
Phosphate Turnover in Various Parts of Nodulated Soybean (Glycine max (L.) Merr.) Plants and the Relation to the Xylem Transport



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Cultivation
2.2. P Treatments
2.3. Analysis of Pi Concentration and Pi Content
2.4. Statistics
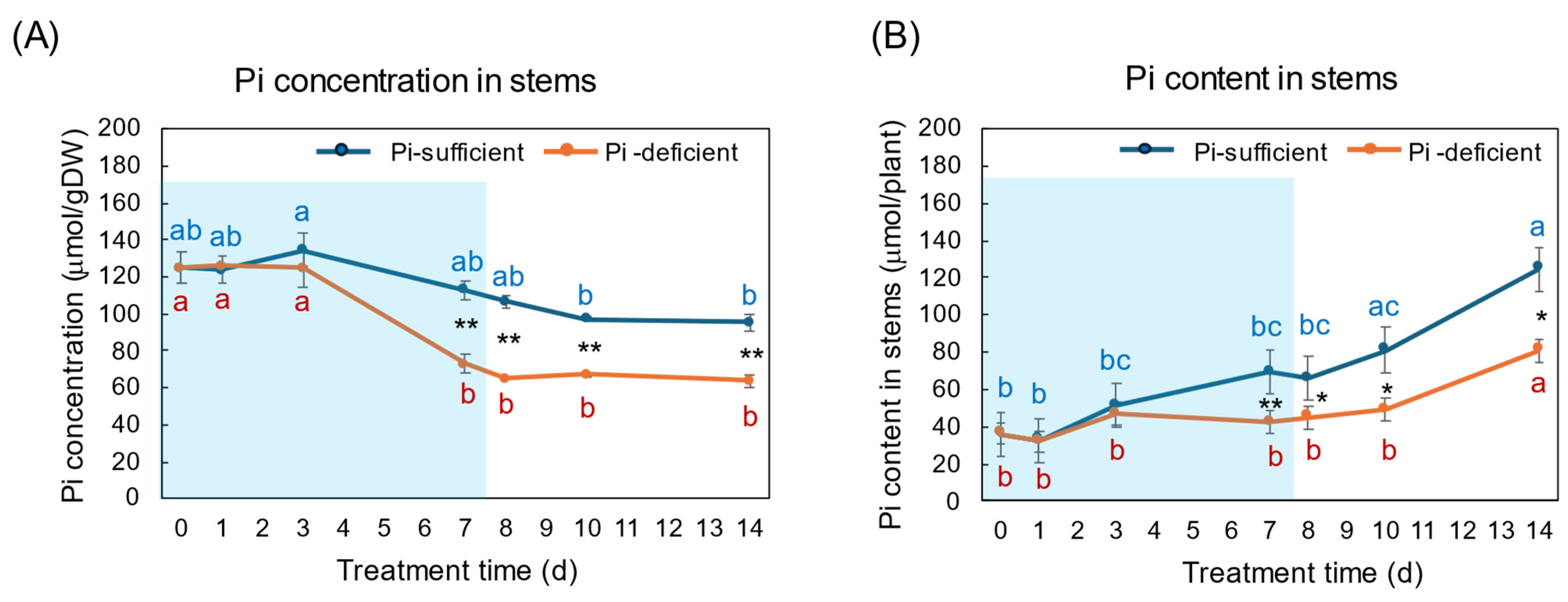
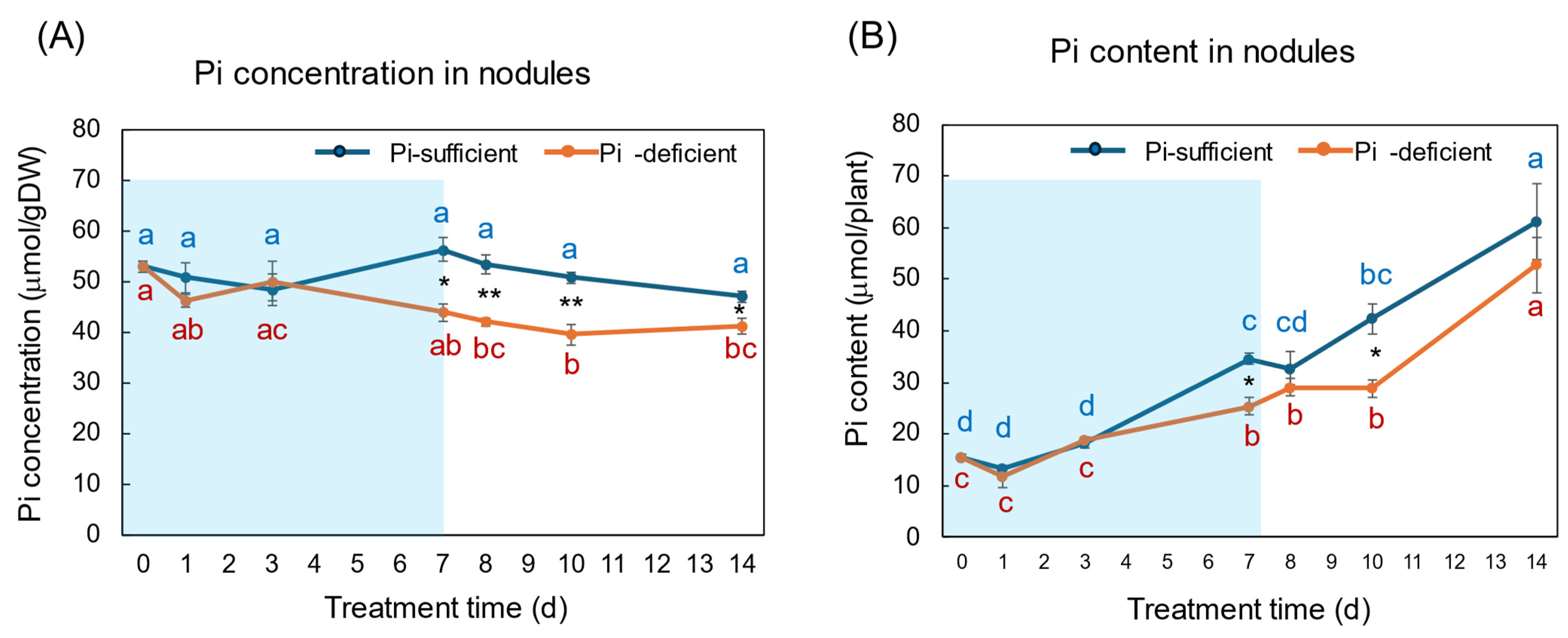
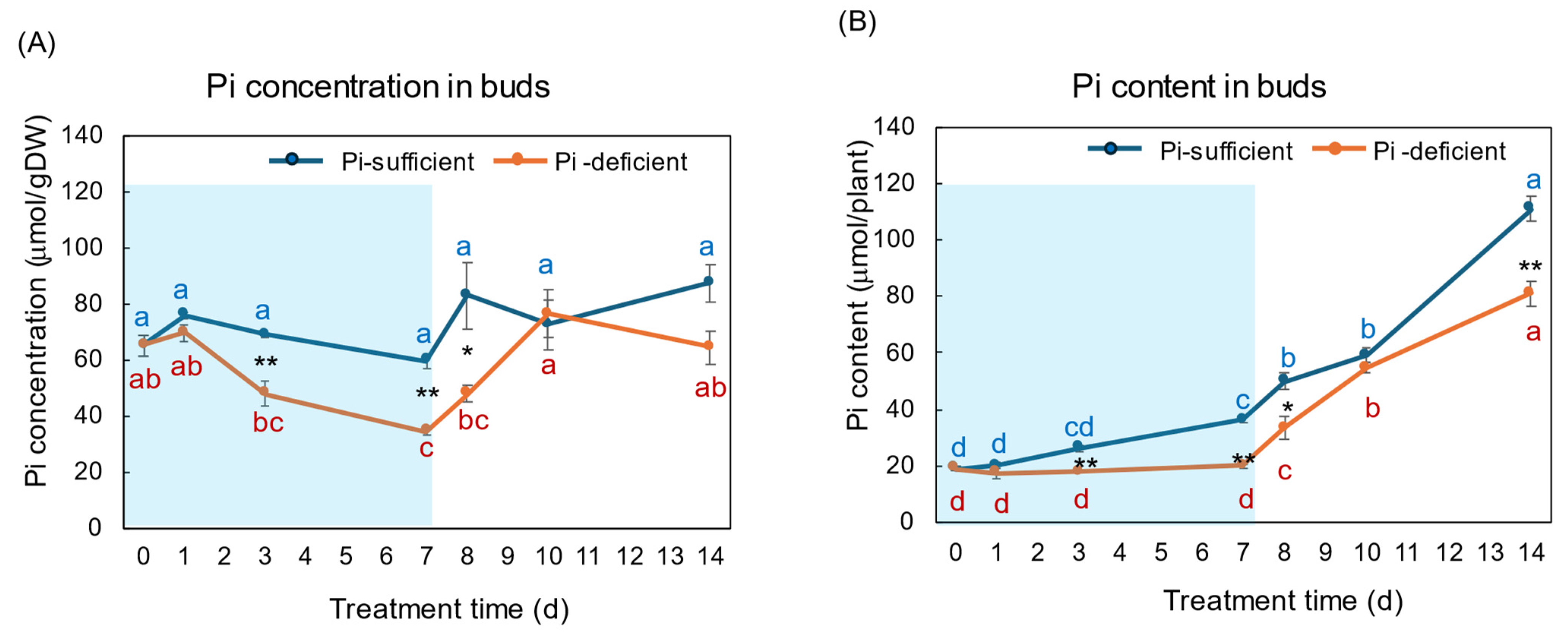
3. Results
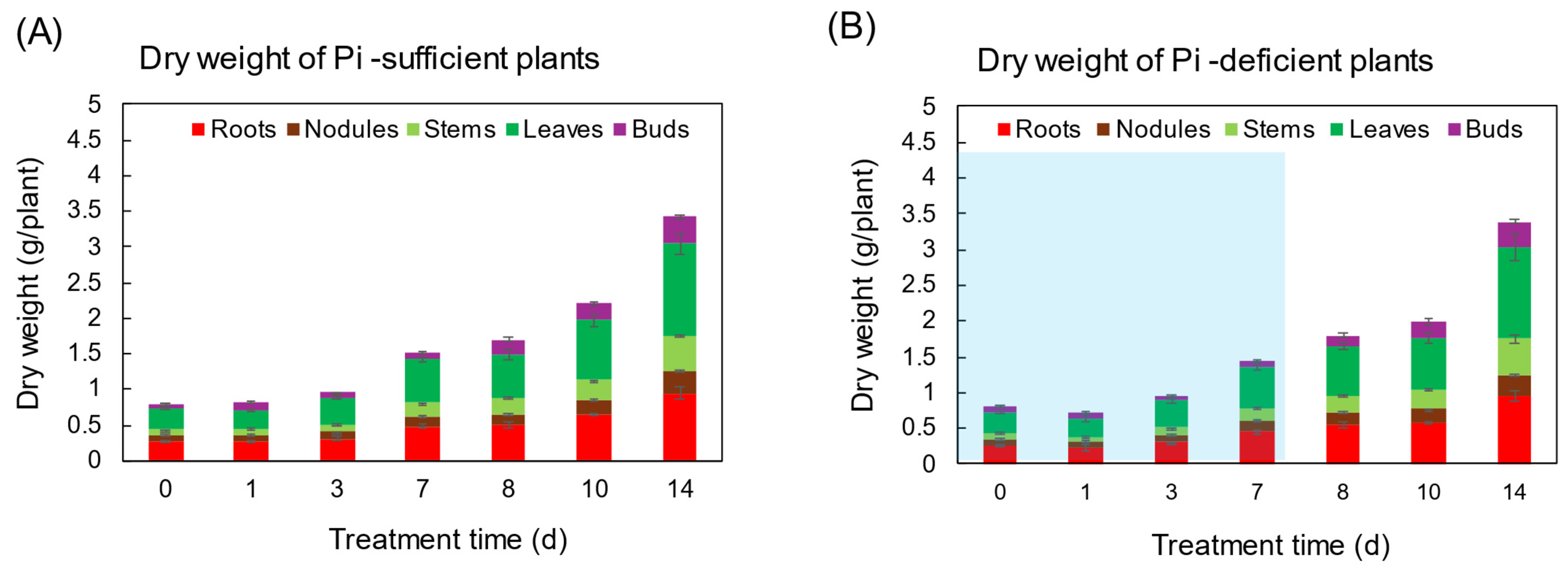
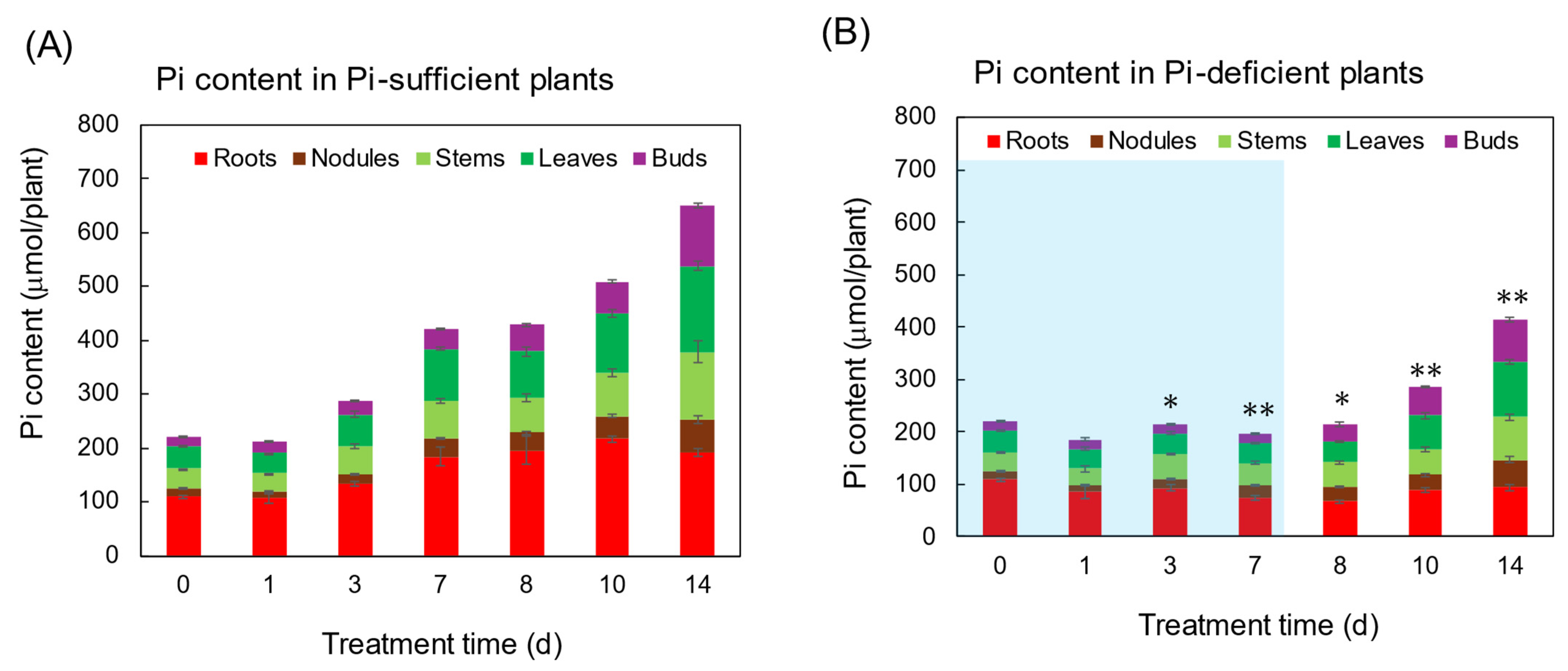
3.1. Comparison of the Growth and the P Status between Pi-Sufficient and Pi-Deficient Treatments
Comparison of the Dry Weight of Each Part
4. Discussion
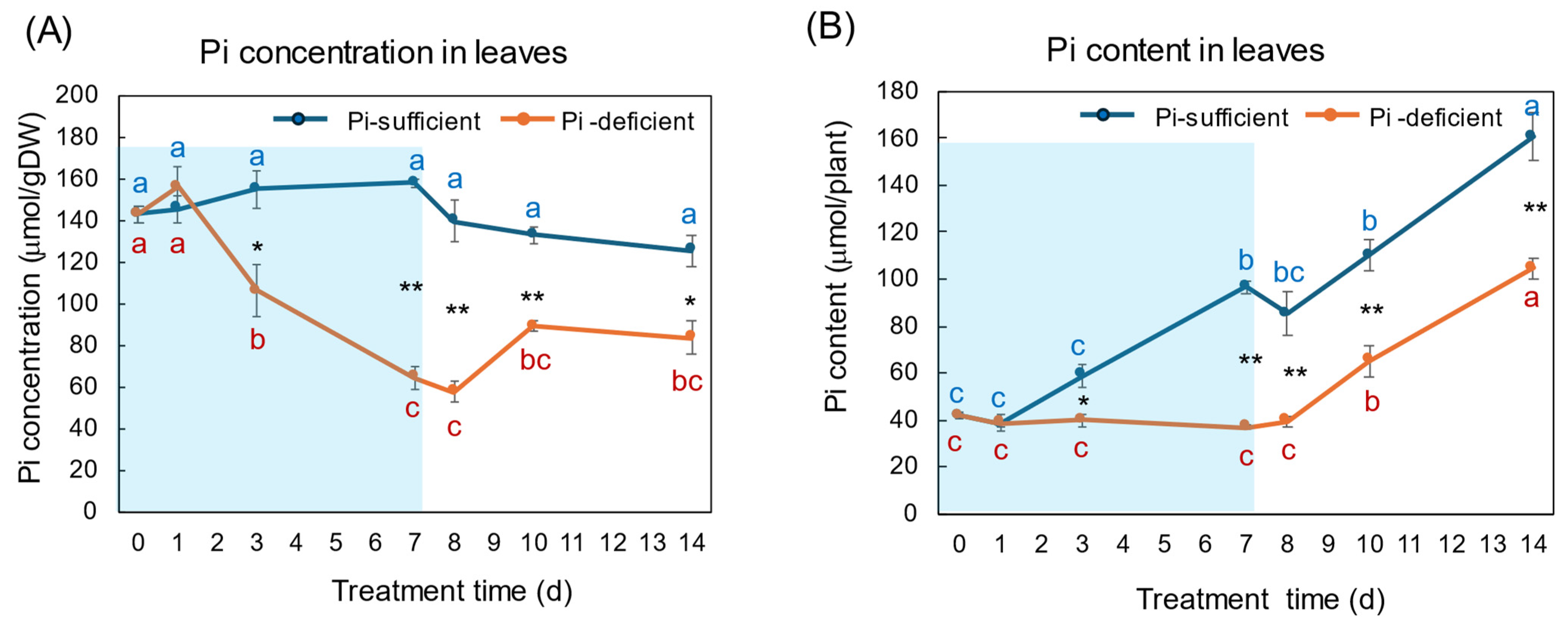
4.1. Changes in the Pi Concentration and Pi Content during Pi-Sufficient and Pi-Deficient Treatments
4.2. Comparison of the Pi Concentration and Pi Content in Each Part during Pi-Sufficient and Pi-Deficient Treatments
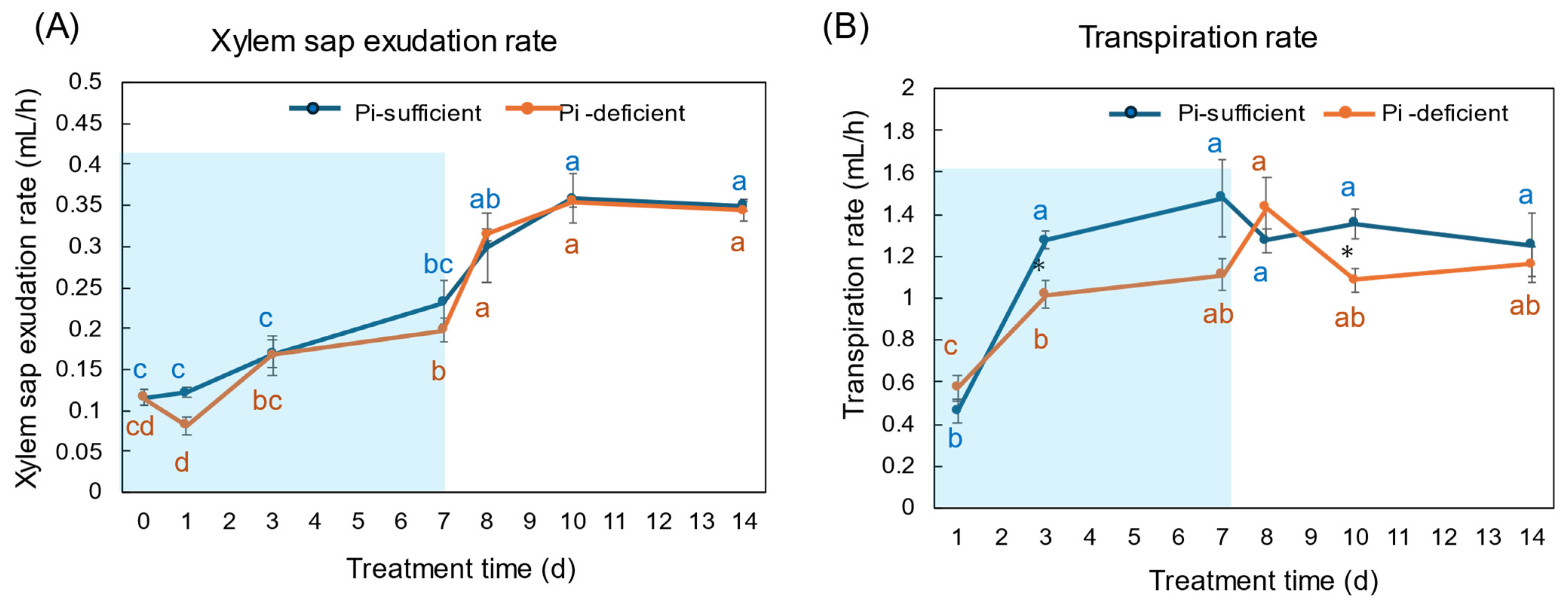
4.3. Changes in the Pi Concentration in Xylem Sap and Estimation of Pi Flux
