
Animal, Herd and Feed Characteristics Associated with Blastocystis Prevalence and Molecular Diversity in Dairy Cattle from the North of France

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
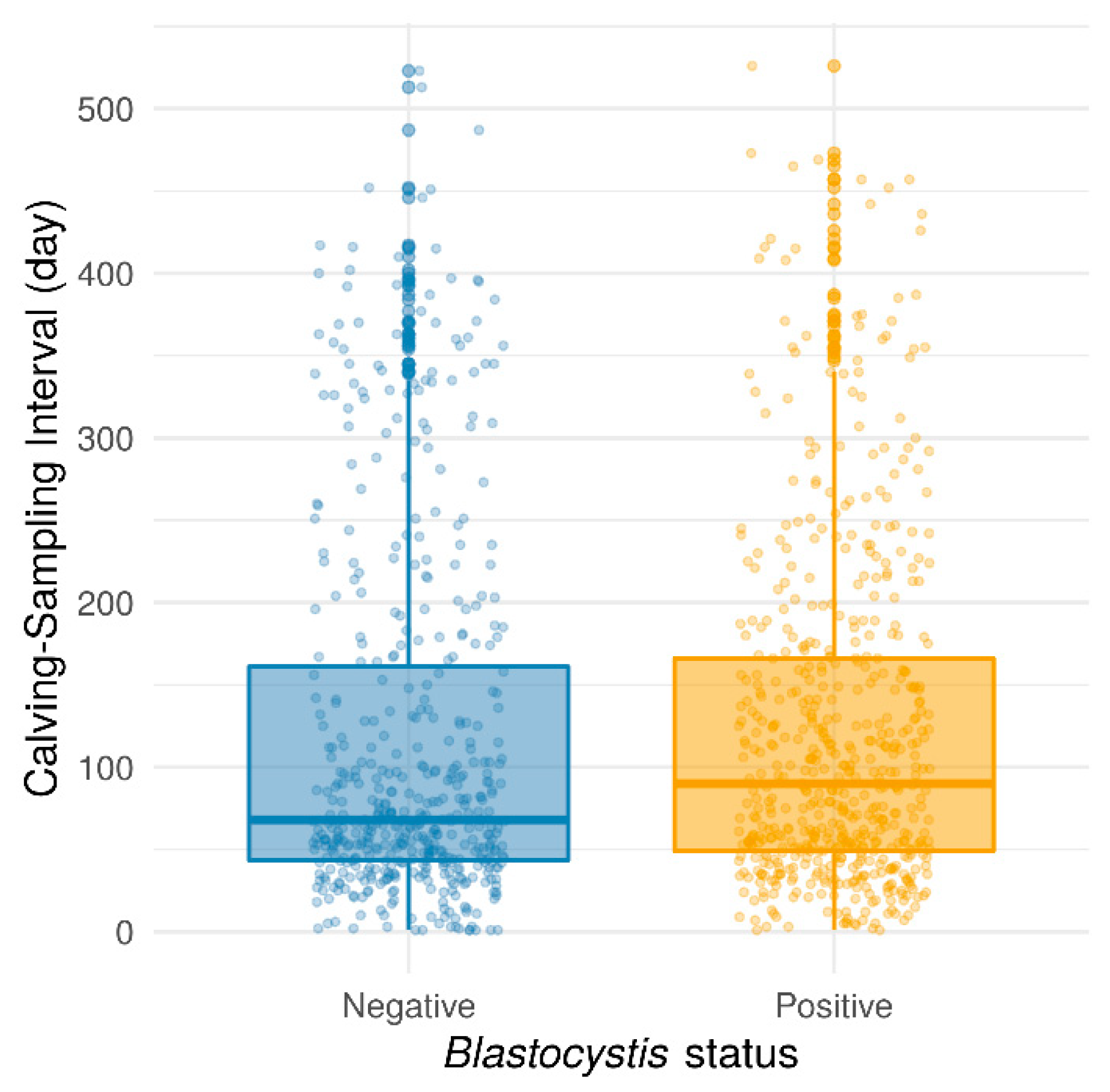
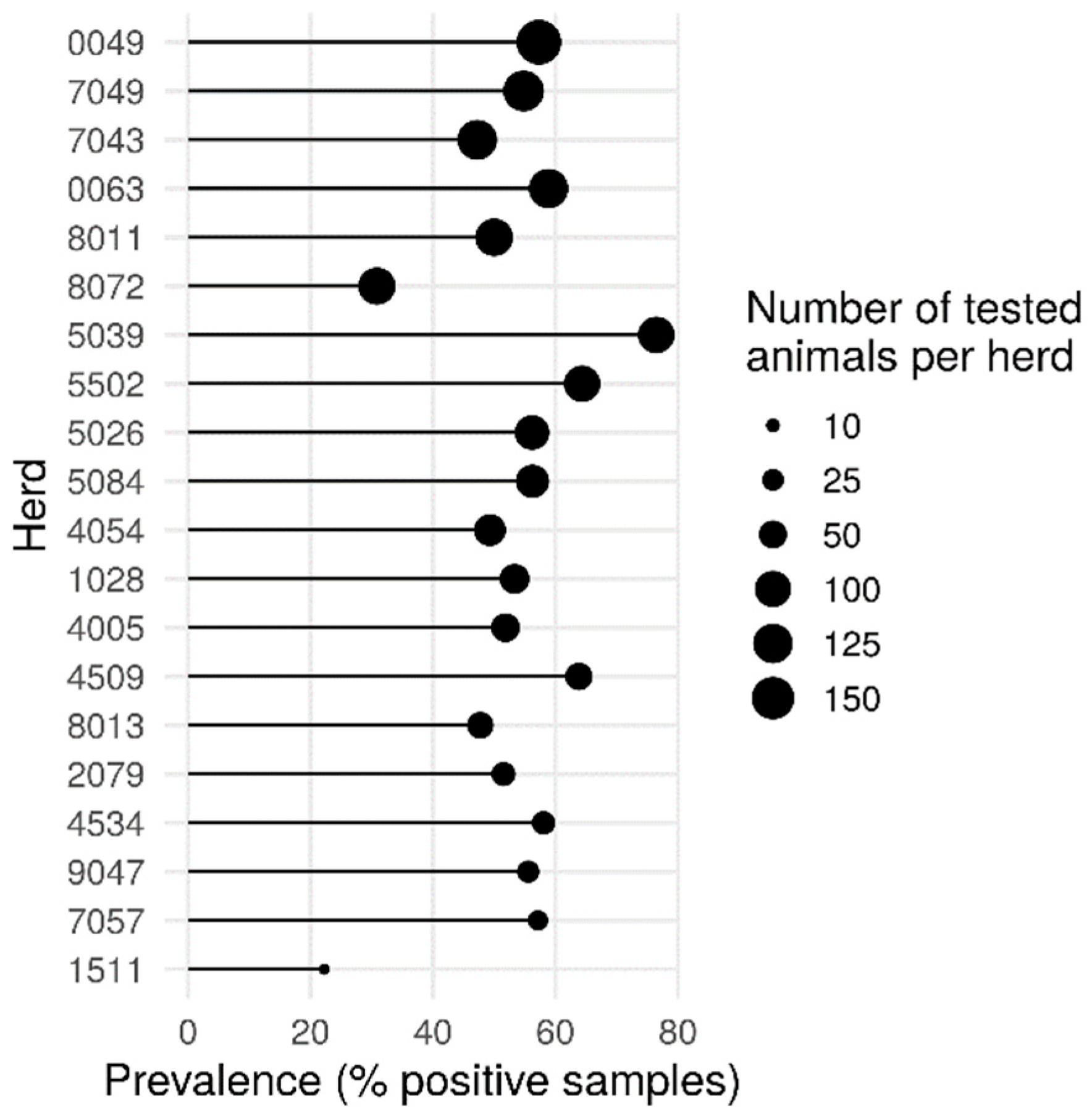
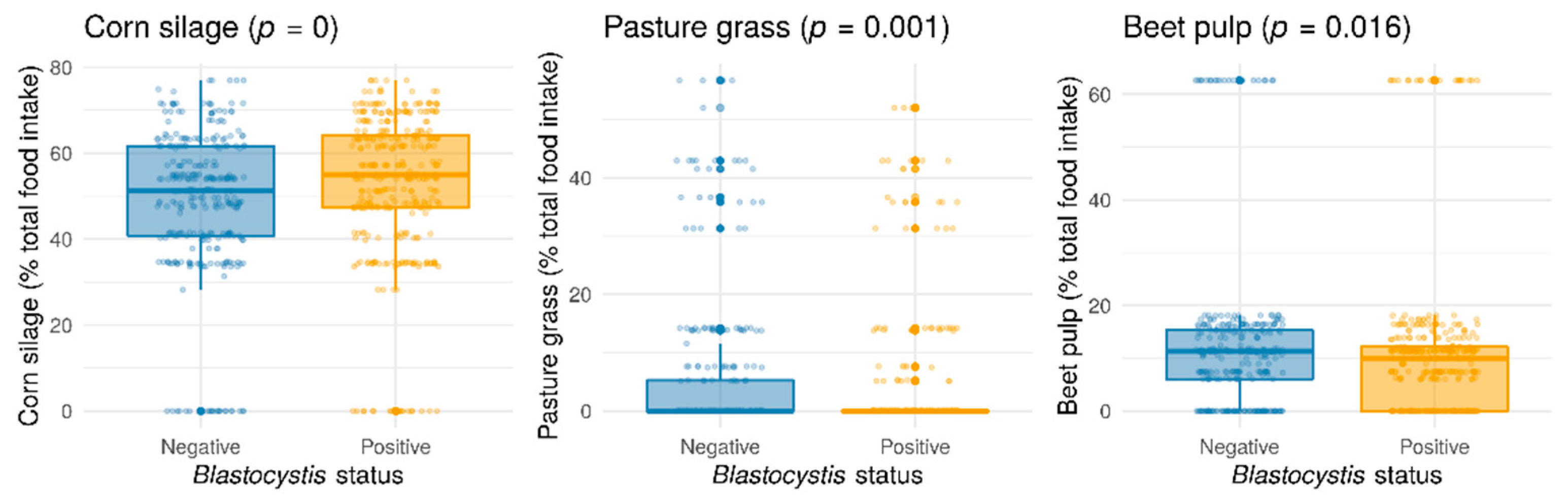
2. Results and Discussion
3. Materials and Methods
3.1. Sampling
3.2. Ethical Issues
3.3. Detection of Blastocystis sp. and Subtyping of the Isolates
3.4. Statistical Tests
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nemati, S.; Zali, M.R.; Johnson, P.; Mirjalali, H.; Karanis, P. Molecular prevalence and subtype distribution of Blastocystis sp. In Asia and in Australia. J. Water Health 2021, 19, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, P.A.; Jaimes, J.E.; Ramírez, J.D. A summary of Blastocystis subtypes in North and South America. Parasites Vectors 2019, 12, 376. [Google Scholar] [CrossRef] [PubMed]
- Rauff-Adedotun, A.A.; Termizi, F.H.M.; Shaari, N.; Lee, I.L. The coexistence of Blastocystis spp. in humans, animals and environmental sources from 2010–2021 in Asia. Biology 2021, 10, 990. [Google Scholar] [CrossRef]
- Tan, K.S.W. New insights on classification, identification, and clinical relevance of Blastocystis spp. Clin. Microbiol. Rev. 2008, 21, 639–665. [Google Scholar] [CrossRef] [Green Version]
- Scanlan, P.D.; Stensvold, C.R.; Rajilić-Stojanović, M.; Heilig, H.G.H.J.; De Vos, W.M.; O’Toole, P.W.; Cotter, P.D. The microbial eukaryote Blastocystis is a prevalent and diverse member of the healthy human gut microbiota. FEMS Microbiol. Ecol. 2014, 90, 326–330. [Google Scholar] [CrossRef] [Green Version]
- Andersen, L.O.; Stensvold, C.R. Blastocystis in Health and Disease: Are We Moving from a Clinical to a Public Health Perspective? J. Clin. Microbiol. 2016, 54, 524–528. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Clark, C.G. Current status of Blastocystis: A personal view. Parasitol. Int. 2016, 65, 763–771. [Google Scholar] [CrossRef]
- Ajjampur, S.S.R.; Tan, K.S.W. Pathogenic mechanisms in Blastocystis spp.—Interpreting results from in vitro and in vivo studies. Parasitol. Int. 2016, 65, 772–779. [Google Scholar] [CrossRef]
