
Comparative Osteology, Phylogeny and Classification of the Eastern South American Catfish Genus Trichomycterus (Siluriformes: Trichomycteridae)


Abstract

1. Introduction
History of Trichomycterus Intrageneric Classification
2. Materials and Methods
2.1. Comparative Osteology
2.2. DNA Sequences
2.3. Phylogenetic Analyses
2.4. Taxonomic Accounts
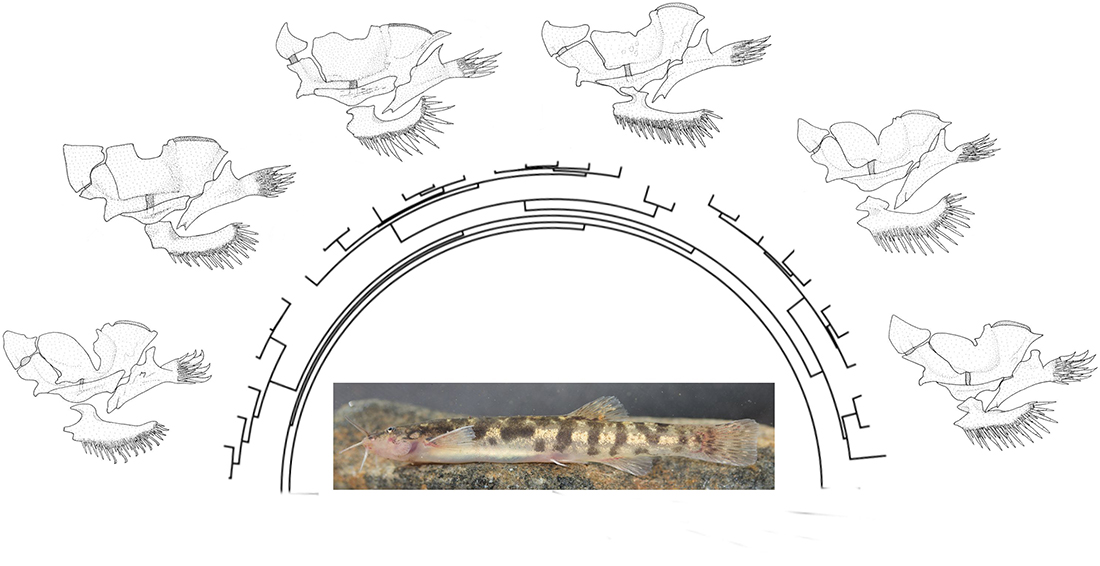
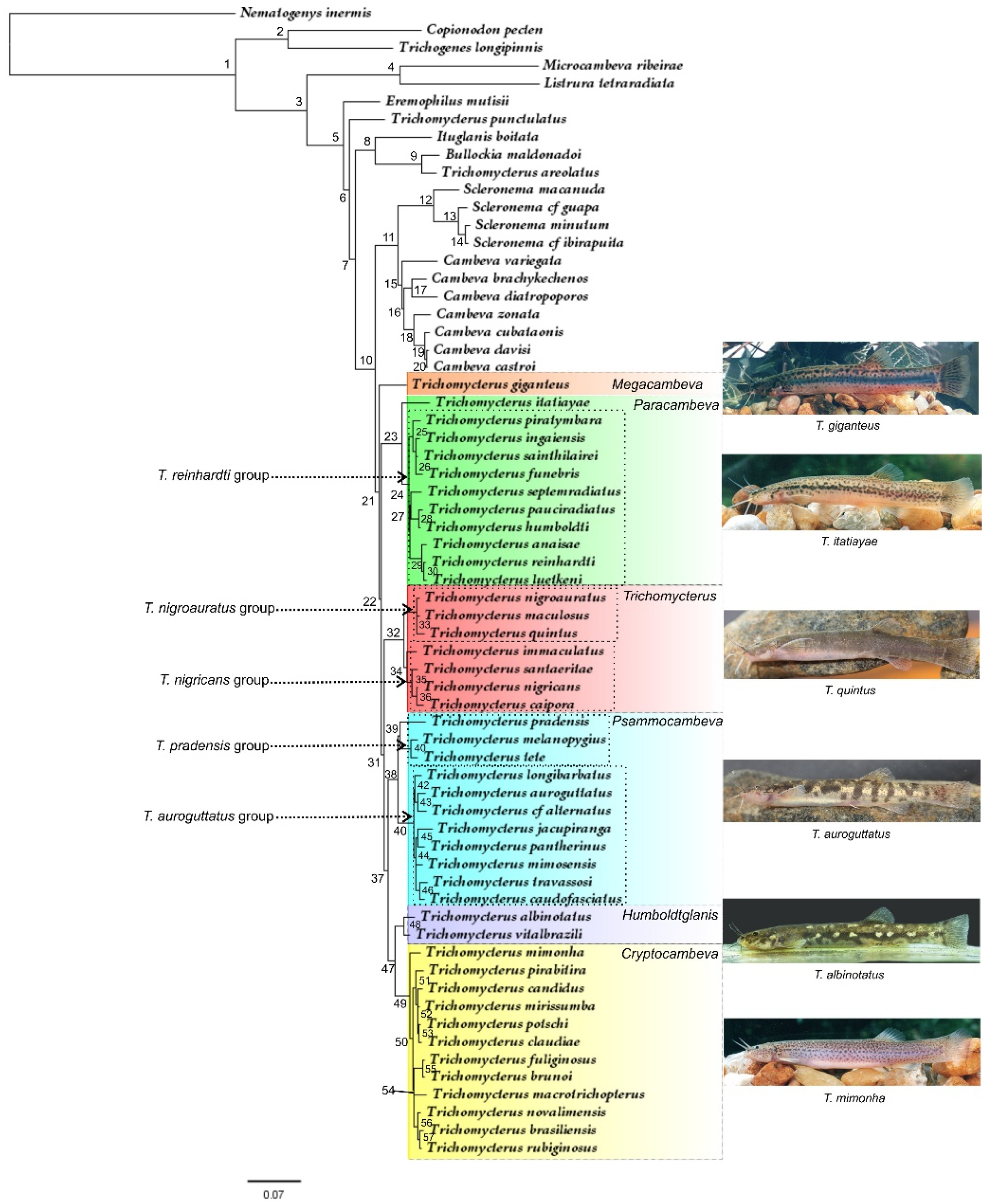
3. Results
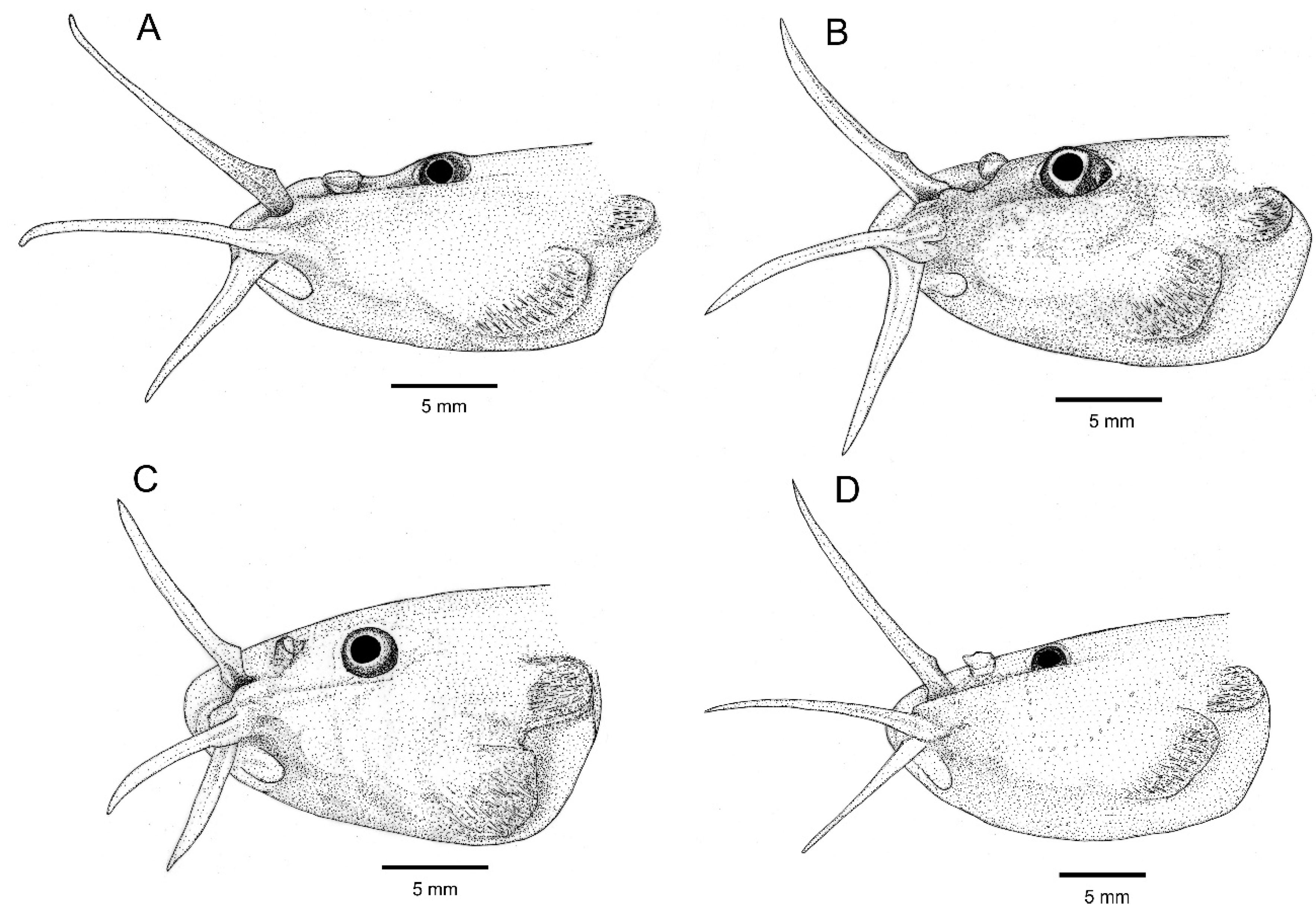
3.1. Comparative Osteology and Phylogeny
3.2. Taxonomic Accounts
3.2.1. Megacambeva, New Subgenus
LSID: urn:lsid:zoobank.org:act:DD5D2BBB-A0E2-4D6F-96F1-229A811ADCE7
3.2.2. Paracambeva, New Subgenus
LSID: urn:lsid:zoobank.org:act:F5037571-D95D-4091-880B-9E598AF9ED9F
3.2.3. Subgenus Trichomycterus Valenciennes 1832
3.2.4. Psammocambeva, New Subgenus
LSID: urn:lsid:zoobank.org:act:4ED967A8-D2DB-43F1-9918-6A3384652165
3.2.5. Humboldtglanis, New Subgenus
LSID: urn:lsid:zoobank.org:act:09886ED2-C1BC-4A84-8824-07E6EEA8B510
3.2.6. Cryptocambeva, New Subgenus
LSID: urn:lsid:zoobank.org:act:E0BE3FF5-ABAE-4881-9875-5FA5C23FE2C4
4. Discussion
4.1. Phylogeny
4.2. Morphological Diversity of Osteological Structures
4.3. CST-Clade and Trichomycterus s.s. Diagnosis
5. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | COI | CYTB | MYH6 | RAG2 |
|---|---|---|---|---|
| Nematogenys inermis | KY857952 | ––––– | KY858107 | KY858182 |
| Copionodon pecten | KY857929 | ––––– | KY858084 | KY858169 |
| Trichogenes longipinnis | MK123682 | MK123704 | MF431104 | MF431117 |
| Microcambeva ribeirae | MN385807 | ––––– | MN385819 | MN385832 |
| Listrura tetraradiata | JQ231083 | JQ231088 | MN385814 | MN385826 |
| Eremophilus mutisii | KY857931 | ––––– | KY858086 | KY858171 |
| ‘Trichomycterus’ punctulatus | KY857983 | ––––– | KY858134 | KY858203 |
| Bullockia maldonadoi | KY857926 | FJ772237 | KY858081 | KY858166 |
| Hatcheria areolata | KY857964 | FJ772214 | ––––– | KY858188 |
| Ituglanis boitata | MK123684 | MK123706 | MF431105 | MK123758 |
| Scleronema minutum | MK123685 | MK123707 | MK123735 | MK123759 |
| Scleronema cf. guapa | MK123687 | MK123709 | MF431107 | MF431118 |
| Scleronema cf. ibirapuita | MK123688 | MK123710 | MK123737 | MK123761 |
| Scleronema macanuda | MK123686 | MK123708 | MK123736 | MK123760 |
| Cambeva variegata | KY857991 | KY858060 | ––––– | KY858211 |
| Cambeva brachykechenos | MN995669 | MK123711 | MF431108 | ––––– |
| Cambeva diatropoporos | KY857996 | KY858065 | KY858147 | KY858213 |
| Cambeva cubataonis | MK123689 | MK123713 | MK123740 | MN385820 |
| Cambeva davisi | KR140345 | MK123714 | MK123741 | MK123762 |
| Cambeva castroi | ––––– | MK123712 | MK123739 | ––––– |
| Cambeva zonata | KY857986 | KY858053 | KY858137 | ––––– |
| Trichomycterus itatiayae | KY857977 | KY858046 | KY858128 | KY858198 |
| Trichomycterus septemradiatus | MK123700 | MK123729 | MK123755 | MW196781 |
| Trichomycterus pauciradiatus | MT941796 | MT941833 | MW196769 | MW196782 |
| Trichomycterus piratymbara | KY857970 | KY858040 | KY858121 | KY858194 |
| Trichomycterus reinhardti | MK123698 | MK123727 | MF431106 | MF431119 |