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Marschner, H. Functions of mineral nutrients: Macronutrients. In Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: London, UK, 1986; pp. 229–312. [Google Scholar]
- Lambers, H. Phosphorus acquisition and utilization in plants. Annu. Rev. Plant Biol. 2022, 73, 17–42. [Google Scholar] [CrossRef]
- Lambers, H.; Plaxton, W.C. Phosphorus: Back to the roots. Annu. Plant Rev. 2015, 48, 3–22. [Google Scholar] [CrossRef]
- Sinclair, J.B. Soybeans. In Nutrient Deficiencies & Toxicities in Crop Plants; Bennett, W.F., Ed.; APS Press: St. Paul, MN, USA, 1993; pp. 99–103. [Google Scholar]
- Poirier, Y.; Bucher, M. Phosphate transport and homeostasis in Arabidopsis. Arab. Book 2002, 2002, e0024. [Google Scholar] [CrossRef] [PubMed]
- López-Arredondo, D.L.; Leyva-González, M.A.; González-Morales, S.I.; López-Bucio, J.; Herrera-Estrella, L. Phosphate nutrition: Improving low-phosphate tolerance in crops. Annu. Rev. Plant Biol. 2014, 65, 95–123. [Google Scholar] [CrossRef]
- Khan, F.; Siddique, A.B.; Shabala, S.; Zhou, M.; Zhao, C. Phosphorus plays key roles in regulating plants’ physiological responses to abiotic stresses. Plants 2023, 12, 2861. [Google Scholar] [CrossRef]