- Hublin, J.S.Y.; Maloney, J.G.; Santin, M. Blastocystis in domesticated and wild mammals and birds. Res. Vet. Sci. 2021, 135, 260–282. [Google Scholar] [CrossRef]
- Shams, M.; Shamsi, L.; Sadrebazzaz, A.; Asghari, A.; Badali, R.; Omidian, M.; Hassanipour, S. A systematic review and meta-analysis on the global prevalence and subtypes distribution of Blastocystis sp. infection in cattle: A zoonotic concern. Comp. Immunol. Microbiol. Infect. Dis. 2021, 76, 101650. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Taner-Mulla, D.; Jacob, A.S.; Imeede, C.A.; Yoshikawa, H.; Stensvold, C.R.; Clark, C.G. Genetic Diversity of Blastocystis in Livestock and Zoo Animals. Protist 2013, 164, 497–509. [Google Scholar] [CrossRef] [Green Version]
- Stensvold, C.R.; Alfellani, M.A.; Nørskov-Lauritsen, S.; Prip, K.; Victory, E.L.; Maddox, C.; Nielsen, H.V.; Clark, C.G. Subtype distribution of Blastocystis isolates from synanthropic and zoo animals and identification of a new subtype. Int. J. Parasitol. 2009, 39, 473–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abarca, N.; Santín, M.; Ortega, S.; Maloney, J.G.; George, N.S.; Molokin, A.; Cardona, G.A.; Dashti, A.; Köster, P.C.; Bailo, B.; et al. Molecular detection and characterization of Blastocystis sp. And Enterocytozoon bieneusi in cattle in northern spain. Vet. Sci. 2021, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Stensvold, C.R.; Clark, C.G. Pre-empting Pandora’s Box: Blastocystis Subtypes Revisited. Trends Parasitol. 2020, 36, 229–232. [Google Scholar] [CrossRef]
- Higuera, A.; Herrera, G.; Jimenez, P.; García-Corredor, D.; Pulido-Medellín, M.; Bulla-Castañeda, D.M.; Pinilla, J.C.; Moreno-Pérez, D.A.; Maloney, J.G.; Santín, M.; et al. Identification of Multiple Blastocystis Subtypes in Domestic Animals from Colombia Using Amplicon-Based Next Generation Sequencing. Front. Vet. Sci. 2021, 8, 732129. [Google Scholar] [CrossRef]
- Maloney, J.G.; Jang, Y.; Molokin, A.; George, N.S.; Santin, M. Wide Genetic Diversity of Blastocystis in White-Tailed Deer (Odocoileus virginianus) from Maryland, USA. Microorganisms 2021, 9, 1343. [Google Scholar] [CrossRef]
