattu, S.N.; Poole, C.F. Recent advances in analytical methods for the determination of citrinin in food matrices. J. Chromatogr. A 2020, 1627, 461399. [Google Scholar] [CrossRef]
- Barreira, M.J.; Alvito, P.; Almeida, C.M.M. Occurrence of patulin in apple-based-foods in Portugal. Food Chem. 2010, 121, 653–658. [Google Scholar] [CrossRef]
- Mahato, D.K.; Kamle, M.; Sharma, B.; Pandhi, S.; Devi, S.; Dhawan, K.; Selvakumar, R.; Mishra, D.; Kumar, A.; Arora, S.; et al. Patulin in food: A mycotoxin concern for human health and its management strategies. Toxicon 2021, 198, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Janik, E.; Niemcewicz, M.; Podogrocki, M.; Ceremuga, M.; Stela, M.; Bijak, M. T-2 Toxin—The Most Toxic Trichothecene Mycotoxin: Metabolism, Toxicity, and Decontamination Strategies. Molecules 2021, 26, 6868. [Google Scholar] [CrossRef]
- Ueno, Y. Toxicological features of T-2 toxin and related trichothecenes. Fundam. Appl. Toxicol. 1984, 4, S124–S132. [Google Scholar] [CrossRef]
- Voss, K.A.; Riley, R.T.; Norred, W.P.; Bacon, C.W.; Meredith, F.I.; Howard, P.C.; Plattner, R.D.; Collins, T.F.; Hansen, D.K.; Porter, J.K. An overview of rodent toxicities: Liver and kidney effects of fumonisins and Fusarium moniliforme. Environ. Health Perspect. 2001, 109, 259–266. [Google Scholar] [CrossRef]
- Chen, J.; Wen, J.; Tang, Y.; Shi, J.; Mu, G.; Yan, R.; Cai, J.; Long, M. Research Progress on Fumonisin B1 Contamination and Toxicity: A Review. Molecules 2021, 26, 5238. [Google Scholar] [CrossRef]
- Wu, Q.-H.; Wang, X.; Yang, W.; Nüssler, A.K.; Xiong, L.-Y.; Kuča, K.; Dohnal, V.; Zhang, X.-J.; Yuan, Z.-H. Oxidative stress-mediated cytotoxicity and metabolism of T-2 toxin and deoxynivalenol in animals and humans: An update. Arch. Toxicol. 2014, 88, 1309–1326. [Google Scholar] [CrossRef]
- Peles, F.; Sipos, P.; Győri, Z.; Pfliegler, W.P.; Giacometti, F.; Serraino, A.; Pagliuca, G.; Gazzotti, T.; Pócsi, I. Adverse Effects, Transformation and Channeling of Aflatoxins Into Food Raw Materials in Livestock. Front. Microbiol. 2019, 10, 2861. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and its metabolites: Effect on human health, metabolism and neutralisation methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef] [PubMed]
- A Meronuck, R.; A Steele, J.; Mirocha, C.J.; Christensen, C.M. Tenuazonic acid, a toxic produced by Alternaria alternata. Appl. Microbiol. 1972, 23, 613–617. [Google Scholar] [CrossRef]
- Hollander, D.D.; Holvoet, C.; Demeyere, K.; De Zutter, N.; Audenaert, K.; Meyer, E.; Croubels, S. Cytotoxic Effects of Alternariol, Alternariol Monomethyl-Ether, and Tenuazonic Acid and Their Relevant Combined Mixtures on Human Enterocytes and Hepatocytes. Front. Microbiol. 2022, 13, 849243. [Google Scholar] [CrossRef]
- Ulrich, S.; Lang, K.; Niessen, L.; Baschien, C.; Kosicki, R.; Twarużek, M.; Straubinger, R.K.; Ebel, F. The Evolution of the Satratoxin and Atranone Gene Clusters of Stachybotrys chartarum. J. Fungi 2022, 8, 340. [Google Scholar] [CrossRef]
- Dyląg, M.; Spychała, K.; Zielinski, J.; Łagowski, D.; Gnat, S. Update on Stachybotrys chartarum—Black Mold Perceived as Toxigenic and Potentially Pathogenic to Humans. Biology 2022, 11, 352. [Google Scholar] [CrossRef]
- Coppock, R.W.; Christian, R.G.; Jacobsen, B.J. Aflatoxins. In Veterinary Toxicology, 3rd ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 983–994. [Google Scholar] [CrossRef]