| Trichomycterus anaisae | MT941782 | MT941820 | ––––– | ––––– |
| Trichomycterus sainthilairei | MT941815 | MT941853 | ––––– | ––––– |
| Trichomycterus luetkeni | MT941793 | MT941831 | KY858148 | KY858214 |
| Trichomycterus ingaiensis | MT941790 | MT941829 | ––––– | ––––– |
| Trichomycterus funebris | MT941785 | MT941823 | KY858121 | KY858194 |
| Trichomycterus humboldti | MT941787 | MT941824 | ––––– | ––––– |
| Trichomycterus giganteus | MK123693 | MK123720 | MK123746 | MT446426 |
| Trichomycterus nigricans | MN813005 | MK123723 | MK123750 | MK123765 |
| Trichomycterus caipora | MN813008 | MN813000 | MN812996 | MN812992 |
| Trichomycterus santaeritae | MN813009 | MN813001 | MN812997 | MN812993 |
| Trichomycterus immaculatus | MK123694 | MK144348 | MK123747 | MF431120 |
| Trichomycterus nigroauratus | MK123696 | MK123724 | MK123751 | MK123766 |
| Trichomycterus maculosus | MN813010 | MN813002 | MN812998 | MN812994 |
| Trichomycterus quintus | MT299917 | MN812999 | MT305242 | ––––– |
| Trichomycterus mimonha | MW196749 | MW196758 | MW196770 | MW196783 |
| Trichomycterus fuliginosus | MW196750 | MW196759 | MW196771 | MW196784 |
| Trichomycterus brunoi | MW196751 | MW196760 | MW196772 | MW196785 |
| Trichomycterus mirissumba | MW196752 | MW196761 | MW196773 | MW196786 |
| Trichomycterus pirabitira | KY857982 | KY858051 | KY858133 | ––––– |
| Trichomycterus macrotrichopterus | MW196753 | MW196762 | MW196774 | MW196787 |
| Trichomycterus brasiliensis | MK123691 | MK123717 | MK123744 | MK123763 |
| Trichomycterus candidus | MK123692 | MK123718 | MK123745 | MW196788 |
| Trichomycterus potschi | ––––– | MW196763 | MW196775 | MW196789 |
| Trichomycterus claudiae | MW196754 | MW196764 | MW196776 | MW196790 |
| Trichomycterus novalimensis | MW196755 | MW196765 | MW196777 | MW196791 |
| Trichomycterus rubiginosus | MK123699 | MK123728 | MK123754 | MK123767 |
| Trichomycterus albinotatus | MN813007 | MK123716 | MK123743 | MN812990 |
| Trichomycterus vitalbrazili | MT435137 | MK748279 | MK748280 | MT446428 |
| Trichomycterus travassosi | MK123701 | MK123730 | MK123757 | ––––– |
| Trichomycterus caudofasciatus | MN812995 | MK123719 | MK123719 | MK123764 |
| Trichomycterus jacupiranga | MK123702 | MK123731 | MK123757 | ––––– |
| Trichomycterus pantherinus | MK123697 | MK123725 | MK123752 | MN812989 |
| Trichomycterus auroguttatus | MT435135 | MT436452 | MT436450 | ––––– |
| Trichomycterus alternatus | MK123690 | MK123715 | MK123742 | MN812991 |
| Trichomycterus melanopygus | KY857976 | KY858045 | KY858127 | KY858197 |
| Trichomycterus pradensis | MN813003 | MK123726 | MK123753 | MN812988 |
| Trichomycterus tete | ––––– | MH620991 | ––––– | ––––– |
| Trichomycterus mimosensis | ––––– | MH621150 | ––––– | ––––– |
| Trichomycterus longibarbatus | ––––– | MH621258 | ––––– | ––––– |
Appendix C
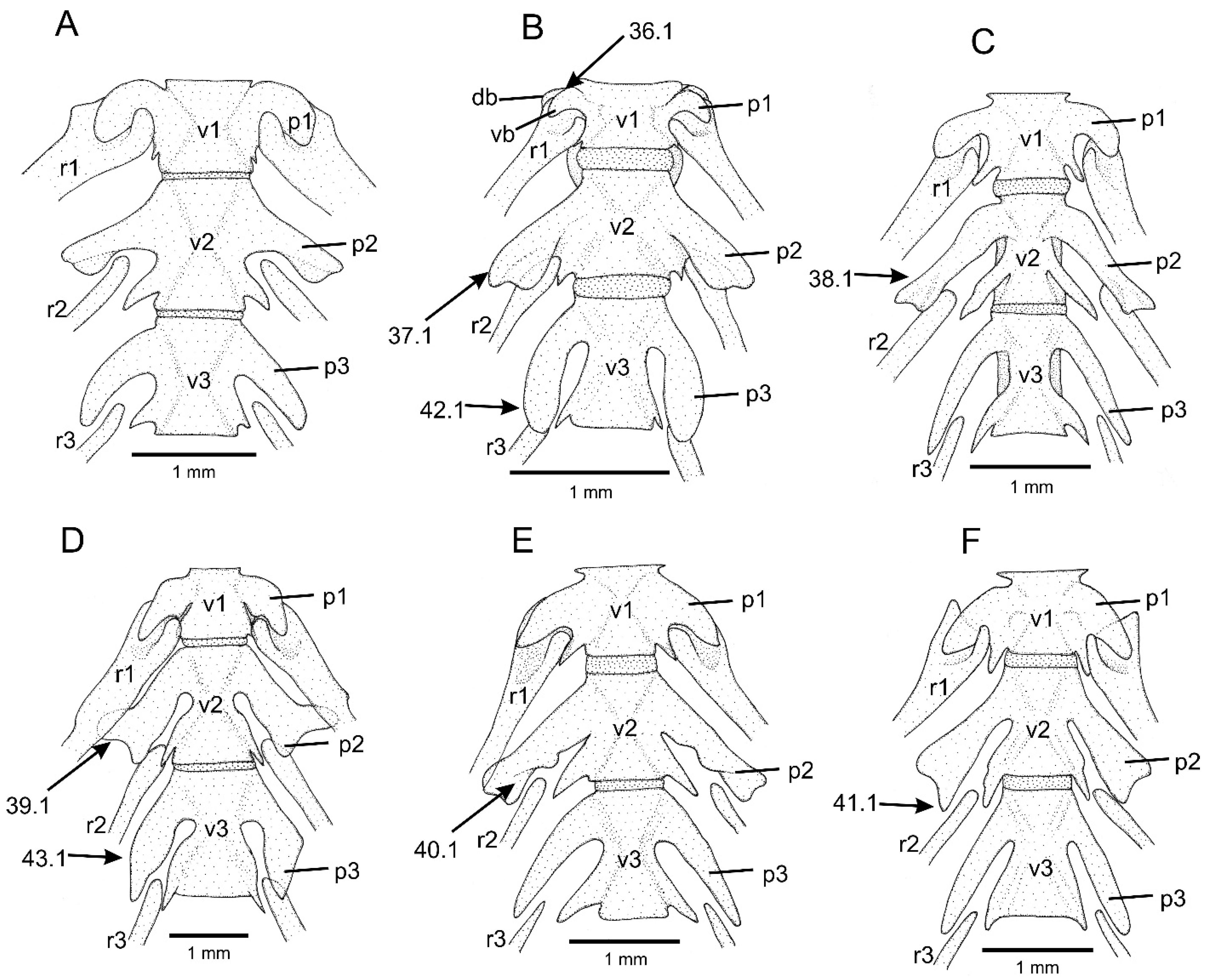
- Autopalatine, dorsal surface, lateral portion, long longitudinal ridge: (0) absent; (1) present.
- Autopalatine, dorsal surface, latero-middle portion, short longitudinal ridge: (0) absent; (1) present.
- Autopalatine, postero-lateral process, development: (0) well-developed; (1) rudimentary.