- Alfellani, M.A.; Stensvold, C.R.; Vidal-Lapiedra, A.; Onuoha, E.S.U.; Fagbenro-Beyioku, A.F.; Clark, C.G. Variable geographic distribution of Blastocystis subtypes and its potential implications. Acta Trop. 2013, 126, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, J.D.; Sánchez, A.; Hernández, C.; Flórez, C.; Bernal, M.C.; Giraldo, J.C.; Reyes, P.; López, M.C.; García, L.; Cooper, P.J.; et al. Geographic distribution of human Blastocystis subtypes in South America. Infect. Genet. Evol. 2016, 41, 32–35. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Ly, A.T.; Senghor, S.; Even, G.; Dautel, E.; Dejager, R.; Sawant, M.; Baydoun, M.; Benamrouz-Vanneste, S.; et al. Prevalence and subtype distribution of Blastocystis sp. in senegalese school children. Microorganisms 2020, 8, 1408. [Google Scholar] [CrossRef]
- Khaled, S.; Gantois, N.; Ayoubi, A.; Even, G.; Sawant, M.; El Houmayraa, J.; Nabot, M.; Benamrouz-Vanneste, S.; Chabé, M.; Certad, G.; et al. Blastocystis sp. Prevalence and subtypes distribution amongst syrian refugee communities living in North Lebanon. Microorganisms 2021, 9, 184. [Google Scholar] [CrossRef]
- Osorio-Pulgarin, M.I.; Higuera, A.; Beltran-álzate, J.C.; Sánchez-Jiménez, M.; Ramírez, J.D. Epidemiological and molecular characterization of Blastocystis infection in children attending daycare centers in Medellín, Colombia. Biology 2021, 10, 669. [Google Scholar] [CrossRef] [PubMed]
- Greige, S.; El Safadi, D.; Khaled, S.; Gantois, N.; Baydoun, M.; Chemaly, M.; Benamrouz-Vanneste, S.; Chabé, M.; Osman, M.; Certad, G.; et al. First report on the prevalence and subtype distribution of Blastocystis sp. in dairy cattle in Lebanon and assessment of zoonotic transmission. Acta Trop. 2019, 194, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Masuda, A.; Sumiyoshi, T.; Ohtaki, T.; Matsumoto, J. Prevalence and molecular subtyping of Blastocystis from dairy cattle in Kanagawa, Japan. Parasitol. Int. 2018, 67, 702–705. [Google Scholar] [CrossRef] [PubMed]
- Udonsom, R.; Prasertbun, R.; Mahittikorn, A.; Mori, H.; Changbunjong, T.; Komalamisra, C.; Pintong, A.; Sukthana, Y.; Popruk, S. Blastocystis infection and subtype distribution in humans, cattle, goats, and pigs in central and western Thailand. Infect. Genet. Evol. 2018, 65, 107–111. [Google Scholar] [CrossRef]
- Ramírez, J.D.; Sánchez, L.V.; Bautista, D.C.; Corredor, A.F.; Flórez, A.C.; Stensvold, C.R. Blastocystis subtypes detected in humans and animals from Colombia. Infect. Genet. Evol. 2014, 22, 223–228. [Google Scholar] [CrossRef]