- Cassman, K.G.; Whitney, A.S.; Stockinger, K.R. Root growth and dry matter distribution of soybean as affected by phosphorus stress, nodulation, and nitrogen source. Crop Sci. 1980, 20, 239–244. [Google Scholar] [CrossRef]
- Israel, D.W. Investigation of the role of phosphorus in symbiotic dinitrogen fixation. Plant Physiol. 1987, 84, 835–840. [Google Scholar] [CrossRef]
- Ohyama, T.; Takayama, K.; Akagi, A.; Saito, A.; Higuchi, K.; Sato, T. Development of an N-free culture solution for cultivation of nodulated soybean with less pH fluctuation by the addition of potassium bicarbonate. Agriculture 2023, 13, 739. [Google Scholar] [CrossRef]
- Puga, M.I.; Rojas-Triana, M.; de Lorenzo, L.; Leyva, A.; Ruvio, V.; Paz-Ares, J. Novel signals in the regulation of Pi starvation responses in plants: Facts and promises. Curr. Opin. Plant Biol. 2017, 39, 40–49. [Google Scholar] [CrossRef]
- Chien, P.-S.; Chiang, C.-P.; Leong, S.J.; Chiou, T.-J. Sensing and signaling of phosphate starvation: From local to long distance. Plant Cell Physiol. 2018, 59, 1714–1722. [Google Scholar] [CrossRef]