- Autopalatine, postero-lateral process, direction: (0) postero-lateral; (1) posterior; (2) lateral.
- Autopalatine, postero-lateral process, length relative to autopalatine longitudinal length excluding postero-lateral process: (0) distinctively shorter; (1) nearly equal or slightly longer.
- Autopalatine in specimens above 45 mm SL, medial margin, shape: (0) concave, sometimes nearly straight; (1) convex; (2) with deep concavity, posteriorly followed by broad convexity.
- Autopalatine, articular facet for lateral ethmoid, ventral flap, shape: (0) small or rudimentary; (1) long.
- Autopalatine, dorsal surface, articular facet for the lateral ethmoid, prominent ridge terminating in process connected to sesamoid supraorbital: (0) absent; (1) present.
- Autopalatine, dorsal surface, articular facet for the lateral ethmoid, robust comma-shaped support: (0) absent; (1) present.
- Metapterygoid, posterior portion, expansion: (0) absent; (1) present.
- Metapterygoid, postero-ventral margin, deep concavity accommodating dorsal expansion of the antero-dorsal process of the quadrate: (0) absent; (1) present.
- Hyomandibula, anterior-outgrowth, shape: (0) deep; (1) shallow.
- Hyomandibula, anterior-outgrowth, dorsal margin, shape: (0) slightly curved, sometimes with weak dorsal concavity; (1) nearly straight and horizontal, with deep U-shaped concavity; (2) step anterior and posterior portions converging to pronounced V-shaped concavity reaching central outgrowth; (3) deep concavity reaching ventral outgrowth.
- Opercle, anterior odontodes, arrangement: (0) transversal; (1) diagonal.
- Opercle and interopercle, largest and internal-most odontodes, distal portion, shape: (0) straight to slightly curved; (1) strongly curved, its tip forming an angle of about 90° with odontode main axis.
- Interopercle, anterior portion, distinctive anterior expansion: (0) absent; (1) present.
- Interopercle, dorsal process, position relative to anterior margin of odontode patch: (0) distant; (1) close.
- Interopercle, dorsal process, anterior margin, shape: (0) slightly concave; (1) prominent V-shaped notch.
- Preopercle, ventral flap, extent relative to ventral bone limit: (0) slightly extending beyond it; (1) expanding ventrally beyond it.
- Maxilla, largest length relative premaxilla largest length: (0) shorter; (1) longer.
- Maxilla, shape: (0) slightly curved; (1) somewhat folded; (2) nearly straight.
- Maxilla, postero-middle process, shape: (0) moderate, slightly projecting beyond bone profile; (1) large, forming pronounced expansion.
- Antorbital, association with anterior segment of the latero-sensory infraorbital canal: (0) associated; (1) separated.
- Sesamoid supraorbital, distinctive robust lateral process: (0) absent; (1) present.
- Mesethmoid, lateral margin, ventral fold: (0) absent; (1) present.
- Lateral ethmoid, articular facet for the autopalatine, folded laminar expansion: (0) absent; (1) present.
- Lateral ethmoid, antero-dorsal extremity, direction: (0) anterior; (1) lateral; (2) medial.
- Lateral ethmoid, medial margin, proximity relative to its paired homologous: (0) in contact; (1) separate.