- Hemalatha, C.; Chandrawathani, P.; Kumar, S.G.; Premaalatha, B.; Geethamalar, S.; Rozita, L.M.H.; Haziqah, F.M.T.; Sabapathy, D.; Ramlan, M. The diagnosis of Blastocystis sp. from animals—An emerging zoonosis. Malays. J. Vet. Res. 2014, 5, 15–22. [Google Scholar]
- Sharifi, Y.; Abbasi, F.; Shahabi, S.; Zaraei, A.; Mikaeili, F.; Sarkari, B. Comparative genotyping of Blastocystis infecting cattle and human in the south of Iran. Comp. Immunol. Microbiol. Infect. Dis. 2020, 72, 101529. [Google Scholar] [CrossRef]
- Rostami, M.; Fasihi-Harandi, M.; Shafiei, R.; Aspatwar, A.; Derakhshan, F.K.; Raeghi, S. Genetic diversity analysis of Blastocystis subtypes and their distribution among the domestic animals and pigeons in northwest of Iran. Infect. Genet. Evol. 2020, 86, 104591. [Google Scholar] [CrossRef]
- Lee, H.; Lee, S.H.; Seo, M.G.; Kim, H.Y.; Kim, J.W.; Lee, Y.R.; Kim, J.H.; Kwon, O.D.; Kwak, D. Occurrence and genetic diversity of Blastocystis in Korean cattle. Vet. Parasitol. 2018, 258, 70–73. [Google Scholar] [CrossRef]
- Zhu, W.; Tao, W.; Gong, B.; Yang, H.; Li, Y.; Song, M.; Lu, Y.; Li, W. First report of Blastocystis infections in cattle in China. Vet. Parasitol. 2017, 246, 38–42. [Google Scholar] [CrossRef]
- Wang, J.; Gong, B.; Yang, F.; Zhang, W.; Zheng, Y.; Liu, A. Subtype distribution and genetic characterizations of Blastocystis in pigs, cattle, sheep and goats in northeastern China’s Heilongjiang Province. Infect. Genet. Evol. 2018, 57, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Ren, M.; Song, J.K.; Yang, F.; Zou, M.; Wang, P.X.; Wang, D.; Zhang, H.J.; Zhao, G.H.; Lin, Q. First genotyping of Blastocystis in yaks from Qinghai Province, northwestern China. Parasites Vectors 2019, 12, 171. [Google Scholar] [CrossRef] [PubMed]
- Badparva, E.; Sadraee, J.; Kheirandish, F. Genetic diversity of Blastocystis isolated from cattle in Khorramabad, Iran. Jundishapur J. Microbiol. 2015, 8, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Aynur, Z.E.; Güçlü, Ö.; Yildiz, I.; Aynur, H.; Hertabaklar, H.; Bozdogan, B.; Ertug, S. Molecular characterization of Blastocystis in cattle in Turkey. Parasitol. Res. 2019, 118, 1055–1059. [Google Scholar] [CrossRef] [PubMed]