- Goldstein, A.; Danon, A.; Baertlein, D.; McDaniel, R. Phosphate starvation inducible metabolism in Lycopersicon esculentum II. characterization of the phosphate starvation inducible excreted acid phosphatase. Plant Physiol. 1988, 87, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root structure and functioning for efficient acquisition of phosphorus: Matching morphological and physiological traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef]
- Bates, T.R.; Lynch, J.P. Stimulation of root hair elongation in Arabidopsis thaliana by low phosphorus availability. Plant Cell Environ. 1996, 19, 529–538. [Google Scholar] [CrossRef]
- Mo, X.; Liu, G.; Zhang, Z.; Liang, C.; Tian, J. Mechanisms underlying soybean response to phosphorus deficiency through integration of omics analysis. Int. J. Mol. Sci. 2022, 23, 4592. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-Y.; Huang, T.-K.; Kuo, H.-F.; Chiou, T.-J. Role of vacuoles in phosphorus storage and remobilization. J. Exper. Bot. 2017, 68, 3045–3055. [Google Scholar] [CrossRef]
- Bieleski, R.L. Effect of phosphorus deficiency on levels of phosphorus compounds in Spirodela. Plant Physiol. 1968, 43, 1309–1316. [Google Scholar] [CrossRef]
- Yamamura, Y.; Higuchi, K.; Saito, A.; Ohyama, T. Absorption and transport of phosphorus in nodulated soybean plants and diagnosis of phosphorus status using xylem sap analysis. Agriculture 2024, 14, 403. [Google Scholar] [CrossRef]
- Lauer, M.J.; Blevins, D.G.; Sierzputowska-Gracz, H. 31P-Nuclear Magnetic Resonance determination of phosphate compartmentation in leaves of reproductive soybeans (Glycine max L.) as affected by phosphate nutrition. Plant Physiol. 1989, 89, 1331–1336. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Lambers, H.; Bragg, J.; Finnegan, P.M.; Lovelock, C.E.; Plaxton, W.C.; Price, C.A.; Scheible, W.-R.; Michael, W.; Shane, M.W.; et al. Opportunities for improving phosphorus-use efficiency in crop plants. New Phytol. 2012, 195, 306–320. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, Y.; Nara, S.; Higuchi, K.; Saito, A.; Ohyama, T. Absorption and Xylem Transport of 33P-Labeled Phosphorus in Nodulated Soybean Plants. Agriculture 2024, 14, 1104. [Google Scholar] [CrossRef]