- Lateral ethmoid, lateral margin, area close to sesamoid supraorbital, lateral projection: (0) absent; (1) present.
- Lateral ethmoid, lateral margin, prominent trapezoidal process: (0) absent; (1) present.
- Lateral ethmoid, lateral margin, widening to form distinct wall limiting fossa aperture: (0) absent; (1) present.
- Sphenotic-prootic-pterosphenoid complex, lateral process, extent relative to main neurocranium body limit: (0) short, slightly extending beyond it; (1) long, strongly projecting beyond it.
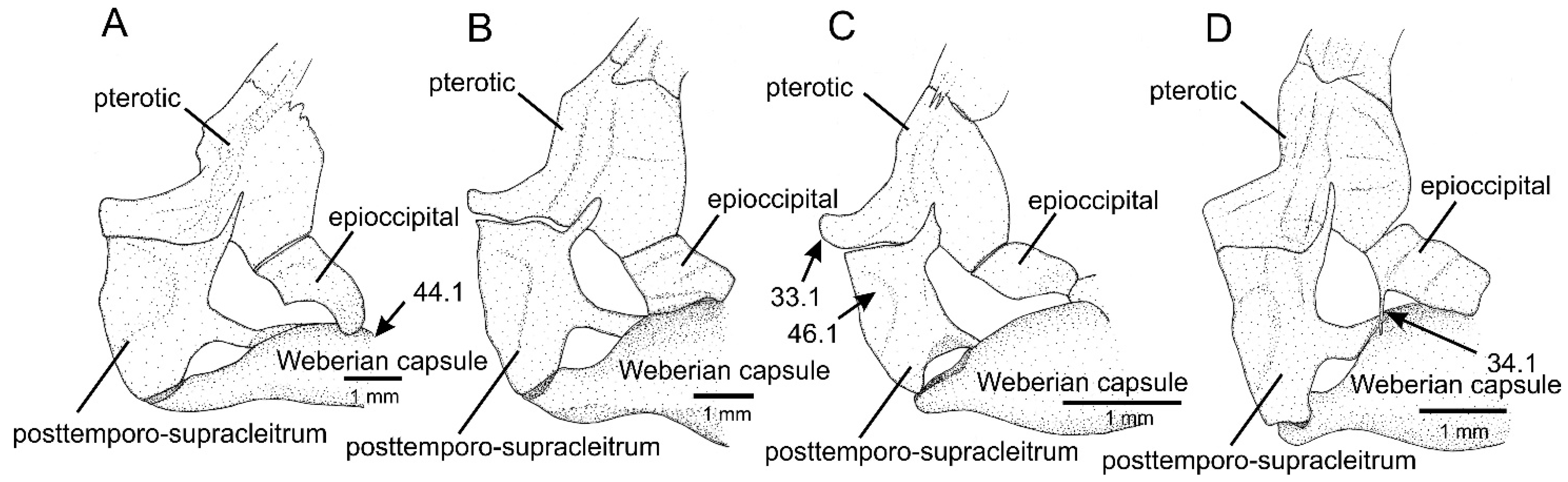
- Pterotic, lateral process, extremity, shape and extent relative to limit of lateral neighbouring bones: (0) truncate, not projecting beyond; (1) rounded, projecting beyond.
- Epioccipital, posterior margin, sharp process posteriorly directed: (0) absent; (1) present.
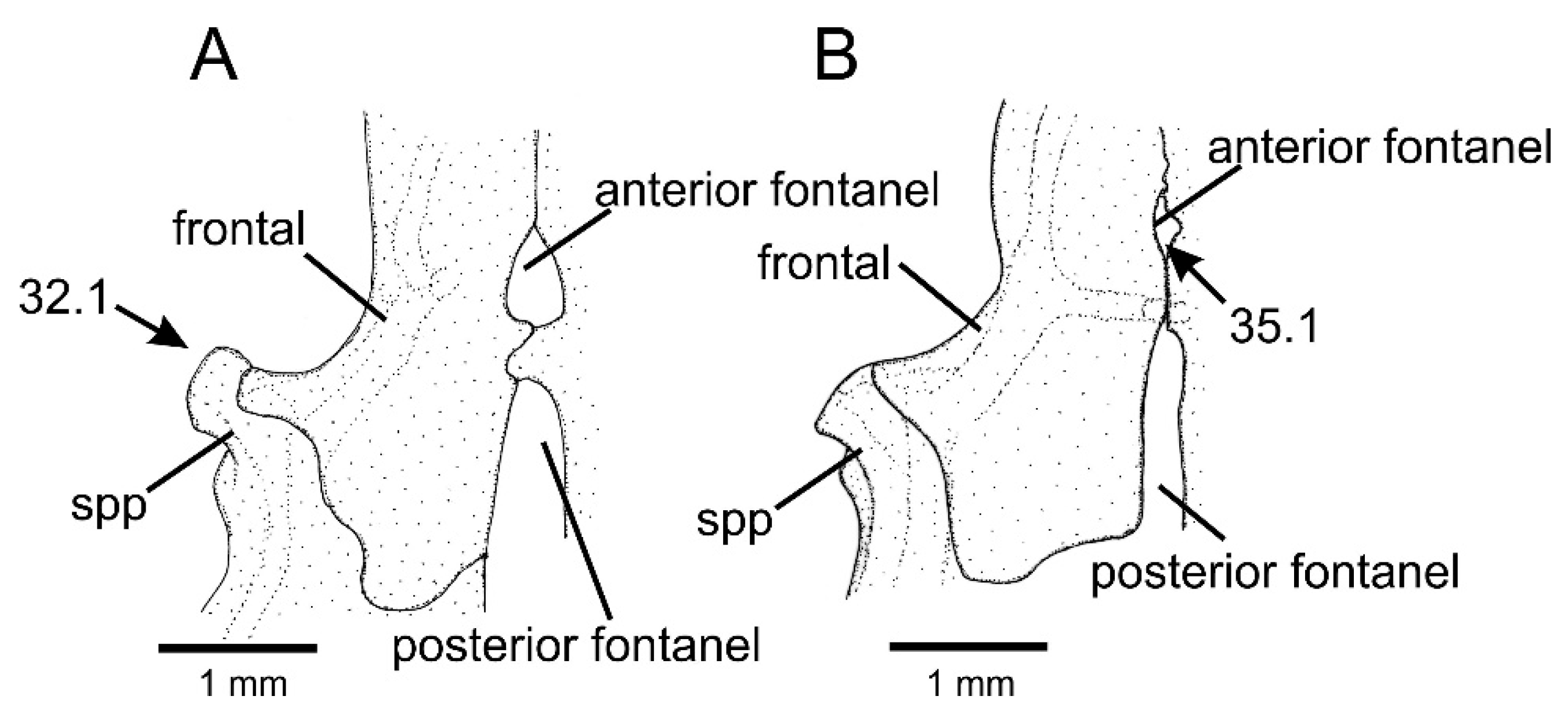
- Frontal, anterior cranial fontanel, shape: (0) well-developed orifice; (1) minute aperture.
- First free vertebra, parapophysis, branching: (0) unbranched; (1) branched.
- Second free vertebra, parapophysis, direction: (0) lateral; (1) posterolateral.
- Second free vertebra, parapophysis, general shape: (0) robust; (1) slender.
- Second free vertebra, parapophysis, antero-distal portion, expansion: (0) absent; (1) present.
- Second free vertebra, parapophysis, posterior margin, small notch: (0) absent; (1) present.
- Second free vertebra, parapophysis, postero-distal portion, expansion: (0) absent; (1) present.
- Third free vertebra, parapophysis, direction: (0) postero-lateral; (1) posterior.
- Third free vertebra, parapophysis, shape: (0) nearly straight; (1) posteriorly curved.
- Weberian capsule, shape: (0) pear-like, abruptly narrowing laterally; (1) slender, subtly narrowing laterally.