- Baranyi, N.; Kocsubé, S.; Varga, J. Aflatoxins: Climate change and biodegradation. Curr. Opin. Food Sci. 2015, 5, 60–66. [Google Scholar] [CrossRef]
- Eshelli, M.; Harvey, L.; Edrada-Ebel, R.; McNeil, B. Metabolomics of the Bio-Degradation Process of Aflatoxin B1 by Actinomycetes at an Initial pH of 6.0. Toxins 2015, 7, 439–456. [Google Scholar] [CrossRef]
- Milićević, D.R.; Spirić, D.; Radičević, T.; Velebit, B.; Stefanović, S.; Milojević, L.; Janković, S. A review of the current situation of aflatoxin M1 in cow’s milk in Serbia: Risk assessment and regulatory aspects. Food Addit. Contam. Part A 2017, 34, 1617–1631. [Google Scholar] [CrossRef]
- Filazi, A.; Tansel, U. Occurrence of Aflatoxins in Food. In Aflatoxins—Recent Advances and Future Prospects; Intechopen: London, UK, 2013. [Google Scholar] [CrossRef]
- Kimanya, M.E.; Routledge, M.N.; Mpolya, E.; Ezekiel, C.N.; Shirima, C.P.; Gong, Y.Y. Estimating the risk of aflatoxin-induced liver cancer in Tanzania based on biomarker data. PLoS ONE 2021, 16, e0247281. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 Years of Research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vettorazzi, A.; van Delft, J.; de Cerain, A.L. A review on ochratoxin A transcriptomic studies. Food Chem. Toxicol. 2013, 59, 766–783. [Google Scholar] [CrossRef] [PubMed]
- Nan, M.; Xue, H.; Bi, Y. Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables. Toxins 2022, 14, 309. [Google Scholar] [CrossRef] [PubMed]
- Sudakin, D.L. Trichothecenes in the environment: Relevance to human health. Toxicol. Lett. 2003, 143, 97–107. [Google Scholar] [CrossRef]
- Foroud, N.A.; Baines, D.; Gagkaeva, T.Y.; Thakor, N.; Badea, A.; Steiner, B.; Bürstmayr, M.; Bürstmayr, H. Trichothecenes in Cereal Grains—An Update. Toxins 2019, 11, 634. [Google Scholar] [CrossRef]
- Proctor, R.; Brown, D.; Busman, M.; Naumann, T. Genomic and Metabolomic Approaches for Detection and Control of Fusarium, Fumonisins and Other Mycotoxins on Corn. Available online: https://www.ars.usda.gov/research/project/?accnNo=430343 (accessed on 20 June 2022).
- Dall’Asta, C.; Battilani, P. Fumonisins and their modified forms, a matter of concern in future scenario? World Mycotoxin J. 2016, 9, 727–739. [Google Scholar] [CrossRef]
- Alexander, J.; Benford, D.; Boobis, A.; Ceccatelli, S.; Cottrill, B.; Cravedi, J.-P.; Di Domenico, A.; Doerge, D.; Dogliotti, E.; Edler, L.; et al. Scientific Opinion on the risks for animal and public health related to the presence ofAlternariatoxins in feed and food. EFSA J. 2011, 9, 2407. [Google Scholar] [CrossRef]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria Mycotoxins in Food and Feed: An Overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef]
- Rodríguez-Carrasco, Y.; Mañes, J.; Berrada, H.; Juan, C. Development and Validation of a LC-ESI-MS/MS Method for the Determination of Alternaria Toxins Alternariol, Alternariol Methyl-Ether and Tentoxin in Tomato and Tomato-Based Products. Toxins 2016, 8, 328. [Google Scholar] [CrossRef]
- De Souza, G.D.; Mithöfer, A.; Daolio, C.; Schneider, B.; Rodrigues-Filho, E. Identification of Alternaria alternata Mycotoxins by LC-SPE-NMR and Their Cytotoxic Effects to Soybean (Glycine max) Cell Suspension Culture. Molecules 2013, 18, 2528–2538. [Google Scholar] [CrossRef]