- Fujikake, H.; Yamazaki, A.; Ohtake, N.; Sueyoshi, K.; Matsuhashi, S.; Ito, T.; Mizuniwa, C.; Kume, T.; Hashimoto, S.; Ishioka, N.-S.; et al. Quick and reversible inhibition of soybean root nodule growth by nitrate involves a decrease in sucrose supply to nodules. J. Exper. Bot. 2003, 54, 1379–1388. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T.; Minagawa, R.; Ishikawa, S.; Yamamoto, M.; Hung, N.V.P.; Ohtake, N.; Sueyoshi, K.; Sato, T.; Nagumo, Y.; Takahashi, Y. Soybean seed production and nitrogen nutrition. In A Comprehensive Survey of International Soybean Research-Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar]
- Sakazume, T.; Tanaka, K.; Aida, H.; Ishikawa, S.; Nagumo, Y.; Takahashi, Y.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Estimation of nitrogen fixation rate of soybean (Glycine max L.) Merr.) by micro-scale relative ureide analysis using root bleeding xylem sap and apoplast fuid in stem. Bull. Facul. Agric. Niigata Univ. 2014, 67, 27–41. [Google Scholar]
- MEdical and PHarmaceutical Statistics (MEPHAS). Available online: www.gen-info.osaka-u.ac.jp/MEPHAS/mokuji1-e.html (accessed on 24 May 2002).