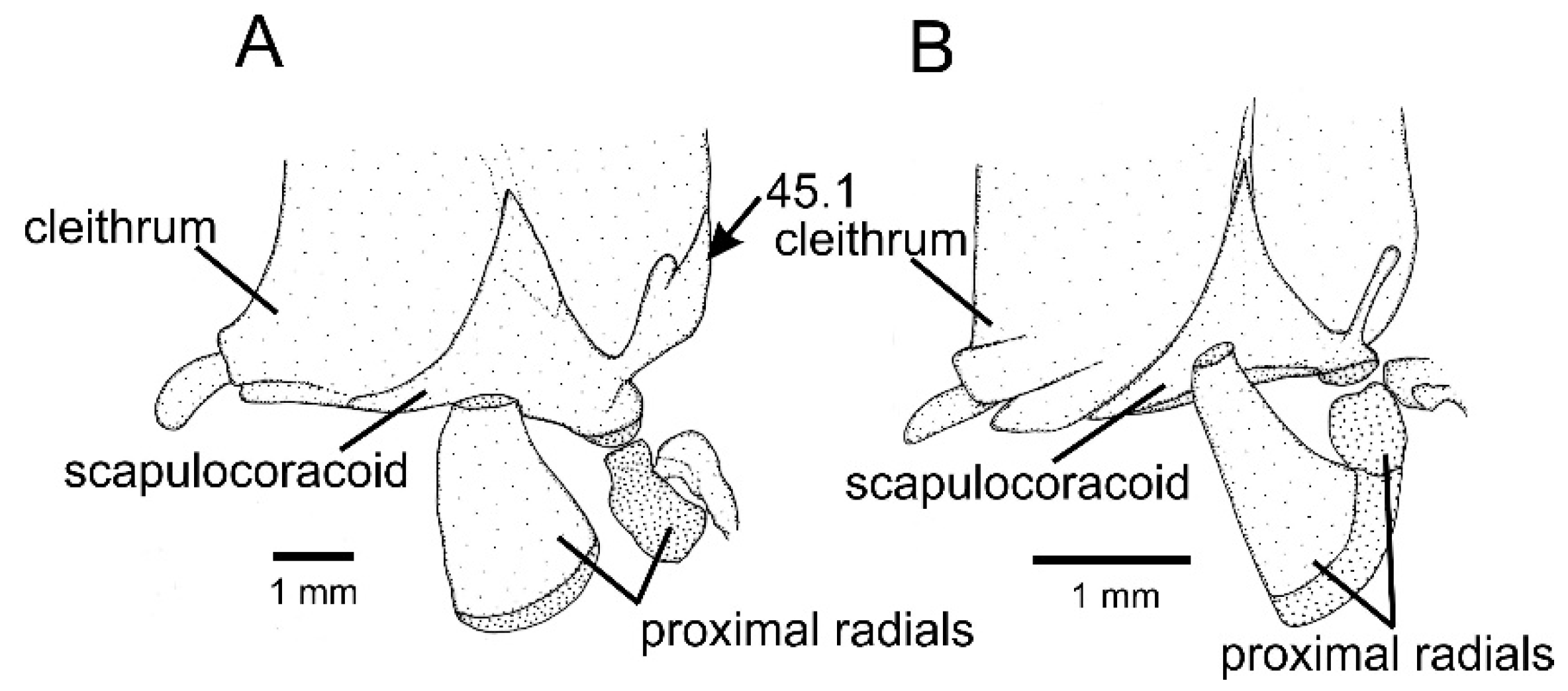
- Scapulo-coracoid, supra-lateral process, shape: (0) needle-shaped; (1) tree-shaped.
- Posttemporo-supracleithrum, size and space around it: (0) large, occupying most space among neighbouring bones; (1) small, surrounded by large space separating from neighbouring bones.
- Pelvic bone, development: (0) well-developed; (1) poorly-developed.
- Pelvic bone, general shape: (0) compact; (1) elongated.
- Pelvic bone, posterior process, development: (0) well-developed; (1) rudimentary. Remarks: taxa with poorly developed pelvic bone or lacking a pelvic bone were coded as (?).
- Pelvic bone, anterior processes, basal region: (0) broad; (1) narrow. Remarks: taxa lacking a pelvic bone were coded as (?).
- Pelvic bone, anterior processes, number: (0) two; (1) three. Remarks: taxa lacking a pelvic bone were coded as (?).
- Pelvic bone, postero-medial portion, small process close to cartilage supporting fin rays: (0) absent; (1) present.
Appendix D
| Taxon | Characters 1–52 |
|---|---|
| Nematogenys inermis | 00???0?0??0?????0??0?00?00000???0?0?000000000?000000 |
| Copionodon pecten | 00???0?0??0?????00?0?00?00000???0000000000000?000000 |
| Trichogenes longipinnis | 00???0?0??0?????00?0?00?00000???0000000000000?00000? |
| Microcambeva ribeirae | 000000?00?????0?0?0100000000000000?0100000000?000000 |
| Listrura tetraradiata | 001??0?00?????0?0?0100000000000000?0100000000??????? |
| Eremophilus mutisii | 00000000000000000000000000001?000000000000000??????? |
| Trichomycterus punctulatus | 00000000000000000000000000000000?0000000000000000000 |
| Bullockia maldonadoi | 0000100000000000001100?00000000000000000000000000000 |
| Trichomycterus areolatus | 0000100000000000001000000000000000000000000000000000 |
| Ituglanis boitata | 0000000000000000000000000000000000?00000001000000000 |
| Scleronema minutum | 0000000000000000100100?00000000000011000010000000000 |
| Scleronema cf guapa | 0000000000000000100100?00000000000011000010000000000 |
| Scleronema cf ibirapuita | 0000000000000000100100?00000000000011000010000000000 |
| Scleronema macanuda | 0000000000000000100100?00000000000011000010000000000 |
| Cambeva variegata | 0000000000000000100000?00000000000011000010000000000 |
| Cambeva brachychenos | 0000000000000000100000?00000000000011000010000000000 |
| Cambeva diatropoporos | 001?000000000000100000000000000000011000010000000000 |
| Cambeva cubataonis | 0000000000000000100000?000000000?0011000010000000000 |
| Cambeva davisi | 0000000000000000100000?000000000?0011000010000000000 |
| Cambeva castroi | 0000000000000000100000?000000000?0011000010000000000 |
| Cambeva zonata | 0000000000000000100000?000000000?0011000010000000000 |
| Trichomycterus giganteus | 0101000100003010000000000000010000001000000110000010 |
| Trichomycterus itatiayae | 001?000000101000100000?00020000000001100000000000001 |
| Trichomycterus septemradiatus | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus pauciradiatus | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus piratymbara | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus reinhardti | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus anaisae | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus funebris | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus humboldti | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus luetkeni | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus ingaiensis | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus sainthilairei | 000000100000000010000010000000000000110000000010?000 |
| Trichomycterus nigricans | 0000000001000000000101000010000100001010001000000000 |
| Trichomycterus caipora | 0000000001000000000101000010000100001010001000000000 |
| Trichomycterus santaeritae | 0000000001000000000101000010000100001000001000000000 |
| Trichomycterus immaculatus | 0000000001000000000001000010000100001010001000000000 |
| Trichomycterus nigroauratus | 0000000001000000000000000110000100001010001000010000 |
| Trichomycterus maculosus | 0000000001000000000000000110000100001010001000010000 |
| Trichomycterus quintus | 0000000001000000000000000110000100001010000000010000 |
| Trichomycterus mimonha | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus fuliginosus | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus brunoi | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus mirissumba | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus pirabitira | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus macrotrichopterus | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus brasiliensis | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus candidus | 100000000000010000000000100110001000100100000100???? |
| Trichomycterus potschi | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus claudiae | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus novalimensis | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus rubiginosus | 1000000000000100000000001001100010001001000001001100 |
| Trichomycterus albinotatus | 1000010000002100000000001001100001001001000000001000 |
| Trichomycterus vitalbrazili | 1000010000002100000000001001100001001001000000001000 |
| Trichomycterus mimosensis | 0000000000000000000100000000001000101000000000000000 |
| Trichomycterus travassosi | 0000000000000000000100000000001000101000000000000000 |
| Trichomycterus caudofasciatus | 0000000000000000000100000000001000101000000000000000 |
| Trichomycterus cf jacupiranga | 0000000000000000000100010000001000101000000000000000 |
| Trichomycterus pantherinus | 0000000000000000000100010000001000101000000000000000 |
| Trichomycterus longibarbatus | 0002000000000000010120000000001000001000000000000000 |
| Trichomycterus auroguttatus | 0000120010000000010110000000001000101000100000000000 |
| Trichomycterus alternatus | 0000100000000000010110000000001000101000100000000000 |
| Trichomycterus melanopygus | 0000000000010000001100000000000000001000000000000000 |
| Trichomycterus pradensis | 0000000000010001001100000000000000001000000000000000 |
| Trichomycterus tete | 0000000000010000001100000000000000001000000000000000 |
Appendix E
| Partition | Base Pairs | Evolutionary Model |
|---|---|---|
| COI 1st | 174 | TNe+G4 |
| COI 2nd | 174 | F81+F+I+G4 |
| COI 3rd | 174 | TIM+F+G4 |
| CYTB 1st | 363 | K2P+I+G4 |
| CYTB 2nd | 363 | TN+F+I |
| CYTB 3rd | 362 | TIM2+F+I+G4 |
| MYH6 1st | 181 | JC+I+G4 |
| MYH6 2nd | 181 | HKY+F+I+G4 |
| MYH6 3rd | 181 | K2P+G4 |