- Meena, M.; Samal, S. Alternaria host-specific (HSTs) toxins: An overview of chemical characterization, target sites, regulation and their toxic effects. Toxicol. Rep. 2019, 6, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Hövelmann, Y.; Hickert, S.; Cramer, B.; Humpf, H.-U. Determination of Exposure to the Alternaria Mycotoxin Tenuazonic Acid and Its Isomer allo-Tenuazonic Acid in a German Population by Stable Isotope Dilution HPLC-MS3. J. Agric. Food Chem. 2016, 64, 6641–6647. [Google Scholar] [CrossRef] [PubMed]
- Islam, Z.; Harkema, J.R.; Pestka, J.J. Satratoxin G from the Black Mold Stachybotrys chartarum Evokes Olfactory Sensory Neuron Loss and Inflammation in the Murine Noseand Brain. Environ. Health Perspect. 2006, 114, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Carey, S.A.; Plopper, C.G.; Hyde, D.M.; Islam, Z.; Pestka, J.J.; Harkema, J.R. Satratoxin-G from the Black Mold Stachybotrys chartarum Induces Rhinitis and Apoptosis of Olfactory Sensory Neurons in the Nasal Airways of Rhesus Monkeys. Toxicol. Pathol. 2012, 40, 887–898. [Google Scholar] [CrossRef]
- Bills, G.F.; Gloer, J.B. Biologically Active Secondary Metabolites from the Fungi. Microbiol. Spectr. 2016, 4, 1087–1119. [Google Scholar] [CrossRef]
- Godtfredsen, W.O.; Jahnsen, S.; Lorck, H.; Roholt, K.; Tybring, L. Fusidic Acid: A New Antibiotic. Nature 1962, 193, 987. [Google Scholar] [CrossRef]
- Newton, G.G.F.; Abraham, E.P. Cephalosporin C, a New Antibiotic containing Sulphur and D-α-Aminoadipic Acid. Nature 1955, 175, 548. [Google Scholar] [CrossRef]
- Novak, R.; Shlaes, D.M. The Pleuromutilin Antibiotics: A New Class for Human Use. Curr. Opin. Investig. Drugs 2010, 11, 182–191. [Google Scholar]
- Jayawardena, R.S.; Hyde, K.D.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Perera, R.H.; de Silva, N.I.; Maharachchikumburua, S.S.N.; Samarakoon, M.C.; Ekanayake, A.H.; et al. One stop shop III: Taxonomic update with molecular phylogeny for important phytopathogenic genera: 51–75 (2019). Fungal Divers. 2019, 98, 77–160. [Google Scholar] [CrossRef]
- Bentley, R. Mycophenolic Acid: A One Hundred Year Odyssey from Antibiotic to Immunosuppressant. Chem. Rev. 2000, 100, 3801–3826. [Google Scholar] [CrossRef]
- Hesterkamp, T. Antibiotics Clinical Development and Pipeline. Curr. Top. Microbiol. 2016, 398, 447–474. [Google Scholar] [CrossRef]
- Denning, D.W. Echinocandins: A new class of antifungal. J. Antimicrob. Chemother. 2002, 49, 889–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anke, H.; Stadler, M.; Mayer, A.; Sterner, O. Secondary metabolites with nematicidal and antimicrobial activity from nematophagous fungi and Ascomycetes. Can. J. Bot. 1995, 73, 932–939. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, Y.; Zhan, H.; Bhatt, P.; Chen, S. An Overview of Strobilurin Fungicide Degradation:Current Status and Future Perspective. Front. Microbiol. 2020, 11, 389. [Google Scholar] [CrossRef]
- Nakanishi, K.; Ohashi, M.; Tada, M.; Yamada, Y. Illudin s (lampterol). Tetrahedron 1965, 21, 1231–1246. [Google Scholar] [CrossRef]
- Movassaghi, M.; Piizzi, G.; Siegel, D.S.; Piersanti, G. Enantioselective Total Synthesis of (−)-Acylfulvene and (−)-Irofulven. Angew. Chem. 2006, 118, 5991–5995. [Google Scholar] [CrossRef]
- Bucknall, R.A.; Moores, H.; Simms, R.; Hesp, B. Antiviral Effects of Aphidicolin, a New Antibiotic Produced by Cephalosporium aphidicola. Antimicrob. Agents Chemother. 1973, 4, 294–298. [Google Scholar] [CrossRef]
- Stierle, A.; Strobel, G.; Stierle, D.; Grothaus, P.; Bignami, G. The Search for a Taxol-Producing Microorganism Among the Endophytic Fungi of the Pacific Yew, Taxus brevifolia. J. Nat. Prod. 1995, 58, 1315–1324. [Google Scholar] [CrossRef]