- Morita, S.; Abe, J. Diurnal and phenological changes of bleeding rate in lowland rice plants. Jpn. J. Crop Sci. 2002, 71, 383–388. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.; Liang, Q.; Lyu, X.; Li, S.; Gong, Z.; Dong, S.; Yan, C.; Ma, C. Regulation of phosphorus supply on nodulation and nitrogen fixation in soybean plants with dual-root systems. Agronomy 2021, 11, 2354. [Google Scholar] [CrossRef]
- Qin, L.; Guo, Y.; Chen, L.; Liang, R.; Gu, M.; Xu, G.; Zhao, J.; Walk, T.; Liao, H. Functional characterization of 14 Pht1 family genes in yeast and their expressions in response to nutrient starvation in soybean. PLoS ONE 2012, 7, e47726. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, M.I.; Adu-Gyamfi, J.J.; Saneoka, H.; Nguyen, N.T.; Suwa, R.; Kanai, S.; El-Shemy, H.A.; Lightfoot, D.A.; Fujita, K. The effect of phosphorus deficiency on nutrient uptake, nitrogen fixation and photosynthetic rate in mashbean, mungbean, and soybean. Acta Physiol. Plant 2008, 30, 537–544. [Google Scholar] [CrossRef]
- Fredeen, A.L.; Rao, I.M.; Terry, N. Influence of phosphorus nutrition on growth and carbon partitioning in Glycine max. Plant Physiol. 1989, 89, 225–230. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamura, Y.; Higuchi, K.; Saito, A.; Ohyama, T. Phosphate Turnover in Various Parts of Nodulated Soybean (Glycine max (L.) Merr.) Plants and the Relation to the Xylem Transport. Crops 2024, 4, 413-425. https://doi.org/10.3390/crops4030029
Yamamura Y, Higuchi K, Saito A, Ohyama T. Phosphate Turnover in Various Parts of Nodulated Soybean (Glycine max (L.) Merr.) Plants and the Relation to the Xylem Transport. Crops. 2024; 4(3):413-425. https://doi.org/10.3390/crops4030029
Chicago/Turabian StyleYamamura, Yoshiaki, Kyoko Higuchi, Akihiro Saito, and Takuji Ohyama. 2024. "Phosphate Turnover in Various Parts of Nodulated Soybean (Glycine max (L.) Merr.) Plants and the Relation to the Xylem Transport" Crops 4, no. 3: 413-425. https://doi.org/10.3390/crops4030029
APA StyleYamamura, Y., Higuchi, K., Saito, A., & Ohyama, T. (2024). Phosphate Turnover in Various Parts of Nodulated Soybean (Glycine max (L.) Merr.) Plants and the Relation to the Xylem Transport. Crops, 4(3), 413-425. https://doi.org/10.3390/crops4030029