| RAG2 1st | 274 | K3P+I |
| RAG2 2nd | 274 | TVMe+G4 |
| RAG2 3rd | 273 | K2P+G4 |
| Osteology | 52 | JC+I+G4 |
References
- Vari, R.P.; Malabarba, L.R. Neotropical ichthyology: An overview. In Phylogeny and Classification of Neotropical Fishes; Malabarba, L.R., Reis, R.E., Vari, R.P., Lucena, Z.M.S., Lucena, C.A.S., Eds.; Edipucrs: Porto Alegre, Brazil, 1998; pp. 1–11. [Google Scholar]
- Reis, R.E.; Kullander, S.O.; Ferraris, C.J., Jr. (Eds.) Check List of the Freshwater Fishes of South and Central America; Edipucrs: Porto Alegre, Brazil, 2003. [Google Scholar]
- Fernández, L.; Schaefer, S.A. Relationships among the Neotropical candirus (Trichomycteridae, Siluriformes) and the evolution of parasitism based on analysis of mitochondrial and nuclear gene sequences. Mol. Phylogenet. Evol. 2009, 52, 416–423. [Google Scholar] [CrossRef] [PubMed]
- Myers, G.S. Two extraordinary new blind nematognath fishes from the Rio Negro, representing a new subfamily of Pygidiidae, with a rearrangement of the genera of the family, and illustrations of some previously described genera and species from Venezuela and Brazil. Calif. Acad. Sci. 1944, 4, 591–602. [Google Scholar]
- Myers, G.S.; Weitzman, S.H. Two remarkable new trichomycterid catfishes from the Amazon basin in Brazil and Colombia. J. Zool. 1966, 149, 277–287. [Google Scholar] [CrossRef]
- Schaefer, S.A.; Provenzano, F.; de Pinna, M.; Baskin, J.N. New and noteworthy Venezuelan glanapterygine catfishes (Siluriformes, Trichomycteridae), with discussion of their biogeography and psammophily. Am. Mus. Novit. 2005, 3496, 1–27. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Henschel, E.; Katz, A.M. Multigene phylogeny reveals convergent evolution in small interstitial catfishes from the Amazon and Atlantic forests (Siluriformes: Trichomycteridae). Zool. Scr. 2020, 49, 159–173. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 11 June 2021).
- Katz, A.M.; Barbosa, M.A.; Mattos, J.L.O.; Costa, W.J.E.M. Multigene analysis of the catfish genus Trichomycterus and description of a new South American trichomycterine genus (Siluriformes, Trichomycteridae). Zoosyst. Evol. 2018, 94, 557–566. [Google Scholar] [CrossRef]
- Eigenmann, C.H. The Pygidiidae, a subfamily of South American catfishes. Mem. Carnegie Mus. 1918, 7, 259–398. [Google Scholar]
- De Pinna, M.C.C.; Wosiacki, W. Trichomycteridae. In Check List of the Freshwater Fishes of South and Central America; Reis, R.E., Kullander, S.O., Ferraris, C.J., Jr., Eds.; Edipucrs: Porto Alegre, Brazil, 2003. [Google Scholar]
- Arratia, G.; Chang, A.; Menu-Marque, S.; Rojas, G. About Bullockia gen. nov., Trichomycterus mendozensis n. sp. and revision of the family Trichomycteridae (Pisces, Siluriformes). Stud. Neotrop. Fauna Environ. 1978, 13, 157–194. [Google Scholar] [CrossRef]
- Costa, W.J.E.M. Description de huit nouvelles espèces du genre Trichomycterus (Siluriformes: Trichomycteridae), du Brésil oriental. Rev. Fr. D’aquariol. 1992, 18, 101–110. [Google Scholar]
- Costa, W.J.E.M.; Bockmann, F.A. Un nouveau genre néotropical de la famille des Trichomycteridae (Siluriformes: Loricarioidei). Rev. Fr. D’aquariol. 1993, 20, 43–46. [Google Scholar]
- Fernández, L.; de Pinna, M.C.C. Phreatic Catfish of the Genus Silvinichthys from southern South America (Teleostei, Siluriformes, Trichomycteridae). Copeia 2005, 2005, 100–108. [Google Scholar] [CrossRef]
- Bichuette, M.E.; Rizzato, P.P. A new species of cave catfish from Brazil, Trichomycterus rubbioli sp. n., from Serra do Ramalho karstic area, São Francisco River basin, Bahia State (Silurifomes: Trichomycteridae). Zootaxa 2012, 3480, 48–66. [Google Scholar] [CrossRef]
- Rizzato, P.P.; Bichuette, M.E. Ituglanis boticario, a new troglomorphic catfish (Teleostei: Siluriformes: Trichomycteridae) from Mambaí karst area, central Brazil. Zoologia 2014, 31, 577–598. [Google Scholar] [CrossRef]
- De Pinna, M.C.C. Phylogenetic relationships of neotropical Siluriformes (Teleostei: Ostariophysi): Historical overview and synthesis of hypotheses. In Phylogeny and Classification of Neotropical Fishes; Malabarba, L.R., Reis, R.E., Vari, R.P., Lucena, Z.M.S., Lucena, C.A.S., Eds.; Edipucrs: Porto Alegre, Brazil, 1998; pp. 279–330. [Google Scholar]
- Arratia, G. Silvinichthys, a new genus of trichomycterid catfishes from the Argentinian Andes, with redescription of Trichomycterus nigricans. Ichthyol. Explor. Freshw. 1998, 9, 347–370. [Google Scholar]
- Bockmann, F.A.; Casatti, L.; de Pinna, M.C.C. A new species of trichomycterid catfish from the Rio Paranapanema, southeastern Brazil (Teleostei; Siluriformes), with comments on the phylogeny of the family. Ichthyol. Explor. Freshw. 2004, 15, 225–242. [Google Scholar]
- Ferrer, J.; Malabarba, L.R. Taxonomic review of the genus Trichomycterus Valenciennes (Siluriformes: Trichomycteridae) from the laguna dos Patos system, Southern Brazil. Neotrop. Ichthyol. 2013, 11, 217–246. [Google Scholar] [CrossRef]
- Ochoa, L.E.; Roxo, F.F.; DoNascimiento, C.; Sabaj, M.H.; Datovo, A.; Alfaro, M.; Oliveira, C. Multilocus analysis of the catfish family Trichomycteridae (Teleostei: Ostariophysi: Siluriformes) supporting a monophyletic Trichomycterinae. Mol. Phylogenet. Evol. 2017, 115, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Costa, W.J.E.M.; Feltrin, C.R.M.; Katz, A.M. Filling distribution gaps: Two new species of the catfish genus Cambeva from southern Brazilian Atlantic Forest (Siluriformes: Trichomycteridae). Zoosyst. Evol. 2021, 97, 147–159. [Google Scholar] [CrossRef]
- Ferrer, J.; Malabarba, L.R. Systematic revision of the Neotropical catfish genus Scleronema (Siluriformes: Trichomycteridae), with descriptions of six new species from Pampa grasslands. Neotrop. Ichthyol. 2020, 18, e190081. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Katz, A.M. Integrative taxonomy supports high species diversity of south-eastern Brazilian mountain catfishes of the T. reinhardti group (Siluriformes: Trichomycteridae). Syst. Biodivers. 2021. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Costa, W.J.E.M. Description of a new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from the rio de Contas basin, northeastern Brazil. Vertebr. Zool. 2011, 61, 307–312. [Google Scholar]