- Stierle, A.A.; Stierle, D.B. Bioactive Secondary Metabolites Produced by the Fungal Endophytes of Conifers. Nat. Prod. Commun. 2015, 10, 1671–1682. [Google Scholar] [CrossRef]
- Thongbai, B.; Rapior, S.; Hyde, K.D.; Wittstein, K.; Stadler, M. Hericium erinaceus, an amazing medicinal mushroom. Mycol. Prog. 2015, 14, 91. [Google Scholar] [CrossRef]
- Strader, C.R.; Pearce, C.J.; Oberlies, N.H. Fingolimod (FTY720): A Recently Approved Multiple Sclerosis Drug Based on a Fungal Secondary Metabolite. J. Nat. Prod. 2011, 74, 900–907. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Panaccione, D.G.; Tudzynski, P. Chapter 2 Ergot Alkaloids—Biology and Molecular Biology. Alkaloids Chem. Biol. 2006, 63, 45–86. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Faust, J.R.; Goldstein, J.L.; Kaneko, I.; Endo, A. Induction of 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase Activity in Human Fibroblasts Incubated with Compactin (ML-236B), a competitive inhibitor of the reductase. J. Biol. Chem. 1978, 253, 1121–1128. [Google Scholar] [CrossRef]
- Endo, A.; Kuroda, M.; Tsujita, Y. ML-236A, ML-236B, and ML-236C, New Inhibitors of Cholesterogenesis Produced by Penicillium Citrinium. J. Antibiot. 1976, 29, 1346–1348. [Google Scholar] [CrossRef] [PubMed]
- Jahromi, M.F.; Liang, J.B.; Ho, Y.W.; Mohamad, R.; Goh, Y.M.; Shokryazdan, P. Lovastatin Production byAspergillus terreusUsing Agro-Biomass as Substrate in Solid State Fermentation. J. Biomed. Biotechnol. 2012, 2012, 196264. [Google Scholar] [CrossRef]
- Wiesinger, D.; Borel, J. Studies on the Mechanism of Action of Cyclosporin A. Immunobiology 1980, 156, 454–463. [Google Scholar] [CrossRef]
- Allison, A.C.; Eugui, E.M. Mycophenolate mofetil and its mechanisms of action. Immunopharmacology 2000, 47, 85–118. [Google Scholar] [CrossRef]
- Singh, S.B.; Ondeyka, J.G.; Tsipouras, N.; Ruby, C.; Sardana, V.; Schulman, M.; Sanchez, M.; Pelaez, F.; Stahlhut, M.W.; Munshi, S.; et al. Hinnuliquinone, a C2-symmetric dimeric non-peptide fungal metabolite inhibitor of HIV-1 protease. Biochem. Biophys. Res. Commun. 2004, 324, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Li, L.; Zhu, T.; Ba, M.; Li, G.; Gu, Q.; Guo, Y.; Li, D. Phenylspirodrimanes with Anti-HIV Activity from the Sponge-Derived Fungus Stachybotrys chartarum MXH-X73. J. Nat. Prod. 2013, 76, 2298–2306. [Google Scholar] [CrossRef]
- Minagawa, K.; Kouzuki, S.; Yoshimoto, J.; Kawamura, Y.; Tani, H.; Iwata, T.; Terui, Y.; Nakai, H.; Yagi, S.; Hattori, N.; et al. Stachyflin and Acetylstachyflin, Novel Anti-influenza A Virus Substances, Produced by Stachybotrys sp. RF-7260. I. Isolation, Structure Elucidation and Biological Activities. J. Antibiot. 2002, 55, 155–164. [Google Scholar] [CrossRef]
- Kaneko, M.; Watashi, K.; Kamisuki, S.; Matsunaga, H.; Iwamoto, M.; Kawai, F.; Ohashi, H.; Tsukuda, S.; Shimura, S.; Suzuki, R.; et al. A Novel Tricyclic Polyketide, Vanitaracin A, Specifically Inhibits the Entry of Hepatitis B and D Viruses by Targeting Sodium Taurocholate Cotransporting Polypeptide. J. Virol. 2015, 89, 11945–11953. [Google Scholar] [CrossRef] [PubMed]
- Sandargo, B.; Michehl, M.; Praditya, D.; Steinmann, E.; Stadler, M.; Surup, F. Antiviral Meroterpenoid Rhodatin and Sesquiterpenoids Rhodocoranes A-E from the Wrinkled Peach Mushroom, Rhodotus Palmatus. Org. Lett. 2019, 21, 3286–3289. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).