- Moura, R.G.F.; de Oliveira-Silva, M.B.; Pedrosa, A.L.; Nascentes, G.A.N.; Cabrine-Santos, M. Occurrence of Blastocystis spp. in domestic animals in triângulo mineiro area of Brazil. Rev. Soc. Bras. Med. Trop. 2018, 51, 240–243. [Google Scholar] [CrossRef] [Green Version]
- AbuOdeh, R.; Ezzedine, S.; Madkour, M.; Stensvold, C.R.; Samie, A.; Nasrallah, G.; AlAbsi, E.; ElBakri, A. Molecular Subtyping of Blastocystis from Diverse Animals in the United Arab Emirates. Protist 2019, 170, 125679. [Google Scholar] [CrossRef]
- Fayer, R.; Santin, M.; MacArisin, D. Detection of concurrent infection of dairy cattle with Blastocystis, Cryptosporidium, Giardia, and Enterocytozoon by molecular and microscopic methods. Parasitol. Res. 2012, 111, 1349–1355. [Google Scholar] [CrossRef]
- Maloney, J.G.; Lombard, J.E.; Urie, N.J.; Shivley, C.B.; Santin, M. Zoonotic and genetically diverse subtypes of Blastocystis in US pre-weaned dairy heifer calves. Parasitol. Res. 2019, 118, 575–582. [Google Scholar] [CrossRef]
- Kamaruddin, S.K.; Mat Yusof, A.; Mohammad, M. Prevalence and subtype distribution of Blastocystis sp. in cattle from Pahang, Malaysia. Trop. Biomed. 2020, 37, 127–141. [Google Scholar]
- Abdullah, D. Molecular Epidemiology, Risk Factors of Zoonotic Enteric Protozoa and Genetic Diversity of Blastocystis Infecting Cattle in Peninsular Malaysia. Ph.D. Thesis, Universiti Putra Malysia, Seri Kembangan, Malaysia, 2017. [Google Scholar]
- Santín, M.; Gómez-Muñoz, M.T.; Solano-Aguilar, G.; Fayer, R. Development of a new PCR protocol to detect and subtype Blastocystis spp. from humans and animals. Parasitol. Res. 2011, 109, 205–212. [Google Scholar] [CrossRef]
- Maloney, J.G.; Santin, M. Mind the gap: New full-length sequences of Blastocystis subtypes generated via oxford nanopore minion sequencing allow for comparisons between full-length and partial sequences of the small subunit of the ribosomal rna gene. Microorganisms 2021, 9, 997. [Google Scholar] [CrossRef] [PubMed]