- Donin, L.M.; Ferrer, J.; Carvalho, T.P. Taxonomical study of Trichomycterus (Siluriformes: Trichomycteridae) from the Ribeira de Iguape River basin reveals a new species recorded in the early 20th century. J. Fish Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.A.; Costa, W.J.E.M. Trichomycterus potschi (Siluriformes: Loricarioidei): A new trichomycterid catfish from coastal streams of southeastern Brazil. Ichthyol. Explor. Freshw. 2003, 14, 281–287. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Seven new species of the catfish genus Trichomycterus (Teleostei: Siluriformes: Trichomycteridae) from southeastern Brazil and redescription of T. brasiliensis. Ichthyol. Explor. Freshw. 2010, 21, 97–122. [Google Scholar]
- Bockmann, F.A.; Sazima, I. Trichomycterus maracaya, a new catfish from the upper rio Paraná, southeastern Brazil (Siluriformes: Trichomycteridae), with notes on the T. brasiliensis species-complex. Neotrop. Ichthyol. 2004, 2, 61–74. [Google Scholar] [CrossRef]
- Ochoa, L.E.; Datovo, A.; DoNascimiento, C.; Roxo, F.F.; Sabaj, M.H.; Chang, J.; Melo, B.F.; Silva Gabriel, S.C.; Foresti, F.; Alfaro, M.; et al. Phylogenomic analysis of trichomycterid catfshes (Teleostei: Siluriformes) inferred from ultraconserved elements. Sci. Rep. 2020, 10, 2697. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Mattos, J.L.O.; Amorim, P.F.; Vilardo, P.J.; Katz, A.M. Relationships of a new species support multiple origin of melanism in Trichomycterus from the Atlantic Forest of south-eastern Brazil (Siluriformes: Trichomycteridae). Zool. Anz. 2020, 288, 74–83. [Google Scholar] [CrossRef]
- Vilardo, P.J.; Katz, A.M.; Costa, W.J.E.M. Relationships and description of a new species of Trichomycterus (Siluriformes: Trichomycteridae) from the Rio Paraíba do Sul basin, south-eastern Brazil. Zool. Stud. 2020, 59, e53. [Google Scholar] [CrossRef]
- Costa, W.J.E.M.; Mattos, J.L.O.; Katz, A.M. Phylogenetic position of Trichomycterus payaya and examination of osteological characters diagnosing the Neotropical catfish genus Ituglanis (Siluriformes: Trichomycteridae). Zool. Stud. 2021, in press. [Google Scholar]
- Barbosa, M.A.; Costa, W.J.E.M. Trichomycterus macrophthalmus (Teleostei: Siluriformes: Trichomycteridae), a new species of catfish from the rio Paraiba do Sul basin, southeastern Brazil. Vertebr. Zool. 2012, 62, 79–82. [Google Scholar]
- Costa, W.J.E.M.; Katz, A.M.; Mattos, J.L.O.; Amorim, P.F.; Mesquita, B.O.; Vilardo, P.J.; Barbosa, M.A. Historical review and redescription of three poorly known species of the catfish genus Trichomycterus from south-eastern Brazil (Siluriformes: Trichomycteridae). J. Nat. Hist. 2020, 53, 2905–2928. [Google Scholar] [CrossRef]
- Taylor, W.R.; Van Dyke, G.C. Revised procedures for staining and clearing small fishes and other vertebrates for bone and cartilage study. Cybium 1985, 9, 107–119. [Google Scholar]
- Baskin, J.N. Structure and Relationships of the Trichomycteridae; City University of New York: New York, NY, USA, 1973; Volume 14, pp. S1–S62. [Google Scholar]
- Arratia, G.; Menu-Marque, S. Revision of the freshwater catfishes of the genus Hatcheria (Siluriformes, Trichomycteridae) with commentaries on ecology and biogeography. Zool. Anz. 1981, 207, 88–111. [Google Scholar]
- Arratia, G. Trichomycterus chungaraensis n. sp. and Trichomycterus laucaensis n. sp. (Pisces, Siluriformes, Trichomycteridae) from the high Andean range. Stud. Neotrop. Fauna Environ. 1983, 18, 65–87. [Google Scholar] [CrossRef]
- Arratia, G. Development and diversity of the suspensorium of trichomycterids and comparison with loricarioids (Teleostei: Siluriformes). J. Morphol. 1990, 205, 193–218. [Google Scholar] [CrossRef]
- Schaefer, S.A.; Fernández, L. Redescription of the Pez Graso, Rhizosomichthys totae (Trichomycteridae) of Lago Tota, Colombia, and aspects of cranial osteology revealed by microtomography. Copeia 2009, 2009, 510–522. [Google Scholar] [CrossRef]
- Carvalho, M.; Bockmann, F.A.; de Carvalho, M.R. Homology of the fifth epibranchial and accessory elements of the ceratobranchials among Gnathostomes: Insights from the development of Ostariophysans. PLoS ONE 2013, 8, e62389. [Google Scholar] [CrossRef]
- Arratia, G.; Schultze, H.-P. The urohyal: Development and homology within Osteichthyans. J. Morphol. 1990, 203, 247–282. [Google Scholar] [CrossRef]
- Britz, R.; Bartsch, P. The myth of dorsal ribs in gnathostome vertebrates. Proc. R. Soc. Lond. Biol. Sci. Ser. B 2003, 270, S1–S4. [Google Scholar] [CrossRef]
- Adriaens, D.; Baskin, J.N.; Coppens, H. Evolutionary morphology of trichomycterid catfishes: About hanging on and digging in. In Origin and Phylogenetic Interrelationships of Teleosts; Nelson, J.S., Schultze, H.P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2010; pp. 337–362. [Google Scholar]
- De Pinna, M.; Reis, V.; Britski, H. A new species of Trichogenes (Siluriformes, Trichomycteridae), with a discussion on the homologies of the anterior orbital bones in trichomycterids and other loricarioids. Am. Mus. Novit. 2020, 3951, 1–27. [Google Scholar] [CrossRef]
- Arratia, G.; Huaquin, L. Morphology of the lateral line system and of the skin of diplomystic and certain primitive loricarioid catfishes and systematic and ecological considerations. Bonn Zool. Monogr. 1995, 36, 1–110. [Google Scholar]
- Arratia, G. Catfish head skeleton, an overview. In Catfishes; Arratia, G., Kapoor, B.G., Chardon, M., Diogo, R., Eds.; Science Publishers: Enfield, UK; Plymouth, UK, 2003; pp. 3–46. [Google Scholar]
- Britski, H.A.; Ortega, H. Trichogenes longipinnis, novo gênero e espéciede Trichomycterinae do sudeste do Brazil (Pisces, Siluriformes). Rev. Bras. Zool. 1983, 1, 211–216. [Google Scholar] [CrossRef]
- Datovo, A.; Bockmann, F.A. Dorsolateral head muscles of the catfish families Nematogenyidae and Trichomycteridae (Siluriformes: Loricarioidei): Comparative anatomy and phylogenetic analysis. Neotrop. Ichthyol. 2010, 8, 193–246. [Google Scholar] [CrossRef]
- Hardman, M.; Page, L.M. Phylogenetic relationships among bullhead catfishes of the genus Ameiurus (Siluriformes: Ictaluridae). Copeia 2003, 2003, 20–33. [Google Scholar] [CrossRef]
- Cramer, C.A.; Bonatto, S.L.; Reis, R.E. Molecular phylogeny of the Neoplecostominae and Hypoptopomatinae (Siluriformes: Loricariidae) using multiple genes. Mol. Phylogenet. Evol. 2011, 59, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ortí, G.; Zhang, G.; Lu, G. A practical approach to phylogenomics: The phylogeny of ray-finned fish (Actinopterygii) as a case study. BMC Evol. Biol. 2007, 7, 44. [Google Scholar] [CrossRef]
- Villa-Verde, L.; Lazzarotto, H.; Lima, S.Q.M. A new glanapterygine catfish of the genus Listrura (Siluriformes: Trichomycteridae) from southeastern Brazil, corroborated by morphological and molecular data. Neotrop. Ichthyol. 2012, 10, 527–538. [Google Scholar] [CrossRef]
- Ward, R.D.; Zemlak, T.S.; Innes, B.H.; Last, P.R.; Hebert, P.D. DNA barcoding Australia’s fish species. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2005, 360, 1847–1857. [Google Scholar] [CrossRef]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7, Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Sereno, P.C. Logical basis for morphological characters in phylogenetics. Cladistics 2007, 23, 565–587. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Anisimova, M.; Gil, M.; Dufayard, J.F.; Dessimoz, C.; Gascuel, O. Survey of branch support methods demonstrates accuracy, power, and robustness of fast likelihood-based approximation schemes. Syst. Biol. 2011, 60, 685–699. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2, Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Costa, W.J.E.M. Description of a new species of catfish from the upper rio Paraíba do Sul basin, south-eastern Brazil (Teleostei: Siluriformes: Trichomycteridae) and re-description of Trichomycterus itatiayae. Aqua Int. J. Ichthyol. 2008, 14, 175–186. [Google Scholar]
- Reis, V.J.C.; de Pinna, M.C.C.; Pessali, T.C. A new species of Trichomycterus Valenciennes, 1832 (Trichomycteridae: Siluriformes) from the Rio Doce drainage with remarkable similarities with Bullockia and a CT-scan survey. J. Fish Biol. 2020, 95, 918–931. [Google Scholar] [CrossRef]
- Lezama, A.Q.; Triques, M.L.; Santos, P.S. Trichomycterus argos (Teleostei: Siluriformes: Trichomycteridae), a new species from the Doce River Basin, Eastern Brazil. Zootaxa 2012, 3352, 60–68. [Google Scholar] [CrossRef]
- De Pinna, M.C.C. A new subfamily of Trichomycteridae (Teleostei, Siluriformes), lower loricarioid relationships and a discussion on the impact of additional taxa for phylogenetic analysis. Zool. J. Linn. Soc. 1992, 106, 175–229. [Google Scholar] [CrossRef]
- Lima, S.M.Q.; Costa, W.J.E.M. Trichomycterus giganteus (Siluriformes: Loricarioidea: Trichomycteridae): A new catfish from the Rio Guandu basin, southeastern Brazil. Zootaxa 2004, 761, 1–6. [Google Scholar] [CrossRef]
- Lima, S.M.Q.; Lazzarotto, H.; Costa, W.J.E.M. A new species of Trichomycterus (Siluriformes: Trichomycteridae) from lagoa Feia drainage, southeastern Brazil. Neotrop. Ichthyol. 2008, 6, 315–322. [Google Scholar] [CrossRef]
- Alencar, A.R.; Costa, W.J.E.M. Description of two new species of the catfish genus Trichomycterus from southeastern Brazil (Siluriformes: Trichomycteridae). Zootaxa 2004, 744, 1–8. [Google Scholar] [CrossRef]
- Barbosa, M.A.; Costa, W.J.E.M. Trichomycterus puriventris (Teleostei: Siluriformes: Trichomycteridae), a new species of catfish from the rio Paraiba do Sul basin, southeastern Brazil. Vertebr. Zool. 2012, 62, 155–160. [Google Scholar]
- Datovo, A.; de Pinna, M.C.C. A new species of Ituglanis representing the southernmost record of the genus, with comments on phylogenetic relationships (Teleostei: Siluriformes: Trichomycteridae). J. Fish Biol. 2014, 84, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Arroyave, J.; Schaefer, S.A. Emerging patterns in phylogenetic studies of trichomycterid catfishes (Teleostei, Siluriformes) and the contribution of Andean diversity. Zool. Scr. 2021. [Google Scholar] [CrossRef]









| Node | ML | MP | ||
|---|---|---|---|---|
| SH-aLRT | aBayes | UFBoot | Bootstrap | |
| 1 | - | - | - | 100 |
| 2 | 97.4 | 1 | 100 | 97 |
| 3 | 100 | 1 | 100 | 99 |
| 4 (Microcambevinae) | 100 | 1 | 100 | 99 |
| 5 (Trichomycterinae) | 98.4 | 1 | 100 | 98 |
| 6 | 84.5 | 1 | 74 | - |
| 7 | 85.8 | 1 | 76 | - |
| 8 | 99.6 | 1 | 100 | 78 |
| 9 | 100 | 1 | 100 | 100 |
| 10 (CST-clade) | 99.8 | 1 | 100 | 95 |
| 11 (Cambeva + Scleronema) | 99.9 | 1 | 100 | 99 |
| 12 (Scleronema) | 100 | 1 | 100 | 100 |
| 13 | 99.9 | 1 | 100 | 100 |
| 14 | 98.6 | 1 | 100 | 100 |
| 15 (Cambeva) | 89.0 | 1 | 95 | 84 |
| 16 | 66.7 | - | 79 | - |
| 17 | 96.6 | 1 | 95 | - |
| 18 | 99.1 | 1 | 100 | 83 |
| 19 | 99.4 | 1 | 100 | 100 |
| 20 | 73.3 | 0.8 | 92 | - |
| 21 (Trichomycterus) | 85.1 | 1 | 92 | 89 |
| 22 | 78.6 | 0.8 | 82 | - |
| 23 (Paracambeva) | 100 | 1 | 100 | 100 |
| 24 (T. reinhardti group) | 97.3 | 1 | 99 | 99 |
| 25 | 98.8 | 1 | 100 | 97 |
| 26 | 96.9 | 1 | 97 | 92 |
| 27 | - | 1 | - | - |
| 28 | 100 | 1 | 100 | 100 |
| 29 | 100 | 1 | 100 | 99 |
| 30 | 83.7 | 0.9 | 97 | 99 |
| 31 | 83.1 | 1 | 91 | - |
| 32 (sg. Trichomycterus) | 100 | 1 | 100 | 100 |
| 33 (T. nigroauratus group) | 100 | 1 | 100 | 100 |
| 34 (T. nigricans group) | 86.2 | 1 | 96 | 72 |
| 35 | 98.3 | 1 | 100 | 94 |
| 36 | 97.6 | 1 | 100 | 100 |
| 37 | 91 | 1 | 93 | - |
| 38 (Psammocambeva) | 98.9 | 1 | 100 | 89 |
| 39 (T. pradensis group) | 50.6 | 0.7 | 80 | - |
| 40 | 87.1 | 1 | 96 | - |
| 41 (T. travassosi group) | 100 | 1 | 100 | 94 |
| 42 | - | - | - | - |
| 43 | 92.3 | 1 | 98 | - |
| 44 | 85.9 | 1 | 87 | - |
| 45 | 92.1 | 1 | 97 | - |
| 46 | - | - | 85 | - |
| 47 | 99.2 | 1 | 100 | 95 |
| 48 (Humboldtglanis) | 99.4 | 1 | 100 | 100 |
| 49 (Cryptocambeva) | 100 | 1 | 100 | 100 |
| 50 | 92.4 | 1 | 99 | 55 |
| 51 | 96.2 | 1 | 99 | 58 |
| 52 | 96 | 1 | 99 | 92 |
| 53 | 89.6 | 1 | 97 | 97 |
| 54 | 73 | - | 95 | - |
| 55 | 100 | 1 | 100 | 100 |
| 56 | 63.6 | - | 94 | 92 |
| 57 | 92.6 | 1 | 99 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, W.J.E.M. Comparative Osteology, Phylogeny and Classification of the Eastern South American Catfish Genus Trichomycterus (Siluriformes: Trichomycteridae). Taxonomy 2021, 1, 160-191. https://doi.org/10.3390/taxonomy1020013
Costa WJEM. Comparative Osteology, Phylogeny and Classification of the Eastern South American Catfish Genus Trichomycterus (Siluriformes: Trichomycteridae). Taxonomy. 2021; 1(2):160-191. https://doi.org/10.3390/taxonomy1020013
Chicago/Turabian StyleCosta, Wilson J. E. M. 2021. "Comparative Osteology, Phylogeny and Classification of the Eastern South American Catfish Genus Trichomycterus (Siluriformes: Trichomycteridae)" Taxonomy 1, no. 2: 160-191. https://doi.org/10.3390/taxonomy1020013
APA StyleCosta, W. J. E. M. (2021). Comparative Osteology, Phylogeny and Classification of the Eastern South American Catfish Genus Trichomycterus (Siluriformes: Trichomycteridae). Taxonomy, 1(2), 160-191. https://doi.org/10.3390/taxonomy1020013