- El Safadi, D.; Cian, A.; Nourrisson, C.; Pereira, B.; Morelle, C.; Bastien, P.; Bellanger, A.P.; Botterel, F.; Candolfi, E.; Desoubeaux, G.; et al. Prevalence, risk factors for infection and subtype distribution of the intestinal parasite Blastocystis sp. from a large-scale multi-center study in France. BMC Infect. Dis. 2016, 16, 451. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on mycotoxin issues in ruminants: Occurrence in forages, effects of mycotoxin ingestion on health status and animal performance and practical strategies to counteract their negative effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Evans, E.; Messerschmidt, U. Review: Sugar beets as a substitute for grain for lactating dairy cattle. J. Anim. Sci. Biotechnol. 2017, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Poirier, P.; Wawrzyniak, I.; Albert, A.; El Alaoui, H.; Delbac, F.; Livrelli, V. Development and evaluation of a real-time PCR assay for detection and quantification of Blastocystis parasites in human stool samples: Prospective study of patients with hematological malignancies. J. Clin. Microbiol. 2011, 49, 975–983. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Audebert, C.; Gantois, N.; Ducrocq, S.; Darras, M.; Merlin, S.; Martel, S.; Viscogliosi, E.; Even, G.; Chabé, M. Animal, Herd and Feed Characteristics Associated with Blastocystis Prevalence and Molecular Diversity in Dairy Cattle from the North of France. Parasitologia 2022, 2, 45-53. https://doi.org/10.3390/parasitologia2010005
Audebert C, Gantois N, Ducrocq S, Darras M, Merlin S, Martel S, Viscogliosi E, Even G, Chabé M. Animal, Herd and Feed Characteristics Associated with Blastocystis Prevalence and Molecular Diversity in Dairy Cattle from the North of France. Parasitologia. 2022; 2(1):45-53. https://doi.org/10.3390/parasitologia2010005
Chicago/Turabian StyleAudebert, Christophe, Nausicaa Gantois, Sébastien Ducrocq, Marianne Darras, Sophie Merlin, Sophie Martel, Eric Viscogliosi, Gaël Even, and Magali Chabé. 2022. "Animal, Herd and Feed Characteristics Associated with Blastocystis Prevalence and Molecular Diversity in Dairy Cattle from the North of France" Parasitologia 2, no. 1: 45-53. https://doi.org/10.3390/parasitologia2010005
APA StyleAudebert, C., Gantois, N., Ducrocq, S., Darras, M., Merlin, S., Martel, S., Viscogliosi, E., Even, G., & Chabé, M. (2022). Animal, Herd and Feed Characteristics Associated with Blastocystis Prevalence and Molecular Diversity in Dairy Cattle from the North of France. Parasitologia, 2(1), 45-53. https://doi.org/10.3390/parasitologia2010005